Paleodemographic analyses of archaeological human populations represent one of the pillars in the reconstruction of the biological and cultural history of our predecessors. Despite their importance, they are often limited by intrinsic and extrinsic factors, which weigh importantly in Maya bioarchaeology. Wright and White (Reference Wright and White1996:156) cogently pointed out that there are very few archaeological skeletal collections large enough to adequately permit “a serious attempt at osteological paleodemography,” mainly (albeit not only) due to underrepresentation of certain ages and sexes (Konigsberg and Frankenberg Reference Konigsberg and Frankenberg1992; Meindl and Russel Reference Meindl and Russell1998; Wood et al. Reference Wood, Milner, Harpending and Weiss1992). Another limiting factor is the lack of information about the age structure of the ancient population at a given time (i.e., the age distribution profile that mimics the reference population; Bocquet-Appel and Masset Reference Bocquet-Appel and Masset1982, Reference Bocquet-Appel and Masset1985, Reference Bocquet-Appel and Masset1996; Hoppa and Vaupel Reference Hoppa, Vaupel, Hoppa and Vaupel2002; Ortega-Muñoz Reference Ortega-Muñoz2004; Sattenspiel and Harpending Reference Sattenspiel and Harpending1983). Another limitation of paleodemography is the reduced reliability of age-at-death estimations among adults, with widening brackets as age progresses beyond the 50-year threshold (Bocquet-Appel and Masset Reference Bocquet-Appel and Masset1982).
Yet probably the single most important limitation is our uncertainty about ancient population growth rates, so that our initial approaches must rely instead on idealized stationary population scenarios, which are distant from ancient reality at best. In such scenarios, the population is static, with equal birth and death rates and a constant growth rate equal to zero (r = 0; no increment or reduction in population over the years). In stationary populations, the mean age-at-death corresponds exactly to life expectancy at birth. Therefore, the distribution of age-at-death will be equivalent to the column of deaths (dx) and can be used to establish a hypothetical life table (Johansson and Horowitz Reference Johansson and Horowitz1986). When a population is not stationary, however, the age-at-death distribution is largely dependent on changes in fertility and only to a smaller degree on mortality (Aguirre Reference Aguirre, Bringas, Hiram and Menkes1999; Sattenspiel and Harpending Reference Sattenspiel and Harpending1983; Wood et al. Reference Wood, Milner, Harpending and Weiss1992).
Since the 1900s, paleodemographic analyses have been conducted on a number of ancient settlement populations of the northern and southern Maya Lowlands (Civera-Cerecedo Reference Civera-Cerecedo, Jaén, López-Alonso, Márquez-Morfín and Hernández-Espinoza1998; Hernández-Espinoza and Márquez-Morfín Reference Hernández-Espinoza, Márquez-Morfín, Hernández-Espinoza and Márquez-Morfín2007, Reference Hernández-Espinoza and Márquez-Morfín2015; Márquez-Morfín et al. Reference Márquez-Morfín, Hernández-Espinoza and González-Licón2006; Márquez-Morfín and Hernández-Espinoza Reference Márquez-Morfín and Hernández-Espinoza2013; Ortega-Muñoz Reference Ortega-Muñoz2007, Reference Ortega-Muñoz and Cucina2015; Saul Reference Saul1972; Storey Reference Storey1992; Tiesler et al. Reference Tiesler, Cucina, Sierra-Sosa, Falla and Meindl2005; Whittington Reference Whittington1991). The results of these studies highlight divergent population dynamics across cultural periods and geographic spans within the larger Maya area, with no clear overall pattern of fertility change throughout the centuries. Such heterogeneity is more evident when comparisons do not take into consideration variables such as settlement size or subsistence patterns, yet, at least among large settlement populations, some rough comparisons seem tenable. From this perspective, Hernández-Espinoza and Márquez-Morfín (Reference Hernández-Espinoza and Márquez-Morfín2015) concluded cautiously that, during the Late to Terminal Classic (AD 550–900), Maya coastal settlement populations experienced a life expectancy at birth that fluctuated between 20 and 42.2 years, with an infant mortality rate around 30% and an overall high fertility of six and more children per woman. On the contrary, the same authors calculated from Classic urban skeletal series like Palenque's or Copan's that, during the same time period, life expectancies at birth were between 25.5 and 26.1 years, infant mortality rates were between 13.3% and 23.3%, and there was a total fertility rate of about five children per woman (Hernández-Espinoza and Márquez-Morfín Reference Hernández-Espinoza and Márquez-Morfín2015).
Within the complex scenario of paleodemographic parameters, we attempt to overcome some of the technique's limitations by focusing on a large and relatively homogeneous burial collection from Xcambó, Yucatan. In contrast to earlier studies for which the use of divergent parameters like time period, subsistence, and geographical location has hampered reliable inferences and comparisons with other site populations, our analysis of one single, continuous occupation during two chronological periods provides the opportunity to delve deeper into the dynamic cultural and biosocial undercurrents affecting population structure.
For about five centuries during the first millennium AD, Xcambó thrived as a small Maya coastal port, nested on the northern shores of the peninsula of Yucatan, Mexico. It was inhabited continuously from the Early to Late Classic period (AD 250–750; Figure 1), only to be abruptly abandoned by AD 750, presumably because of shifts in the regional political sphere of influence (Sierra-Sosa Reference Sierra-Sosa2004). This prehispanic settlement was excavated intensively and extensively between 1996 and 1999 (Sierra-Sosa Reference Sierra-Sosa2004). The location of Xcambó and the characteristics of the soil made possible the preservation of some 600 skeletons of both sexes and representative of all ranges of age-at-death. Moreover, the detailed contextual and chronological information permitted the hypothetical archaeological re-creation of population density and growth during the Early (AD 250–550) and Late Classic (AD 550–750; Ortega-Muñoz et al. Reference Ortega-Muñoz, Andrea Cucina and Sierra-Sosa2018). In turn, this imbricated contextual reconstruction provides the basis for addressing the structural paleodemography of Xcambó's settlers, based on the age-at-death and sex distribution of the skeletal sample.
Figure 1. Map of the peninsula of Yucatan, indicating the location of the site of Xcambó and other regional important sites.
