


Introduction: The History of The Find
The British Museum (Natural History), London, received the partial remains of ‘Tilbury Man’ in 1884, following its discovery in September 1883 during the construction of new docking facilities for the East and West India Dock Company (Lankester Reference Lankester1904, 235) (Figs 1 & 2). The skeleton was presented to the museum by the Company directors, through Donald S. Baynes, the project engineer. A section provided by August Manning and Baynes, published by Richard Owen (Reference Owen1884), shows its position at a depth of some 34½ ft, or 10.5 m, in the uppermost 0.5 m of a (possibly redeposited) Devensian sand layer, underlying an alternating sequence of peats – of which there are three distinct bands – and estuarine silts and clays (Table 1; Figs 3 & 4) (Devoy Reference Devoy1980). Observations of the in situ skeleton were also made during a site visit by Whitaker and Spurrell (Reid Reference Reid1913). Given the circumstances of its discovery, no information is available on the position in which the body lay, nor were any associated finds reportedFootnote 1.

Fig. 1 The Lower Thames region, with selected site locations mentioned in the text. The −9 m OD bathymetric line is near the probable Late Mesolithic coastline at c. 6000 bc

Fig. 2 ‘Tilbury Dock, Dry Dock Caisson’, Tilbury Dock under construction c. 1885 (Hasler Collection, No. 59, Photo R. Hider)
Table 1 TILBURY DOCKS SECTION AT THE LOCATION OF THE SKELETON, FROM DATA PROVIDED BY MANNING & BAYNES (IN OWEN Reference Owen1884)


Fig. 3 ‘Illustration of Peat Stratum’ taken in 1855 with man standing beside railtracks against cutting with c. 1 m thickness peat, with clays above and below (Hasler Collection, History/Tilbury Dock/H.110)

Fig. 4 Tilbury Docks section. The skeleton was found in the upper sands of layer 8 (from section drawn by Manning & Baynes, reproduced in Owen (Reference Owen1884), redrawn by Alison Wilkins)
As was common for many deeply buried human skeletons found in the 19th century, Tilbury Man was originally considered by Sir Richard Owen (Reference Owen1884, 14–5) to be of Palaeolithic age, contemporary with the bones of extinct mammals such as mammoth that were being found in the Thames gravels. This was immediately and strongly criticised by T. Vincent Holmes, president of the Essex Field Club: ‘It is evident, however, that the illustrious and venerable naturalist [Owen] has never visited the Docks himself. Consequently he does not appreciate the difference of age between the older alluvial deposits of the Thames Valley … and the newer alluvium of Tilbury …’ (1885, 146; see also 1884). Holmes appears to have had considerable familiarity with the sedimentary history of the lower Thames Valley, and was adamant that the position of the skeleton, despite its great depth (or, indeed, partly because of it), must be of more recent origin, though still prehistoric (ie, pre-Roman). He was also in communication with William Whitaker, who recorded many well-sections in Essex (Whitaker Reference Whitaker1886), culminating in his Geology of London, in which he concurred with Holmes regarding the skeleton's age (Whitaker Reference Whitaker1889, 467). Sir Arthur Keith (Reference Keith1915) similarly thought the find to be of ‘Neolithic’ age, though it should be noted that, as the concept of the Mesolithic was not yet widely accepted at this time (Rowley-Conwy Reference Rowley-Conwy1996), he considered this to range from 8000 to 2000 bc. With widespread acceptance of the existence of a distinct period between the ‘Old’ and ‘New Stone Age’ following Clark's seminal Reference Clark1932 publication The Mesolithic Age in Britain, King and Oakley (Reference King and Oakley1936, 69) attributed the skeleton to the Late Mesolithic based on the stratigraphy at the find location (see also Wells Reference Wells1959). Churchill (Reference Churchill1963) was able to bring additional data to bear, analysing pollen from muds in the medullary cavity of the right femur of the Tilbury skeleton, as well as of the Thatcham humerus from Wymer's (1962) excavations, discussed further below. On this basis, he attributed Tilbury to pollen zone VIIb (eg, the Subboreal, from c. 4000 bc and hence post-Mesolithic).
As with a number of other early finds of unassociated human remains claimed to be of some antiquity, interest in Tilbury waned in the following decades, not least because of Churchill's conclusions regarding its more recent age (cf. Newell et al. Reference Newell, Constandse-Westermann and Meiklejohn1979). At the same time, many supposedly early human skeletons had been found to belong to considerably later periods when radiocarbon dated. The Galley Hill skeleton, for example, was once considered a type-fossil for the Palaeolithic (Keith Reference Keith1915), yet already in 1948 newly developed fluorine relative dating tests indicated that it was probably intrusive into the gravel terrace in which it was found in 1888 (Oakley & Montagu Reference Oakley and Montagu1949). This was subsequently confirmed when radiocarbon dating placed it in the Bronze Age (BM-86, 3310 ± 150 bp: 2010–1220 cal bc) (Barker & Mackey Reference Barker and Mackey1961). ‘Hunstanton Woman’, found in 1897 in a Norfolk gravel pit and overlain by some 2 m of apparently undisturbed gravels, was initially described by Keith and reported as Palaeolithic or Mesolithic. The skeleton has now been shown to be Anglo-Saxon (OxA-4386, 1265 ± 65 bp: cal ad 650–933) (Hoare & Sweet Reference Hoare and Sweet1994). Many other examples could be given (see Meiklejohn et al. Reference Meiklejohn, Chamberlain and Schulting2011; Oakley et al. Reference Oakley, Campbell and Molleson1971). However, it cannot be assumed that all such attributions are mistaken, and so it is important to revisit earlier collections.
The Skeleton
The Tilbury skeleton was partly destroyed before Baynes’ attention was drawn to the find. Nevertheless, it appears that much of the complete skeleton was originally present, as seen by the presence of at least some small bones of the hands and feet (Owen Reference Owen1884, 4). Baynes’ account notes that the workers damaged the pelvis with a pick, though Owen (ibid., 12) commented that the surviving fragments were of ‘male proportions’. The major elements listed by Owen (ibid., 4) are those currently in the Natural History Museum, though an unspecified element was sent at that time to a colleague at what was then the British Museum, Natural History for destructive analysis (ibid., 13). Unfortunately, it has not been possible to locate three teeth – an incisor, canine, and premolar – illustrated in Owen's publication (1884, fig. 2) (Fig. 5).

