


1. Introduction
The Betic Cordillera (SE Spain) is part of the European Alpine Belt, which formed as a result of the relative movements of the African, Iberian and European plates (Weijermars, Reference Weijermars1987a, Reference Weijermars1991; Vergés, Fernàndez & Martínez, Reference Vergés, Fernàndez and Martínez2002; Fernández-Fernández et al. Reference Fernández-Fernández, Jabaloy-Sánchez, Nieto and González-Lodeiro2007). Uplift of the Betic Cordillera is related to the collision of the African and Iberian plates in Santonian/Campanian times (De Jong, Reference De Jong1990; Weijermars, Reference Weijermars1991; Vergés, Fernàndez & Martínez, Reference Vergés, Fernàndez and Martínez2002; Reicherter & Peters, Reference Reicherter and Peters2005). The NNE–SSW-elongated orogen can be subdivided into an External Zone (N), an Internal Zone (SE) and a Flysch Zone (SW; Fig. 1; Huibregtse et al. Reference Huibregtse, van Alebeek, Zaal and Biermann1998; Braga, Martín & Quesada, Reference Braga, Martín and Quesada2003; Reicherter & Peters, Reference Reicherter and Peters2005; Fernández-Fernández et al. Reference Fernández-Fernández, Jabaloy-Sánchez, Nieto and González-Lodeiro2007; Jabaloy-Sánchez, Fernández-Fernández & González-Lodeiro, Reference Jabaloy-Sánchez, Fernández-Fernández and González-Lodeiro2007). In consequence of the ongoing uplift of the Betic Cordillera during Neogene times, intermontane pull-apart basins developed in the External Zone (with occasional linkage to the Atlantic Ocean) and the Internal Zone (linked to the Mediterranean Sea; Fig. 1b and Fig. 2; Krijgsman et al. Reference Krijgsman, Fortuin, Hilgen and Sierro2001; Braga, Martín & Quesada, Reference Braga, Martín and Quesada2003; Puga-Bernabéu et al. Reference Puga-Bernabéu, Martín, Braga and Aguirre2014). The Neogene- to Quaternary-age basins of the Betic Cordillera include the Sorbas, Nijar and Vera basins (Sanz de Galdeano & Vera, Reference Sanz de Galdeano and Vera1992; Martin & Braga, Reference Martin, Braga, Friend and Dabrio1996; Martín et al. Reference Martín, Braga, Betzler and Brachert1996; Martín, Braga & Riding, Reference Martín, Braga and Riding1997; Krijgsman et al. Reference Krijgsman, Fortuin, Hilgen and Sierro2001; Braga, Martín & Quesada, Reference Braga, Martín and Quesada2003; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003; Puga-Bernabéu et al. Reference Puga-Bernabéu, Martín, Braga and Aguirre2014; Do Couto et al. Reference Do Couto, Gumiaux, Jolivet, Augier, Lebret, Folcher, Jouannic, Suc and Gorini2015) separated by metamorphic domes (c.f. Augier et al. Reference Augier, Agard, Monié, Jolivet, Robin and Booth-Rea2005; Pedrera et al. Reference Pedrera, Pérez-Peña, Galindo-Zaldívar, Azañón and Azor2009; Fig. 2) as well as by normal and strike-slip faults (e.g. the Palomares Fault Zone (PFZ) in the Vera Basin; Fig. 1b; Egeler & Simon, Reference Egeler and Simon1969; Bousquet, Reference Bousquet1979; García-Hernández et al. Reference García-Hernández, López-Garrido, Rivas, Sanz de Galdeano and Vera1980; Weijermars, Reference Weijermars1987b; Huibregtse et al. Reference Huibregtse, van Alebeek, Zaal and Biermann1998; Jonk & Biermann, Reference Jonk and Biermann2002; Faulkner, Lewis & Rutter, Reference Faulkner, Lewis and Rutter2003; Booth-Rea et al. Reference Booth-Rea, Azañón, Azor and Garcı́a-Dueñas2004; Martínez-Díaz & Hernández-Enrile, Reference Martínez-Díaz and Hernández-Enrile2004; Krijgsman et al. Reference Krijgsman, Leewis, Garcés, Kouwenhoven, Kuiper and Sierro2006). The tectonostratigraphic histories of the basins are broadly similar, while their evolution parallels that of the Alboran Sea (Banks & Warburton, Reference Banks and Warburton1991; Braga et al. Reference Braga, Martín, Riding, Aguirre, Sánchez-Almazo and Dinarès-Turell2006; Giaconia et al. Reference Giaconia, Booth-Rea, Martínez-Martínez, Azañón, Storti and Artoni2014; Mancilla et al. Reference Mancilla, Booth-Rea, Stich, Pérez-Peña, Morales, Azañón, Martin and Giaconia2015).

Figure 1. Geological overview of the Betic Cordillera: (a) location and orientation on the Iberian peninsula, including the main structural units of the orogen (e.g. External Zone, Internal Zone, Flysch Trough Units) and the Neogene-age basins (Atlantic or Mediterranean-linked basins); (b) detailed view of the western part of the Betic Cordillera, showing the main fault systems (CFZ – Crevillente Fault Zone; GF – Gafariollos Fault; PFZ – Palomares Fault Zone; CF – Carboneras Fault) and the Mediterranean-linked Neogene-age basins (from west to east): T – Tabernas Basin; N – Nijar Basin; S – Sorbas Basin; and V – Vera Basin. The sierras surrounding the basins are (from west to east): SG – Sierra Gador; SA1 – Sierra Alhamilla; SF – Sierra de los Filabres; SCG – Sierra del Cabo de Gata; SC – Sierra Cabrera; SA2 – Sierra Almagro; and SA3 – Sierra Almagrera (modified after Huibregtse et al. Reference Huibregtse, van Alebeek, Zaal and Biermann1998; Faulkner, Lewis & Rutter, Reference Faulkner, Lewis and Rutter2003; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003; Jabaloy-Sánchez, Fernández-Fernández & González-Lodeiro, Reference Jabaloy-Sánchez, Fernández-Fernández and González-Lodeiro2007; Mancilla et al. Reference Mancilla, Booth-Rea, Stich, Pérez-Peña, Morales, Azañón, Martin and Giaconia2015).
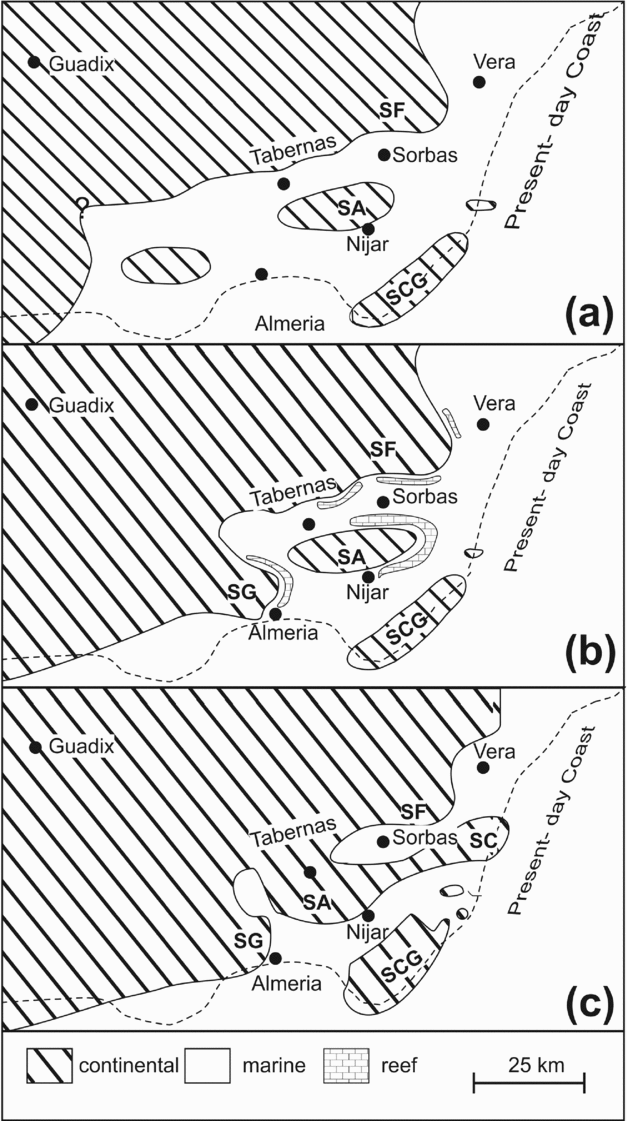
Figure 2. Palaeogeography of SE Spain, showing the development of the connection between the Neogene-age basins and the progressive uplift of the surrounding sierras during the deposition of the Abad Member: (a) latest Tortonian to earliest Messinian times, representing the setting during the accumulation of the older Abad Member deposits; (b) early Messinian times, including the middle to late Messinian-aged reefs; and (c) the end of the Messinian, i.e. subsequent to the reflooding at the end of the Messinian Salinity Crisis. Modified after Braga, Martín & Quesada (Reference Braga, Martín and Quesada2003) and Fortuin & Krijgsman (Reference Fortuin and Krijgsman2003). Abbreviations: SF – Sierra de los Filabres; SA – Sierra Alhamilla; SG – Sierra Gador; SCG – Sierra del Cabo de Gata; SC – Sierra Cabrera
The Vera Basin, located in the eastern part of the Betic Orogen, is connected to the south (via the Almocáizar Corridor) with the Sorbas Basin, and to the north (via the Pulpí Corridor) with the Lorca Basin. The sedimentary succession of the Vera Basin, comprises > 1 km of Burdigalian- to Pliocene-age deposits and consequently includes evidence of the Messinian Salinity Crisis (MSC) (Montenat et al. Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976; Bellón et al. Reference Bellón, Mosser, Roquin and Pardo1994; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995; Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Booth-Rea et al. Reference Booth-Rea, Azañón, Garcıá-Dueñas and Augier2003; Stokes, Reference Stokes2008). The succession can thus be broadly subdivided into pre- and post-MSC units (Völk & Rondeel, Reference Völk and Rondeel1964; Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Booth-Rea et al. Reference Booth-Rea, Azañón, Garcıá-Dueñas and Augier2003; Stokes, Reference Stokes2008). The succession records a dramatic deepening event (continental to deep marine) relatively early during the basin evolution. The aim of the present study is to investigate, in detail, the evidence for such a deepening event, via sedimentological fieldwork, and including detailed ichnological analysis, which has, to date, not yet been carried out within this area. Focusing on the corridor area (i.e. Almocáizar Corridor; Montenat et al. Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976) between the Sorbas and Vera basins, as well as the central area of the Vera Basin (located immediately to the north-east; Fig. 3a) allows the closely related histories of the basin successions (Vera, Sorbas) to be compared and contrasted, while also investigating the possible reasons why this event appears to be restricted to the Vera Basin.

Figure 3. (a) Geological map of the central and southern parts of the Vera Basin showing the main stratigraphic units (Burdigallian–Pleistocene) within the basin (modified after Bellón et al. Reference Bellón, Mosser, Roquin and Pardo1994; Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Braga et al. Reference Braga, Martín, Riding, Aguirre, Sánchez-Almazo and Dinarès-Turell2006; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003; Stokes, Reference Stokes2008; Booth-Rea et al. Reference Booth-Rea, Azañón, Azor and Garcı́a-Dueñas2004 and including the results of new fieldwork carried out as part of this study). (b, c) Detailed views of the sampling locations (I/II/III/IV). Broad sampling areas are marked by circles while the locations of the sedimentological profiles are marked by stars. Section coordinates: GPS – UTM European 1950, 30S, (1.1) 586677/4109472; (1.2) 588331/410947; (2.1) 596200/4116972; (2.2) 595914/4118305. PFZ – Palomares Fault Zone.
2. Pre-Messinian stratigraphy of the Vera, Sorbas & Nijar basins
As noted above, the Vera Basin is one of several Neogene-age basins in the Almeria area of SE Spain (Fig. 1b) which were closely linked during the early stages of their formation, to the extent that some of them (e.g. Vera, Nijar and Sorbas basins) could at this initial stage be considered as a single basin (albeit with localized sub-depocentres; Fig. 2a). Successive uplift of the Betic Cordillera led to increasing restriction, in terms of sediment movement and transport, between the Neogene-age basins, particularly from late Messinian times onwards (Fig. 2a–c; Völk & Rondeel, Reference Völk and Rondeel1964; Sanz de Galdeano, Reference Sanz de Galdeano1987; Benson & Bied, Reference Benson and Bied1991; Bellón et al. Reference Bellón, Mosser, Roquin and Pardo1994; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995; Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Booth-Rea et al. Reference Booth-Rea, Azañón, Garcıá-Dueñas and Augier2003; Braga, Martín & Quesada, Reference Braga, Martín and Quesada2003; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003). Owing to this level of interconnection, stratigraphic correlation (especially between the Sorbas, Nijar and Vera basins) of the older (Burdigalian – upper Messinian) sediments is possible. Subsequent uplift, however, resulted in increasing basin isolation leading to the development of considerable variations in the post-late Messinian sedimentary successions within the various basins (Fig. 2b, c; Sanz de Galdeano & Vera, Reference Sanz de Galdeano and Vera1992; Soria, Fernández & Viseras, Reference Soria, Fernández and Viseras1999; Baggley, Reference Baggley2000; Stokes & Mather, Reference Stokes and Mather2000; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003; Puga-Bernabéu et al. Reference Puga-Bernabéu, Martín, Braga and Aguirre2014; Do Couto et al. Reference Do Couto, Gumiaux, Jolivet, Augier, Lebret, Folcher, Jouannic, Suc and Gorini2015).