Relying on Ortega-Muñoz and coauthors’ (Reference Ortega-Muñoz, Andrea Cucina and Sierra-Sosa2018) archaeodemographic approach, and given the considerable size, documentation, and preservation of this skeletal series, this article reconstructs and discusses the demographic dynamics that occurred at the site during the two phases of occupation. As we argue in detail later, these two phases, despite being continuous in time, are characterized by different socioeconomic and living conditions (Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014). By exploring two different scenarios of low and high growth rates for each period of occupation (Ortega-Muñoz et al. Reference Ortega-Muñoz, Andrea Cucina and Sierra-Sosa2018), our research design is able to assess whether and to what extent the site's socioeconomic shift from a low-density population settlement, devoted to fishing and salt production, to a densely settled, relatively autonomous administrative center of salt production, administration, and trade affected the demographic characteristics of the local population.
Xcambó
Xcambó was a settlement built over a small artificial peten (mound) within the coastal marshlands about 1 km from the northern shores of the peninsula of Yucatan, Mexico. Despite the zoonotically charged (disease-prone) marshland environs, the mound appears to have been a favorable settlement location, given its available drinking water (provided by a chultun and five waters springs), and the ready availability of perishable and nonperishable construction materials (Sierra-Sosa Reference Sierra-Sosa2004:94–96). The horizontal dimensions of the mound are 500 × 1,800 m; the settlement occupied 700 m from east–west and 150 m from north–south (Sierra-Sosa Reference Sierra-Sosa2004:97).
The buildings are arranged along the east–west axis of the mound, and their pattern is nucleated. During the Early Classic (AD 250–550), the settlement had modest civic, religious, and administrative structures in the Megalithic style. The main square was erected in the middle of the mound. Around it, residential dwellings were built, along with numerous warehouses.
During the Late Classic (AD 550–700/750), the square was eventually surrounded by 11 important structures, in which were performed civic, religious, and administrative activities, and the number of residential mounds increased. Some 250 m to the east, three public structures, which were built during the Late Classic period, form a smaller square. A sacbe (unpaved white road) goes from this smaller square to a residential area at the easternmost side and limits of the settlement. The warehouses that characterized the early phase of occupation were relocated near the salt mines (Sierra-Sosa Reference Sierra-Sosa2004:96, 97).
People of a somewhat high economic standing lived in its large dwellings around the main square. There is no archaeological evidence for small perishable huts, which would have been located in small localities close to the site, such as Xtampu, Misnay, and San Bruno. People living in Xcambó must have been intensively engaged in trading salt and in maritime procurement. A pier was built at the estuary that served as an anchoring spot and refuge for boats (Sierra-Sosa Reference Sierra-Sosa2004). Apparently, crops came in from Cemul and Misnay, two localities near Xcambó, because no direct evidence of agricultural activities in and around the settlement was found. An additional source of food procurement was fishing, the remnants of which were extensively documented during the archaeological explorations. Salt production was carried out in the nearby salt mines of Xtampac (Sierra-Sosa Reference Sierra-Sosa2004:106, 107, 115).
By the Late Classic (ca. AD 550), the number of coastal settlements had increased, and more people inhabited them (Sierra-Sosa Reference Sierra-Sosa2004:107). Likely because of its role as a salt production and administrative center, Xcambó became an important political, economic, and commercial center. As indicated by diversified isotopic profiles in dental strontium, the site experienced immigration from both inside and outside of the peninsula (Cucina et al. Reference Cucina, Sierra-Sosa, Tiesler, Charlotte Arnauld, Pereira and Beekman2021; Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014). The inhabitants reused earlier structures, relocating warehouses close to the salt mines outside the mound, and expanded the settlement to occupy almost the whole mound (Sierra-Sosa Reference Sierra-Sosa2004). Its social organization was more complex, and administrative activities gained importance, especially for local men (Maggiano et al. Reference Maggiano, Schultz, Kierdof, Sierra-Sosa, Maggiano and Tiesler2008).
By around AD 700–750, the region saw the rise of Isla Cerritos, a settlement ruled by Chichen Itza that functioned as a major trade center in connection with different territories along the Gulf of Mexico and the Mexican and Guatemalan Petén. It represented a major shift in the sociopolitical sphere of influence in the region, which led to the sudden abandonment of Xcambó, as we concluded in earlier work (Sierra-Sosa Reference Sierra-Sosa2004:164–165).
The people of Xcambó were not free from infectious diseases (Cucina Reference Cucina2011; Cucina et al. Reference Cucina, Tiesler and Sierra-Sosa2003; Méndez-Collí et al. Reference Méndez-Collí, Sierra-Sosa, Tiesler and Cucina2009) but enjoyed a balanced diet, as indicated by a relative lack of metabolic disease. Xcambo's male population did not engage in much strenuous physical activity, which seems to indicate a high level of involvement in administrative and service activities, a trend particularly noticeable during the Late Classic period (Maggiano et al. Reference Maggiano, Schultz, Kierdof, Sierra-Sosa, Maggiano and Tiesler2008). Some changes in dietary patterns might have occurred over time. In the Early Classic, the diet was likely based on marine resources and crops. In the Late Classic, the significant increase in carious lesions both for males and females suggests the introduction of new cariogenic foods in the diet, such as honey (Cucina, Perera, et al. Reference Cucina, Perera, Sierra-Sosa and Tiesler2011); this increase in carious lesions was not due to a heavy reliance on maize. Inhabitants tended to manifest closer biological affinities with other coastal sites, rather than with inland populations of the peninsula (Cucina Reference Cucina and Cucina2015). During the Late Classic, the population of the site must have been more diverse than during the previous time span. This idea is supported by the diversification in strontium isotope ratios, trace elements, dental morphology (Cucina, Tiesler, et al. Reference Cucina, Tiesler, Sierra-Sosa and Neff2011; Cucina et al. Reference Cucina, Sierra-Sosa, Tiesler, Charlotte Arnauld, Pereira and Beekman2021; Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014), and artificial cranial forms (Tiesler Reference Tiesler2005, Reference Tiesler and Cucina2015), confirming a heterogeneous collective setup in terms of identity and geographic origins.
Material and Methods
An assemblage of about 600 well-documented skeletons, excavated during the 1996–1999 field seasons, was used to elaborate a standard life table. Sex was determined based on the extent of sexual dimorphism in the coxal bone and cranial vault (Buikstra and Ubelaker Reference Buikstra and Ubelaker1994). Juvenile and subadults could not be sexed. Age-at-death of subadults was estimated by dental crown and root formation and eruption (Ubelaker Reference Ubelaker1978), whereas for the adult segment of the population, pubic symphysis, the auricular surface of the ilium, and cranial vault were analyzed (Buikstra and Ubelaker Reference Buikstra and Ubelaker1994). Coxal bones and cranial vaults were seriated by the degree of remodeling to adjust the estimated age-at-death distribution (Lovejoy et al. Reference Lovejoy, Meindl, Mensforth and Barton1985).