Fig. 5 ‘Tilbury Man’ cranium and mandible as illustrated in Owen (Reference Owen1884). The three teeth shown have not been located in the NHM. Note that the 1884 printing has reversed the image, ie, what is shown as the left side of the skull is in fact the right, and vice versa. Lower right is part of the occipital
The extant cranial remains consist of the partial calvarium of an older adult male, together with the left half of the mandible. In the absence of the posterior dentition and the pelvis, ageing is based on the state of closure of the cranial sutures, further supported by the largely edentulous mandible fragment (Fig. 6). Sexing, again in the absence of the pelvis, is based on the overall morphology of the cranium, and in particular the pronounced supraorbital ridges, dull orbital margins, pronounced nuchal region, and sloping forehead, all of which indicate male (Ubelaker Reference Ubelaker1978). Indeed, although its brow ridges are, in fact, only moderately pronounced relative to many other European populations of comparable age, at the time the Tilbury calvarium evoked a comparison with that of the recently discovered (1856) Neanderthal skeleton, with Holmes (Reference Holmes1885, 146) commenting that with the publication of the plates in Owen's (1884) report it could be seen that ‘… the Neanderthal-like appearance of the Tilbury skull is very strikingly shown.’ (That Holmes would make this comparison is odd, since, as noted above, in the same paper he argues vigorously for a Holocene age for Tilbury Man.) The cranial length and breadth measurements are 172 mm and 136 mm respectively, yielding a ‘mesocephalic’ cranial index of 78.2. On the very limited available evidence, the Mesolithic population of Britain includes both mesocephalic and dolichocephalic crania (Fawcett Reference Fawcett1922; Humphrey & Stringer Reference Humphrey and Stringer2002; Schulting Reference Schulting2005). The roofs of the Tilbury cranium's orbits exhibit possible slight healed cribra orbitalia scars, not uncommon in prehistoric skeletal assemblages (Roberts & Cox Reference Roberts and Cox2003), and generally thought to be the result of childhood anaemia (Stuart-Macadam Reference Stuart-Macadam1992).
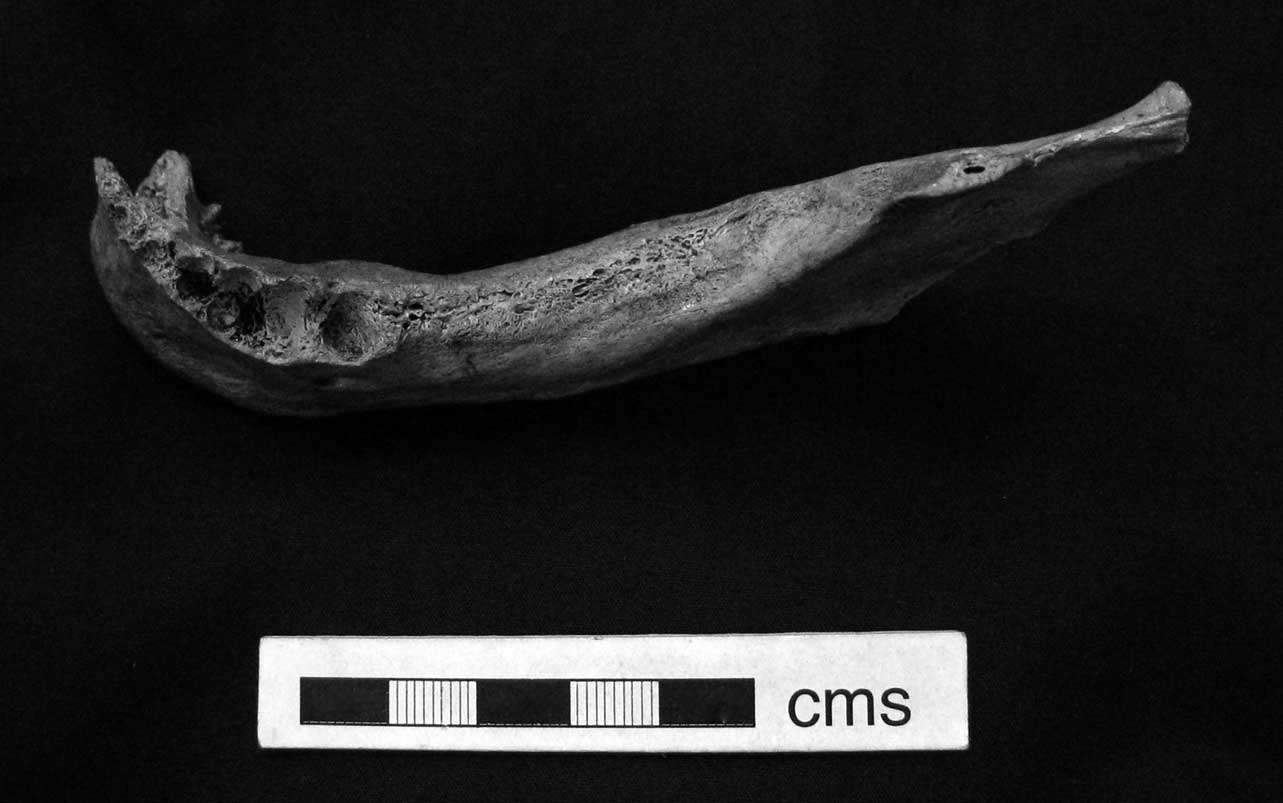
Fig. 6 Tilbury mandible (Natural History Museum, London, PA SK 9). Photo by author
The postcranial remains are limited to the major longbones, and include the shafts of both femora and tibiae, as well as one-third of a fibula shaft (Table 2; Fig. 7). All are missing their articular ends; less than half of the right tibia is represented, but the cleanly cut distal end indicates that this is the source of the tube of powdered bone present in the same box (see below). The upper limbs are represented by most of a right humerus, the proximal half (minus the proximal end) of the left humerus, a portion of the distal right ulna, and most of the left ulna and radius. The latter retain a portion of their proximal ends, but lack their distal ends entirely.