The pre-Messinian stratigraphy of the Sorbas, Vera and Nijar basins is summarized in Figure 4. The succession comprises five formations, namely, from base to top (i.e. the onset of the MSC succession): the Alamo, Gomara, Umbria, Chozas and Turre formations. The oldest described sediments (i.e. Alamo, Gomara and Umbria formations of Völk & Rondeel, Reference Völk and Rondeel1964) are found in the Sorbas and Vera basins (Sanz de Galdeano & Vera, Reference Sanz de Galdeano and Vera1992; Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Booth-Rea et al. Reference Booth-Rea, Azañón, Garcıá-Dueñas and Augier2003; Braga et al. Reference Braga, Martín, Riding, Aguirre, Sánchez-Almazo and Dinarès-Turell2006; Do Couto et al. Reference Do Couto, Gumiaux, Jolivet, Augier, Lebret, Folcher, Jouannic, Suc and Gorini2015). These comprise deformed continental conglomerates as well as shallow-marine conglomerates, sandstones and marls and are of Burdigallian – early Tortonian age. The continental conglomerates are locally derived and concentrated along the basin margins where they prograde into the marine areas where they become shallow-marine conglomerates. In contrast, the shallow-marine marls are widespread across the basinal areas, extending from the Sorbas Basin into the Vera Basin (Fig. 4; Jonk & Biermann, Reference Jonk and Biermann2002). Whether these sediments are also present within the Nijar Basin is unclear owing to a lack of outcrop.

Figure 4. (a) Schematic stratigraphic overview for the sedimentary succession of the pre-Messinian Salinity Crisis (Burdigalian to the end of Messinian times) sedimentary succession of the Vera Basin, showing the relationship between the individual formations and, in detail, the correlation of the three members of the Turre Formation (i.e. Azagador, Abad and Cantera members). (b) Comparison of the stratigraphy of the Vera Basin with the adjacent Sorbas and Nijar basins. Note the occurrence of the same formations/members within these adjoining basins. Modified after Montenat et al. (Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976), Braga, Martín & Wood (Reference Braga, Martín and Wood2001), Fortuin & Krijgsman (Reference Fortuin and Krijgsman2003), Puga-Bernabéu et al. (Reference Puga-Bernabéu, Martín, Braga and Aguirre2014) and including the results of field mapping from this study.
The basal sedimentary succession is unconformably overlain by the lower Tortonian-age Chozas Fm (Fig. 4; Völk & Rondeel, Reference Völk and Rondeel1964; Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Booth-Rea et al. Reference Booth-Rea, Azañón, Azor and Garcı́a-Dueñas2004), which comprises marls, limestones and sandstones and represents a deepening sequence within the Neogene-age basin infill (Martín et al. Reference Martín, Braga, Betzler and Brachert1996; Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Jonk & Biermann, Reference Jonk and Biermann2002). The unconformably overlying upper Tortonian to upper Messinian-age Turre Fm (Fig. 4; Martín et al. Reference Martín, Braga, Betzler and Brachert1996; Braga & Martín, Reference Braga, Martín, Riding and Awramik2000; Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Krijgsman et al. Reference Krijgsman, Fortuin, Hilgen and Sierro2001; Sierro et al. Reference Sierro, Hilgen, Krijgsman and Flores2001; Jonk & Biermann, Reference Jonk and Biermann2002; Booth-Rea et al. Reference Booth-Rea, Azañón, Azor and Garcı́a-Dueñas2004) comprises three members. The first two of these are the Azagador Member, a proximal, shallow-marine unit containing siliciclastic and carbonate sediments, and the closely related reef complex (Cantera Member), which is best developed in late Messinian times, and is particularly well exposed in the Sorbas and Nijar basins (Fig. 4; Sanz de Galdeano & Vera, Reference Sanz de Galdeano and Vera1992; Martin & Braga, Reference Martin, Braga, Friend and Dabrio1996; Martín et al. Reference Martín, Braga, Betzler and Brachert1996; Martín, Braga & Riding, Reference Martín, Braga and Riding1997; Krijgsman et al. Reference Krijgsman, Fortuin, Hilgen and Sierro2001; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003; Puga-Bernabéu et al. Reference Puga-Bernabéu, Martín, Braga and Aguirre2014). Within the Vera Basin, the Cantera member is only exposed in a restricted area in the north-west (Fig. 3). These two members (Azagador and Cantera Member) interdigitate with the distal marls and intercalated sandstones of the Abad Member.
The development of transgressive successions in the upper part of the Turre Fm was related to sea-level variations, which commenced at the Tortonian–Messinian boundary, coupled with a coeval increase in tectonic activity (Fig. 4). This latter event was related to uplift of the sierras surrounding the basins in the region (Fig. 2c; Braga, Martín & Quesada, Reference Braga, Martín and Quesada2003; Booth-Rea et al. Reference Booth-Rea, Azañón, Azor and Garcı́a-Dueñas2004) as well as increased activity along the strike-slip fault systems. The end result was a phase of rapid basin subsidence as well as increasing isolation of the Sorbas and Nijar basins, both from the Mediterranean Sea as well as from the Vera Basin. As a consequence of these changes, the Azagador and Abad members became increasingly lithologically disparate between the Sorbas/Nijar and Vera basins (Cloetingh, Reference Cloetingh, Müller, McKenzie and Weissert1991; Baggley, Reference Baggley2000; Krijgsman et al. Reference Krijgsman, Fortuin, Hilgen and Sierro2001; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003; Braga et al. Reference Braga, Martín, Riding, Aguirre, Sánchez-Almazo and Dinarès-Turell2006).
The sediments of the Abad Member (Vera Basin) form the basis for this study. Within the basin, the succession comprises 200–300 m of grey silty marls intercalated with sandstone beds (Fig. 4–6). The percentage of sandstone beds increases upwards, thus roughly subdividing the member into a sand-poor lower part (sandstone:marl = 1:10) and a sand-rich upper part (1:2). This lithological subdivision of the Abad Member correlates broadly with the foraminiferal assemblages present, with Globorotalia mediterranea and Globorotalia multiloba present in the lower part, and Neogloborotalia acostaensis in the upper part (Sierro et al. Reference Sierro, Flores, Zamarreño, Vázquez, Utrilla, Francés, Hilgen and Krijgsman1999; Baggley, Reference Baggley2000). Booth-Rea et al. (Reference Booth-Rea, Azañón, Azor and Garcı́a-Dueñas2004) have suggested that there is an unconformity (related to tectonic activity) present within the member (although the precise location of this, if present, is unclear).
The sand-rich part of the Abad Member (i.e. Santiago Beds; Völk & Rondeel, Reference Völk and Rondeel1964; Benson & Bied, Reference Benson and Bied1991; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995) is considered to represent a marker unit deposited prior to the onset of the MSC. In the Vera Basin, the Santiago Beds are interpreted as turbidites based largely on the presence of Bouma sequences (Völk & Rondeel, Reference Völk and Rondeel1964; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995; Huibregtse et al. Reference Huibregtse, van Alebeek, Zaal and Biermann1998) as well as the ichnofossil Paleodictyon.
In the Sorbas and Nijar basins, the Abad Member can also be subdivided into two units based on foraminiferal biostratigraphy as well as sedimentary characteristics (Martín et al. Reference Martín, Braga, Betzler and Brachert1996; Krijgsman et al. Reference Krijgsman, Fortuin, Hilgen and Sierro2001; Sierro et al. Reference Sierro, Hilgen, Krijgsman and Flores2001; Braga, Martín & Quesada, Reference Braga, Martín and Quesada2003; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003). During the deposition of the Abad Member, palaeoenvironmental conditions were stable with the relatively low energy system (Krijgsman et al. Reference Krijgsman, Hilgen, Raffi, Sierro and Wilson1999, Reference Krijgsman, Fortuin, Hilgen and Sierro2001; Baggley, Reference Baggley2000; Sierro et al. Reference Sierro, Hilgen, Krijgsman and Flores2001; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003). In contrast, the deposition of the Upper Abad Member occurred during a period of fluctuating salinity accompanied by the deposition of sapropels and slump activity all of which suggest ongoing restriction between the basins (i.e. Sorbas/Nijar and Vera) and the Mediterranean Sea (Fig. 4, Krijgsman et al. Reference Krijgsman, Hilgen, Raffi, Sierro and Wilson1999, Reference Krijgsman, Fortuin, Hilgen and Sierro2001; Sierro et al. Reference Sierro, Flores, Zamarreño, Vázquez, Utrilla, Francés, Hilgen and Krijgsman1999, Reference Sierro, Hilgen, Krijgsman and Flores2001).
The subsequent phase of deposition, which included the MSC, differs markedly between the Sorbas/Nijar and Vera basins as a result of their increasing separation Fig. 2c and the development of individual palaeogeographical settings (e.g. Riding et al. Reference Riding, Braga, Martín and Sánchez-Almazo1998; Krijgsman et al. Reference Krijgsman, Hilgen, Raffi, Sierro and Wilson1999; Sierro et al. Reference Sierro, Flores, Zamarreño, Vázquez, Utrilla, Francés, Hilgen and Krijgsman1999, Reference Sierro, Hilgen, Krijgsman and Flores2001; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003; Augier, Jolivet & Robin, Reference Augier, Jolivet and Robin2005; Braga et al. Reference Braga, Martín, Riding, Aguirre, Sánchez-Almazo and Dinarès-Turell2006; Manzi et al. Reference Manzi, Gennari, Hilgen, Krijgsman, Lugli, Roveri and Sierro2013). In the Vera Basin, the MSC is mainly preserved as an erosional hiatus (Benson & Bied, Reference Benson and Bied1991; Bellón et al. Reference Bellón, Mosser, Roquin and Pardo1994; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995), while in the Sorbas and Nijar basins the period is represented by the deposition of extensive gypsum units (Yesares Member; Martín, Braga & Riding, Reference Martín, Braga and Riding1997; Conesa & Babinot, Reference Conesa and Babinot1999; Krijgsman et al. Reference Krijgsman, Hilgen, Raffi, Sierro and Wilson1999, Reference Krijgsman, Fortuin, Hilgen and Sierro2001; Jonk & Biermann, Reference Jonk and Biermann2002; Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003; Braga et al. Reference Braga, Martín, Riding, Aguirre, Sánchez-Almazo and Dinarès-Turell2006; Do Couto et al. Reference Do Couto, Gumiaux, Jolivet, Augier, Lebret, Folcher, Jouannic, Suc and Gorini2015).
3. Description of the studied succession
The uppermost Tortonian- to Messinian-aged Abad Member crops out in the southern and central parts of the Vera Basin (Fig. 3a). The sediments in the southern part of the basin are stratigraphically older than those in the central parts. Four representative sedimentary profiles, two from each area, were selected for detailed study (Fig. 3b, c). The sediments of the Abad Member comprise marls with interbedded sandstones. The former is the dominant lithology within the member, while the coarser-grained sandstones are present throughout, although there are marked changes both in frequency and bed thickness. In general, the stratigraphically younger part of the succession contains relatively greater amounts of sandstone (i.e. Santiago Beds, see above).
3.a. Lithologies
3.a.1. Marl
As noted above, marls are the dominant lithofacies of the Abad Member (up to 70%). This lithofacies comprises a grey-coloured, apparently homogenous, and well-sorted mixture of carbonate and siliciclastic material, with individual beds ranging in thickness from 0.5–20 cm. The grain size ranges from clay to silt (classified after Wentworth, Reference Wentworth1922). In addition, the marls in the southern part of the Vera Basin are coarser (mainly silt) than in the central part (dominantly clay). The siliciclastic fraction (c. 50%) comprises quartz and clay minerals as well as up to 10% of mica (dominantly muscovite), while the carbonate fraction consists mainly of micrite and microfossils. The latter are predominantly foraminifera, such as Globorotalia mediterranea (Benson & Bied, Reference Benson and Bied1991; Baggley, Reference Baggley2000; Braga, Martín & Wood, Reference Braga, Martín and Wood2001). Furthermore, isolated brachiopods (e.g. Maltaja pajaudi) and fragments of red algae were noted. Internally, rare parallel and wavy lamination was observed (< 10% of the entire succession). However, the presence of diffuse microbioturbation in some areas would suggest that primary sedimentary structures were originally present but destroyed as a result of biological activity. Occasionally, the marls show a darker grey-coloured amorphous pattern, which is made up of concentrations of several dark-grey spots (c. 1 mm in diameter), suggesting an enrichment in organic material.
3.a.2. Sandstone
Light-grey to beige-coloured beds of coarse- to fine-grained sandstones, ranging in thickness from 0.5–20 cm, are interbedded within the marls. The coarse- to medium-grained sandstones tend to crop out more frequently in the southern part of the Vera Basin, while the central part of the basin is characterized more by the medium- to fine-grained sandstones. The sandstones comprise a mixture of siliciclastic and carbonate material.