Table 1 shows the size of the samples by chronology. In the initial assessment, 130 individuals could not be assigned specifically to the Early or Late Classic period but were assigned instead to a more general “Classic” period; eventually these individuals were proportionally distributed into the Early and Late Classic groups. Similarly, 31 of the 81 individuals dated to the Early Classic sample could not be sexed, nor could their age be estimated, or both; the same problem occurred in 166 individuals belonging to the Late Classic. We then distributed proportionally all individuals of undetermined sex or age-at-death to the subadult or adult age classes within their own chronological period. Finally, to develop life tables, we applied the  ${1 / {16}}$ technique to smooth the curve (Wunsch Reference Wunsch1992). This resulted in a total of 201 males (38 assigned to the Early Classic and 163 to the Late Classic) and 169 females (25 and 144, respectively, for the Early and Late Classic).
${1 / {16}}$ technique to smooth the curve (Wunsch Reference Wunsch1992). This resulted in a total of 201 males (38 assigned to the Early Classic and 163 to the Late Classic) and 169 females (25 and 144, respectively, for the Early and Late Classic).
Table 1. Original Data and Corrected Age-at-Death Distributions of Early and Late Classic Series by Age Groups.
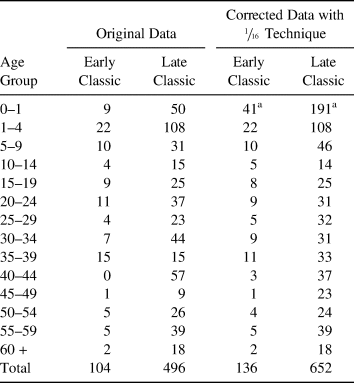
a Data elevated at 30% of total deaths.
We decided to start our study with life tables, because they are the most useful tool in formal demography (Gage Reference Gage1988; Gómez de León Reference Gómez de León, Bronfman and de León1988, Reference Gómez de León, Márquez-Morfín and de León1998; Moore et al. Reference Moore, Swedlund, Armelagos and Swedlund1975; Pressat Reference Pressat1967). For this study, we elaborated life tables using 5-year age intervals up to age 60 and beyond. Theoretically, the deaths of individuals younger than 5 years of age-at-death represent between 36% and 43% of all deaths (Woods Reference Woods2007), and demographic analyses have shown that in past societies the infant mortality rates did not change drastically until the eighteenth century (Alesan et al. Reference Alesan, Malgosa and Simó1999; Bourgeois-Pichat Reference Bourgeois-Pichat1951; Brothwell Reference Brothwell1987; Gage Reference Gage1990). As Table 1 shows, the group of individuals dying from birth to 5 years of age is 29.7% for the Early Classic (with 8.7% of individuals dying before age 1) and 31.9% for the Late Classic (with 10.1% of individuals dying younger than age 1). We decided to increase the number of individuals younger than age 5 years for that age group to represent 46% of the whole population; doing so permitted the 0–1 year age class to represent 30% of all deaths, which is in agreement with a pre-transitional demographic profile.
Growth rates were applied to build life tables of stable populations. The growth rates were based on the archaeodemographic analysis of Xcambó, which estimated the total volume of the population based on the number and distribution of archaeological households (Ortega-Muñoz et al. Reference Ortega-Muñoz, Andrea Cucina and Sierra-Sosa2018). The authors estimated a minimum and a maximum population volume for both periods of occupation, calculating growth rates of 0.08% to 0.11% in the Early Classic period and of 0.15% to 0.20% in the Late Classic (Table 2). Adding to these scenarios, we calculated a third model under the scenario of a stationary population. By applying such growth rates to life tables, we calculated life expectancy at every age, infant mortality rate (IMR), gross reproduction rate (GRR), net reproduction rate (NRR), total fertility rate (TFR), crude death rate (CDR), and crude birth rate (CBR) based on Weiss's model (Reference Weiss1973).
Table 2. Demographic Simulations with Life Tables of Both Sexes.

Notes: The scenarios follow the model of high growth rates (r = 0.11 and 0.20 for Early and Late Classic collections, respectively), stationary populations (r = 0), and low growth rates (r = 0.08 and 0.15 for Early and Late Classic collections, respectively).
Subsequently, we contrasted Weiss's (Reference Weiss1973) classic models with a new methodological approach based on the charts developed by Séguy and Buchet (Reference Séguy and Buchet2013). This calculation procedure is founded on general life tables, but calculates the probability of death (nqx) by applying the P indicator, which corresponds to the ratio between individuals who died in the 5–19 year range and those who died at age 20 and older (Boquet-Appel Reference Bocquet-Appel2002:639); in this way, it obtains the juvenile index, life expectancy at birth and at age 20 for the whole population and by sex, the growth rate (r), and the crude birth rate.
Results
According to our archaeodemographic reconstruction, the Early Classic period is characterized by lower demographic growth than the Late Classic (Ortega-Muñoz et al. Reference Ortega-Muñoz, Andrea Cucina and Sierra-Sosa2018), a trend that matches the archaeological evidence (Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014). However, application of the P indicator produces higher r values (Tables 2 and 3). The corresponding survival curve (Sx) shows that both periods produce similar mortality patterns until age 10 to 15 years (Figure 2). After this age range, and all the way to age 60, the curves tend to separate, because Late Classic individuals apparently present higher rates of survival than their Early Classic counterparts.
Table 3. Demographic Simulation according to the Model Proposed by Séguy and Buchet (Reference Séguy and Buchet2013) and Bocquet-Appel (Reference Bocquet-Appel2002).


Figure 2. Survival curve by age for the Early and Late Classic periods at Xcambó.