Table 2 OSTEOMETRIC DATA FOR THE EXTANT POSTCRANIAL REMAINS OF ‘TILBURY MAN’ (PV M 1913)

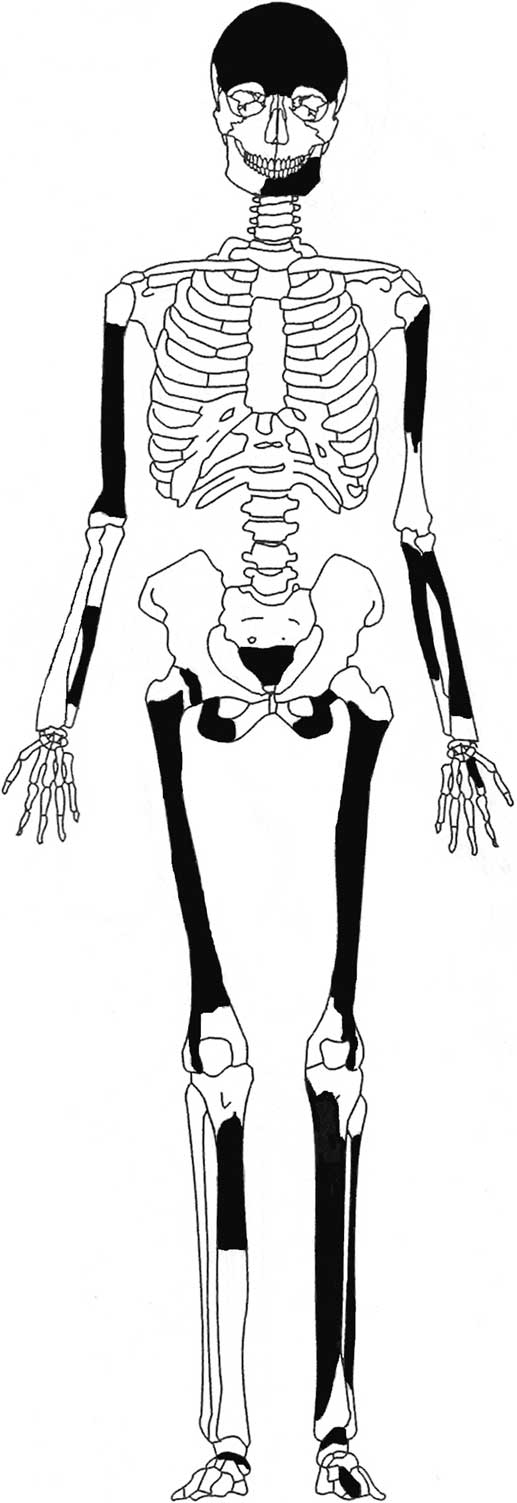
Fig. 7 Extant skeletal elements of ‘Tilbury Man’ (Natural History Museum, London, PA SK 9, PV M 1913)
Estimating the length of the incomplete right femur as c. 40.2 cm allows the calculation of a living stature of 158.8 ± 4 cm (or just over 5 ft 2 in) (using Trotter and Gleser's formula for Caucasian males, in Bass Reference Bass1987, 221). While this is shorter than the Early Mesolithic ‘Cheddar Man’ from Gough's Cave, Somerset, with an estimated stature of c. 166 cm (Holliday & Churchill Reference Holliday and Churchill2003), both individuals fit comfortably within the range for male stature of 161.3 ± 5.8 cm for the western European Mesolithic (Formicola & Giannecchini Reference Formicola and Giannecchini1999, tab. 5). Though not particularly large, the postcranial remains are those of a moderately robust individual, with well-defined muscle attachment sites. The femoral lineas aspera are very prominent, as are the interproximal crests of the tibiae, suggesting habitual use of the legs for strenuous activity. Both the femora and tibiae are flattened (platymeric and platycnemic, respectively), further supporting an active lifestyleFootnote 2 (Ruff et al. Reference Ruff, Larsen and Hayes1984; Stock Reference Stock2006). The surviving left radial tuberosity is prominent and rugous, with a central depression for the attachment of a tendon of biceps brachii, indicating repeated, powerful movements of the forearm (Hawkey & Merbs Reference Hawkey and Merbs1995). The surviving portion of the matching proximal ulna is similarly well developed.
In the absence of most of the articular ends of the long bones, as well as the vertebrae, it is not possible to assess the degree of degenerative wear on the skeleton, but considering the individual's apparently relatively advanced age, and the evidence for an active lifestyle, it is probable that such changes were present. There are slight indications of lipping on the surviving ulnar notch. The loss of the posterior dentition (including all molars and the second premolar) on the surviving left mandible is probably also age-related, since the loss of these teeth is almost certainly the result of the exposure of the pulp chambers through attrition, with subsequent abscessing and eventual loss. The original illustrations of the now lost teeth show that the premolar had been worn to an enamel ring (the accuracy of the etching is confirmed by a comparison with the surviving cranium and mandible). Caries can also lead to abscessing, but are extremely rare in north-west European Mesolithic populations likely due to the rarity of carbohydrates and sugars in the diet (Meiklejohn & Zvelebil Reference Meiklejohn and Zvelebil1991; Schulting Reference Schulting2005).
There is a small, well-healed depressed fracture on the right frontal, c. 5.8 cm above the middle of the right orbit (Fig. 8). The depression itself is round and smooth-walled, with no signs of active remodelling. It measures c. 10 mm in diameter and 1.0 mm in depth, leaving the internal vault unaffected. A second, larger depression is found on the left orbit, measuring c. 24.0 × 12.2 mm; its depth is difficult to measure due to its position across the supraorbital ridge, but it is at least 4 mm deep. Interestingly, these two features were accurately depicted on the 1884 illustrations (Fig. 5), though Owen did not comment on them.
