INTRODUCTION
Diverse fossil footprints are abundant and widely distributed in late Pleistocene coastal eolianites and cemented foreshore deposits, preserved along the Cape south coast of South Africa (Fig. 1). These cemented paleodune deposits, which form the ~100,000-yr-old Waenhuiskrans Formation along the Cape south coast, have been surveyed on foot between 2007 and 2019 by the present authors (mainly CWH) from Arniston in the west to Robberg in the east, a distance of ~350 km (Fig. 1).

Figure 1. (color online) Locality map showing the location of track-bearing Waenhuiskrans Formation outcrops in the Still Bay area of South Africa. Site 1 is the more westerly, and site 2 the more easterly of the two sites, marked with stars. The inset at lower right shows the iSimangaliso Wetland Park World Heritage Site northern KwaZulu-Natal, near St. Lucia, known as a nesting area for extant sea turtles.
These tracksite discoveries have generated a recent spate of publications detailing a variety of tracks of mammals and other tetrapods. Most notable have been the reports of hominin tracks (Helm et al., Reference Helm, McCrea, Cawthra, Cowling, Lockley, Marean, Thesen, Pigeon and Hattingh2018b, Reference Helm, Lockley, Cole., Noakes and McCrea2019b), bird tracks (Helm et al., Reference Helm, Anderson, Buckley, Cawthra and De Vynck2017), and various large mammal tracks (Roberts et al., Reference Roberts, Bateman, Murray-Wallace, Carr and Holmes2008; Helm et al., Reference Helm, Cawthra, Cowling, De Vynck, Marean, McCrea and Rust2018a, Reference Helm, Cawthra, de Vynck, Lockley, McCrea and Venter2019a). All these studies stress the vulnerability of track-bearing blocks because of proximity to the ocean and the inevitable damage caused by continuous wave action and cliff collapse.
Although it has been inferred that the Cape south coast track record may be biased toward the tracks of larger, heavier trackmakers, especially mammals (Helm et al., Reference Helm, Cawthra, Cowling, De Vynck, Marean, McCrea and Rust2018a, Reference Helm, McCrea, Cawthra, Cowling, Lockley, Marean, Thesen, Pigeon and Hattingh2018b, Reference Helm, McCrea, Cawthra, Thesen and Mwankunda2018c, Reference Helm, Cawthra, de Vynck, Lockley, McCrea and Venter2019a, Reference Helm, Lockley, Cole., Noakes and McCrea2019b), some tracks, including those of birds (Helm et al., Reference Helm, Anderson, Buckley, Cawthra and De Vynck2017), small mammals (Helm et al., Reference Helm, McCrea, Cawthra, Thesen and Mwankunda2018c), and the hatchling sea turtles reported here, show the potential for registration and preservation of smaller tracks. The only other possible reptile tracks, putatively made by tortoises, and observed in small numbers at three localities (Dana Bay, Gericke's Point, and Goukamma), are morphologically quite distinct from hatchling sea turtle tracks and are as yet undescribed and will be reported elsewhere.
The purpose of this article is to describe a number of highly distinctive and diagnostic tracks (trails) attributed to sea turtle hatchlings. These tracks have considerable ichnological and paleobiological significance for the following reasons: (1) these are the first sea turtle hatchling tracks known from the fossil record; (2) this is the first documentation of fossil reptile tracks from South Africa's Cape south coast, in an ichnofauna otherwise dominated by mammal and bird tracks; (3) from a paleobiological viewpoint these tracks evidently capture the behavior of sea turtle hatchlings in the first few minutes of postemergence activity, the only time they will make terrestrial tracks at this stage of ontogenetic development; (4) from a paleogeographic and paleoecological viewpoint the tracks tell us about the breeding range of sea turtles in the Pleistocene; and (5) the tracks represent two new and highly distinctive and morphologically diagnostic categories of fossil turtle tracks, which require the naming of two new ichnotaxa in order to distinguish them from other formally named turtle track morphotypes.
With reference to the fact that hatchling sea turtle tracks have never previously been reported, it is pertinent to compare them with other previously described tracks of sea and terrestrial turtles from the fossil record, as well as hatchling sea turtle tracks made on southern African beaches today. The fossil trackways are known mostly from the Mesozoic of the Northern Hemisphere and include evidence of both swimming and walking behaviors of presumed mature adults. These are associated with different paleoenvironmental settings from the coastal beach-dune facies described here. However, the morphology of these various Mesozoic trackways, reported with increased frequency in recent years (Bernier et al., Reference Bernier, Barale, Bourseau, Buffetaut, Demathieu, Gaillard and Gall1982; Thulborn, Reference Thulborn, Gillette and Lockley1989, Reference Thulborn1990; Gaillard et al., Reference Gaillard, Bernier, Barale, Bourseau, Buffetaut, Ezquerra, Gall, de Lapparent de Broin, Renous and Wenz2003; Avanzini et al., Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005; Lockley et al., Reference Lockley, McCrea, Buckley and King2018a, Reference Lockley, Smith and King2018b, Reference Lockley, Xing and Xu2019), also has ichnotaxonomic significance, which is reviewed here and discussed in relation to a coherent classification of the fossil tracks and trackways of turtles.
GEOLOGIC SETTING AND STRATIGRAPHIC CONTEXT
The track-bearing Pleistocene coastal dunes and beach sands of the Cape south coast have been described by Roberts et al. (Reference Roberts, Karkanas, Jacobs, Marean and Roberts2012) and Cawthra et al. (Reference Cawthra, Jacobs, Compton, Fisher, Karkanas and Marean2018) based on the Great Brak River outcrops as including three types of Pleistocene facies that can look quite similar. These can be broadly characterized as eolianite facies characteristic of the Waenhuiskrans Formation (Malan, Reference Malan1989) and foreshore and shoreface facies that form part of the Klein Brak Formation (Malan, Reference Malan1991). These are most likely dated from Marine Oxygen Isotope Stage (MIS) 5, between ~130 and ~90 ka (see Helm et al. [2018b] for discussion and the remote possibility of older MIS 11 dates). These facies intergrade to various degrees and are characterized locally by high-angle cross bedding, dune sets, and low-angle to planar bedded foreshore facies.
The track-bearing surfaces are referred to as turtle tracksites 1 and 2. Turtle tracksite 1 was exposed by the fall of a large block, about 2.5 m thick, from the cliffs above. A possible source of the block is ~15 m above its present position (Fig. 2). The block came to rest in a position that, when examined in 2017, exposed the track-bearing surface in a near-vertical orientation. The surface is on the underside of the block where the planar bedded beach sands split along an apparent stratigraphic plane of weakness to expose a very flat surface with maximum dimensions of about 3.0 × 5.6 m, representing a surface of no more than 17.0 m2, on which multiple elongate hatchling turtle trails were registered (Figs. 2–5). The sides of the block reveal that the sand was horizontally bedded or part of a very low-angle sequence, with what appear to be at least two tracks in cross section (Fig. 3B), at a horizon separated from the track-bearing surface by about 40 cm. These tracks appear as “v-shaped depressions” with at least 6–7 cm of relief, but as they do not indent the horizontal layers at the apex of the “v,” they are best referred to as track or bioturbation “features” and not depressions.