Figure 3 shows the quartile distribution of age-at-death in the Early and Late Classic. In both periods, the first quartile (75%) indicates that one-fourth of the population died before reaching age 1 (age 0.84 and 0.89 years for the Early and Late Classic, respectively). The Late Classic group reaches a slightly older age at the fiftieth quartile (50%), even though in both cases half of the population died before reaching age 10 (at 8.32 and 9.15 years of age for the earlier and later phases of occupation, respectively). A more noticeable difference can be appreciated only at the last survival quartile (when only 25% of the initial population is still alive): both periods are separated by about 7 years in average remnant life span (31.31 years for the Early Classic vs. 38.02 for the Late Classic). It is important to emphasize that the artificial adjustment in the categories of age between 0 to 5 years tends to level out only the differences between periods within the first quartile: it does not affect the differences that can be highlighted in the other quartiles.

Figure 3. Quartiles and median age-at-death of survivorship for the Early and Late Classic periods at Xcambó.
The criterion of mortality force (μx) indicates the magnitude of mortality. During the Early Classic, mortality was higher than in the Late Classic in almost all ages from 5 to 45 years. The deaths between 15 to 40 years increase steadily and are proportionally larger for the adults of the Early Classic period. The increase in deaths among adults can be explained by associating it with maternal mortality in females in the Early Classic. The mortality force is similar for both periods from age 45 to the few individuals who survived beyond 50 years of age (Figure 4).

Figure 4. Mortality force by age-at-death for the Early and Late Classic periods at Xcambó.
As displayed in Figure 5, the pattern of life expectancy (ex) is fairly similar between the periods, even though the Late Classic period presents higher values between birth and age 35. After this age, the two periods score similarly. That said, life expectancy at birth is noticeably different between the time periods when calculated under stable population assumptions (with positive r assumed as minimum and maximum growth rates). In both cases, Late Classic newborns experience about three more years in expected average life than the Early Classic newborns. Life expectancy simulated under a stationary scenario (r = 0) shows an increase of 2.6 years during the Late Classic (16.2 vs. 18.8 years; Table 2).

Figure 5. Life expectancy by age-at-death for the Early and Late Classic periods at Xcambó.
Finally, by applying Séguy and Buchet's methodological proposal, which considers a growth rate of 1%, Xcambó's Late Classic dwellers are expected to live 5.3 years more than their Early Classic counterparts (Table 3). With the same methodology and in a stationary scenario, expected life expectancy drops to 14.2 years at birth for the Early Classic and to 18.8 years for the Late Classic. Regardless of which estimated growth rate is applied, all simulations show better life expectancy during the Late Classic period, when each individual is predicted to live between 18 and 23 years, in comparison with the Early Classic, a time when people are expected to live roughly between 14 to 18 years.
Regarding life expectancy in adults, measured by e20 under each scenario, Late Classic adults are expected to survive about four years longer than adults during the Early Classic. As Tables 2 and 3 show, e20 ranges from 16.7 to 19.1 in the earlier phase of occupation, compared to 20.7 to 23.6 in the later period.
Infant mortality rate (IMR) is consistently higher in the Early Classic cohort, as we see in both of the paleodemographic scenarios (Tables 2 and 3). The differences are higher when Séguy and Buchet's demographic simulation is applied, with about 40 deaths per thousand more in the earlier phase (Table 3). When the other growth rates are applied, differences range between 8 and 13 deaths per thousand (Table 2). According to this pattern, CDR was higher in general during the Early Classic, with eight to nine more deaths than in the Late Classic both under stable and stationary scenarios (Table 2).
The age-at-death distribution and the age-at-death mean explain more about the pattern of fertility than the pattern of mortality (Sattenspiel and Harpending Reference Sattenspiel and Harpending1983; Wood et al. Reference Wood, Milner, Harpending and Weiss1992). Despite a demographic “explosion” during the Late Classic, as documented by Sierra-Sosa and coauthors (Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014) and by Ortega-Muñoz and colleagues (Reference Ortega-Muñoz, Andrea Cucina and Sierra-Sosa2018), the distribution of age-at-death during the Late Classic displays a lower CBR than in the Early Classic (52.99 births per thousand inhabitants vs. 61.43 per thousand under a stable population scenario when the higher growth rates are applied). The other simulations (stationary and with Bocquet-Appel's P indicator) are similarly patterned (Tables 2 and 3). GRR and TFR are in agreement with CBR, with fertility decreasing between 0.7 and 1.4 children per woman in the Late Classic. However, TFR, GRR, and NRR indicate that, in both periods, fertility granted population replacement (Table 2). Like fertility, the mean age-at-death at 20 years (a20) is higher in the Late Classic segment of the skeletal population (41.8 years) than in the Early Classic one (37.7 years; Table 3). The mean adult age-at-death is more differentiated, showing a distinctive scenario when analyzed by sex. Although the mean age-at-death of males did not change through time, females increased a20 from 35 years to almost 45 years of age. This is likely due to the presence of a series of very old females clustering in the later phase of occupation, a segment of the population that was clearly absent in the Early Classic sample.
Discussion
Paleodemographers have scrupulously emphasized the need for a set of basic conditions before engaging paleodemographic simulations through age-at-death distributions (Hoppa and Vaupel Reference Hoppa, Vaupel, Hoppa and Vaupel2002; Séguy and Buchet Reference Séguy and Buchet2013). One of these conditions is how well the skeletal collections represent the ancient living population. Second, the population or population segment occupying the cemetery has to be traceable back to the living population (Séguy and Buchet Reference Séguy and Buchet2013:14). Finally, reliable anthropological methods of age-at-death estimation are needed. The skeletal population from the marshland platform of Xcambó fulfills these paleodemographic conditions.
Life tables have received two main critiques from scholars: the underrepresentation of infant remains, which modifies survivorship values, and the use of a stationary scenario that is prone to introduce errors and biases into the elaboration of such tables (Civera-Cerecedo and Márquez-Morfín Reference Civera-Cerecedo, Márquez-Morfín, Márquez-Morfín and de León1998). In response to these critiques, Moore and colleagues (Reference Moore, Swedlund, Armelagos and Swedlund1975) pointed out that life expectancy and the probabilities of death do not alter mathematically and can therefore be used for valid interpretations. Johansson and Horowitz (Reference Johansson and Horowitz1986) have also demonstrated that, when the simulations are focused on large time spans, the deviations caused by an r that is equal to zero are minimal. As an additional guarantee, paleodemographers have compared scenarios with different growth rates to select the best “fit” (Hernández-Espinoza and Márquez-Morfín Reference Hernández-Espinoza and Márquez-Morfín2015; Séguy and Buchet Reference Séguy and Buchet2013).