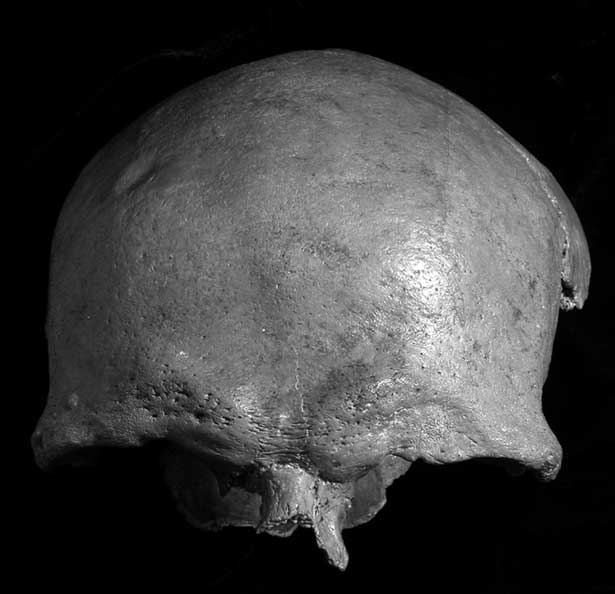
Fig. 8 Tilbury calvarium (Natural History Museum, London, PA SK 9). Note small depression on the right frontal, and depressed area above left orbit. Photo by author
Both depressions are typical of blunt force cranial injuries; very similar defects have been described for the Danish Mesolithic, for example on an adult male from Korsør Nor (Bennike Reference Bennike1997), and from the British Neolithic (Schulting & Wysocki Reference Schulting and Wysocki2005), as well as elsewhere in Europe (see papers in Schulting & Fibiger Reference Schulting and Fibiger2012). An element of conflict resolution within or between communities has often been proposed for such non-lethal cranial injuries (Lambert Reference Lambert1997; Schulting & Wysocki Reference Schulting and Wysocki2005; Standen & Arriaza Reference Standen and Arriaza2000; Walker Reference Walker1989), and may well be the case here, though of course other scenarios are plausible, such as survival of an injury of lethal intent. Nor can accidental injury be entirely ruled out, though it is not seen as the most likely explanation: falls rarely impact the cranium, and when they do they more typically result in linear fractures (Given & Williams Reference Given and Williams2002; Lambert Reference Lambert1997); in this case, the concentration of force implied by the location and shape of the injury implies two blows struck with a club-like implement. Both injuries are well healed, and it is not possible to determine whether they occurred at the same time, or on two separate occasions. The injury above the left orbit has distorted the shape of the orbit compared to the right side. It is likely that, unlike the small depression on the right frontal, it would have been visible on the living individual, and indeed may have affected his eyesight.
The condition of the surviving bone surfaces is generally good, with no indication of surface exfoliation resulting from sub-aerial weathering, nor of water rolling. Whether the remains can be said to represent an intentional burial on this basis is unclear, though it is highly probable, or even certain according to Oakley (Reference Oakley1963). Based on the skeleton's location within riverine sands, Holmes (Reference Holmes1884, 395) opts for another possibility: ‘The man probably floated down the stream, and his remains were deposited somewhere near the shore, where the current was sluggish.’ But of course a grave could equally have been dug into the sands near the river's shore, or some distance from it, as the river channel continued to migrate southwards to cut into the chalk of the Kent shore, leaving a broad alluvial plain on the north side that was to become the Essex marshes (Holmes Reference Holmes1885; Whitaker Reference Whitaker1889). There can be no definitive conclusion, given the circumstances of the discovery and the current state of the skeleton itself. If the body did float down the river, it must not have been in the water for long before becoming buried in the sands. Decomposition is greatly accelerated in well-aerated water, and the degree of articulation evident from the presence of at least parts of all the major longbones, as well as the mandible, would not be expected to persist for long (Haglund & Sorg Reference Haglund and Sorg2002). Nor is there any evidence for scavenging. The presence of some of the small bones of the hands and feet does suggest that a complete skeleton was present, which would support the interpretation of an intentional burial. The absence of grave offerings – assuming the 19th century observations are reliable – would be uncommon, though not unheard of, in a European Mesolithic context. Accepting Tilbury as such, no definite examples of other intentional burials are known in Britain from this period, with the possible exception of Gough's Cave 1, ‘Cheddar Man’, also with no recorded grave offerings, and Aveline's Hole, Somerset, Britain's only example of a sizeable Mesolithic cemetery (dating to c. 8300 cal bc). Unfortunately, the early date of the latter's investigation (from its discovery in 1797 onwards) and the loss of much material and records from later excavations during World War II means that the documentation for the site is poor. While not abundant, perforated shell beads, tooth pendants, fossil ammonites, and other objects likely constitute grave offerings, although with one possible exception, specific associations with human remains have been lost (Schulting Reference Schulting2005).
AMS Radiocarbon and Stable Isotope Analysis
Most of the bone, and particularly the calvarium, has been heavily treated with a shellac-like preservative, probably applied not long after the skeleton was accessioned. Notably, however, the right tibia shaft fragment appears untreated; as mentioned above, the distal end of this element has been cleanly cut, and is the source of the powered bone sample found in a clearly labelled glass tube inside the same box as the skeleton. This may be the remnant of the sample sent by Owen (Reference Owen1884, 13) to Dr Flight of the British Museum, Natural History for chemical tests to determine the bone's degree of fossilisation, and hence estimate its age. Alternatively, the sample may have been taken for fluorine, uranium, and nitrogen analysis in the 1950s or 1960s, a procedure to which many purportedly early skeletons were subjected at the Natural History Museum. However, the sample is much larger than would have normally been taken for this purpose and, while Tilbury is mentioned, no results for it are reported in the comprehensive Catalogue of Fossil Hominids (Oakley et al. Reference Oakley, Campbell and Molleson1971, 41–2). Thus, the former scenario is the more likely, with this being the sample sent by Owen. This powdered bone provided the material used for AMS dating and stable carbon and nitrogen isotope analysis, since 1) it derived from an untreated element, and 2) it avoided the need to take a new sample and so preserved the surviving elements.