The coarser-grained sandstone beds are light grey in colour and graded with the grain size ranging from coarse grained (10%) through to medium grained (60%) and on to fine grained (30%). In addition to the internal grading, cross- and parallel-lamination were also observed. These internal structures can be classified according to the scheme of Bouma (Reference Bouma1962). Thus, the sandstones are predominantly Tbc (c. 40%) and Tab (c. 30%), while Tac (c. 20%), Tad (c. 5%) and Ta (c. 5%) were also recognized. Bed bases show evidence of loading and flute casts (270°, n = 7). Bed tops show evidence of current ripples (Tc). The coarse- to medium-grained sandstones are dominated by the siliciclastic fraction (up to 65%), mainly quartz, mica, lithic fragments (dominantly mica schist and carbonates) and clay minerals. The remaining 35% of the sandstones comprises micrite, shell fragments and foraminifera.
The finer-grained sandstone beds are light grey to beige in colour and graded, ranging from medium grained (60%) through to fine grained (40%). Bed thickness varies from 1–15 cm. Internal grading as well as cross- and parallel-lamination were observed. The sandstones are predominantly Tac (c. 30%), while Ta (15%), Tad (c. 10%), Tbc (10%) and Tcd (5%) were also recognized. Bed bases show evidence of loading while the bed tops are often rippled (Tc). The finer- and coarser-grained beds are lithologically similar, although the former show higher percentages of carbonate content (up to 50%), as well as an enrichment of clay within the clastic fraction, resulting in the beige colour of the sediments.
3.b. Sedimentary profiles
Four representative sedimentary sections were measured in the deposits of the Abad Member within the study area. Two of these are older, cf from a stratigraphically lower location in the Almocáizar Corridor area (Fig. 5), while two sections from the basin centre are younger, cf from a stratigraphically higher position (Fig. 6).

Figure 5. Measured sections of the Abad Member succession. 1.1 and 1.2 – sections within the Almocáizar Corridor (i.e. older Abad deposits). Grain size: M – mud; S – silt; Sst – sandstone (f – fine; m – medium; c – coarse; vc – very coarse). For locations see Figure 3.

Figure 6. Measured sections of the Abad Member succession. 2.1 and 2.2 – sections from the basin centre (i.e. 2.1 younger Abad deposits and 2.2 Lago Mare deposits). Grain size: M – mud; S – silt; Sst – sandstone (f – fine; m – medium; c – coarse; vc – very coarse). For locations see Figure 3.
3.b.1. Sections 1.1 and 1.2 (Fig. 5)
The older profiles (Figs 4a,b; 5) are characterized by thick beds (ranging from 0.07–8 m, but predominantly c. 0.7 m) of silt-rich marl, alternating with coarser-grained sandstone beds. These latter vary in thickness, from 5–20 cm (generally c. 8 cm; c. 60%). The boundaries (bases and tops) with the intercalated marls are very sharp and slightly wavy to straight. The frequency of the sandstone beds varies, and this can be related to the thickness of the marl beds, varying from several metres (e.g. 3.8 m thick bed in Fig. 5) to tens of centimetres.
3.b.2. Sections 2.1 and 2.2 (Fig. 6)
The marls and sandstone sediments of the younger profiles (Figs 4a,c; 6) in the central part of the basin comprise mainly finer-grained material. In addition, the marls in this area are more beige in colour as a result of the higher content of clay minerals. In section 2.1, the frequency of the interbedded sandstone beds is higher than in the older profiles (Fig. 5; 1.1 and 1.2). Marl thickness is generally c. 30 cm (range 1–90 cm). The boundaries of the medium- to fine-grained, normally graded sandstones are very sharp and slightly wavy and, occasionally (c. 10%), the tops grade into the overlying marl. Trace fossils are mainly preserved on the bases of the sandstone beds.
In section 2.2, there is an increase in the sandstone bed frequency, while the marl beds are mainly c. 20 cm thick (ranging from 5–55 cm). Additionally, the sandstones are generally finer grained than in the other measured profiles in the region. In this section no ichnofossils were observed, nor were any ichnofossils noted in the surrounding area (e.g. weathered outcrops).
4. Systematic ichnology
The ichnofossil assemblage of the Vera Basin is presented in the following section in alphabetical order. Preservational terminology follows that adopted by Ekdale (Reference Ekdale, Ekdale, Bromley and Pemberton1984) and Knaust (Reference Knaust, Knaust and Bromley2012), while the internal structures of sandstone beds use the terminology of Bouma (Reference Bouma1962), Collinson, Mountney & Thompson (Reference Collinson, Mountney and Thompson2006) and McCann & Manchego (Reference McCann and Manchego2015).
The ichnofossils were observed and collected from two main outcrop areas in the Vera Basin: the older one in the southern part of the basin (Almocáizar Corridor) and the younger one in the central part (Fig. 3). In the stratigraphically lower area (i.e. Almocáizar Corridor, three locations (I, II and III in Fig. 3b) were examined in detail, while in the younger, area (i.e. Basin Centre) work was concentrated in one location (section 2.1 in Fig. 3c). In all four of these locations, suites of ichnofossils were collected for subsequent examination. Uncollectable samples were examined and photographed in the field.
Ichnogenus Circulichnis Vialov, Reference Vialov1971
Circulichnis montanus Vialov, Reference Vialov1971 by original designation
Figures 7e, 8c
Material. Five specimens and numerous observations in the field.
Description. Smooth ellipsoidal traces (both partial and complete ellipses) preserved in convex hyporelief on the soles of 1.5–1.8 cm thick, medium-grained sandstone beds with internal cross-lamination (Tac). Burrows are cylindrical, unlined. Infill is identical to that of the host rock. Diameters of the long axes vary between 10.7 and 14.6 mm and of the short axes between 0.5 and 10.3 mm.

Figure 7. Ichnofossils from the Abad Member, Vera Basin. (a) View of the base of a 2 cm thick, medium-grained sandstone bed showing a variety of ichnofossils preserved in convex hyporelief. These are viewed in more detail in (b) and (c). (b) Desmograpton dertonensis (Dd) and Paleodictyon minimum, showing both the deep (Pmin-d) and shallow (Pmin-s) forms; (c) deep form of Paleodictyon minimum (Pmin-d) and D. dertonensis (Dd). (d) D. ichthyforme (Di), Helminthopsis tenuis (Ht) and Paleodictyon minimum preserved as shallow (Pmin-s) and deep (Pmin-d) form in convex hyporelief on the base of a 2 cm thick, medium-grained sandstone. (e) Glockerichnus alata (G) and an incomplete specimen of Circulichnis montanus (C) preserved in convex hyporelief on the base on the base of a 1.8 cm thick, medium-grained sandstone. Scale bar = 1 cm.

Figure 8. Ichnofossils from the Abad Member, Vera Basin. (a) Helminthoida labyrinthica (Hl) and Paleodictyon miocenicum (Pmi) preserved in convex hyporelief on the base of a 1.8 cm thick, medium-grained sandstone. (b) Phycodes bilix (Ph) and Palaeophycus tabularis preserved in convex hyporelief on the base of a 1.1 cm thick, medium-grained sandstone. (c) Circulichnis montanus (C) and Helminthoida labyrinthica (Hl) preserved in convex hyporelief on the base of a 1.8 cm thick, medium-grained sandstone. (d) Thalassinoides suevicus (T) preserved in convex hyporelief on the base of a 3 cm thick, medium-grained sandstone. (e) Helminthoida labyrinthica (Hl) and Mammillichnis aggeris? (M) preserved in convex hyporelief on the base of a 2.9 cm thick, medium-grained sandstone. (f) Helminthoida labyrinthica (Hl) and Helminthopsis tenuis (Ht) preserved in convex hyporelief on the base of a 3.4 cm thick, medium-grained sandstone. (g) Palaeophycus herberti (Ph) and Palaeophycus tabularis (Pt) preserved in convex hyporelief on the base of a 2 cm thick, medium-grained sandstone. Scale bar = 1 cm.
Remarks. The samples of the Vera Basin conform well to those of McCann (Reference McCann1993) and Fillion & Pickerill (Reference Fillion and Pickerill1984), who discussed the original description of Vialov (Reference Vialov1971) in detail and suggested that C. montanus is a post-depositional burrow.
Ichnogenus Desmograpton Fuchs, Reference Fuchs1895
Desmograpton dertonensis Sacco, Reference Sacco1888
Figure 7b, c
Material. Three specimens and rarely noted in the field.
Description. Narrow J-shaped semi-meanders (1.2–1.5 mm in diameter of the meander) with parallel to sub-parallel connecting bars. Burrows are up to 0.8 mm wide. The ichnofossils are preserved on the bases of up to 2.5 cm thick, medium-grained sandstones with internal grading and parallel-lamination (Tab).
Remarks. As noted by Uchman (Reference Uchman1995, Reference Uchman1998), the ichnotaxonomic and ichnospecies revisions of Desmograpton are problematic. His revisions helped to clarify some of the extant problems, but have also resulted in new ones. For example, both Desmograpton dertonensis and D. alternum contain U- or J-shaped semi-meanders according to the descriptions of Uchman (Reference Uchman1998). Indeed, the only difference between them is the presence of the connecting bars, which according to Uchman (Reference Uchman1995) are parallel or sub-parallel in D. dertonensis, while in D. alternum (= D. geometricum of Książkiewicz, Reference Książkiewicz1977) they are oblique and form a zig-zag pattern (Uchman, Reference Uchman1995, Reference Uchman1998). However, the illustrations provided by Uchman (Reference Uchman1998) of both ichnospecies differ from those illustrated by Seilacher (Reference Seilacher, Crimes and Harper1977) whose D. geometricum (= D. alternum of Uchman, Reference Uchman1998, see his fig. 97) appears to be quite different. Seilacher's (Reference Seilacher, Crimes and Harper1977) D. inversum most closely resembles the current sample, termed D. dertonensis by Uchman (Reference Uchman1998) after the original illustration by Sacco (Reference Sacco1888). Thus, while the ichnogenus is still clearly in need of taxonomic revision, the current designation as D. dertonensis fits best with the observed features of the samples.
Desmograpton ichthyforme Macsotay, Reference Macsotay1967
Figures 7d, 9f, 10c
Material. Five specimens and rarely noted in the field.
Description. Smooth burrow system preserved in convex hyporelief at the base of 1–2.5 cm thick, fine- and medium-grained sandstones which are internally parallel- and cross-laminated (Tab, Tbc, Tcd). Length of the structures range from 1–2.5 cm, with burrow diameters ranging from 0.2–1.5 mm. The narrow, roughly straight and sub-parallel closely spaced (1 mm) string-sized burrows show a clear swelling in the median zone of the individual strings. Occasional transverse links (0.2–1 mm in length) were observed.
Remarks. The samples of the Vera Basin conform to the description of Desmograpton fuchsi as described by Książkiewicz (Reference Książkiewicz1977) and McCann (Reference McCann1989), and classified as D. ichthyforme by Seilacher (Reference Seilacher, Crimes and Harper1977). The latter author noted the pronounced length of the transverse elements, although this need not be the case (McCann, Reference McCann1989). The partial review of Desmograpton undertaken by Uchman (Reference Uchman1995) noted that the name D. ichthyforme takes preference over D. fuchsi. Although, the transverse links between the burrows in the current examples are barely preserved, the characteristic swelling of the medial zone is present allowing the specimen to be clearly classified as D. ichthyforme (cf. Książkiewicz, Reference Książkiewicz1977)
Ichnogenus Glockerichnus Pickerill, Reference Pickerill1982
Glockerichnus alata Seilacher, Reference Seilacher, Crimes and Harper1977
Figure 7e
Material. One specimen.
Description. The 18.1 mm bilateral structure is made up of radially elongated ridges (1.2–1.9 mm in diameter). These ridges are made up of 6 mm wide U-shaped burrows. The smooth burrows are preserved in convex hyporelief on the base of a 1.8 cm thick, medium-grained and internally cross-laminated (Tc) sandstone bed with a rippled top.
Remarks. As noted by Weber et al. (Reference Weber, Hu, Steiner and Zhao2012), several Glockerichnus ichnospecies have been described (e.g. Książkiewicz, Reference Książkiewicz1968, Reference Książkiewicz1977; Seilacher, Reference Seilacher, Crimes and Harper1977). All of these ichnospecies show the characteristic stellate morphology of Glockerichnus where numerous ribs run radially from a diffuse central field, often with dichotomous branching along the distal parts of some of the larger main ribs (Uchman, Reference Uchman1998). Specimens of Glockerichnus range from c. 15 cm (some of the larger Carpathian examples, e.g. Glockerichnus glockeri, Książkiewicz, Reference Książkiewicz1977) down to 2–3 cm (e.g. ‘Glockeria’ parvula = Glockerichnus parvula, Książkiewicz, Reference Książkiewicz1968). The current example, however, contains distinct loops in the central part of the trace, thus more closely resembling Glockerichnus alata (= Glockeria alata Seilacher, Reference Seilacher, Crimes and Harper1977). The main problem, however, is the relative size of the specimens described by Seilacher (Reference Seilacher, Crimes and Harper1977) and figured in Seilacher (Reference Seilacher2007). In detail, the ribs are described as having diameters of up to 7 mm, with the entire burrow system having a diameter of up to 50 cm. The current example is considerably smaller. Thus, the assignation may represent an ontogenetic variation of the specimens described by Seilacher (Reference Seilacher, Crimes and Harper1977) or another ichnospecies. At the present time, however, we choose to suggest that the recorded ichnospecies belongs to G. alata.