Differential mortuary treatments are a principal concern among those Mayanists studying burial populations. Commoners and people of higher ranks were buried (and often reburied) in diverse manners not in cemeteries but underneath residential (and sometimes ceremonial) structures. In regional archaeology, this burial practice has hampered the homologous collective excavation and registry of such skeletal series (Andrews and Fash Reference Andrews and Fash1992; Fash Reference Fash, Leventhal and Kolata1982; Haviland Reference Haviland1972; Tourtellot et al. Reference Tourtellot, Sabloff, Carmean, Chase and Chase1992). At Xcambó, burial practices were more homogeneous, with individual primary deposits and simple cists prevailing (Sierra-Sosa Reference Sierra-Sosa2004). Lacking any distinguishable evidence of a political elite, Sierra-Sosa (Reference Sierra-Sosa2004) concluded that the population remained relatively homogeneous, although the wealth of this salt production center must have increased during the Late Classic. The intensive and extensive archaeological exploration of Xcambó's subsoil has been instrumental in the collective recovery of this burial population, permitting the chronological assignment of the great majority of its skeletons (Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014). Fortunately for this study, the geochemical conditions of its waterlogged subsoil led to acceptable preservation levels among all skeletal age groups and both sexes. We can therefore make a good case for the Xcambó's mortuary series being representative of its living population.