The AMS radiocarbon determination of 6065–5912 cal bc (OxA-18781: 7120 ± 38 bp) places the skeleton within the British Late Mesolithic (Barton & Roberts Reference Barton and Roberts2004). The standard means of assessing collagen quality are all within acceptable limits (collagen yield = 6.8%, %C = 44.0%, %N = 16.0%, C:N = 3.2) (DeNiro Reference DeNiro1985; van Klinken Reference Klinken1999). Stable carbon (δ13C) and nitrogen (δ15N) isotopes were measured separately at the Research Laboratory for Archaeology and the History of Art, Oxford, with the averages of three runs reported here. The δ13C value of −19.3‰, while slightly elevated compared to a purely terrestrial endpoint in north-temperate Europe (eg, −20 to −21‰), does not show any significant consumption of marine foods when compared to results on humans of both equivalent and earlier date from Foxhole Cave and Caldey Island in south Wales (Reference Schulting, Murray, McLaughlin, Walker, Macphail, Price and FibigerSchulting et al. in press; Schulting & Richards Reference Schulting and Richards2002a), or from Oronsay on the west coast of Scotland, dating to the very end of the Mesolithic (Richards & Sheridan Reference Richards and Sheridan2000; Schulting & Richards Reference Schulting and Richards2002b). However, the δ15N value of 11.7‰ is sufficiently high to suggest some contribution of freshwater protein sources, such as fish and/or waterfowl, though it is likely that this was relatively modest (eg, compared with Mesolithic individuals from the Iron Gates, often above 14‰ (Bonsall et al. Reference Bonsall, Cook, Hedges, Higham, Pickard and Radovanovíc2004)). There is also the possibility of some minor use of marine foods, although given the environmental context of the find, estuarine species may be the most likely candidates for the slight elevation seen in both isotopes. Any significant contribution of freshwater, marine, or estuarine protein raises the possibility of a reservoir effect on the radiocarbon determination (Cook et al. Reference Cook, Bonsall, Hedges, McSweeney, Boronean and Pettitt2001; Keaveney & Reimer Reference Keaveney and Reimer2012), making the individual somewhat too old, but it is not possible to quantify the effect. However, it is considered unlikely to be large, both because of the comparatively small elevation in δ15N and because of the supporting chronostratigraphic evidence, discussed below.
The Environmental Context
There is a great deal of palaeoenvironmental information available from the immediate environs of Tilbury, since a series of well-sections recorded in the late 19th century (Whitaker 1885) together with a number of boreholes taken in the 1960s and 1970s form the basis of one of the classic sequences for the Lower Thames. Indeed, the Thames estuary type-site, borehole SB21/1, was taken approximately 200 m from the World's End Tavern at Tilbury (Devoy Reference Devoy1979, 364; Reference Devoy1982), and is also the site of one of the well-sections reported by Holmes (Reference Holmes1884). The borehole lies approximately 1 km east of the Tilbury Docks where the skeleton was discovered. A series of five biogenic layers are defined (Table 3). The arboreal pollen in the lowest, Tilbury I (TI), is dominated by Quercus, Alnus, and Corylus, but also present are Pinus, Ulmus, and low amounts of Betula. Devoy (Reference Devoy1979, 368) notes that the early and considerable presence of Alnus in TI, which commences at c. 8170 radiocarbon years bp, clashes with evidence elsewhere in the country, but reasonably accounts for this as a localised expansion given the favourable conditions for alder in the low-lying valley next to a large river. As well as the arboreal pollen, there is abundant evidence for wetland grasses and herbs, with Phragmites being particularly well-represented.
TABLE 3 RADIOCARBON DATED BIOGENIC SEQUENCE AT WORLD'S END TAVERN, TILBURY

After Devoy (Reference Devoy1979, table 1). Re-calibrated in OxCal 4.0 using IntCal 2009
Tilbury II (TII) is most directly relevant to the present discussion, as this layer is found at and above the depth of the skeleton, c. −9.44 m OD (Devoy Reference Devoy1980) (Figs 9 & 10). Since the skeleton lay within sands underlying the peat layer, there is a good possibility that, if the body was intentionally buried, TII represents the land surface at the time. TII shows a decrease in pine and the first consistent appearance of Tilia, with the other species, both arboreal and non-arboreal, from TI remaining well represented (Devoy Reference Devoy1979, fig. 6). Alder (Alnus glutinosa) continues to be a significant presence. A high occurrence of this species was also noted from the medullary cavity of the Tilbury femur (Churchill Reference Churchill1963); in fact, with the exception of the low presence of oak and the absence of Fagus and Plantago – present as single pollen grains in the femur – the pollen from TII actually matches that taken from the skeleton quite well.