Ichnogenus Helminthoida Schaufhäutl, Reference Schafhäutl1851
Helminthoida labyrinthica Heer, Reference Heer1865
Figure 8a, c, e, f
Material. Three specimens and numerous observations in the field.
Description. Smooth meandering burrow preserved in convex hyporelief on the base of 1.9–3.2 cm thick, medium- and fine-grained sandstones, which are internally graded and parallel-laminated (Ta, Tb). The burrow diameters range from 1.3–1.5 mm, and loop separation is 1–3 mm. The burrows show evidence of weak coiling.
Remarks. Helminthoida is a variable ichnogenus (Książkiewicz, Reference Książkiewicz1977) with closely spaced and parallel burrows (Crimes & Anderson, Reference Crimes and Anderson1985), although they can be extremely variable (Häntzschel, Reference Häntzschel and Teichert1975). The observed variety led Książkiewicz (Reference Książkiewicz1977) to subdivide the ichnogenus into two groups, although it is doubtful whether such a subdivision is necessary. Additionally, Seilacher (Reference Seilacher, Crimes and Harper1977) erected the new ichnogenus Helminthorhaphe to separate Helminthoida crassa Schafhäutl, Reference Schafhäutl1851 from H. labyrinthica, which he considered to be preservational variations. Both Crimes, Goldring & Homewood (Reference Crimes, Goldring and Homewood1981) and Crimes & Anderson (Reference Crimes and Anderson1985) have discussed this new ichnogenus and deemed it unnecessary. Uchman (Reference Uchman1998), in a detailed revision of the collection of Marian Książkiewicz, also noted the problematic nature of the ichnogenus, and assigned H. labyrintica to Nereites irregularis Schafhäutl, Reference Schafhäutl1851. However, Nereites is clearly defined as having a median, back-filled tunnel (core) enveloped by an even to lobate zone of reworked sediment, with commonly only the external part of the mantle being preserved as a densely packed chain of uni- or multi-serial depressions or pustules (Uchman, Reference Uchman1995, Reference Uchman1998). A close reading of the description of Nereites irregularis Schafhäutl, Reference Schafhäutl1851 reveals that the burrow comprises a light-coloured core and an indistinct backfill feature, both of which are clearly absent in the present samples, suggesting that the smooth meandering trace of Helminthoida is clearly not a Nereites. Additionally, Uchman (Reference Uchman1998) notes that Nereites irregularis occurs mainly in deep-marine, often calcareous planktonic sediments (Seilacher, Reference Seilacher, Nitecki and Kitchell1986), which is clearly not the case in the present samples, which are demonstrably turbiditic and siliciclastic. The problematic nature of the ichnogenus, however, remains. In the absence of any real synonymy with Nereites and the absence of any real reason to use Helminthorhaphe, we prefer to use the general term Helminthoida until such time as the genus can be fully revised.
Ichnogenus Helminthopsis Heer, Reference Heer1877
Helminthopsis tenuis Książkiewicz, Reference Książkiewicz1968
Figures 7d, 8f
Material. Two specimens and rarely noted in the field.
Description. Simple, unbranched, elongate simple tubes (0.9 mm in diameter) with wide, shallow meanders up to 1 cm wide, and narrow but obtuse meanders, up to 4 cm wide. The burrow depth varies from 0.1–0.3 mm. The narrow meanders are slightly deeper than the wider ones.
Remarks. Distinguishing between the two ichnospecies Helminthopsis tenuis and H. abeli is difficult. Han & Pickerill (Reference Han and Pickerill1995) considered them synonyms, and in their emended diagnosis noted that the winding style should be considered the main distinguishing feature between ichnospecies of Helminthopsis. Uchman (Reference Uchman1998) has noted that specimens of H. tenuis generally display wide, shallow meanders and deeper, narrow but obtuse meanders, in comparison to the bulging, horseshoe-shaped meanders exhibited by H. abeli. The variations in meander form exhibited by the present specimens suggest that they should be classified as H. tenuis.
Ichnogenus Mammillichnis Chamberlain, Reference Chamberlain1971
Mammillichnis aggeris? Chamberlain, Reference Chamberlain1971
Figure 8e
Material. Four specimens and rarely noted in the field.
Description. Hypichnial elliptical mound with a depression in its centre preserved in hypichnial relief on the bases of 1.5–2.0 cm thick laminated sandstones (Ta, Tab, Tb). The structures are 2.8–3.0 mm wide and 2 mm in height.
Remarks. According to Uchman (Reference Uchman1998) Mammillichnis aggeris is the upper part of a thin vertical or sub-vertical cylindrical burrow. While the elliptical structure, including a central depression, of the samples within the Vera Basin coincides well with the description of Książkiewicz (Reference Książkiewicz1977) and Uchman (Reference Uchman1998), the size seems to be more problematic. The present specimens are markedly smaller in size than those described by the aforementioned authors, who describe specimens of Mammillichnis aggeris ranging from 8–10 mm in diameter and 3–4 mm in height. The present specimens are thus smaller than those described by Książkiewicz (Reference Książkiewicz1977) and Uchman (Reference Uchman1998), but in terms of their morphology, they conform well with both the ichnogenus and ichnospecies.
Ichnogenus Palaeophycus Hall, Reference Hall1847
Palaeophycus herberti Saporta, Reference Saporta1872
Figure 8g
Material. One specimen and rarely noted in the field.
Description. Smooth cylindrical burrow preserved in full relief on top of a 2 cm thick graded, medium-grained sandstone, with internal parallel- and cross-lamination (Tbc). The burrow, which is straight, is 2.9 mm in height and 4.2 mm in diameter, with the cylindrical inner part eroded and partly collapsed; the central tube is 1.9 mm in diameter
Remarks. The specimen corresponds well with the description of Pemberton & Frey (Reference Pemberton and Frey1982) and Blissett & Pickerill (Reference Blissett and Pickerill2004); the internal tube of the specimen is, as noted above, partly collapsed and owing to the poor preservation difficult to distinguish.
Palaeophycus serratus McCann, Reference McCann1993
Figure 10e
Material. Two specimens and rarely noted in the field.
Description. Straight to slightly winding, lined burrows preserved in convex and concave hyporelief on the base of a 1.7 cm thick, medium-grained sandstone, with internal grading, cross-lamination and with ripples preserved at the top (Tac). The burrows are 4.5–5.3 mm in diameter with an annulation of a maximum 2 per millimetre.
Remarks. The present specimen corresponds well with the description of McCann (Reference McCann1993), who distinguished P. serratus from other ichnospecies of Palaeophycus. Characteristic for this ichnospecies is the presence of a transverse annulation, which is clearly and consistently observable in both specimens from the Vera Basin.
Palaeophycus tabularis Hall, Reference Hall1847
Figures 8g, 9a, 10c
Material. Thirty-eight specimens and numerous observations in the field.
Description. Straight to slightly winding, unbranched and smooth burrows on the base of 1.0–2.9 cm thick sandstones, which are medium to very fine grained and internally parallel- and cross-laminated (Tab, Tac, Tacd, Tbc, Tc). The burrows, which are preserved in convex hyporelief, range from 0.7–2.9 mm in diameter and from 0.37 cm to 4.3 cm in length. The burrow fill is identical to that of the host rock; no internal structures are observable.

Figure 9. Ichnofossils from the Abad Member, Vera Basin. (a) Palaeophycus tabularis (Pt) and Paleodictyon (Glenodictyon) miocenicum (Pmi) preserved in convex hyporelief on the base of a 2.5 cm thick, medium-grained sandstone. (b) Paleodictyon (Glenodictyon) maximum (Pmax) preserved in convex hyporelief on the base of a 3.4 cm thick, medium-grained sandstone. (c) Paleodictyon (Glenodictyon) gomezi (Pg) preserved in convex hyporelief on the base of a 1.5 cm thick, medium-grained sandstone. (d) Paleodictyon intermedium (Pi) preserved in convex hyporelief on the base of a 2.4 cm thick, medium-grained sandstone. (e) Paleodictyon (Glenodictyon) majus (Pm) preserved in convex hyporelief on the base of a 1.1 cm thick, medium-grained sandstone. (f) Desmograpton ichthyforme (Di), Palaeophycus tabularis (Pt) and Paleodictyon (Glenodictyon) majus (Pm) preserved in convex hyporelief on the base of a 2 cm thick, medium-grained sandstone. Scale bar = 1 cm.

Figure 10. Ichnofossils from the Abad Member, Vera Basin. (a) Urohelminthoida dertonensis (U) preserved in convex hyporelief on the base of a 2.2 cm thick, medium-grained sandstone. (b) Scolicia prisca (Spr) preserved in convex hyporelief on the base of a 2 cm thick, medium-grained sandstone. (c) Palaeophycus tabularis (Pt) and Desmograpton ichthyforme (Di) preserved in convex hyporelief on the base of a 1 cm thick, medium-grained sandstone. (d) Scolicia plana (Spli) preserved in convex hyporelief on the base of a 2.2 cm thick, medium-grained sandstone. (e) Palaeophycus serratus preserved in convex (Ps-v) and concave (Ps-c) hyporelief on the base of a 1.7 cm thick, medium-grained sandstone. Scale bar = 1 cm.
Remarks: According to Pemberton & Frey (Reference Pemberton and Frey1982), burrows of Palaeophycus tabularis can be characterized as branched or unbranched, smooth or ornamented, lined, cylindrical and mainly horizontal trace fossils. This rather broad description demonstrates the diversity of this particular ichnofossil, and is consistent with a variety of specimens in the samples collected from the Vera Basin. Palaeophycus is a eurybathic facies-crossing ichnogenus, produced probably by polychaetes (after Pemberton & Frey, Reference Pemberton and Frey1982).
Ichnogenus Paleodictyon Meneghini in Savi & Meneghini, Reference Savi and Meneghini1851
Subichnogenus Glenodictyon van der Marck, Reference van der Marck1863
Diagnosis. Three-dimensional network ichnofossil consisting of a horizontal net composed of regular to irregular hexagonal meshes and vertical outlets (after Uchman, Reference Uchman1995, Reference Uchman1998).
Paleodictyon (Glenodictyon) gomezi Azpeitia Moros, Reference Azpeitia Moros1933
Figure 9c
Material. Three specimens and numerous observations in the field.
Description. Smooth, irregular hexagonal (sometimes pentagonal), very large network preserved in convex hyporelief on the bases of 1.6–2.75 cm thick and internally graded medium- to fine-grained, structureless, parallel- and cross-laminated (Tab, Tac, Tbc) sandstone beds. The strings are 2.0–2.8 mm in diameter and the mesh size ranges from 10.5–20.5 cm.
Remarks. The specimens correspond well with the description of P. gomezi of Wetzel (Reference Wetzel2000), Uchman (Reference Uchman1998) and Crimes & McCall (Reference Crimes and McCall1995) and, in addition, are very similar to the specimens described by Crimes (Reference Crimes, Crimes and Harper1977; e.g. his pl. 4c, e, p. 85). According to Wetzel (Reference Wetzel2000) the penetration depth of this very large type of Paleodictyon is distinctly deeper than the normal shallow burrowing depth of Paleodictyon, suggesting a positive correlation between the size and the penetration depth of this ichnogenus. This trend corresponds well with the present samples, since P. gomezi represents the highest relief in comparison with other traces on the same slab. The maximum depth of P. gomezi of the collected samples is 0.58 mm and hence, noticeably greater than any other collected Paleodictyon specimens from the Abad Member.
Paleodictyon (Glenodictyon) intermedium Książkiewicz, Reference Książkiewicz, Crimes and Harper1970
Figure 9d
Material. Eight specimens and numerous observations in the field.
Description. Medium-sized Glenodictyon with strings 0.9–1.3 mm in diameter and mesh diameters ranging from 2.2–4.7 mm. These smooth, consistently hexagonal networks are preserved in convex hyporelief on the bases of 1.8–3.2 cm thick, medium- to fine-grained sandstones, with evidence of internal grading, as well as internal parallel- and cross-lamination (Tab/Ta,c/Tbc).
Remarks. Ichnospecies of Paleodictyon are assigned on the basis of mesh size and string diameter. Accordingly, the current specimens, with their mesh width of 2.2–4.7 mm and string diameters of 0.9–1.3 mm, can be classified as Paleodictyon (Glenodictyon) intermedium. This is based on the dimension values provided by Książkiewicz (Reference Książkiewicz1977) in his table 16. In the accompanying descriptions, however, Książkiewicz (Reference Książkiewicz1977) notes that P. intermedium generally have regular hexagonal-shaped meshes and that the overall size of the ichnofossil rarely exceeds c. 5 cm². These latter features aid in distinguishing between P. intermedium and P. latum (given that there is a small degree of overlap between the two ichnospecies, as noted in his table 16). Uchman (Reference Uchman1998), in his review of Książkiewicz's material, included P. intermedium under P. strozzii. However, Książkiewicz (Reference Książkiewicz1977) clearly distinguished between the ichnospecies P. strozzii and P. intermedium on the basis of their size, noting that the former is distinctly larger (mesh sizes in P. strozzii range from 2.5–5.5 mm and riblets between 0.3 and 1.0 mm, whereas in P. intermedium they are 2.0–3.5 mm and 0.5–1.0 mm, respectively). However, Książkiewicz (Reference Książkiewicz1977) also noted the degree of irregularity in terms of mesh morphology between P. strozzii and P. intermedium, with the former showing a range of different mesh forms ranging from roughly hexagonal, pentagonal or rhomboidal or even elongated in one direction. Uchman (Reference Uchman1998), however, considered that P. intermedium was a junior synonym of P. strozzii. This, however, is clearly not the case (based on the original descriptions of Książkiewicz, Reference Książkiewicz, Crimes and Harper1970, Reference Książkiewicz1977), and we herein assign the current specimen to P. intermedium.