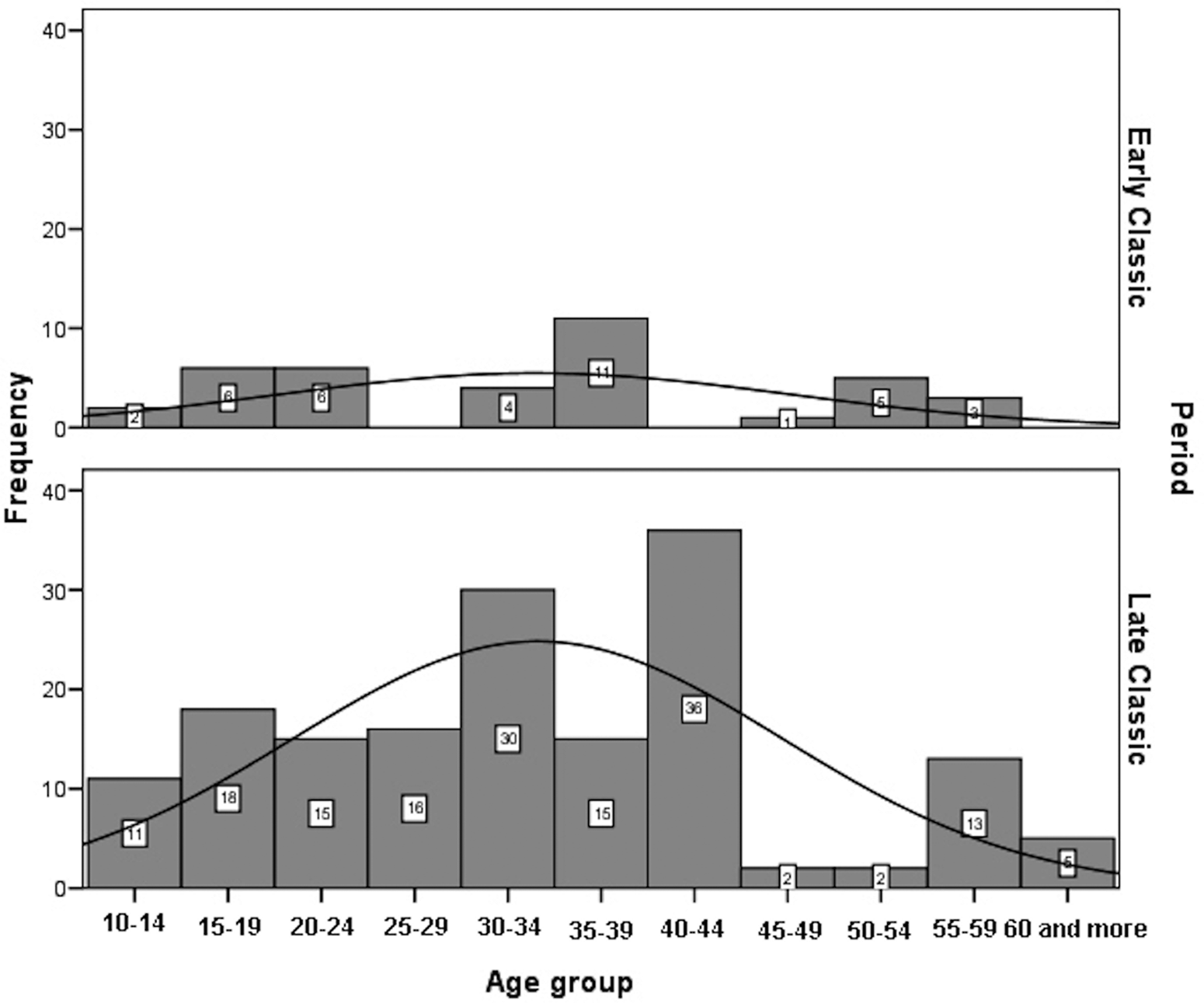
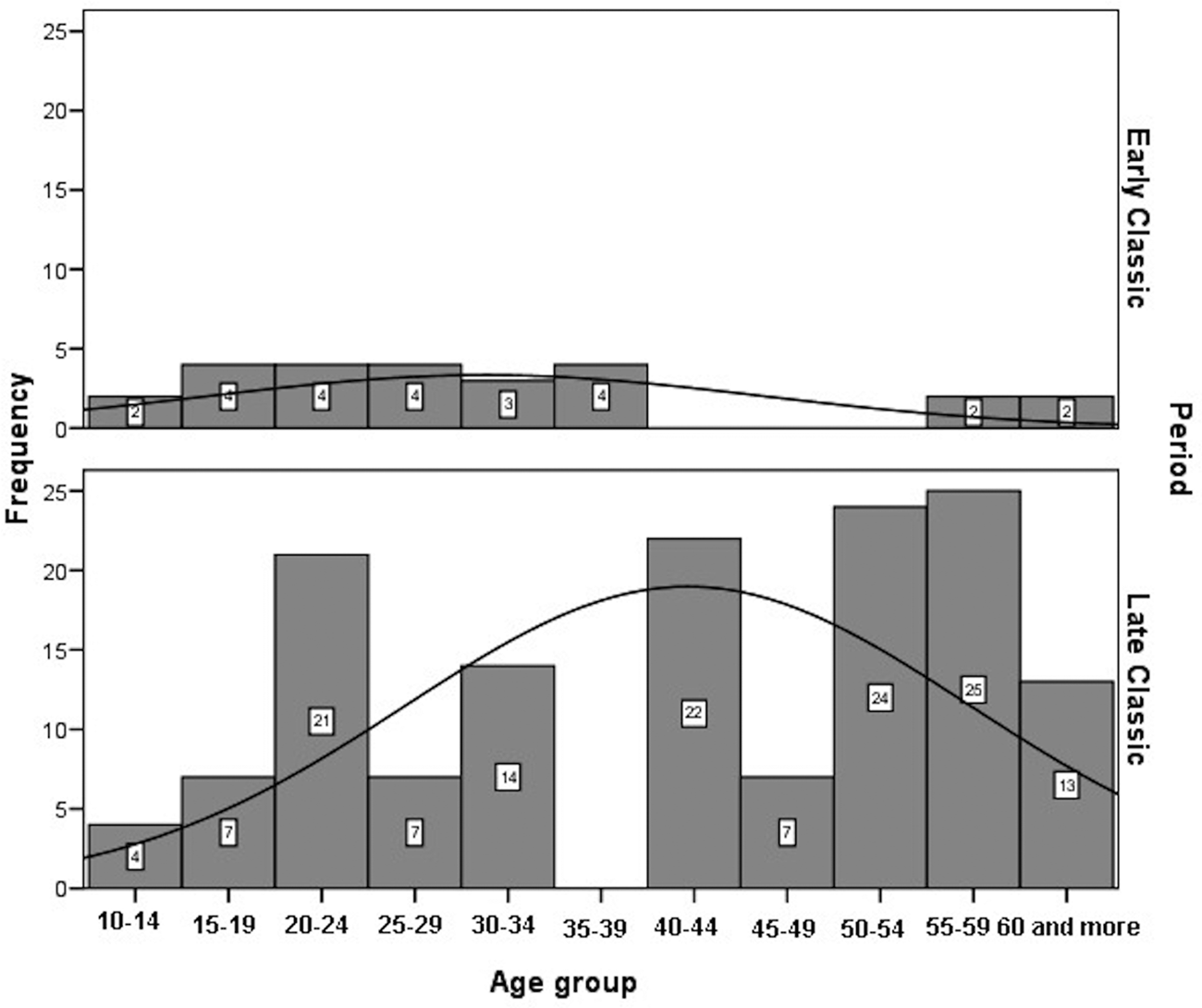
Age-at-death was estimated using a wide range of macroscopic and histomorphometric techniques (Suzuki et al. Reference Suzuki, Andrea Cucina, Sierra-Sosa, Laporte, Arroyo and Mejía2009). In addition, all the available auricular surfaces were seriated to improve the accuracy of age estimates (Tiesler et al. Reference Tiesler, Cucina, Sierra-Sosa and Meindl2004, Reference Tiesler, Cucina, Sierra-Sosa, Falla and Meindl2005). The age-at-death distributions for adults by sex, after having smoothed the original data, are significantly different between periods (males: χ2 = 38.676; df = 10; p < 0.00; females: χ2 = 43.331; df = 10; p < 0.00). Male deaths are more frequent between 15 and 44 years of age during the Late Classic, whereas no specific pattern can be highlighted for the Early Classic sample (Figure 6). Female deaths in Early Classic period are concentrated between 15 and 39, coinciding with females’ most fertile age. Female deaths in Late Classic show three modal peaks: at age 20–24 years, at 40–44 years (for reproductive ages), and 50–59 years (Figure 7).
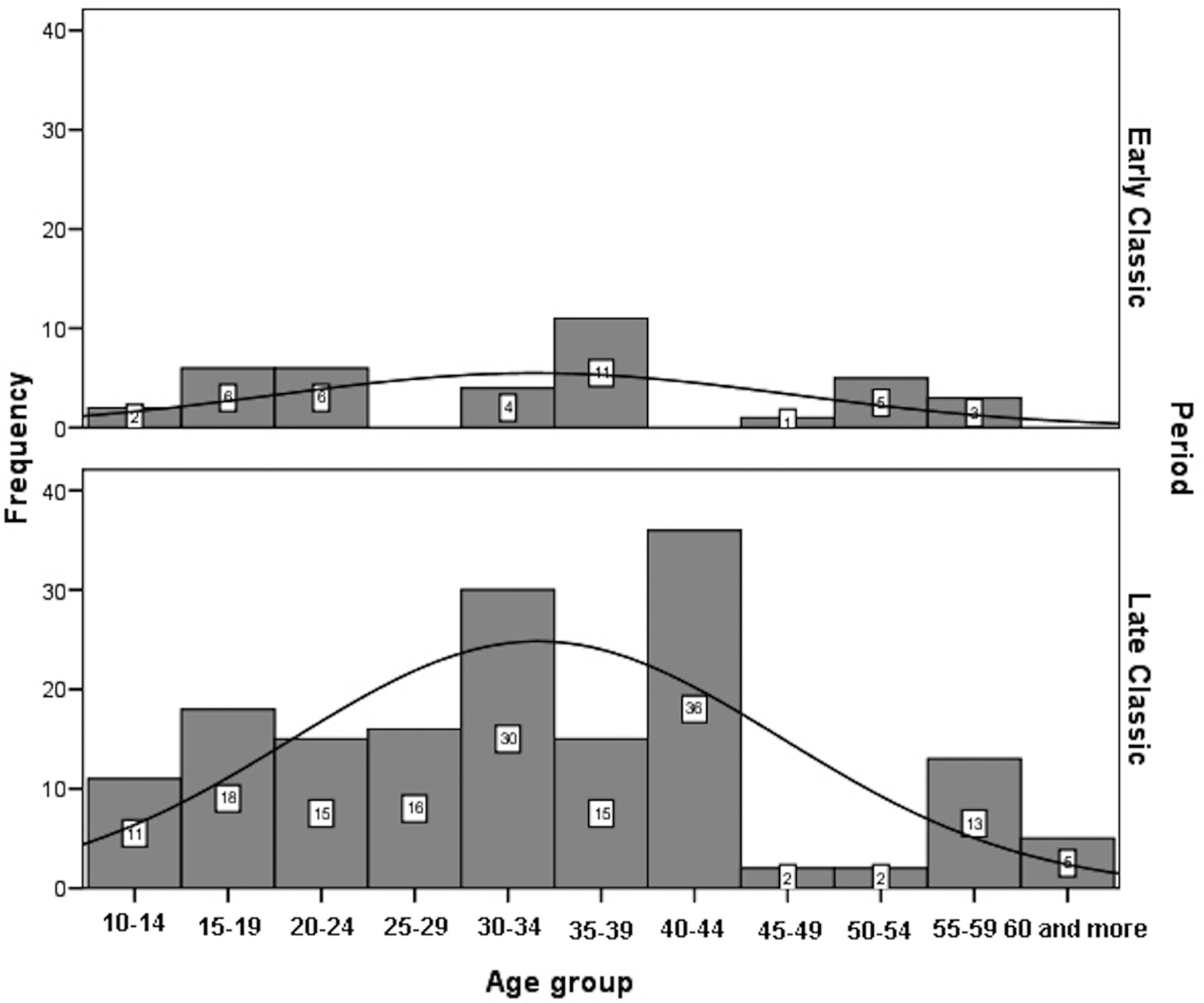
Figure 6. Histogram of distribution of adult males at Xcambó by chronological periods.