Figure 2. Large sandstone block showing parallel sea turtle trackways (black arrows) preserved in concave epirelief on the track-bearing surface. Large and small red squares correspond to line drawings in Figure 5A and C. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)

Figure 3. (color online) (A) Three sea turtle hatchling trackways (Australochelichnus agulhasii) from central area of slab shown in Figure 2. Note that two trackways (right) overlap. (B) Stratigraphic section of track-bearing block, showing the track-bearing surface in vertical orientation (up section to left) and presumed tracks in cross section, center, in layers just left of 15-cm scale bar.

Figure 4. Photogrammetric image of turtle trackway shown in left side of large box in Figure 2 and on left side of Figure 3. This trackway is designated as the holotype of Australochelichnus agulhasii, and represented by a “hard copy” three-dimensional (3D) print (MGL 600 and UCM 230.281) of the representative segment shown in red box. See text for details. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)

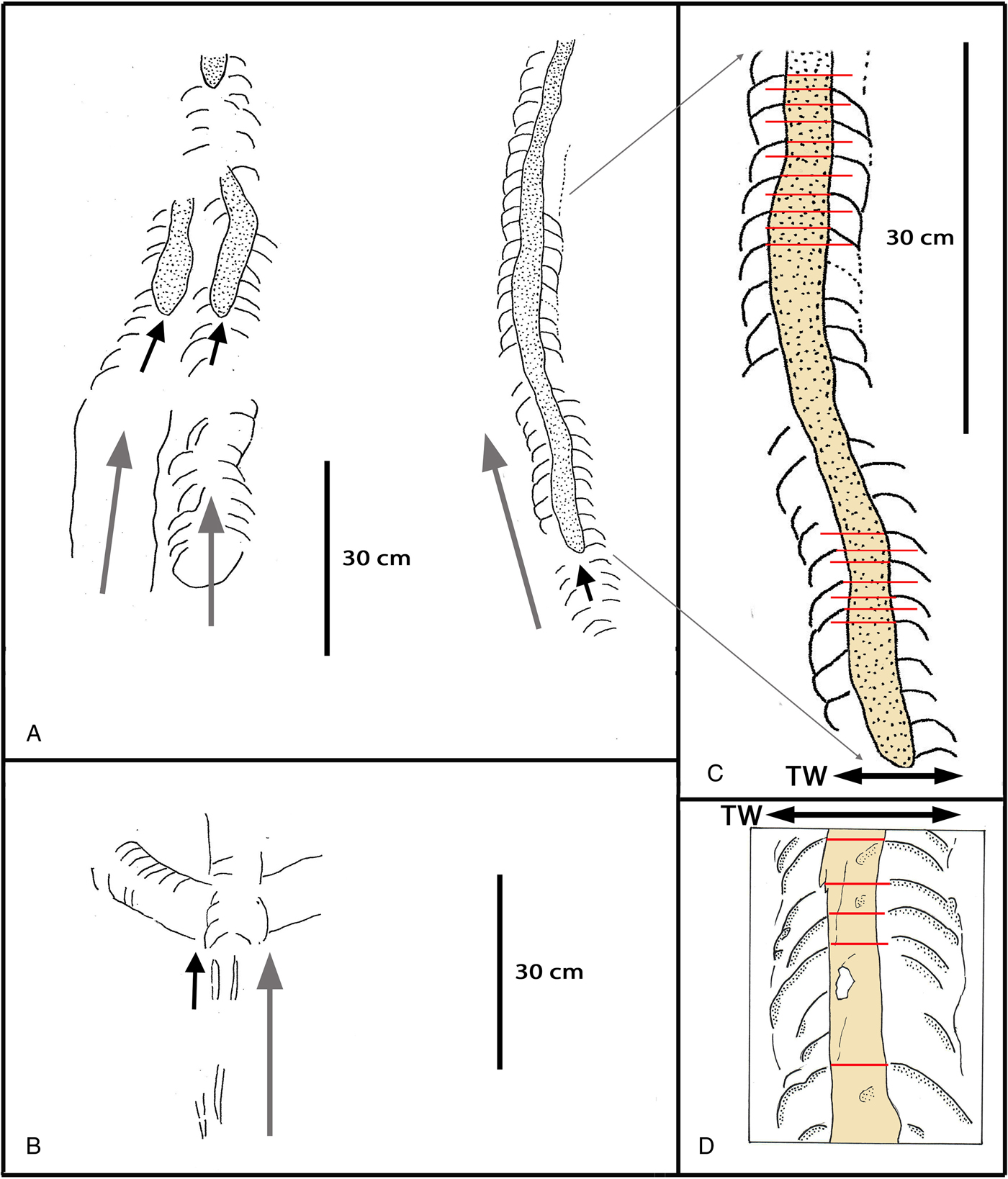
Figure 5. (A) Three subparallel sea turtle trackways (A. agulhasii), inferred to represent leatherback hatchlings: gray arrows show direction of progression. Note appearance of a median ridge (small black arrows) after appearance of paddle traces. (B) Paired trackway-parallel traces before appearance of paddle traces. (C) Detail of trackway shown in panel A to illustrate that paddle traces alternate (left and right), as shown by plotting transverse lines (red) across trackway axis from anterior border of each paddle trace. (D) Sea turtle trackway (Marinerichnus latus), inferred to represent a loggerhead hatchling. Note that left and right paddle traces (red lines) registered symmetrically about the trackway midline. Note that panels C and D are shown at the same scale to show that the M. latus trackway width (TW) is much wider. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
The substrate, on which the trackways were registered, was sand, now consolidated and yellowish brown in color. Short dark streaks on the upper part of this surface at the junction with the upper surface of the block reflect continuation of the dark-brown to black coloration on the upper surface, which is one of the sides where the block is seen in cross section (Fig. 2). Newly exposed surfaces in this area are light yellowish brown, but after prolonged exposure to weathering, exposed surfaces become dark brown or black and, in places, are covered in a biofilm layer, which attaches preferentially to convex and irregular surfaces (Helm et al., Reference Helm, Anderson, Buckley, Cawthra and De Vynck2017). This biofilm also eventually tends to develop on flat surfaces. This dark-brown upper layer, which implies prolonged exposure, when viewed in the context of the source cliffs above, allows the inference that the trackways we describe were heading south in a seaward direction. The trails on the track-bearing surface appear as elongate, slightly curved tripartite furrows with the middle part a raised ridge (convex epirelief) about one-third of trail width, with the two side furrows showing long sequences of transverse paddle traces in concave epirelief. Thus, the trackways are the original natural impressions (concave epireliefs) not the infilling casts (Figs. 2–4). Unfortunately, the counterpart (convex hyporelief) of this surface or the in situ surface from which the block fell is not exposed or accessible for study. When revisited in 2018, the block had split at a stratigraphic level below the track-bearing surface and was tilted seaward away from the near-vertical inclination to an angle of ~60°.