Paleodictyon (Glenodictyon) majus Savi & Meneghini, Reference Savi and Meneghini1851
Figure 9e, f
Material. Two specimens and several observations in the field.
Description. Six-sided netlike structures preserved in convex hyporelief at the base of 0.6–2.5 mm thick sandstones. Internally, the sandstones show evidence of grading, parallel-lamination and cross-lamination (Tab, Tbc). Strings are 1.5–2.0 mm in diameter and mesh diameters range from 10–15 mm.
Remarks. Paleodictyon majus was originally named in Savi & Meneghini (Reference Savi and Meneghini1851), although neither illustration nor description were provided. However, both De Stefani (Reference De Stefani1879) and Peruzzi (Reference Peruzzi1880) noted that Savi & Meneghini (Reference Savi and Meneghini1851) were the authors of this form, and as Książkiewicz (Reference Książkiewicz1977) writes, it was probably noted in a collection but not described in detail. Indeed, the first written description was by Peruzzi (Reference Peruzzi1880). Paleodictyon majus can be distinguished from P. tauricum on the basis of its thinner riblets.
Paleodictyon (Glenodictyon) maximum Eichwald, Reference Eichwald1868
Figure 9b
Material. Two specimens and several observations in the field.
Description. Smooth, six-sided netlike structures with meshes ranging in size from 0.54–1.18 cm, while the strings range in diameter from 2.6–4.3 mm. The traces are preserved in convex hyporelief on the bases of 2.0–4.3 cm thick, medium- to coarse-grained, structureless or graded, parallel and cross-laminated (Tac) sandstone beds.
Remarks. The dimensional values of the current specimen accord most closely with those of Paleodictyon maximum, which according to Uchman (Reference Uchman1998) comprises Glenodictyon morphologies with riblets of up to 2.8 mm (i.e. smaller than the current specimens) and mesh sizes of up to 1.4 cm (again smaller than the current specimens). The Vera Basin specimens can be distinguished from Paleodictyon gomezi on the basis of its larger mesh size (> 4.0 cm).
Paleodictyon (Glenodictyon) miocenicum Sacco, Reference Sacco1888
Figure 9a
Material. Two specimens and rarely noted in the field.
Description. Six-sided netlike structure preserved in hypichnial relief at the base of 1.2–2.9 cm thick normally graded, medium- to very fine-grained sandstones beds. Internally, parallel- and cross-lamination (Tacd, Tbc) were also noted. Strings are c. 1.0 mm in diameter, with some swelling noted in the middle, while the mesh diameter is 5–7 mm.
Remarks. The samples from the Vera Basin coincide well with the descriptions provided by Książkiewicz (Reference Książkiewicz1977). However, the present samples do not show any evidence of tubercles in the mesh centres (e.g. the crossing points of the burrows within the network), which are noted as characteristic of P. miocenicum by Książkiewicz (Reference Książkiewicz1977). This particular feature, however, was not deemed to be of sufficient importance in the assignation of the ichnospecies.
Paleodictyon (Glenodictyon) minimum Sacco, Reference Sacco1888
Figure 7b–d
Material. Four specimens and several observations in the field.
Description. Smooth, netlike trace fossil preserved in convex hyporelief on the bases of 2.0–2.5 cm thick, medium-grained sandstone beds, with evidence of normal grading and parallel-lamination (Tab). This very small Glenodictyon consists of strings which are 0.4–0.5 mm in diameter while the mesh diameters range from 0.5–0.7 mm. Two specimens are preserved in highly convex and prominent hyporelief, resulting in a protruding plate-like appearance.
Remarks. The samples from the Vera Basin coincide well with the descriptions provided by Uchman (Reference Uchman1995). According to Seilacher (Reference Seilacher, Crimes and Harper1977, Reference Seilacher2007), P. minimum is typically distinguished by the presence of clear burrow systems, which are deeper than those of other Paleodictyon species, leading to the formation of a particularly prominent network. Monaco (Reference Monaco2008) qualified this characterization suggesting that there are two types of P. minimum, namely a shallow and a deep type. Both of these types are recognizable in the samples from the Vera Basin.
Ichnogenus Phycodes Richter, Reference Richter1850
Phycodes bilix Książkiewicz, Reference Książkiewicz1977
Figure 8b
Material. One collected specimen and rarely noted in the field.
Description. Bundled and branching burrows (i.e. flabellate), occasionally falcate on the top of 0.9–1.1 cm thick, medium-grained sandstone beds with internal cross-lamination. The burrows are up to 1 cm in diameter, branching from a central burrow into, up to three, slightly curved burrows separated from the main burrow at angles of c. 50°. Occasionally the edges of the burrows are elevated, leading to the formation of a median depression.
Remarks. Although Han & Pickerill (Reference Han and Pickerill1994) provided a detailed overview of this ichnogenus, Uchman (Reference Uchman1998) has noted the need for ichnogeneric reform, suggesting that Książkiewicz's (Reference Książkiewicz1977) Butholrephis bilix should be included in the Phycodes ichnogenus. Comparing the current specimens with the illustrations provided by both Han & Pickerill (Reference Han and Pickerill1994) and Uchman (Reference Uchman1998) would suggest that the current specimen best conforms to the ichnospecies P. bilix on the basis of the median depression observed in the samples from the Vera Basin. This depression was also described by Książkiewicz (Reference Książkiewicz1977; i.e. Butholrephis bilix) and Uchman (Reference Uchman1998), with partial collapse of the thick-walled burrows being suggested as the cause of the central depression.
Ichnogenus Scolicia de Quatrefages, Reference de Quatrefages1849
Scolicia prisca de Quatrefages, Reference de Quatrefages1849
Figure 10b
Material. Four samples and rarely noted in the field
Description. Tripartite winding trails or burrows, broadly U-shaped in transverse section, preserved in concave epirelief on the surface of 1.5–2.0 cm thick, cross-bedded and medium-grained sandstone beds (Tc). The median ridge, which is 3–5 mm in width, is convex and ribbed (10–11 ribs per centimetre). The ridge is bordered by two concave furrows, which are up to 2 mm deep and have a width of 0.9–1.8 mm. In parts, asymmetrical and looser packed ribs (5–6 ribs per cm) are preserved along the outer slope/margins of the trails/burrows.
Remarks. The samples from the Vera Basin compare well with the Scolicia prisca descriptions provided by Książkiewicz (Reference Książkiewicz1977) and Uchman (Reference Uchman1998). However, the ribs are weakly developed in the current specimens, which may be a result of preservation. Despite this, the samples include the main characteristics of S. prisca (i.e. fine, packed transverse ribs along the median lobe and looser asymmetrical ribs along the slopes/margins, cf. Uchman, Reference Uchman1998).
Scolicia plana Książkiewicz, Reference Książkiewicz, Crimes and Harper1970
Figure 10d
Material. Three samples collected and rarely noted in the field.
Description. Three-lobed trace containing deep marginal grooves. The flat median lobe is 6–9 mm in width, while the grooves range from 2.1–2.4 mm in width. The median lobe is ridged, with 8 ribs/cm present. Ribs are slightly concave to straight and oriented at 90° to the axis of the trail. A second form is also trilobate, albeit with shallow marginal grooves. The flat median lobe is 9–10 mm in width, while the grooves are 3–6 mm in width. The ribs are straight (10/cm) and oriented at 90° to the axis of the trail. Both forms are epichnial on the top of 1.9–2.2 cm thick, medium-grained and normally graded sandstones. Internally, the sandstones show evidence of cross-lamination (Tc).
Remarks. Książkiewicz (Reference Książkiewicz1977) noted that there was a degree of variability in this ichnospecies, suggesting that there were two forms. His ‘Form A’ comprises an epichnial trilobate furrow with a low flat median lobe, densely striated and bordered by less densely ribbed fringes. The median lobe may be bordered by narrow rims and longitudinally transected by a narrow trench (Książkiewicz, Reference Książkiewicz1977). ‘Form B’ is similar, although lateral strings are present (Książkiewicz, Reference Książkiewicz1977). The lack of these lateral strings suggests that the forms present within the Vera Basin conform to Książkiewicz's ‘Form A’. In the illustration of the morphologies which occur within ‘Form A’, Książkiewicz (Reference Książkiewicz1977) notes morphologies with both deep and shallow marginal grooves, both of which occur in the Scolicia plana specimens from the Vera Basin.
Ichnogenus Thalassinoides Rieth, Reference Rieth1932
?Thalassinoides suevicus Rieth, Reference Rieth1932
Figure 8d
Material. One collected specimen and rarely noted in the field.
Description. Horizontal and smooth trace preserved in a convex hyporelief on the base of a 3 cm thick, parallel- and cross-laminated (Tbc) medium- to fine-grained, normally graded sandstone. The Y-shaped burrow is 1.6 mm in diameter and 16 mm in length. It is enlarged (to 2.2 mm) at the point of bifurcation.
Remarks. The sample from the Vera Basin coincides well with the Thalassinoides descriptions of Uchman (Reference Uchman1998) and Carvalho, Viegas & Cachão (Reference Carvalho, Viegas and Cachão2007). It is noticeable, however, that the present specimens are distinctly smaller than those described by Uchman (Reference Uchman1998; i.e. 5–30 mm in diameter), but, Carvalho, Viegas & Cachão (Reference Carvalho, Viegas and Cachão2007) noted that the size of T. suevicus can vary markedly (up to 110 mm in diameter) without providing a minimum size.
Ichnogenus Urohelminthoida Sacco, Reference Sacco1888
Urohelminthoida dertonensis Sacco, Reference Sacco1888
Figure 10a
Material. One collected specimen and numerous observations in the field.
Description. Convex hyporelief on a 2.2 cm thick, medium-grained, normal-graded sandstone with internal parallel- and cross-lamination (Tbc). The trace is preserved as a string-sized (2.3 mm in diameter) meandering form. Lateral appendages are up to 10.7 mm in length and pass straight into one arm of the meander, while an angle (c. 50°) to the second arm is present.
Remarks. The samples of the Vera Basin are consistent with the description of Urohelminthoida dertonensis by Uchman (Reference Uchman1995, Reference Uchman1998) and Książkiewicz (Reference Książkiewicz1977).
4.a. Ichnofossil distribution
In total 12 ichnogenera, comprising 21 ichnospecies were recognized from the sedimentary succession in the southern part of the Vera Basin. The ichnoassemblages clearly belong to the Nereites ichnofacies (Seilacher, Reference Seilacher, Crimes and Harper1977, Reference Seilacher2007), given that they contain many classic ichnogenera (e.g. Desmograpton, Helminthoida, Helminthopsis, Paleodictyon, Scolicia and Urohelminthoida) typical of deep-marine settings (e.g. Książkiewicz, Reference Książkiewicz1977; Seilacher, Reference Seilacher2007; Uchman, Reference Uchman and Miller III2007; MacEachern et al. Reference MacEachern, Bann, Gingras, Zonneveld, Dashtgard, Pemberton, Knaust and Bromley2012). As noted above, the Abad Member extends across the southern and central parts of the basin. The ichnofossil assemblages collected from these two areas differ in terms of both the ichnogenera and ichnospecies present (Tables 1, 2). While the southern area contains mainly Desmograpton dertonensis, D. ichthyforme, Paleodictyon minimum and Urohelminthoida dertonensis, the central area is generally characterized by Paleodictyon maximum and Scolicia plana. However, in the northern part of the central region (Fig. 3c, location of profiles 2.1 and 2.2), all traces of biological activity disappear. This apparent absence of bioturbation occurs both in the complex ichnogenera (e.g. Desmograpton, Paleodictyon, Scolicia or Urohelminthoida), as well as in the relatively common and simple ichnogenera (e.g. Palaeophycus). The precise reasons for this will be discussed below.
Table 1. Relative frequencies of the ichnogenera from the Abad Member of the Vera Basin

Frequencies range from rare (x) through to relatively abundant (xxxxx).
Table 2. Relative frequencies of the ichnospecies from the Abad Member of the Vera Basin

Frequencies range from rare (x) through to relatively abundant (xxxxx).
5. Discussion
The Abad Member crops out in the Vera Basin, extending from the southern basin margins where it interdigitates with the shallow-marine sediments of the Azagador Member through to the central parts of the basin. There is also a corresponding increase in member thickness from the margins through to the basin centre. As noted above, the Abad Member is dominated by marls, with medium- to coarse-grained sandstones concentrated in the southern parts of the basin (i.e. Almocáizar Corridor, which is also the stratigraphically oldest part of the Abad Member, as suggested by field mapping), while fine- to medium-grained sandstones characterize the central parts of the basin.