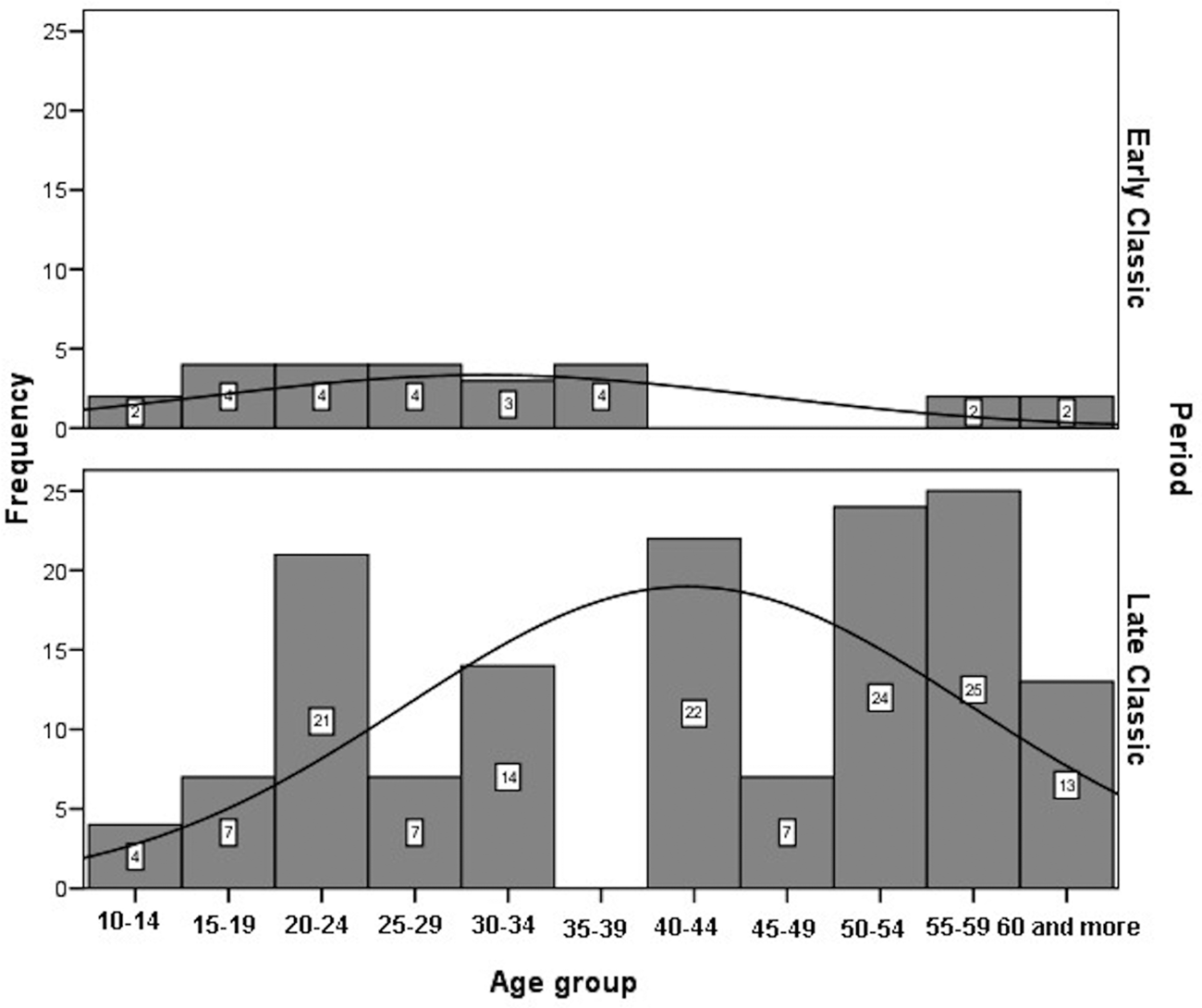
Figure 7. Histogram of distribution of adult females at Xcambó by chronological periods.
Brothwell (Reference Brothwell1987) pointed to the difficulties inherent in inferring infant mortality rates (IMR) in ancient populations because of the limitations imposed by taphonomic, environmental, and epidemiological factors. In validated skeletal populations, IMR has been considered a sensitive indicator of precarious living conditions, poverty, and a lack of healthy routines precisely because of infants’ higher vulnerability to disease and death (Bronfman Reference Bronfman1990; Eberstein Reference Eberstein1989; Gortmaker and Wise Reference Gortmaker and Wise1997; Juárez Reference Juárez1992; Márquez-Morfín and Sosa-Márquez Reference Márquez-Morfín and Sosa-Márquez2016; Schroeder and Brown Reference Schroeder and Brown1994; Tomkins Reference Tomkins2000). Factors such as a limited access to potable water (Gómez de León Reference Gómez de León, Bronfman and de León1988, Reference Gómez de León, Márquez-Morfín and de León1998; Mojarro and Nuñez Reference Mojarro and Núñez1988; Morelos Reference Morelos1996:423), infectious respiratory and gastrointestinal diseases, complications during pregnancy and childbirth, and maternal illness (Márquez-Morfín and Sosa-Márquez Reference Márquez-Morfín and Sosa-Márquez2016; Monterrubio-Gómez and González-Morales Reference Monterrubio-Goméz and González-Morales1995) increased mortality risk among infants. In addition, the age of the mother at childbirth can affect IMR. The curvilinear relation between the mother's age and IMR displays increased death rates toward the extremes of fertile life: younger than age 20 and older than 34 years (Bronfman Reference Bronfman1990:337).