Turtle tracksite 2 is situated 4.4 km east of the previously identified site 1 (turtle tracksite 1) and consists of a subrectangular, fallen sandstone slab 160 cm in maximum length, 90 cm in maximum width, and 32 cm in maximum thickness (Fig. 6). Bedding appears parallel or subparallel, without clear cross bedding. Bedding planes vary in thickness from 2 to 10 mm. The track-bearing surface represents a single bedding plane with the natural impression (concave epirelief) of a single trackway (Figs. 6 and 7). Shell fragments or other coarse material were not observed. The slab is located above the intertidal splash zone and ~ 5m from the bottom of steep coastal cliffs. It is not possible to reliably determine from which layer in the cliffs it originated, and the counterpart slab has not been located. It is also not possible to determine the original bearing of the trackway relative to the coastline. Moreover, the slab is wedged almost vertically between two much larger rocks and is reasonably well protected from erosion, at least in the short term.

Figure 6. (A) The Marinerichnus latus trackway block wedged in between much larger fallen blocks. (B) Close-up view of trackway with arcuate red lines to show curvature of trackway and approximate lengths of inner (concave) and outer (convex) margins at 600 mm and 830 mm, respectively. (C) Photogrammetric model with color vertical profile of leatherback turtle trackway east of Still Bay, courtesy of Carina Helm (see “Material and Methods”). Vertical scale is in meters. Horizontal scale bar is 0.25 m. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)

Figure 7. (color online) (A) Close-up photograph of Marinerichnus latus trackway segment showing measurements of major morphological features made directly form the trackway (see “Description”). (B) Line drawing of the same area showing midline ridge, margins of trackway, and symmetrical arrangement of paddle traces (see “Description”). Compare with Figures 5D and 6.
Tectonic activity is inferred to have been minimal on the Southern Coastal Plain during the Pleistocene (Fleming et al., Reference Fleming, Johnston, Zwartz, Yokoyama, Lambeck and Chappell1998). Thus, the planar bedded strata seen throughout the block can be inferred to represent strata previously situated in situ somewhere in the overlying cliff strata, where the bedding planes reflected inclinations close to their original angle of deposition.
MATERIAL AND METHODS
The tracks at sites 1 and 2 were inspected and photographed in the field in order to obtain both two-dimensional and three-dimensional (3D) images (Figs. 2–7). Two-dimensional images were obtained from photographs, and tracings made on clear acetate film. Tracings were made of three representative site 1 trackway segments (Fig. 5) after chalking the outlines of the main features. The tracings are reposited in the University of Colorado Museum of Natural History as T 1781. Photogrammetric images of the trackways from turtle tracksite 1 were obtained using a Canon PowerShot ELFPH 340 HS camera by one of us (JCDV). Point clouds and digital terrain models were compiled (by RTM) using Agisoft Photoscan Professional (v. 1.0.4), and color topographic profiles were created with CloudCompare (v. 2.6.3.beta) to produce an image of the trackway shown in Figure 4. Attempting to obtain photogrammetric images of the trackways from turtle tracksite 2 proved challenging because of the lack of camera access in cramped space beside the trackway. However, oblique and close-up photos of trackway segments were obtained, from which measurements were procured (Figs. 6–7), and a photogrammetric model was also obtained (Fig. 6C), courtesy of Carina Helm with Agisoft MetaShape Professional (v. 1.0.4) using 27 images from Canon PowerShot ELPH 110 HS with 4608 × 3456 resolution and pixel size of 1.34 × 1.34 µm using a 4.3 mm focal length. The average camera altitude was 0.22 m. The reprojection error of the model is 0.71 pix.
The 3D digital image data for the site 1 trackway were used to generate a 3D print of a segment of the trackway, shown in Figure 4, which is reposited in the fossil footprint collections of Moab Giants Museum (MGL), Utah, as “hard copy” replica MGL 600. A second hard copy replica is reposited at the University of Colorado Museum of Natural History (UCM) as specimen UCM 230.281.
While this article was in review, a third tracksite yielding a hatching turtle trackway was discovered (in April 2019), in a coastal section of the Garden Route National Park, by Andre and Emily Brink. This site is 110 km east of sites 1 and 2 and consists of an isolated loose slab ~60 cm × 60 cm wide and long and ~20 cm thick (Fig. 8). The slab was found on a dune surface below vegetated slopes lacking nearby visible outcrops to suggest the stratigraphic level from which it came. Faint horizontal bedding is evident, parallel to that of the track-bearing surface, which appears to have been exposed and weathered/eroded for some time. Nonetheless, the natural mold of one clear turtle trackway with an alternating paddle trace gait is similar in size and morphology to the site 1 trackways inferred to represent a hatchling loggerhead turtle (Caretta caretta).

Figure 8. (color online) Turtle trackway site 3, Garden Route National Park area. (A) View of isolated track-bearing slab on beach. (B) Detail of slab with clear, diagnostic trackway segment (long arrow) and unclear trace (short arrow left) with opposite sense of progression. Trackway segments occur on either side of an uneven area that might represent remains of a nest. See text for details.
The diagnostic trackway segment, 25 cm long and 9 cm wide, is preserved in the middle of the slab with evidence of forward motion away from a disturbed (bioturbated) area (left to right in Fig. 8) and toward a broken area (far right in Fig. 8). On the opposite side of the uneven area from the diagnostic trackway segment, other paddle-like traces, with curved edges, consistent with hatchling turtle trackmaker morphology are evident and apparently suggest motion in a direction ~180° opposite to that of the main trackway. This ichnological evidence is difficult to interpret but could possibly suggest at least two trackways emanating from a nest site. Ichnologically, the trackways from sites 1 and 2 can be classified as epifaunal trails, whereas a nest would be an infaunal trace, and potentially more easily preserved than surface trails, and potentially amenable to study through sectioning of the sedimentary rock comprising the slab. However, although the site 3 specimen (Fig. 8) is portable and can be removed to the safety of a repository for further study, it is premature to undertake any potentially damaging analysis.
A fourth turtle trackway site was reported to us (see “Acknowledgments”). However, details of this fourth site, although indicated by photographs, are insufficiently known to allow us to describe the material adequately without further study of the field evidence.
ICHNOTAXONOMY
General ichnotaxonomic observations
According to the latest taxonomic revisions (Rhodin et al., Reference Rhodin, Iverson, Bour, Fritz, Georges, Shaffer and Van Dijk2017), all tortoises, a subgroup within the turtles (Chelonia), are terrestrial. Thus, all tortoises are turtles, but not all turtles are tortoises. Nontortoise turtles are mostly aquatic and include both terrestrial, freshwater turtles and sea turtles. In the discussion that follows, we use the term turtle to exclude terrestrial tortoises. This is convenient when discussing the fossil track record of chelonians, as there are no confirmed reports of the tracks of terrestrial tortoises, even though a few questionable Cretaceous occurrences have been suggested (Fiorillo, Reference Fiorillo2005; Pascual-Arribas and Hernández-Medrano, Reference Pascual-Arribas and Hernández-Medrano2015), and we have recently observed a few as yet undescribed tortoise tracks in the coastal dune and beach deposits described here.