5.a. Palaeoenvironmental setting
The palaeoenvironmental setting of the Abad Member can be examined from two perspectives: (1) the sediments, and (2) the ichnoassemblages. Both provide important and useful information for the characterization of the depositional setting. As noted above, the sandstone beds of the Abad Member can be uniformly described according to the Bouma (Reference Bouma1962) scheme, and can thus be termed classical turbidites (albeit thinner Ta-c, Tbc turbidites, i.e. Facies C2.2, 2.3 of Pickering et al. Reference Pickering, Stow, Watson and Hiscott1986). The presence of sandstone turbidites records the depositional passage of turbidity currents, which can be found in a variety of areas, including deep-marine settings, ramps or slope aprons but also in lacustrine settings (e.g. Stelting, Bouma & Stone, Reference Stelting, Bouma, Stone, Bouma and Stone2000; Mulder & Alexander, Reference Mulder and Alexander2001; Talling et al. Reference Talling, Masson, Sumner and Malgesini2012; Moernaut et al. Reference Moernaut, van Daele, Heirman, Fontijn, Strasser, Pino, Urrutia and Batist2014; Pickering & Hiscott, Reference Pickering and Hiscott2015). Thus, while the presence of turbidites does not automatically imply a deep-marine setting, the presence of interdigitated muddy to silty structureless marls and associated marine fossils (e.g. microfauna, macrofauna, ichnofauna) would suggest a marine setting for the sediments of the Abad Member. Indeed, the relative thicknesses of the turbiditic sandstone beds would imply a broadly distal setting, although whether this can be classified and integrated into an existing submarine fan model (e.g. Pickering et al. Reference Pickering, Stow, Watson and Hiscott1986; Mulder & Alexander, Reference Mulder and Alexander2001; Talling et al. Reference Talling, Masson, Sumner and Malgesini2012; Pickering & Hiscott, Reference Pickering and Hiscott2015) is questionable.
Based on the ichnofossils present, the sediments of the Abad Member are considered to have been deposited in a setting represented by the classical Nereites ichnofacies (cf. Seilacher, Reference Seilacher1964), with a mixture of facies-crossing and facies-controlled ichnogenera present. Seilacher (Reference Seilacher1964) introduced the concept of ichnofacies, using them to reconstruct specific palaeoenvironmental conditions, with the Nereites ichnofacies representing a deep-marine depositional environment. In detail, this ichnofacies is characterized by the ethologies of the trace-producing organisms and their related trophic styles. These styles are described as complex grazing and patterned feeding or dwelling structures. They occur below the sediment/water interface, and form as a result of organized and efficient feeding strategies (c.f. table 1 of MacEachern et al. Reference MacEachern, Bann, Gingras, Zonneveld, Dashtgard, Pemberton, Knaust and Bromley2012). These features result in the formation of a suite of characteristic traces that represent the ichnofacies (i.e. graphoglyptids: meanders and networks). The majority of the ichnogenera present within the Abad Member (e.g. Desmograpton, Glockerichnus, Helminthoida, Helminthopsis, Paleodictyon, Scolicia and Urohelminthoida) are considered to be characteristic for the Nereites ichnofacies (Seilacher, Reference Seilacher1964, Reference Seilacher2007; Leszczyński & Seilacher, Reference Leszczyński and Seilacher1991; Uchman, Reference Uchman and Miller III2007; MacEachern et al. Reference MacEachern, Bann, Gingras, Zonneveld, Dashtgard, Pemberton, Knaust and Bromley2012; Uchman & Wetzel, Reference Uchman, Wetzel, Knaust and Bromley2012). Additionally, the interpreted palaeoenvironmental conditions as determined by the detailed analysis of the sedimentary facies and facies associations within the Abad Member (i.e. muddy to silty marls with interstratified sandy turbidites) would support, the typical conditions of such environments (i.e. muddy sediments as a result of very slow sedimentation with intercalated sandy turbidites; MacEachern et al. Reference MacEachern, Bann, Gingras, Zonneveld, Dashtgard, Pemberton, Knaust and Bromley2012). Thus, a possible depositional setting within a lower slope or basin floor environment (and related fan/lobe system) would successfully integrate both the sedimentary and ichnological observations (cf. MacEachern et al. Reference MacEachern, Bann, Gingras, Zonneveld, Dashtgard, Pemberton, Knaust and Bromley2012).
The Nereites ichnofacies has been revised by a number of workers (e.g. Seilacher, Reference Seilacher, Crimes and Harper1977; Uchman, Reference Uchman2001, Reference Uchman and Miller III2007; Heard & Pickering, Reference Heard and Pickering2008; Knaust, Reference Knaust2009; Uchman & Wetzel, Reference Uchman, Wetzel, Knaust and Bromley2012) who subdivided it into three sub-ichnofacies (Nereites, Paleodictyon and Ophiomorpha rudis) based primarily on the lithological characteristics within the individual environmental settings. This subdivision allows more subtle ichnoassemblages to be recognized and classified, and in particular it better reflects the bathymetric differentiation of a typical deep-sea fan system (i.e. inner to outer fan) as well as lateral changes within such a system (i.e. interchannel areas, channel axis, levee and overbank deposits; cf. Seilacher, Reference Seilacher, Crimes and Harper1977; Uchman, Reference Uchman2001, Reference Uchman and Miller III2007; Heard & Pickering, Reference Heard and Pickering2008; Knaust, Reference Knaust2009; Uchman & Wetzel, Reference Uchman, Wetzel, Knaust and Bromley2012).
The ichnoassemblages present within the Abad Member, together with the sedimentary successions, suggest a depositional setting corresponding to the Paleodictyon sub-ichnofacies (i.e. medium- to thin-bedded sandstone turbidites in a mud-rich succession; cf. Uchman & Wetzel, Reference Uchman, Wetzel, Knaust and Bromley2012), and suggest an intermediate setting in terms of both bathymetric and lateral differentiation of the Nereites ichnofacies. Such a setting could include overbank, interchannel, interlobe or distal depositional-lobe environments (Wetzel & Uchman, Reference Wetzel and Uchman1997; Uchman, Reference Uchman and Miller III2007; MacEachern et al. Reference MacEachern, Bann, Gingras, Zonneveld, Dashtgard, Pemberton, Knaust and Bromley2012; Uchman & Wetzel, Reference Uchman, Wetzel, Knaust and Bromley2012).
However, the absence of obvious channel sediments (both, main or distributary channel) in the outcrop area, suggests a possible depositional-lobe-type setting. The palaeogeographic setting of the Almocáizar Corridor, and its extension into the Vera Basin, is that of a mixed siliciclastic–carbonate shelf extending into a deeper-marine environment (Fig. 11). The shelf is oriented E–W and has been classified as relatively narrow, with the shelf-slope break located close to the shoreline (cf. Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Martín et al. Reference Martín, Braga, Sánchez-Almazo, Aguirre, Mutti, Piller and Betzler2012). In this respect, it was probably similar to other published examples of narrow shelf areas (e.g. the Oligocene of New Zealand, Carter & Lindqvist, Reference Carter and Lindqvist1975; present-day Albany Shelf, Australia, James & Bone, Reference James and Bone2010). Along the northern margin of the shelf (i.e. Sierra de los Filabres) coastal (including beach systems) and inner-platform sediments were deposited, while small deltas also formed along the coast (e.g. in the area of Almocáizar; Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Martín et al. Reference Martín, Braga, Sánchez-Almazo, Aguirre, Mutti, Piller and Betzler2012). In areas where delta development is marked, the shelf is more clastic (with sediment also being transported parallel to the shoreline by ENE-directed longshore currents), whereas away from the areas of delta formation, the shelf sediments are mixed (i.e. carbonate/siliciclastic).
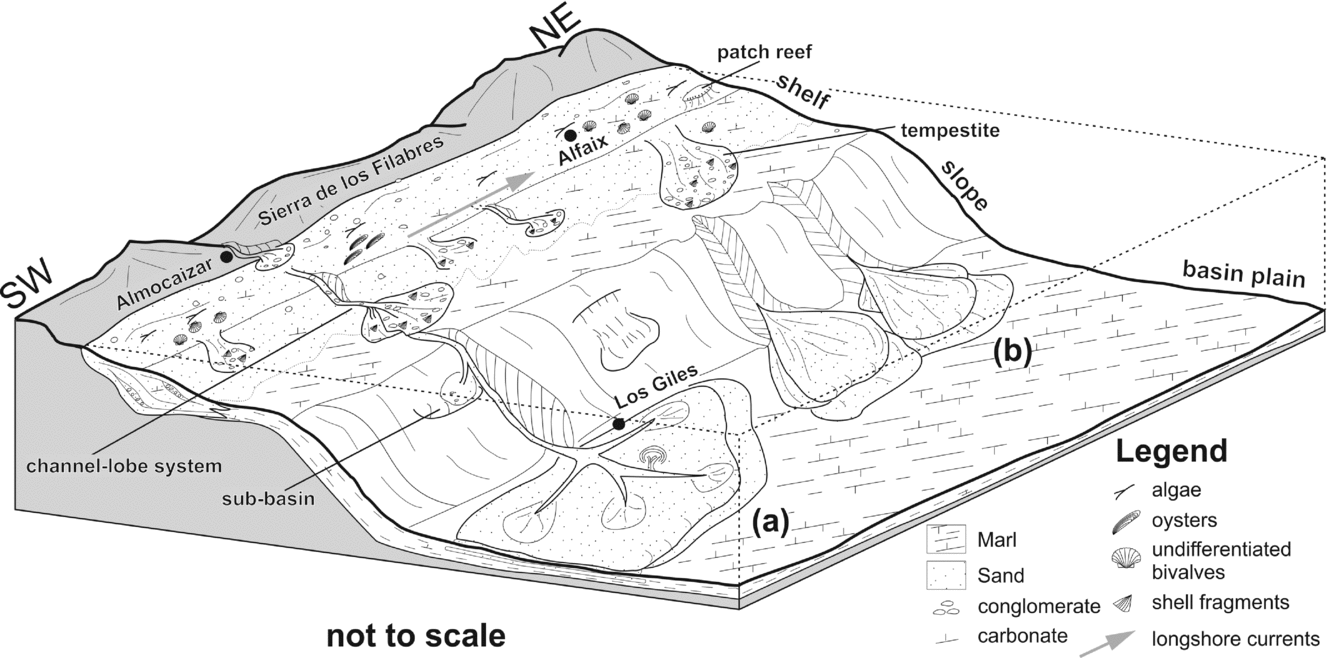
Figure 11. Block diagram reconstructing the depositional setting of the Almocáizar Corridor, Vera Basin (SE Spain) in early Messinian times. This schematic model illustrates the palaeogeography at this time, with the Sierra de los Filabres located in the north, and the shelf/slope/deep-marine areas located further south. Two possible scenarios are envisaged: (a) where a channel-fan system developed downcurrent of a shelf/slope channel system, and (b) a detached lobe system developed basinwards of the narrow shelf with sediment supplied from turbidity currents generated by storms on the shelf. Additionally, a third model would combine aspects of these two scenarios (see text for details). (Shelf morphology and facies modified after Martín et al. Reference Martín, Braga, Sánchez-Almazo, Aguirre, Mutti, Piller and Betzler2012).
The coastal systems prograded southwards into shallow shelf deposits, comprising a mixed siliciclastic–carbonate succession, which has been interpreted in terms of a ramp (with distal steepening; Martín et al. Reference Martín, Braga, Sánchez-Almazo, Aguirre, Mutti, Piller and Betzler2012) or platform (Braga, Martín & Wood, Reference Braga, Martín and Wood2001) system. Subaqueous sand-filled channels (Fig. 11) cross-cutting the inner-platform sediments, and located at the mouths of fluvial systems, have also been noted (Braga, Martín & Wood, Reference Braga, Martín and Wood2001; Martín et al. Reference Martín, Braga, Sánchez-Almazo, Aguirre, Mutti, Piller and Betzler2012). Areas adjacent to these channels were presumably subjected to bypass, with sediment being transported across the narrow shelf and deposited along the shelf-slope break, or indeed, being directly transported into deeper-marine areas (i.e. the area forming the subject of this study). Braga, Martín & Wood (Reference Braga, Martín and Wood2001) suggested that lobe deposits in the deeper-marine area are fed by sediment originating in the Sierra de los Filabres (based on clast analysis) and transported across the shelf.
However, the preserved submarine channel/lobe systems described above (i.e. from Braga, Martín & Wood, Reference Braga, Martín and Wood2001) do not appear to be correlatable with the turbidites described as part of this study. Certainly, there are problems both with regard to (a) the precise location (i.e. Braga, Martín & Wood, Reference Braga, Martín and Wood2001 describe lobes from the area to the south of Turre, see Fig. 3 for location, whereas the present lobes occur in the area of the Almocáizar Corridor), and (b) the stratigraphic position (i.e. the lobes described by Braga, Martín & Wood, Reference Braga, Martín and Wood2001 would appear to be younger in age and to have been deposited in a shallower marine setting). In addition, these lobes appear to be smaller in scale and they comprise mainly coarse-grained sediments (sandstones, conglomerates). The lobes in the present study are composed of finer-grained sediments (sandstones). Additionally, there would appear to be a clear correlation between the channels, extending from the fluvial systems north of the Almocáizar Corridor and crossing the narrow shelf, and the deep-marine lobes to the south. This would suggest that the lobes of the Almocáizar Corridor region were directly fed from erosion of the Sierra de los Filabres, with sediment being transported across the shelf and deposited seawards of the shelf-slope break (Fig. 11).