Fig. 9 Radiocarbon determinations for the Tilbury SB21/1 core (Devoy Reference Devoy1979) and Tilbury skeleton

Fig. 10 Age-depth model for the Tilbury SB21/1 core (data from Devoy Reference Devoy1979). The two earliest dates (Q-1426 and 1427) define the Tilbury I regression, followed by the Thames I transgression. Above this, the next two dates (Q-1428 and 1429) mark the Tilbury II regression, which is of the most immediate relevance here, as this is the period associated with the Tilbury skeleton
Immediately underlying TII is a layer of silts and clays marking the Thames I marine transgression, dated by Devoy to 8200–6970 bp (c. 7200–5850 cal bc). There was a rapid rise in relative sea-level in the early Holocene: between 8500 and 7000 bp (c. 7550–5900 cal bc), mean high water rose from −25.5 m OD to −8.9 m OD (Devoy Reference Devoy1979), resulting in the submergence of extensive lowland areas, including, of course, the entire landmass of Doggerland (Coles Reference Coles1998; Gaffney et al. Reference Gaffney, Thomson and Fitch2007; Weninger et al. Reference Weninger, Schulting, Bradtmöller, Clare, Collard, Edinborough, Hilpert, Jöris, Niekus, Rohling and Wagner2008). A strong brackish water influence is seen in the diatoms at Tilbury from early in this sequence, pre-dating the skeleton and continuing up to its corresponding age/depth. Devoy (Reference Devoy1979, 369) interprets this as indicative of a typical littoral or estuarine saltmarsh environment. The earliest date from the overlying peat of TII is 6020–5800 cal bc (Q-1428, 7050 ± 100 bp), which corresponds very well with the dating of the Tilbury skeleton at 6065–5912 cal bc. Their depths OD also correspond reasonably well, particularly considering that the two locations are separated by a kilometre and minor local variations can be expected (see also Long Reference Long1995; Marsland Reference Marsland1986; Sidell et al. Reference Sidell, Wilkinson, Scaife and Cameron2000). The regression seems to have been quite rapid, based on a sharp decline in marine/brackish water diatoms (Devoy Reference Devoy1979). Relative sea-level in the inner Thames estuary is estimated to have been c. −12 m OD (Haggart Reference Haggart1995, fig. 79), though it is not straightforward to place the coastline very precisely, other than to note that it would probably have lain at least 10 km further east. About a millennium later, at c. 6000 bp (c. 4900 cal bc), there was another transgression, Thames II, that extended well upstream from Tilbury, with three others following over the subsequent millennia (Devoy Reference Devoy1979; cf. Bates & Barham Reference Bates and Barham1995; Stafford et al. Reference Stafford, Goodburn and Bates2012). The 6000 bp transgression resulted in the drowning of large tracts of forest, traces of which can be found at various points along the Lower Thames foreshore, including Purfleet, Erith, and Rainham, located 8–14 km upstream from Tilbury (Spurrell Reference Spurrell1899; Wilkinson & Murphy Reference Wilkinson and Murphy1995).
The above information provides us with some sense of the environment at the time of ‘Tilbury Man’. It would not have been long, perhaps a few generations, since the mouth of the estuary had migrated eastwards from Tilbury. Given the apparent rapidity of the regression, it is likely that people would have been very aware of the changes in their environment. As the marine influence receded, first saltmarsh and then Phragmites reedswamps appeared. Trees quickly colonised the low-lying areas, the more so as one progressed upstream from Tilbury, with a fen woodland of oak, alder, and lime, and abundant hazel. Reedswamps would have remained plentiful in the hollows and along watercourses, of which there were no doubt many, providing a rich mosaic of habitats. Using the channels of the Thames and its tributaries would have brought a whole range of wetland habitats within easy reach of a dugout canoe, from the estuary to the east, to increasingly developed woodlands to the west. Faunal remains collected from Tilbury II deposits in the 19th century include the usual panoply of Holocene large mammals: aurochs, red deer, wild pig, and wolf, as well as beaver (Spurrell Reference Spurrell1885), which would have been particularly well-suited to this habitat (Coles Reference Coles2006); indeed, Spurrell (1889, 218) notes the finding of a complete beaver skeleton underneath a tree trunk in the buried forest bed at Crossness. A range of freshwater and estuarine fish species would have been accessible, as would wildfowl, with migratory species being particularly abundant at certain times of the year (eg, large numbers of Brent geese and various ducks still overwinter on the Thames estuary today: Boorman & Ranwell Reference Boorman and Ranwell1977). Plant foods would also have been varied and abundant on the floodplains and in the fen woodlands. The alluvial flats extended for some 3 km to the north, with higher ground beyond. Across the river to the south were the chalklands of Kent, offering a different range of plant resources than those available on the alluvial flats. In short, this was a good place for broad-spectrum hunter-fisher-gatherers to live (cf. Jacobi Reference Jacobi1978).
As discussed above, the stable isotope data do not allow for any significant contribution from marine protein in Tilbury Man's diet. There are only two other Mesolithic humans for comparison from the lowlands of central and eastern England: Staythorpe, Nottinghamshire and Thatcham, Berkshire, each represented by only single elements. The δ13C and δ15N values for Staythorpe are −20.4‰ and 9.3‰ respectively (Richards Reference Richards2001), while those for Thatcham are −21.9‰ and 8.4‰ (Schulting & Richards Reference Schulting and Richards2000). Thus both provide entirely ‘terrestrial’ isotopic signatures, though it must be emphasised that this does not necessarily distinguish between protein from terrestrial sources proper, and freshwater sources. Marine sources do not enter into the equation at either site, since Staythorpe is 80 km from even the modern coastline, while Thatcham is 65 km from the south coast. Anadromous salmon would be accessible in rivers such as the Trent and Kennet, but are elevated in δ13C and so would be expected to affect human bone collagen measurements if they were being consumed in any quantity. The fact that this does not appear to have been the case, either here or in inland locations in Ireland (Woodman Reference Woodman2004) is interesting, and presents a strong contrast to the importance of salmon to the classic complex fisher-hunter-gatherers of the northwest coast of North America (Ames & Maschner Reference Ames and Maschner1999; Chisholm et al. Reference Chisholm, Nelson and Schwarz1982). Furthermore, the δ15N values are also comparatively low at Staythorpe and Thatcham, and clearly do not suggest a high contribution from freshwater fish, which should result in substantially higher values (>12‰) for human consumers (Bonsall et al. Reference Bonsall, Cook, Hedges, Higham, Pickard and Radovanovíc2004; Lillie & Jacobs Reference Lillie and Jacobs2006). As both finds are from riverside locations, this is perhaps surprising, but then the availability of fish must be balanced against the available terrestrial resource opportunities (cf. Winterhalder Reference Winterhalder2001).