Fossil turtle tracks are known from the Mesozoic and Cenozoic, with some evidence that the earliest known tracks, from the Early Triassic, are older than the oldest known skeletal remains (Lichtig et al., Reference Lichtig, Lucas, Klein and Lovelace2018). The turtle track record covers a variable range of morphologies reflecting the locomotion of different-sized marine and freshwater turtles while either swimming or progressing on land. In most cases there are clear-cut differences between the trackway configurations made by swimmers and walkers, and also between the facies associations of marine or sea turtle traces (Fig. 9) and those associated with freshwater settings. There are also at least four widely used ichnogenus names applied to turtle tracks. These include Chelonichnium, originally applied to an undiagnostic isolated track (C. vogesiacum) from the Early Triassic of Germany (Schimper, Reference Schimper1850). As reviewed by Avanzini et al. (Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005), Abel (Reference Abel1935), and Haubold (Reference Haubold1971), this ichnotaxon was declared dubious (a nomen dubium). This led Demathieu and Gaillard (1982, in Bernier et al., Reference Bernier, Barale, Bourseau, Buffetaut, Demathieu, Gaillard and Gall1982), to propose adopting (transferring) the ichnogenus name to label large sea turtle tracks from the Late Jurassic of France. These are preserved in a long trail (Fig. 10), which they named Chelonichnium cerinense. These authors used the potentially ambiguous term “turtle” throughout their study, which we take to mean nontortoise turtles. (However, they may have wished to imply tortoise.) They explicitly described the Chelonichnium trackway as progressing from a drier to wetter substrate in an area with what they described as “soft sediment slides” confirming a substrate at least partly immersed, subaqueously, in shallow marginal marine facies. Interestingly, as in the case noted previously where Early Triassic turtle tracks may predate the oldest body fossil remains, so these Late Jurassic sea turtle tracks apparently also predate the oldest known body fossil remains.

Figure 9. (color online) (A–C) Modern hatchling loggerhead turtles en route to sea, registering trackways on sandy substrates with variable surface textures. Note scale bars in panels A and C for comparison with Figure 7. (D) Adult loggerhead digging nest. Trackway (~1.0 m wide) indicates onshore (up-beach) progression and shows distance from sea. Photographs in panel A (November 2015) and panel D (2013) courtesy of Linda Harris. Photographs in panels B and C (January 2019) courtesy of Diane Le Gouvello. All photographs from Bhana Nek, in iSimangaliso Wetland Park World Heritage Site northern KwaZulu-Natal, South Africa.

Figure 10. (color online) Sea turtle (A–D) and terrestrial turtle (E–G) trackways. (A–C) From the Jurassic of France. (A) Chelonichnium cerinense (after Bernier et al., Reference Bernier, Barale, Bourseau, Buffetaut, Demathieu, Gaillard and Gall1982). (B) Saltosauropus latus (after Bernier et al., Reference Bernier, Barale, Bourseau, Buffetaut, Demathieu, Gaillard, Gall and Wenz1984). (C) Unnamed Saltosauropus-like giant turtle tracks (modified after Gaillard et al., Reference Gaillard, Bernier, Barale, Bourseau, Buffetaut, Ezquerra, Gall, de Lapparent de Broin, Renous and Wenz2003). (D) Australochelichnus agulhasii from South Africa. (E) Chelonipus plieningeri from the Upper Triassic of Germany (after Haubold, Reference Haubold1971, Reference Haubold1984). (F) Chelonipus isp. from the Cretaceous of Utah (after Lockley et al., Reference Lockley, McCrea, Buckley and King2018a). (G) Turtle swim track assemblage T 1491 from the Cretaceous of Colorado (after Lockley et al., Reference Lockley, Smith and King2018b).
Bernier and his colleagues also named Saltosauropus latus (Bernier et al., Reference Bernier, Barale, Bourseau, Buffetaut, Demathieu, Gaillard, Gall and Wenz1984), which they initially misinterpreted as tracks of a hopping dinosaur. These tracks were convincingly reinterpreted as those of large swimming marine turtles, which provided trace evidence of large paddles used in synchronous swimming strokes (Thulborn, Reference Thulborn, Gillette and Lockley1989, Reference Thulborn1990). This interpretation has been supported by additional finds of trackways “attributed to giant turtles” (Gaillard et al., Reference Gaillard, Bernier, Barale, Bourseau, Buffetaut, Ezquerra, Gall, de Lapparent de Broin, Renous and Wenz2003, p. 315) that resemble Saltosauropus (Fig. 9). Thus, in the sense used by Bernier and his colleagues, Chelonichnium and Saltosauropus respectively represent the progression of a marine turtle, in the former case over a wet partly submerged substrate, and in the latter case, swimming more freely, but still touching the substrate with paddle strokes.
Because freshwater turtles, associated with fluviolacustrine facies in terrestrial deposits, may also walk or swim, we can expect to find trackways representing both modes of progression. This is exactly what we find in the track record. Generally, the traces of freshwater turtles have been labelled Chelonipus (Rühle von Lilienstern, Reference Rühle von Lilienstern1939), although the ichnogenus name Emidhypus (Fuentes Vidarte et al., Reference Fuentes Vidarte, Meijede Calvo, Meijede Fuentes Calvo, Meijede Fuentes Calvo and Perez-Lorente2003) has also been used (see Avanzini et al., Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005, for review of ichnotaxonomy). The record of Chelonipus extends from the Early and Late Triassic of Europe (Rühle von Lilienstern, Reference Rühle von Lilienstern1939, and Haubold, Reference Haubold1971, respectively) to the few sites in the Jurassic of North America (Foster et al., Reference Foster, Lockley and Brockett1999; Lockley and Foster, Reference Lockley and Foster2006) and multiple tracksites in the Cretaceous of North America (Lockley et al., Reference Lockley, McCrea, Buckley and King2018a, Reference Lockley, Smith and King2018b). The label Emidhypus has been used to describe Middle Jurassic (Klein et al., Reference Klein, Lagnaoui, Gierliński, Saber, Lallensack, Oukassou and Charrière2018) and Late Jurassic turtle tracks from Spain (Avanzini et al., Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005) and Morocco, as well as the original tracks described from the Early Cretaceous of Spain (Fuentes Vidarte et al., Reference Fuentes Vidarte, Meijede Calvo, Meijede Fuentes Calvo, Meijede Fuentes Calvo and Perez-Lorente2003). The difference between the type specimens of Chelonipus and Emidhypus is slight, based on more “parallel ungual traces” in the manus (front foot), turned slightly inward, and “apparently always away in respect to” the pes (hind foot) (Avanzini et al., Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005, p. 748). These authors considered that Emidhypus might be a “junior synonym” of Chelonipus but did not opt for this conclusion because the trackway configurations were different. Fuentes Vidarte et al. (Reference Fuentes Vidarte, Meijede Calvo, Meijede Fuentes Calvo, Meijede Fuentes Calvo and Perez-Lorente2003) had failed to compare their Emidhypus with Chelonipus but were “saved” by Avanzini et al. (Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005) who demonstrated diagnostic differences justifying both names as valid. Note here that the differences in trackway configuration are considered sufficient to validate the naming of different ichnogenera.