Two hypotheses are proposed for the precise depositional setting of the Almocáizar lobes. In the first situation (Fig. 11a), sediments were transported down the slope via a submarine channel leading to the establishment of a small mud-rich channel-fan system (cf. Richards, Bowman & Reading, Reference Richards, Bowman and Reading1998). In such a system, a number of features would be anticipated including, for example, clear proximal–distal changes, the presence of a range of turbiditic facies, as well as clear evidence of channelization within the channel-fan system. However, as noted above, the sediments in the Almocáizar Corridor area do not contain evidence of any of these features. The fact that a pronounced channel system was developed on the shelf would certainly suggest that at the shelf-slope break the system would extend basinwards into the deep-marine area. The absence of any evidence of a channel-fan system in the deep-marine parts of the basin does not imply that it never existed, but rather that it was, not preserved or that the current outcrop situation is not extensive enough to allow full recognition of the entire system.
A second option (Fig. 11b) is that the shelf channel system played a minor role, in that the sediments were transported from the coastal areas and deposited in the outer shelf area. In such a scenario, storms on the narrow shelf would have resulted in the generation of slope aprons, and associated turbidity flows, transporting the sediment into the deep-marine area and resulting in the formation of lobe deposits. These lobes, detached from a direct source, would have been formed from individual turbidity currents generated in the shelf/slope region. The fact that the lobes in the present study are generally of similar grain size would suggest pre-sorting in the source area of the transported sediment, an aspect which might correlate with outer shelf deposition. Indeed, the presence of tempestites in the shelf area would support the idea of storm-generated sediment bodies on the shelf.
An alternative option (in addition to the two described above) is that the lobes were produced by a combination of both models, i.e. possible channelization on the slope but with the development of detached lobes, rather than a channel-fan system (i.e. Fig. 11a), coupled with detached lobes developing from slope aprons generated by storm deposits on the shelf (i.e. Fig. 11b). In such a system, the emphasis would be on the lobe morphology and composition (i.e. lobate forms extending basinwards of the slope; finer-grained sediments) rather than the direct presence of feeder systems. The current outcrop situation, however, does not allow any one model to be favoured.
5.b. Bathymetry of the Abad Member
As noted above, the succession is dominated by turbiditic sandstones and marls, although the presence of the former says nothing definitively per se about absolute depths in the depositional area. Seilacher's (Reference Seilacher1964) ichnofacies, while useful for generalized water depth determinations in the fossil record, are, however, not always directly applicable and, indeed, may lead to a false interpretation (e.g. Fürsich, Taheri & Wilmsen, Reference Fürsich, Taheri and Wilmsen2007). MacEachern et al. (Reference MacEachern, Bann, Gingras, Zonneveld, Dashtgard, Pemberton, Knaust and Bromley2012) have suggested that neo-ichnology plays an important role in determining precise depth ranges (or maxima) for organisms in particular settings. For example, Helminthoida-like traces have been recorded at depths of 5248 m from the Japan Trench (Fujioka et al. Reference Fujioka, Taira, Kobayashi, Nakamura, Ilyama, Cadet, Lallemand and Girard1987), while Scolicia-like trails have been noted from 3830 m from the Taira deep-sea fan and from 3835 m from the Tenryu deep-sea fan (Fujioka et al. Reference Fujioka, Taira, Kobayashi, Nakamura, Ilyama, Cadet, Lallemand and Girard1987). The possible water depths at which these organisms can exist, therefore, are deep. While similar water depths for the Abad Member are not probable (based on palaeogeographical evidence, see below), it does suggest that similar conditions (e.g. very low energy and related slow rates of deposition) can be presumed. When extrapolating to the fossil record, however, the problematics of depth determination become more obvious. While estimated bathymetries are, at best, imprecise, some ichnofossils (particularly those considered to be facies-controlled, e.g. Desmograpton) can be used to provide rough depth estimates of the depositional setting.
As noted above, ichnofacies models have been determined on the basis of trace-fossil morphology, and this is controlled by the feeding behaviour of the trace-producing organisms (Książkiewicz, Reference Książkiewicz1977). Thus, the pattern of the traces and their complexity is dependent on the amount of food available within the sediment. Consequently, with increasing depth a decrease in nutrient availability can be generally assumed (e.g. Thibodeau et al. Reference Thibodeau, Lehmann, Kowarzyk, Mucci, Gélinas, Gilbert, Maranger and Alkhatib2010; Moore et al. Reference Moore, Mills, Arrigo, Berman-Frank, Bopp, Boyd, Galbraith, Geider, Guieu, Jaccard, Jickells, La Roche, Lenton, Mahowald, Maranon, Marinov, Moore, Nakatsuka, Oschlies, Saito, Thingstad, Tsuda and Ulloa2013), resulting in the development of more complex trace patterns (e.g. graphoglyptids or networks; Książkiewicz, Reference Książkiewicz1977; Seilacher, Reference Seilacher, Crimes and Harper1977, Reference Seilacher2007; Uchman & Wetzel, Reference Uchman, Wetzel, Knaust and Bromley2012). Furthermore, the type of sediment (e.g. turbidites or deep-sea muds) and the composition of the water (e.g. oxygenation levels) may influence both the form and amount of nutrients present within the sediments of the deep sea. These variations may be independent of depth, thus suggesting that palaeobathymetric reconstructions need to take a number of factors into account and not just the morphology of the particular ichnofossils. Additionally, with the Nereites ichnofacies broadly considered to represent ‘deep-marine’ settings, there is an increasing body of work which suggests otherwise (e.g. Häntzschel, Reference Häntzschel and Teichert1975; Ernst & Zander, Reference Ernst, Zander, Abbate, Sagri and Sassi1993; Fürsich, Taheri & Wilmsen, Reference Fürsich, Taheri and Wilmsen2007). Paleodictyon, for example, has been reported from shallow-marine locations in the Lower Palaeozoic (Stanley & Pickerill, Reference Stanley and Pickerill1993), while Gierlowski-Kordesch & Ernst (Reference Gierlowski-Kordesch, Ernst, Matheis and Schandelmeier1987) and Ernst & Zander (Reference Ernst, Zander, Abbate, Sagri and Sassi1993) have reported it in Upper Cretaceous-age midshelf deposits where it was associated with other ichnofossils considered typical of the Nereites ichnofacies (e.g. Cosmorhaphe, Spirorhaphe, Urohelminthoida). Other evidence for comparatively shallow-marine Paleodictyon has been provided by Hantzpergue & Branger (Reference Hantzpergue and Branger1992), who reported the ichnofossil from an upper Oxfordian limestone unit which they suggested was deposited at a depth of not > 100 m. Depths of 200–300 m have been suggested by Uchman, Janbu & Nemec (Reference Uchman, Janbu and Nemec2004) from the Cretaceous–Eocene-age central Pontides of Turkey. Considered together, these data question the assumption that the Nereites ichnofacies can always be considered as ‘deep marine’. Indeed Fürsich, Taheri & Wilmsen (Reference Fürsich, Taheri and Wilmsen2007) have gone further, suggesting that the bathymetric range of Paleodictyon was wider than generally assumed, and that its dominance in deep-marine turbiditic successions is at least partly a preservational effect. Furthermore, Fürsich, Taheri & Wilmsen (Reference Fürsich, Taheri and Wilmsen2007) suggested that bathymetric interpretations are best made on the basis of complete ichnoassemblages rather than single ichnogenera/ichnospecies.
This approach, i.e. using complete ichnoassemblages, but also combining them with other factors, was used by Książkiewicz (Reference Książkiewicz1977) in order to provide a broad bathymetric range for a series of particular ichnospecies found within Cenozoic-age sediments (and which are comparable with the samples from this study). According to these results, the palaeobathymetry of the Abad Member would fall within the epibathyal zone (200–600 m of Książkiewicz, Reference Książkiewicz1977; cf. his table 11) and extending down to the onset of the mesobathyal zone (600–1000 m of Książkiewicz, Reference Książkiewicz1977; cf. his table 11; p. 40). Interestingly, the water depth estimates suggested by the foraminiferal assemblage present within the Abad Member correlate well with those of the ichnofossils. Baggley (Reference Baggley2000) analysed foraminiferal assemblages (predominantly benthic) from several sections within the Sorbas Basin and the Almocáizar Corridor (connecting the Sorbas and Vera basins, and overlapping with the sampling location (I) of this study, Fig. 3a). Nine samples were collected from an 84.5 m thick section (i.e. the ‘Barranco de Los Giles (BG)’ section, which is located within the older sediments of this study (i.e. within the Almocáizar Corridor) and c. 500–600 m to the south of our sampling area; Baggley, Reference Baggley2000). The foraminiferal assemblage includes Eponides pusillus, Pullenia bulloides and Quinqueloculina venusta suggesting a lower epibathyal to upper mesobathyal depth (500–1300 m; Baggley, Reference Baggley2000), and thus broadly encompassing the 200–1000 m as suggested by the ichnoassemblage.
5.c. Variations between the Vera Basin and the Sorbas/Nijar basins
The Turre Formation (i.e. Azagador and Abad members, Figs 3, 4) crops out in the Vera Basin, as well as in the adjacent Sorbas and Nijar basins (Fig. 4). In all three basins, the Azagador Member is similar in terms of its lithological composition and appearance. However, the overlying Abad Member differs markedly between the Vera Basin, where it contains turbiditic sandstones, and the Sorbas and Nijar basins where it does not. Additionally, foraminiferal studies reflect a clear lateral deepening of the Abad Member towards the Vera Basin (Baggley, Reference Baggley2000). Thus, the depositional history of the Abad Member in the Vera Basin marks a caesura in terms of the regional sedimentary evolution. The question, as to why deepening occurred within the Vera Basin, is problematic. In contrast, the deposits of the basins to the south represent a clear shallowing trend.
The tectonic evolution of the area in Messinian times resulted in basin separation during the period of the deposition of the upper Abad Member leading to the tectonic restriction and isolation of the Sorbas and Nijar basins (Fig. 2; Braga et al. Reference Braga, Martín, Riding, Aguirre, Sánchez-Almazo and Dinarès-Turell2006). Coevally, extensive carbonate platforms and reefs (e.g. Cantera Member) along the basin margin topographic highs in the Sorbas and Nijar basins developed (Fig. 2b). However, this development did not occur in the Vera Basin, except for some rare and minor occurrences along the central-western margin of the basin (Fig. 3, cropping out to the west of Antas). The reefal growth, especially in the Sorbas Basin (Fig. 2b), would also have resulted in the formation of a topographic barrier to the free movement of water bodies between the Vera Basin to the north (with open contact to the Mediterranean Sea) and the Sorbas and Nijar basins to the south-west (Fortuin & Krijgsman, Reference Fortuin and Krijgsman2003; Krijgsman et al. Reference Krijgsman, Leewis, Garcés, Kouwenhoven, Kuiper and Sierro2006). These barriers would thus have reinforced the basin separation between the two regions, which from this point in time underwent differing depositional evolutions (cf. see above/regional geology; Fig. 2).
5.d. The lower Abad Member (rapid rise in sea level)
As noted above, there are clear changes in terms of the ichnoassemblage between the older part of the Abad Member (i.e. Almocáizar Corridor) and the younger sediments in the central part of the Vera Basin. This change is characterized by a marked decrease in the amount of recognizable ichnospecies and ichnogenera (Tables 1, 2). In addition, typical ichnofossils of the mesobathyal zone (i.e. Desmograpton ichthyforme; Helminthoida labyrintica) are no longer present. The changes in terms of ichnoassemblage between the two locations can be interpreted in terms of a temporal change in bathymetry from a deeper (upper meso- to lower epibathyal: i.e. older deposits) setting in the basin margin/older deposits through to a shallower setting (mainly epibathyal; i.e. younger deposits) in the basin centre. Such a shallowing trend was also noted by Baggley (Reference Baggley2000) in his study of the benthic foraminifera, where a deep-to-shallow trend could also be seen in both the Sorbas and Vera basins. In addition, sequence-stratigraphic models for the Vera Basin (Braga, Martin & Wood Reference Braga, Martín and Wood2001), as well as for the Sorbas Basin (Martin et al. Reference Martín, Braga, Betzler and Brachert1996), support the recognized deep-shallow trend obviously. These models were established on the basis of the Azagador Member deposits immediately underlying (and sometimes interdigitating with) the Abad Member, which comprise shallow-marine sandstones and temperate carbonates. Such sediments are generally deposited in shelf regions at depths of < 200 m (e.g. James & Bone, Reference James and Bone2010). The presumed depth of the upper part of the Abad Member, therefore, would involve a sudden and geologically rapid increase in water depth to a maximum depth of c. 1100 m. The question is how such a rapid increase in water depth could be explained.
The Azagador Member was deposited along the basin margin, while the Abad Member extended towards the basin centre (Figs 3, 4). However, it cannot be stated with any certainty that a specific stratigraphic level within either member corresponds to a specific stratigraphic time since it is entirely probable that the finer-grained sediments in the basin centre were deposited relatively slowly, in comparison with the coarser basin margin units. Thus, while the stratigraphic interval represented by the joint succession (i.e. basin margin and basin centre) ranges in age from upper Tortonian to uppermost Messinian, the precise ages of the respective successions (i.e. Azagador Member, Abad Member; Fig. 4) and their relationship to one another is unclear.