Tilbury, on the other hand, is much nearer the sea, and, while its δ13C value is slightly elevated compared to either Staythorpe or Thatcham, and therefore may indicate a small contribution of marine protein, it can still be characterised as strongly ‘terrestrial’ (cf. Schulting Reference Schulting2011). This is not necessarily surprising: as the estuary receded eastwards, the area from Tilbury upstream would have been a productive freshwater wetland, with abundant fish and wildfowl. Eel runs, for which the Thames was well-known historically, may have been particularly important seasonally, given their large numbers during migrations, their high fat content, and the ease with which they may be preserved by smoking at least for brief periods (Wheeler Reference Wheeler1979). Further downstream, the size of the Thames would still make it the dominant influence, creating brackish estuarine conditions with relatively low salinity. Species feeding in estuaries are highly variable in their stable carbon isotope signatures, but in the absence of a ‘hardwater’ carbonate effect they are always intermediate between freshwater and marine values (Deegan & Garritt Reference Deegan and Garritt1997; Fry Reference Fry2002; Thornton & McManus Reference Thornton and McManus1994). With the organic input of the Thames, it is likely that any organisms from its waters even near the mouth of the estuary were relatively depleted in δ13C compared to more open marine conditions. Thus there may be a stronger contribution from fish in the Tilbury human isotope measurements than is at first apparent. But whether they dominated the protein component of the diet in this individual is another matter, since the rich riverside woodlands would have provided abundant alternatives, and in much larger packages. Aurochs, for example, would favour such low-lying habitats. There would have been many plant foods as well, including starchy tubers such as bulrush (Typha latifolia). Their contribution, however, is under-represented in stable isotope measurements on bone collagen, which is biased towards the protein component of the diet (Ambrose & Norr Reference Ambrose and Norr1993; Jim et al. Reference Jim, Jones, Ambrose and Evershed2006), and starchy tubers generally score low in this regard. Bulrush pollen is also edible (as is the stalk), and would be expected to contain higher protein levels. In fact, cattail (the more common North American term for Typha latifolia) was so important to some Native American groups that wars were fought over controlling access to the most productive marshes (Liptay Reference Liptay1989).
The Archaeological Context
Despite its potential attractiveness for hunter-gatherers, there are few Mesolithic sites known from the lower Thames. The depth – c. 10 m – at which the Tilbury skeleton was found suggests why: the relevant land surfaces are deeply buried under many metres of alluvium (cf. Rackham Reference Rackham1994). A dugout canoe and two paddles were reportedly found (not together) in the first half of the 20th century in wetlands at Lion Point, between the rivers Stour and Colne. The paddles were described as being very similar to Danish Mesolithic examples from Holmegaard (Warren et al. Reference Warren, Piggott, Clark, Burkitt and Godwin1936); however, there are only so many ways to make a paddle, and one found in 1983 has been dated to the Bronze Age (Wilkinson & Murphy Reference Wilkinson and Murphy1995). Other undated logboats are known from Erith, Woolwich, Murston, and the marshes near Maidstone in Kent, while a paddle was found near Swanscombe together with two tranchet flint axes, or ‘Thames picks’ (Ashbee Reference Ashbee2005, 49, 79). Regardless of the dating of these specific finds, dugouts were doubtless indispensable for the exploitation of this watery world, and are well-documented in the Danish Mesolithic (Christensen Reference Christensen1997). Unfortunately, Mesolithic finds near the river are rare, and are generally found in disturbed contexts during building projects (eg, Lavender Reference Lavender1998; Lewis Reference Lewis2000). A more substantial flint assemblage including microliths, though mixed with Neolithic flints and pottery, comes from Ebbsfleet, just across the river from Tilbury (Burchell Reference Burchell1938; Sieveking Reference Sieveking1960). More recent is the discovery of a large Late Mesolithic flint assemblage – suggesting a specialised production site for tranchet axes – on sands under 2 m of peats along the Bronze Age Way, Erith, dated to c. 4550–4265 cal bc (Beta-88688, 5570 ± 70 bp) (Bennell Reference Bennell1998; Lewis Reference Lewis2000). Away from the valley, and its covering alluvium, large early collections of microliths are known from the hills to the north-east, around Rayleigh and Thundersley, found during the course of sand and gravel extraction (Jacobi Reference Jacobi1980; Warren et al. Reference Warren, Piggott, Clark, Burkitt and Godwin1936). More recent lithic collections of Mesolithic character are also known from this area (eg, Crowe Reference Crowe1992). Further to the east, Mesolithic flints have been recorded in and around Greater London (Lewis Reference Lewis2000; Sidell et al. Reference Sidell, Cotton, Rayner and Wheeler2002). From the river itself have been dredged a number of ‘Thames picks’ (Lacaille Reference Lacaille1961) and bone and antler harpoons and mattocks, examples of which have been directly dated to the Mesolithic (Bonsall & Smith Reference Bonsall and Smith1990).
Nearer the present coast, the land–sea interface has been active throughout the Holocene, and sea-level rise and continuing coastal erosion together have no doubt led to the inundation or destruction of many sites. The timing of the opening of the English Channel is a complex issue, but can probably be placed within the range 7000–6500 cal bc (Lambeck Reference Lambeck1995; Weninger et al. Reference Weninger, Schulting, Bradtmöller, Clare, Collard, Edinborough, Hilpert, Jöris, Niekus, Rohling and Wagner2008). This is important, as it suggests that the Thames would have been emptying directly to the east by the time of ‘Tilbury Man’, rather than turning south. Some sites have been discovered on the east Essex coast, most notably at Hullbridge, where Reader (1911, in Heppell et al. Reference Heppell, Brown and Murphy2004) reported a large Mesolithic assemblage from the intertidal zone. A large Mesolithic flint assemblage and hearths from Lower Halstow in Kent may include a Late Mesolithic component (Burchell Reference Burchell1928), in which case it would be near-coastal, but the collection is difficult to assess given the circumstances and early date of its discovery, and the highly selective recovery of larger objects (Jacobi Reference Jacobi1982). Situated on the bank of a palaeochannel of the Medway, the site was buried under 1.83 m of peats and marsh clays. Deeper still, charcoal concentrations, fire-cracked flint, and struck flints were found at c. −9 m OD during construction of the Medway Tunnel, with dates from this level of c. 6200–5800 cal bc (Beta-66456: 6930 ± 70 bp and Beta-66457: 7290 ± 80 bp) (Barham et al. Reference Barham, Bates, Pine and Williamson1995), again highlighting the difficulty of finding low-lying sites in this landscape.