Avanzini et al. (Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005) explicitly interpreted the type material of both ichnogenera as examples of walking behavior and supported this conclusion by comparing the fossil trackway with those of living turtles, including the tortoise Testudo and other nontortoise turtle genera. Reolid et al. (Reference Reolid, Márquez-Aliaga, Belinchón, García-Forner, Villena and Martínez- Pérez2018) described trackway evidence of what they called “semi-aquatic locomotion” but indicated that tracks could not be given a “concrete assignment” to either Chelonipus or Emidhypus. Thus, these two ichnogenera may be used as labels for walking trackways, in the type specimens, but may elsewhere be difficult to distinguish and may additionally indicate semiaquatic locomotion.
Turtles are capable of subaqueous bottom walking and, depending on substrate consistency (firmness), may register very indistinct trackways that do not closely reflect foot morphology (Avanzini et al., Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005), or they may register quite regular trackway patterns that are difficult to distinguish from those made when walking on emergent subaerial substrates (Foster et al., Reference Foster, Lockley and Brockett1999; Avanzini et al., Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005; Lockley et al., Reference Lockley, Cart, Martin, Prunty, Houck, Hups, Lim, Kim, Houck and Gierlinski2014). In other paleoenvironmental settings, high densities of parallel to subparallel, incomplete tracks (mostly distal toe traces) with no discrete or recognizable trackways configurations have been registered (Lockley et al., Reference Lockley, McCrea, Buckley and King2018a, Reference Lockley, Xing and Xu2019). The inability of observers to recognize trackways may be because of multiple overprinting. These suggest swimming, with toes touching the substrate under the influence of unidirectional or preferred orientation currents, or some other paleoenvironmental influence on the direction of progression of multiple individuals.
The South African tracks named here are morphologically quite distinct from any turtle trackways previously described but exhibit some general similarities to the turtle traces named Chelonichnium cerinense (Demathieu and Gaillard, 1982, in Bernier et al. Reference Bernier, Barale, Bourseau, Buffetaut, Demathieu, Gaillard and Gall1982). For these reasons, we erect the new ichnotaxa Australochelichnus agulhasii ichnogen. et ichnosp. nov. and Marinerichnus latus ichnogen. et ichnosp. nov., which recognize the “chelonian” and “mariner” affinities of the trackmakers and also recognize that the two trackways registered quite different “terrestrial” gaits (i.e., alternating in A. agulhasii and symmetrical in M. latus). As these different gaits are unequivocally tied to loggerhead (genus Caretta) and leatherback (genus Dermatochelys) turtles, respectively, it is ichnotaxonomically consistent to place them in separate ichnogenera. However, both represent extant sea turtles, or close relatives, and can be united in a single ichnofamily. Because of the opinion of various ichnologists that Chelonichnium is a nomen dubium (Avanzini et al., Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005, and references therein), we resist using this ichnogenus name as the basis of an ichnofamily. Instead, we erect the new ichnofamily Marineropodidae (derived from mariner, meaning seafarer: Spanish/French/English).
Formal ichnotaxonmy
Ichnofamily Marineropodidae
Referred material
Australochelichnus agulhasii ichnogen. et ichnosp. nov. MGL 600/UCM 230.281, this study. Marinerichnus latus ichnogen. et ichnosp. nov., this study.
Narrow, elongate subsymmetrical trackways (trails) characterized by a trilobed configuration consisting of a central ridge, or groove, with serial, posteriorly divergent paddle traces oriented postero-laterally on both sides of the trackway midline. Paddle impressions may register as mirror image traces, or with slight alternation on either side of the central ridge.
Discussion of ichnofamily features
Where paddle traces are deeper than the trace of the ventral plastron, because of the lifting of the trackmaker's body above the deepest paddle registration points, the central or midline trace between the paddle marks will appear as a raised ridge in natural impressions (a convex epirelief ridge separated by concave lateral grooves with serial paddle traces. A natural cast of the natural impression will show all features in reverse—that is, an indented groove (concave hyporelief groove separated by convex lateral ridges with serial paddle trace casts). The height (or depth) of the central ridge (groove) in natural impressions will vary depending on the substrate and the ability of the trackmaker to raise its plastron enough to reduce or minimize contact with the substrate.
Australochelichnus ichnogen. nov.
Diagnosis. Continuous subsymmetrical trails about 10 cm wide with median groove averaging about 35–40% as wide as whole trail separating very regular serial, posteriorly angled rhomb-shaped “paddle” traces, with anteriorly convex margins. Anterior margins of paddle traces show an alternating pattern. Median groove may be locally absent.
Type material. Holotype and paratype trails occur as natural impressions on a large fallen block of consolidated beach sandstone. Holotype preserved as photo series used to create 3D image (Fig. 3).
Age of type material. Middle to late Pleistocene.
Type horizon and locality. Klein Brak Formation, east of Still Bay, ~4.4 km west of M. latus type locality.
Derivation of ichnogenus name. “Southern turtle trail,” Australo meaning southern, chel referring to chelonian or turtle, and ichnus meaning trace or trail.
Australochelichnus agulhasii ichnosp. nov., Figures 2–5
Type material. As for ichnogenus. MGL 600/UCM 230.281.
Age of type material. As for ichnogenus.
Type horizon and locality. As for ichnogenus.
Derivation of ichnospecies name. Referring to Agulhas current, which may have influenced turtle breeding range in the study area.
Description. Continuous subsymmetrical trails about 10 cm wide with median ridge ranging from 25% to 50% as wide as whole trail but averaging 35–40% of trail width. Median ridge separating posteriorly angled rhomb-shaped flipper or paddle traces slightly wider than long, where length is measured parallel to trail axis. Left and right paddle traces show regular alternating pattern, with anterior margins on each side offset about half a paddle trace length relative to opposite side. In cross section median ridge appears in convex epirelief, whereas lateral paddle traces are subhorizontal to slightly concave epireliefs. Anterior margins of paddle traces are crescent to semicircular in shape and anteriorly convex, with posterior margins marked by identical anteriorly convex traces of previously registered paddle marks, which repeat to create a highly regular series of traces on each side. Median margins of paddle traces are angled at about 110° to direction of travel marked by trackway midline, and curving postero-laterally to create outer trackway margin.
Some trails show segments that registered only lateral paddle traces, followed continuously by segments where the median ridge is also present. This suggests a change in locomotor style or substrate consistency that is reflected in differences in the configuration of the trail.
Marinerichnus ichnogen. nov.