The rapid change in sea level (i.e. water depth reconstructions between the Azagador and the Abad members) may have been related to tectonic activity in the region. An increase in subsidence of the Vera Basin during the time of deposition of the Abad Member would have coincided with the onset of uplift in the eastern most part of the Sierra de Filabres, which began in late Tortonian times (Fig. 2; Braga, Martín & Quesada, Reference Braga, Martín and Quesada2003). This would have been coeval with the initial phase of deposition of the Azagador Member in the southwest. By mid-Messinian times, uplift of the crystalline massifs surrounding the basin had progressed to the point that the Sierra Cabrera was uplifting (Braga, Martín & Quesada, Reference Braga, Martín and Quesada2003; Booth-Rea et al. Reference Booth-Rea, Azañón, Azor and Garcı́a-Dueñas2004). Additionally, there is a clear correlation between the initial activity of the Palomares Fault Zone (in late Tortonian times) and uplift activity of the Sierra Cabrera in the Vera Basin (c.f. Booth-Rea et al. Reference Booth-Rea, Azañón, Azor and Garcı́a-Dueñas2004).
5.e. The uppermost Abad Member (end of full marine conditions)
As noted above, there is an obvious change in terms of the ichnofossil assemblages present within the succession between two of the measured profiles (i.e. sections 2.1 and 2.2; Fig. 6) in the central part of the basin. The former area contains an assemblage corresponding to the Nereites ichnofacies, while the latter is characterized by the complete absence of ichnofossils. The latter section (i.e. 2.2) is stratigraphically younger, and comprises finer-grained sediments, including both marls and turbiditic sandstones. A possible explanation for these observed changes may be owing to variations in the palaeoenvironmental conditions, related to temperature, nutrient availability, oxygen level and/or salinity (cf. Wetzel & Uchman, Reference Wetzel and Uchman1998). All of these changes would suggest a transition in terms of the environmental conditions from one favouring the presence of trace-producing organisms to one that could be considered a non-viable zone. In the case of the Vera Basin, which was affected by the Messinian Salinity Crises in latest Messinian times (i.e. uppermost Abad Member), a change in the degree of salinity is the most probable cause. A number of authors have recorded similar facies (i.e. marls and turbidites including Paleodictyon and marls with interbedded very thin turbidites) in the eastern Vera Basin (e.g. Cuevas de Almanzora Section; Montenat et al. Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976; Gonzalez Donoso & Serrano, Reference Gonzalez Donoso and Serrano1978; Cita, Schilling & Bossio, Reference Cita, Schilling and Bossio1980; Benson & Bied, Reference Benson and Bied1991; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995). Within the Cuevas del Almanzora profile, for example, Fortuin, Kelling & Roep (Reference Fortuin, Kelling and Roep1995) recognized the occurrence of ostracods (e.g. Cyprideis sp.) and the algae Chara, both indicative of brackish conditions. These fossils are found close to the boundary between the Messinian and the Pliocene, and would appear to correlate with the change in facies as noted in the southern part of the Vera Basin (i.e. from sediments containing a Nereites ichnofacies assemblage to sediments which are barren in terms of ichnofossils). Thus, the observed radical change in the ichnofossil assemblages may be related to a shift from full marine to brackish conditions, and thus reflect the changes wrought on the basin by the onset of the Messinian event.
A variety of foraminiferal studies have been carried out for stratigraphical purposes in the area (e.g. Montenat et al. Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976; Gonzalez Donoso & Serrano, Reference Gonzalez Donoso and Serrano1978; Cita, Schilling & Bossio, Reference Cita, Schilling and Bossio1980; Benson & Bied, Reference Benson and Bied1991; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995; Sierro et al. Reference Sierro, Hilgen, Krijgsman and Flores2001). These biostratigraphic analyses were mainly based on changes (quantitative and qualitative) in planktonic foraminifera. The aim of these studies was to provide a detailed stratigraphic subdivision of the Tortonian- to Pliocene-aged deposits within the Vera Basin (and the adjacent Sorbas Basin, e.g. Sierro et al. Reference Sierro, Hilgen, Krijgsman and Flores2001), which are dominated by similar, and almost indistinguishable, marl deposits (cf. in the Vera Basin the marls of the Tortonian-aged Chozas Formation, Tortonian- to Messinian-aged Abad Member and the Pliocene-aged Cuevas Formation). Based on these studies, the Tortonian was subdivided into two biostratigraphic units: Tortonian I (of Montenat et al. Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976) characterized by Neogloboquadrina acostaensis and the stratigraphically younger Tortonian II characterized by Globorotalia (G.) miocenica. The onset of the Messinian was defined by the occurrence of G. mediterranea and G. conomiozea. The onset of the youngest marine period in the regional basins (i.e. Pliocene) is characterized by the presence of G. margaritae and G. puncticulata (c.f. Montenat et al. Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976; Benson & Bied, Reference Benson and Bied1991; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995).
The biostratigraphic subdivisions were supplemented and supported by studies on magnetostratigraphy and geochemistry (e.g. Montenat et al. Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976; Benson & Bied, Reference Benson and Bied1991; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995; Sierro et al. Reference Sierro, Hilgen, Krijgsman and Flores2001). The extensive analyses carried out at the Cuevas de Almanzora Section (located in the northeastern part of the Vera Basin), which is a representative section for the Messinian to Pliocene transition, were integrated into high-resolution stratigraphical studies of the region. The description of Montenat et al. (Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976), Benson & Bied (Reference Benson and Bied1991) and Fortuin, Kelling & Roep (Reference Fortuin, Kelling and Roep1995) of the Cuevas de Almanzora Section sediments (grain size, internal structures and bed thickness) as well as their general descriptions of ichnofaunal observations (e.g. presence of Paleodictyon isp. compared to absence of ichnofossils) matches the observations within the deposits of the younger sediments of this study (e.g. sections 2.1 and 2.2; Fig. 6).
In addition to the stratigraphical work, a number of studies focused on palaeoenvironmental reconstructions. Examination of the foraminiferal and ostracod assemblages showed clear evidence of changes in salinity (e.g. Montenat et al. Reference Montenat, Bizon, Bizon, Carbonnel, Muller and Reneville1976; Benson & Bied, Reference Benson and Bied1991; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995; Sierro et al. Reference Sierro, Hilgen, Krijgsman and Flores2001; Rouchy & Caruso, Reference Rouchy and Caruso2006; Bourillot et al. Reference Bourillot, Vennin, Rouchy, Blanc-Valleron, Caruso and Durlet2010). Indeed, three different phases of the MSC (5.971–5.33 Ma, cf. Roveri et al. Reference Roveri, Lugli, Manzi, Gennari and Schreiber2014) could be identified across the entire Mediterranean area. These three phases were interpreted as representing: (1) the onset of the MSC, (2) the isolation of the Mediterranean area from the Atlantic Ocean, and (3) the reflooding of the Mediterranean Sea (Rouchy & Martin, Reference Rouchy and Martin1992; Krijgsman et al. Reference Krijgsman, Hilgen, Raffi, Sierro and Wilson1999; Bourillot et al. Reference Bourillot, Vennin, Rouchy, Blanc-Valleron, Caruso and Durlet2010; Manzi et al. Reference Manzi, Gennari, Hilgen, Krijgsman, Lugli, Roveri and Sierro2013; Do Couto et al. Reference Do Couto, Popescu, Suc, Melinte-Dobrinescu, Barhoun, Gorini, Jolivet, Poort, Jouannic and Auxietre2014; Roveri et al. Reference Roveri, Lugli, Manzi, Gennari and Schreiber2014; Flecker et al. Reference Flecker, Krijgsman, Capella, Castro Martíns, de, Dmitrieva, Mayser, Marzocchi, Modestou, Ochoa, Simon, Tulbure, van den Berg, van der Schee, Lange, de, Ellam, Govers, Gutjahr, Hilgen, Kouwenhoven, Lofi, Meijer, Sierro, Bachiri, Barhoun, Alami, Chacon, Flores, Gregory, Howard, Lunt, Ochoa, Pancost, Vincent and Yousfi2015). The second of these phases encompasses the Lago Mare Facies (5.55–5.32 Ma, Stoica et al. Reference Stoica, Krijgsman, Fortuin and Gliozzi2016), interpreted as representing predominantly brackish conditions, and which is widely observed in the Mediterranean region (e.g. Deckker, Chivas & Shelley, Reference Deckker, Chivas and Shelley1988; Rouchy et al. Reference Rouchy, Orszag-Sperber, Blanc-Valleron, Pierre, Rivière, Combourieu-Nebout and Panayides2001; Warny, Bart & Suc, Reference Warny, Bart and Suc2003; Clauzon et al. Reference Clauzon, Suc, Popescu, Marunteanu, Rubino, Marinescu and Melinte2005; Orszag-Sperber, Reference Orszag-Sperber2006; Do Couto et al. Reference Do Couto, Popescu, Suc, Melinte-Dobrinescu, Barhoun, Gorini, Jolivet, Poort, Jouannic and Auxietre2014; Roveri et al. Reference Roveri, Lugli, Manzi, Gennari and Schreiber2014; Marzocchi et al. Reference Marzocchi, Flecker, van Baak, Lunt and Krijgsman2016; Stoica et al. Reference Stoica, Krijgsman, Fortuin and Gliozzi2016). This particular phase occurred during the time of the continued separation of the Mediterranean Sea from the Atlantic. Thus, waters entering the Vera Basin (and related basins) were derived from freshwater influx from the surrounding region. The marine influence at this time was confined to occasional influxes from the Paratethys, as evidenced from a diverse assemblage of Paratethyan ostracod species (e.g. Cyprideis and Loxoconcha; Benson & Bied, Reference Benson and Bied1991; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995; Do Couto et al. Reference Do Couto, Popescu, Suc, Melinte-Dobrinescu, Barhoun, Gorini, Jolivet, Poort, Jouannic and Auxietre2014; Stoica et al. Reference Stoica, Krijgsman, Fortuin and Gliozzi2016). The combination of the ichnofossil (i.e. absence of bioturbation) and microfossil (ostracods and foraminifera) records from the Abad Member sediments of the central part of the Vera Basin thus suggest that brackish conditions were active at this particular time. This would suggest a possible correlation with the extant Lago Mare Facies elsewhere in the Mediterranean region (e.g. Benson & Bied, Reference Benson and Bied1991; Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995; Do Couto et al. Reference Do Couto, Popescu, Suc, Melinte-Dobrinescu, Barhoun, Gorini, Jolivet, Poort, Jouannic and Auxietre2014; Stoica et al. Reference Stoica, Krijgsman, Fortuin and Gliozzi2016), and that the Abad Member succession in the Vera Basin represents a shift from full marine through to early post-MSC deposition. Thus, the correlation of the sediments of section 2.2 with the Lago Mare Facies would imply that these particular deposits should not be assigned to the Abad Member. A clear stratigraphical assignment to the period of the Messinian Salinity Crises and the corresponding Lago Mare Facies (Fortuin, Kelling & Roep, Reference Fortuin, Kelling and Roep1995; Do Couto et al. Reference Do Couto, Popescu, Suc, Melinte-Dobrinescu, Barhoun, Gorini, Jolivet, Poort, Jouannic and Auxietre2014; Stoica et al. Reference Stoica, Krijgsman, Fortuin and Gliozzi2016) is necessary.
6. Conclusions
The succession of the Tortonian- to Messinian-aged Abad Member of the Vera Basin is characterized by distal, low-energy background sedimentation (i.e. marl) with intercalated mixed (i.e. siliciclastic–carbonate) medium- to fine-grained turbidites, which may have been deposited in a deep-marine depositional-lobe-type setting of a channel-fan system and/or detached lobes developing from slope aprons generated by storm deposits on the shelf. The ichnofaunal assemblage of the Abad Member can be assigned to the Nereites ichnofacies of Seilacher (Reference Seilacher1964) and, in detail, to the Paleodictyon sub-ichnofacies of Seilacher (Reference Seilacher2007) and Uchman (Reference Uchman and Miller III2007). The range of ichnofossils (12 ichnogenera, 21 ichnospecies) recorded in this study includes facies-crossing and facies-controlled taxa, with the latter commonly considered as typical of deep-marine depositional settings. Bathymetric analysis of the ichnoassemblages suggests that deposition occurred at epi- to mesobathyal depths, with a deep-to-shallow trend from the older parts of the Abad Member (i.e. Almocáizar Corridor) through to the younger parts of the member (i.e. basin centre). The youngest sediments are considered to be Messinian in age and these record the abrupt disappearance of the ichnofossils. This disappearance heralded the onset of the MSC in the region and the onset of sedimentation under restricted conditions (i.e. brackish/Lago Mare phase of the Mediterranean region).
Acknowledgements
The authors would like to thank the reviewers, Franz Fürsich and an anonymous reviewer for the excellent comments which greatly helped to improve the manuscript. We would also like to thank Eduouad Grigowski and Bettina Krumbiegel for preparing the ichnofossil and graphic diagrams. Additionally, the comments made by Geological Magazine Editor Katarzyna Piper resulted in a much improved and consistent manuscript.