Roger Jacobi provides useful reviews of the Mesolithic archaeology of both Essex (1980) and Kent (1982) as known in the late 1970s. Wymer's (Reference Wymer1977) gazetteer records numerous find locations. Very little Mesolithic archaeology was discovered during the building of the Channel Tunnel Rail Link, largely confined to a few isolated lithics diagnostic of the period (eg, Wessex Archaeology 2000). Ashbee (Reference Ashbee2005, 77–86) provides a recent overview of Mesolithic sites in Kent.
Burial in the British Mesolithic
Assuming, as seems likely, that Tilbury represents an intentional burial, what does this tell us about mortuary practices at this time? Aside from the scattered and fragmentary finds from the Oronsay shell middens (Meiklejohn et al. Reference Meiklejohn, Merrett, Nolan, Richards and Mellars2005), the Late Mesolithic is a period for which human skeletal finds are very rare in Britain (Fig. 11). In fact, counter-intuitively, there are far more human remains known from the Early Mesolithic, primarily due to the long history of interest in, and investigation of, caves in the Mendip hills, Somerset and along the south coast of Wales (Schulting Reference Schulting2005; Reference Schulting2009; Schulting et al. Reference Schulting, Gardiner, Hawkes and Murray2010; Schulting & Richards 2002; see overview in Meiklejohn et al. Reference Meiklejohn, Chamberlain and Schulting2011). If the practice of using river valley locations both for settlement and for burial was widespread in the Late Mesolithic, it may go some way towards accounting for their paucity in the archaeological record (cf. Blockley Reference Blockley2005). The degree of alluviation and other depositional processes at Tilbury resulted in the burying of an old land surface under some 10 m of sediment. While this may be an extreme case, similar processes occurring further upriver, and along smaller rivers, may have resulted in destruction or invisibility either through the erosion of riverside burials on the one hand, or their burial under metres of sediments on the other. If coastal settlement was emphasised at this time – and there are good reasons for supposing this to be the case – then the inland population density is likely to have been lower, as well as perhaps being more mobile. What this does not account for, of course, is the paucity of Late Mesolithic human remains in caves near the coast in western Britain (though there are rare exceptions, such as Foxhole Cave, south Wales: Reference Schulting, Murray, McLaughlin, Walker, Macphail, Price and FibigerSchulting et al. in press).

Fig. 11 Map showing the location sites with directly dated (and assumed, in the case of Thatcham) Mesolithic human remains in Britain and Ireland. There is some question over the status of Bower Farm (see Meiklejoen et al. 2011, 34)
The possibility of riverside burial in the Late Mesolithic receives further tentative support from an isolated but well-preserved and uneroded human adult (female?) left femur from a palaeochannel of the River Trent, near Staythorpe, Nottinghamshire. The femur has been directly dated to 5733–5630 cal bc (Beta-144016: 6790 ± 40 bp; Myers Reference Myers2006), placing it slightly later than Tilbury. That the practice of riverside burial may not have been restricted to the Late Mesolithic is suggested by an isolated adult left humerus recovered during excavations at the Early Mesolithic site of Thatcham in the Kennet valley, Berkshire (Wymer Reference Wymer1962). While radiocarbon dates from the site as a whole span c. 10,000–8000 cal bc (Ambers & Bowman Reference Ambers and Bowman1998; Burleigh et al. Reference Burleigh, Ambers and Matthews1982; Churchill Reference Churchill1962; Gowlett et al. Reference Gowlett, Hedges, Law and Perry1987; Hedges et al. Reference Hedges, Housley, Law and Perry1988), many individual determinations are problematic, either in terms of the material dated (eg, bulk charcoal) or its associations. In any case, the humerus itself has never been directly AMS dated (Meiklejohn et al. Reference Meiklejohn, Chamberlain and Schulting2011). Emphasising the need for this is the fact that the bone was actually found in redeposited shell marl overlying the peat deposits that contained most of the cultural remains (Churchill Reference Churchill1962, 364). Also worth noting is this context is the recent dating of two human crania from Greylake, Somerset, to the Early Mesolithic, c. 8450–8300 cal bc (Wk-30930: 9118 ± 37 bp; Wk-30931: 9134 ± 37 bp) (Brunning & Firth Reference Brunning and Firth2012). These and other human remains, some now lost, were recovered during quarrying on what would have been a small islet in the floodplain of the Somerset Levels during the early Holocene.
Aside from the above-mentioned exceptions, Mesolithic human remains in Britain and Ireland have been primarily found in caves and rockshelters (Chamberlain Reference Chamberlain1996; Dowd Reference Dowd2008; Schulting Reference Schulting2005; Schulting et al. Reference Schulting, Gardiner, Hawkes and Murray2010; in press; Schulting & Richards Reference Schulting and Richards2002a). The Oronsay and Ferriter's Cove shell middens from the west coast of Scotland and south-west Ireland respectively, provide another context for isolated human bone finds, some of which may be the disturbed remains of burials, while others may reflect the practice of excarnation (Conneller Reference Conneller2006; Meiklejohn et al. Reference Meiklejohn, Merrett, Nolan, Richards and Mellars2005). It is very likely that more human remains would be known from shell middens were more of the contemporary coastline available. Eastern England is poorly served in both cases: for the most part it lacks caves, and rising sea-levels combined with relatively shallow waters have resulted in the loss of the Early and Mid-Holocene coastlines. In addition, soft-shore coastlines may not provide the more strongly localised concentrations of shellfish found on rocky shores that encourage the repeated exploitation of specific locales, and hence midden accumulation. But away from the coasts, and in the absence of caves, burial practices in the lowlands of Britain would probably have focussed on the river valleys that were the main foci of life as well as of death. Chance finds like Tilbury provide a brief glimpse into this world.
Acknowledgments
Many thanks to Margaret Clegg and Robert Kruszynski of the Natural History Museum, London, for facilitating access to the Tilbury skeleton, and to the Museum for permitting sampling. Figures 2 and 3 are used with kind permission of the Museum of London, Docklands. Thanks also to Tom Higham and Peter Ditchfield for AMS radiocarbon dating and mass spectrometry. Funding for laboratory analysis was provided through the Leverhulme Trust (grant no. F/08 735/E).