Diagnosis. Continuous subsymmetrical trails about 14–15 cm wide with median ridge averaging about 30% as wide as whole trail separating very regular serial, posteriorly angled rhomb-shaped “paddle” traces, with anteriorly convex margins. Anterior margins of paddle traces show mirror image configuration about trackway midline.
Type material. Holotype trails occur as natural impressions on fallen block of consolidated beach sandstone (Figs. 5D, 6, and 7).
Age of type material. Middle to late Pleistocene.
Type horizon and locality. Klein Brak Formation, east of Still Bay, ~4.4 km east of A. agulhasii type locality.
Derivation of ichnogenus name. “Seafarer turtle trail,” mariner meaning one who goes to sea, and ichnus meaning trace or trail.
Marinerichnus latus ichnosp. nov., Figures 6–7
Type material. As for ichnogenus.
Age of type material. As for ichnogenus.
Type horizon and locality. As for ichnogenus.
Derivation of ichnospecies name. Referring to latus, the width of the trackway.
Description. A continuous symmetrical trail about 14–15 cm wide and ~70 cm long as measured along curved midline, with median ridge about 30% as wide as whole trail. Median ridge separating posteriorly angled arcuate paddle traces, longer than wide, where length is measured parallel to trail axis. Left and right paddle traces show symmetrical arrangement about the median ridge/trackway midline. In cross section median ridge appears in convex epirelief whereas lateral paddle traces are concave epireliefs, up to 15 mm deep. Anterior margins of paddle traces are arcuate anteriorly and antero-laterally with posterior margins marked by identical anteriorly convex traces of previously registered paddle marks. However, the length of paddle traces measured parallel to the trackway axis varies, averaging ~2.5 cm (range ~1.5–2.5 cm). Median margins of paddle traces are angled at about 130° to direction of travel marked by trackway midline and curve postero-laterally to create outer trackway margin.
Comparisons between A. agulhasii and M. latus
A. agulhasii and M. latus can be differentiated by a number of distinctive morphological features of the traces, which appear to have unequivocal ichnotaxonomic significance. For example, M. latus is a wider (14.0–15.0 cm) trackway with larger, more posteriorly oriented paddle traces, arranged symmetrically about the trackway midline. This makes the median ridge narrower in proportion to the trackway width (~30%, compared with 35–40% in A. agulhasii). This is in marked contrast to the narrower trackway of A. agulhasii (~10.0 cm, thus only ~67–71% as wide as M. latus), which has alternating and shorter, less posteriorly oriented paddle traces. All features of both trackways are consistent with inferring that A. agulhasii was made by a loggerhead turtle hatchling and M. latus by a leatherback turtle hatchling. The M. latus trackway is curved, and there is greater variation in the length of the paddle traces as measured parallel to the midline. In contrast, the alternating A. agulhasii paddle traces are very regular in size and shape.
Ichnological observations and interpretations
In order to place the A. agulhasii trails (trackways), as well as the M. latus trackway, in their paleoenvironmental context, it is important to note that in the case of A. agulhasii there are multiple parallel trackways on a single block (~5.6 x 3.0 m: ~17.0 m2), which we interpret as a beach deposit (see Discussion). Five of these trackways are marked with arrows (Fig. 2), two are shown in Figure 3A, one (the holotype) is shown as a 3D image (Fig. 4), and four are illustrated with line drawings (Fig. 5). Distinctive features of these trackways are their continuous and subsymmetrical configurations and the very regular serial pattern created by the paddle marks (Figs. 3A, 4, and 5). However, as noted in the description, trackway segments where only lateral paddle traces were registered are, in places, followed by segments where a median ridge occurs. This suggests either (a) that the trackmaker carried the animal's central carapace clear of the substrate (with an elevated posture) for a certain distance before lowering to the point where the carapace left a central groove, or (b) that substrate conditions changed subtly as the trackmaker progressed. The central ridge appears at approximately the same place in the three parallel trackways illustrated in Figure 5A, which potentially suggests the latter interpretation (b) is the more parsimonious.
Given this latter possibility, differences in the registration of traces might reflect the passing of the trackmakers across substrate areas with different physical properties (consolidation, moisture content, etc.), as would be expected across a beach, especially perpendicular to the shoreline. In this regard, we note that the trails occur as natural impressions on the once upward-facing surface of a large fallen block from which the matching surface, the “counterpart” natural casts, are lost. The block is several meters thick, and the lower portion is composed of planar bedded sandstone of the Klein Brak Formation. This bedding style indicates that the block represents beach deposits rather than dune deposits of the Waenhuiskrans Formation, which elsewhere in the formation are represented by cross-bedded facies. This interpretation is also consistent with the occurrence of sea turtle hatchling trackways.
Given the subparallel trackway pattern that, as noted previously, we infer to be shore-perpendicular, it is possible to suggest that the tracks were not made in, or radiating out from, the immediate vicinity of a nest, likely in dry sand above the high tide mark. Rather they likely represent down shore movement from some point near the previous high-tide mark, with sand wet enough to leave well-defined tracks. There is no unambiguous evidence to reveal the mechanisms that allowed tracks to be buried and preserved without being eroded or washed out. However, a very plausible mechanism would be the blowing or drifting of sand over the wet surface without the power to cause deflation. If this was the preservation mechanism, it is possible that the covering layer of blown sand was more susceptible to postconsolidation and postexhumation erosion, perhaps because of being drier and less prone to cementation. Such a scenario would help explain why the impression surfaces are preserved and as yet no infilling cast surfaces have been found—that is, the lack of casts could be because of the preferential weathering away of the infilling layer to more readily expose the more resistant surface with the natural trackway impressions.
The Marinerichnus latus trackway is not associated with any other recognizable tetrapod traces, although some invertebrate traces are evident (Fig. 6B). The trackway appears “suddenly” relative to the bedding plane surface of the block. It is tempting to suggest that the trackmaker was emerging from the sand, possibly its nest, but as noted previously, the apparent sudden appearance may represent the passage of the trackmaker from one layer of sand to another more suitable for registering traces. Thus, the emergence possibility, although not entirely ruled out, is speculative. However, the discovery of two sea turtle hatchling trackway morphotypes (representing at least six individuals), within a geographic area of paleoshoreline less than 5 km wide, suggests that more sea turtle tracks and traces, including nests, could be found. This inference is supported by the aforementioned suggestion that the site 3 specimen (Fig. 8) might represent a nest, which, as an infaunal trace, potentially has higher if more localized preservation potential than epifaunal surface trackways. As illustrated in Fig. 9, the shore perpendicular zones of nest excavation and trackway registration are quite localized on the scale of tens of meters. Tetrapod nests and nesting grounds are not uncommon in the fossil record at least as early as the Mesozoic (Carpenter et al., Reference Carpenter, Hirsch, Horner, Carpenter, Hirsch and Horner1994; Carpenter, Reference Carpenter1999).
DISCUSSION
Neoichnology of sea turtle tracks
There seems little doubt that the A. agulhasii and M. latus trackways are unequivocally attributable to hatchling sea turtles, moving over exposed surfaces (i.e., creating epifaunal trails). Prior to making this determination, we considered the need to eliminate the possibility that the trackways might conceivably represent infaunal sand-swimming traces of a genus like Eremitalpa from the fossorial golden mole family (Chrysochloridae) (Stuart and Stuart, Reference Stuart and Stuart2000). However, such traces are not compatible with the features seen here. Less likely possibilities, including crawling penguins during times of preterminal distress leaving tramline impressions with their wings, are incompatible with the evidence reported here of seven similar subparallel trackways, all too small to permit the unlikely conjecture that they could be attributable to distressed, beached birds. Other “tramline trackmakers” such as crawling, rather than hopping anurans (frogs or toads) or invertebrates such as scorpions or crabs, known to inhabit beach and sand dune settings, are all easily discounted because of their known trackway configurations being quite different in size and morphology (Häntzschel, Reference Häntzschel1975, and multiple references to invertebrate trails therein). Likewise we can discount the clawed tracks of other reptiles such as terrestrial tortoises, terrapins, and crocodylians, which again leave distinctive trackways described from both modern and ancient sediments (e.g., Milàn et al., Reference Milàn, Lucas, Lockley and Spielmann2010).
The formality of excluding the aforementioned list of potential trackmakers for A. agulhasii and M. latus leaves the sea turtle hatchling interpretation as the only realistic possibility. This is strongly supported by several compelling lines of evidence. First, the tracks are morphologically highly similar to those of extant sea turtles. Second, they occur in coastal deposits, which represent beach environments seaward of sand dunes. Third, the presence of seven parallel to subparallel trackways, in an area less than 6.0 m wide, indicates a large number of individuals moving in the same direction in a small area, evidently at the same time. Fourth, the similarity in trackway configuration suggests they moved at the same speeds. This is the behavior expected of hatchling sea turtles heading across a beach toward the sea. The fact that we have two morphologically diagnostic morphotypes that are compellingly attributed to loggerheads and leatherbacks not only supports the sea turtle interpretation generally, but also points to the possibility of deriving paleoecological and paleogeographic interpretations pertaining to two major sea turtle groups.
Online images of sea turtle hatchling trackways and videos of baby turtles scrambling to the sea after hatching are abundant and easily found but pertain to a wide range of geographic localities. These include various images that show parallel tracks and trackways that change in appearance as the substrate the trackmakers cross also changes in consistency (moisture content, firmness of sand) in a seaward direction. Many of the images show a great variety of turtle trackway configurations including patterns similar to those recorded here. For example, a trackway from Turtle Sanctuary Beach, Palau Perhentian Besar, Malaysia, is practically identical to those recorded here (Fig. 9). The variation seen in modern sea turtle hatchling trackways can be attributed to differences in species morphology such as relative size of larger anterior paddles and smaller hind feet.
All trackways generally share the common characteristic of having been made on sandy substrates with anterior paddle traces on the lateral margins. However, although the central portions of some trackways show traces of body or carapace drag traces, others do not, but instead show the traces of the hind limbs or other indications that the carapace did not touch the substrate. These differences may also be attributed to the speed of progression of individuals, which in turn may be influenced by substrate consistency.
Paleoichnology of sea turtle tracks
To the best of our knowledge, there are no previous reports of sea turtle hatchling tracks from the fossil record. The only reports of sea turtle tracks come from Jurassic limestones from the Cerin region of France. These fall into the category of swimming traces attributed to a large marine turtle (Thulborn, Reference Thulborn, Gillette and Lockley1989, Reference Thulborn1990; Lockley and Meyer, Reference Lockley and Meyer2000; Gaillard et al., Reference Gaillard, Bernier, Barale, Bourseau, Buffetaut, Ezquerra, Gall, de Lapparent de Broin, Renous and Wenz2003; Fig. 10). These trackways were initially, and controversially, interpreted as evidence of hopping dinosaurs (Bernier et al., Reference Bernier, Barale, Bourseau, Buffetaut, Demathieu, Gaillard, Gall and Wenz1984). It was argued that the symmetrical configuration of footprints, each with three-digit traces, supported such inferences. So, the tracks were named Saltosauropus latus, implying a hopping or jumping reptile (Fig. 10). These paired tracks were subsequently reinterpreted as the result of synchronous registration of the paddles of a large marine turtle (Thulborn, Reference Thulborn, Gillette and Lockley1989, Reference Thulborn1990) swimming in shallow water over a calcareous mud substrate. Thulborn's interpretation was substantiated by further discoveries of swimming sea turtle tracks by Gaillard et al. (Reference Gaillard, Bernier, Barale, Bourseau, Buffetaut, Ezquerra, Gall, de Lapparent de Broin, Renous and Wenz2003), which they claimed represented the oldest evidence of marine turtles. These Jurassic trackways are quite different from those described here from the Pleistocene Klein Brak Formation, in all relevant ichnological details. First, the trackways are much larger, up to 1.5 m wide. Second, they represent swimming behavior and show only paddle or flipper marks with no central body trace. Third, they occur in very fine-grained calcareous and subaqueously deposited substrates.
As noted previously, prior to the incorrect hopping dinosaur interpretation proposed by Bernier et al. (Reference Bernier, Barale, Bourseau, Buffetaut, Demathieu, Gaillard, Gall and Wenz1984), Bernier et al. (Reference Bernier, Barale, Bourseau, Buffetaut, Demathieu, Gaillard and Gall1982) had described a turtle (chelonian) trackway they formally named Chelonichnium cerinense, using the ichnogenus name Chelonichnium originally proposed by Schimper (Reference Schimper1850) (see Avanzini et al., Reference Avanzini, García-Ramos, Lires, Menegon, Piñuela and Fernández2005, for discussion). Bernier et al. (1982, p. 447) interpreted the C. cerinensis trackway (Fig. 10A) not as that of a sea turtle but as a “terrestrial” chelonian walking over a “very wet, superficially soft, and sloping” substrate interpreted as a carbonate “platform” behind a ribbon of islands. They also considered that the animal was progressing down a decreasingly steep slope and over an increasingly wet substrate. Despite the suggestion that this was a terrestrial turtle (? tortoise) the slope interpretation begs the question: Was this in fact the trackway of a marine turtle going to sea? Gaillard et al. (Reference Gaillard, Bernier, Barale, Bourseau, Buffetaut, Ezquerra, Gall, de Lapparent de Broin, Renous and Wenz2003) hint at this possibility without explicitly reinterpreting the Chelonichnium trackway. They note that large marine turtles likely came into very shallow water and may have nested in the area.
Literature on sea turtle locomotion discusses the development of the hypertrophied, hydrofoil- or wing-like front limbs (paddles or flippers) and the modified coordination of the four limbs as part of the secondary adaptation of land-based tortoises (chelonians) to aquatic marine and freshwater environments (Davenport et al., Reference Davenport, Munks and Oxford1984; Renous et al., Reference Renous, Bels and Davenport1989). Of interest here are the differences between the alternating (left-right) gaits of walkers on land, the perhaps alternating strokes made by some freshwater swimmers, and the synchronous strokes made by large marine turtles when swimming (and also on beaches), as inferred for the large Jurassic swim trackways (Thulborn, Reference Thulborn, Gillette and Lockley1989, Reference Thulborn1990; Lockley and Meyer, Reference Lockley and Meyer2000; Gaillard et al., Reference Gaillard, Bernier, Barale, Bourseau, Buffetaut, Ezquerra, Gall, de Lapparent de Broin, Renous and Wenz2003). Smaller modern sea turtles may swim with alternating strokes, but given that all fossil trackways attributable to marine turtles are Jurassic in age, conjectures on the swimming styles of all fossil turtles are necessarily speculative.
Paleoenvironmental implications
The Pleistocene beaches represented by the late Pleistocene Klein Brak Formation sediments would have provided suitable nesting sites for sea turtles. Basic requirements for successful nesting incubation of extant sea turtles are as follows: beach sand above the high-tide mark that is deep enough (50–80 cm) to allow for burial of eggs; sufficient sand moisture so that the leathery eggs do not dry out; sand that is not so wet as to drown eggs by prohibiting gas exchange; temperatures between 25°C and 35°C to allow for successful incubation. Sand temperatures below the pivotal temperature of ~29.3°C generate mostly male hatchlings, and above this pivotal temperature mostly females (Maxwell et al., Reference Maxwell, Morata and Frank1988; Davenport, Reference Davenport1997). Compared with other extant sea turtles, loggerhead and leatherback turtles are able to nest successfully in subtropical, rather than tropical regions.
In southern Africa, these conditions exist most consistently more than 1200 km to the northeast along the beaches of northern KwaZulu-Natal, as a result of the Agulhas Current generating warm conditions along the shore for successful incubation. The rookeries of loggerhead and leatherback turtles on the KwaZulu-Natal coastline, in the iSimangaliso Wetland Park World Heritage Site just south of the border with Mozambique, represent the southernmost regular sea turtle breeding sites in the world (see St. Lucia area: Fig. 1 inset). However, occasionally nests of either loggerhead or leatherbacks are reported alongshore between St. Lucia to Cape Town, either at the beginning (October) or end (January or February) of the nesting season, suggesting that female turtles had an early or late clutch still to lay on the migration between the foraging and nesting grounds. Of these nests, one near Alexandria (500 km east of the Pleistocene site described here) was reported to have incubated successfully. Depending on the seasonal temperature and assuming the clutch is not deposited in a dune shade or under vegetation, with 55–80 days of warm weather available (i.e., the incubation period for turtle nests), occasional turtle nests could conceivably incubate along most parts of the coast east of Cape Agulhas. However, this assumes near-ideal incubation conditions.
We are aware of two anecdotal reports of recent nesting turtles (Dermochelys) on the Cape south coast at Still Bay in 2005 (du Plessis, J., personal communication, 2018) and at Buffelsbaai in 2007 (Combrink, X., Baker, N., personal communication, 2018), as well as reports and photos of tracks and clutch of eggs from Nautilus Bay in 2011 (Scholtz Reference Scholtz2011; Heyns, K., personal communication, 2018) and a report of a leatherback turtle at Kleinemonde in January 2018. Although these reports indicated failed nests, they are relevant to the evidence for fossil nesting just east of Still Bay, because they indicate the overlap, at least marginally, if not extensively with the paleogeographic and geographic regions where extant sea turtle nesting occurs. The A. agulhasii trackways reported here appear consistent with the alternating gait tracks of extant loggerhead hatchlings (Caretta caretta) (personal observation by RN; see Fig. 9). The occurrence of M. latus trackways representing hatchlings, in the same region, confirms that leatherbacks also nested here in the Pleistocene. MIS 5e was characterized by warmer global temperatures, with sea levels 5–6 m higher than present levels. These warmer temperatures could conceivably have allowed turtles to nest on the Cape south coast. The existence of both loggerhead and leatherback hatchling turtle trackways within a 5 km stretch of coastline on the Cape south coast suggests a similar phenomenon to that which currently occurs in the iSimangaliso Wetland Park World Heritage Site near St. Lucia (Figs. 1 and 9), with the two species nesting in proximity to one another.
CONCLUSIONS
Fossil trackways of hatchling sea turtles have never previously been reported from the ancient track record. The multiple trackways reported here from the Klein Brak Formation and named Australochelichnus agulhasii ichnogen. and ichnosp. nov. are therefore a novel and highly distinctive addition to the tetrapod paleoichnological record. The trackways are unequivocally diagnostic of hatchling loggerhead turtles, which left traces that are morphologically quite distinct from formally named fossil trackways of large adult sea turtles, currently known only from the Jurassic of the Northern Hemisphere. The occurrence of multiple subparallel trackways, all representing similarly sized hatchlings, is also of behavioral significance in indicating that they were made postemergence from their nests, during a downshore “run for the sea.” The finding of a trackway (Marinerichnus latus) consistent with those of hatchling leatherback turtles, less than 5 km from the loggerhead turtle tracksite, appears to duplicate the current nesting situation in a subtropical coastal environment farther to the northeast, where the two species breed in proximity to one another. Their occurrence in Pleistocene deposits of the Cape south coast, likely dated from the MIS 5 stratigraphic interval, is the first direct evidence of the antiquity of sea turtle nesting in this region and is consistent with an extension of the present-day range of successful sea turtle nesting sites. The occurrence of a third site with an Australochelichnus-like trackway associated with a possible nest trace serves to underscore the importance of this area for the study of Pleistocene sea turtle paleobiology, also suggesting that their traces may be more common than previously supposed. As such traces of hatchling sea turtle trackways have both paleobiological and paleogeographic significance and utility in comparison with the traces of their extant descendants.
ACKNOWLEDGMENTS
We acknowledge the support of Xander Combrink, Guy Gardner, Linda Helm, Kei Heyns, Johan Huisamen, Christina Mars, Gerrit Mars, Peter Todd, and Boy van Rensburg. We thank Andre and Emily Brink for finding and reporting the trackway slab herein referred to as site 3 (Garden Route National Park area). Mark Dixon also reported a fourth turtle tracksite quite close to sites 1 and 2, for which he provided supporting photographic evidence. However, these tracks are at a difficult-to-access location and cannot be described here in detail without further field investigations. Linda Harris and Diane Le Gouvello kindly provided the photographs used in Figure 9. The photogrammetric image in Figure 6C was processed and provided courtesy of Carina Helm. The three-dimensional print of the A. agulhasii holotype (MGL 600/UCM 230.281) was made with the help of Luis Sanchez Vega (University of Colorado Denver), Neffra Matthews, National Operations Center (Denver, Colorado), Dinosaur Designs LLC (Golden, Colorado), and the offices of the University of Colorado Denver Dinosaur Trackers Research Group.