Under a stable population scenario, Xcambó's IMR identifies values of 296.1 per thousand births and 282.4 per thousand births during the Early and the Late Classic cohorts, respectively (Table 2). These values are consistent with those of other Maya coastal groups that range between 233 and 333 at Chac Mool and Jaina, respectively, during the Late-Terminal Classic period (Hernández-Espinoza and Márquez-Morfín Reference Hernández-Espinoza and Márquez-Morfín2015). If we accept these figures, the IMR at Xcambó was relatively high, just as among other Mexican preindustrial populations, although it decreased gradually toward the end of its occupation (Camposortega-Cruz Reference Camposortega-Cruz1997; Oliver Reference Oliver, Muriá and Olveda1992; Ortega-Muñoz Reference Ortega-Muñoz2004, Reference Ortega-Muñoz2012:175; Velasco Reference Velasco and del Pilar1992).
Rebecca Storey (Reference Storey, Whittington and Reed1997) documented infants from elite residential contexts at urban Copán and interpreted their high morbidity and mortality as evidence of a lack of buffering systems against unhealthy environments in the city's cramped central living quarters. Sierra-Sosa (Reference Sierra-Sosa2004) and Sierra-Sosa and coauthors (Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014) argued that the population of Xcambó was composed of homogeneously wealthy settlers but without the upper echelons of society, because no separate acropolis or any royal-like tomb was unearthed at this site. Given the Xcambó placement—in the midst of swampy marshland close to the coast— Xcambó's infants may have experienced similar difficulty in buffering negative environmental impacts, similar to those described at urban Copán. Detailed analyses of linear enamel hypoplasia (Cucina Reference Cucina2011) indicate that all of Xcambó's population had undergone intense physiological stress episodes during childhood. On average, four to five horizontal hypoplastic lines per individual were recorded; individuals with central values (from three to eight defects per person) could be assigned to both subadults and adults. None of the recorded subadults manifested less than two defects, and subadults exhibited the highest numbers of defects per individual recorded. This stands in contrast to those individuals who did survive to maturity (Cucina Reference Cucina2011).
We suggest that the decreasing IMR was not related to the improved social conditions that may have granted better health, because diet did not appear to have shifted notably through time (Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014), nor did we note any major changes in general pathological conditions between time periods. Therefore, the reason for that decrease must be sought in other factors. The adult populations’ overall parameters of fertility may provide one explanation, because IMR is a direct consequence of fertility (Sattenspiel and Harpending Reference Sattenspiel and Harpending1983).
On a global scale, human fertility increased with the onset of agriculture and was coupled with a shorter life span among women, which was likely triggered by maternal mortality (Eshed et al. Reference Eshed, Avi Gopher, Gage and Hershkovitz2004). Our results show high fertility levels at Early Classic Xcambó, with a considerable death toll among females during their reproductive ages. Their mortality is concentrated between the 15–19 and 20–29 age groups at age-at-death, a trend that is consistent with the higher mortality risks associated with childbirth, just as Boldsen and Paine (Reference Boldsen, Paine, Jeune and Vaupel1995) and Eshed and coauthors (Reference Eshed, Avi Gopher, Gage and Hershkovitz2004) reported during the transition to agriculture. By contrast, our results suggest a decrease in fertility during the Late Classic, when female peaks of mortality, within fertile ages, occurred in the 20–24 to 40–44 years age groups (another noticeable peak occurred in the postmenopausal class of 50–59 years of age; Figure 7).
The patterns of IMR and fertility, associated with the women's mortality profiles, are indeed indicative of changes in the patterns of childbirth: earlier pregnancies and more births in the Early Classic shifted to later pregnancies and, of course, fewer births in the following period. Fertility could be manipulated intentionally or unintentionally during daily routines. Gurri and Ortega-Muñoz (Reference Gurri and Ortega-Muñoz2015) reported that, in agricultural populations that share the same environment, increased female workloads, inadequate access to food staples, and prolonged breastfeeding could all result in reduced fertility levels among women (Cohen Reference Cohen1989; Lee Reference Lee, Cohen, Malpass and Klein1980; Watkins and Van de Walle Reference Watkins, de Walle, Rotberg and Rabb1990). In such a scenario, however, and contrary to what we may expect, the skeletal occupational indicators at Xcambó did not show accountable differences in women's daily activity levels between the Early and the Late Classic periods (Maggiano et al. Reference Maggiano, Schultz, Kierdof, Sierra-Sosa, Maggiano and Tiesler2008), nor did their diet appear to have changed (Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014): females accessed “exogenous” foodstuff during the Late Classic just like their male counterparts (Cucina, Perera, et al. Reference Cucina, Perera, Sierra-Sosa and Tiesler2011). It seems unlikely that either unfavorable living conditions or culturally induced practices (infanticide or abortion) affected fertility during the later phase of occupation, because we have no skeletal evidence to indicate either factor.
Infanticide usually is tied more to baby girls than to baby boys and therefore leads to an unbalanced sex ratio with a male dominance. However, in contrast to other Maya settlement populations in which males appear to predominate systematically over females (see, for example, Krejci and Culbert Reference Krejci, Patrick Culbert and Grube1995; Tiesler Reference Tiesler1999; Welsh Reference Welsh1988), the sexes are fairly balanced among Xcambó's adult population segment; this argues against the possibility of infanticide. Furthermore, the presence of neonates at the site and the location of the burials in the residential compounds discount the possibility that reduced fertility might be attributable to the archaeological excavation not having recovered this demographic fraction. Many perinatal and neonates were unearthed from garbage mounds at the site; although we cannot rule out the possibility that they were the product of infanticide, they might well indicate a different burial practice from that reserved for the older mortals of the local population. Moreover, even in the case that they represent infanticide, their presence in the skeletal collection would imply that we do not need to compensate our life table for this eventuality.
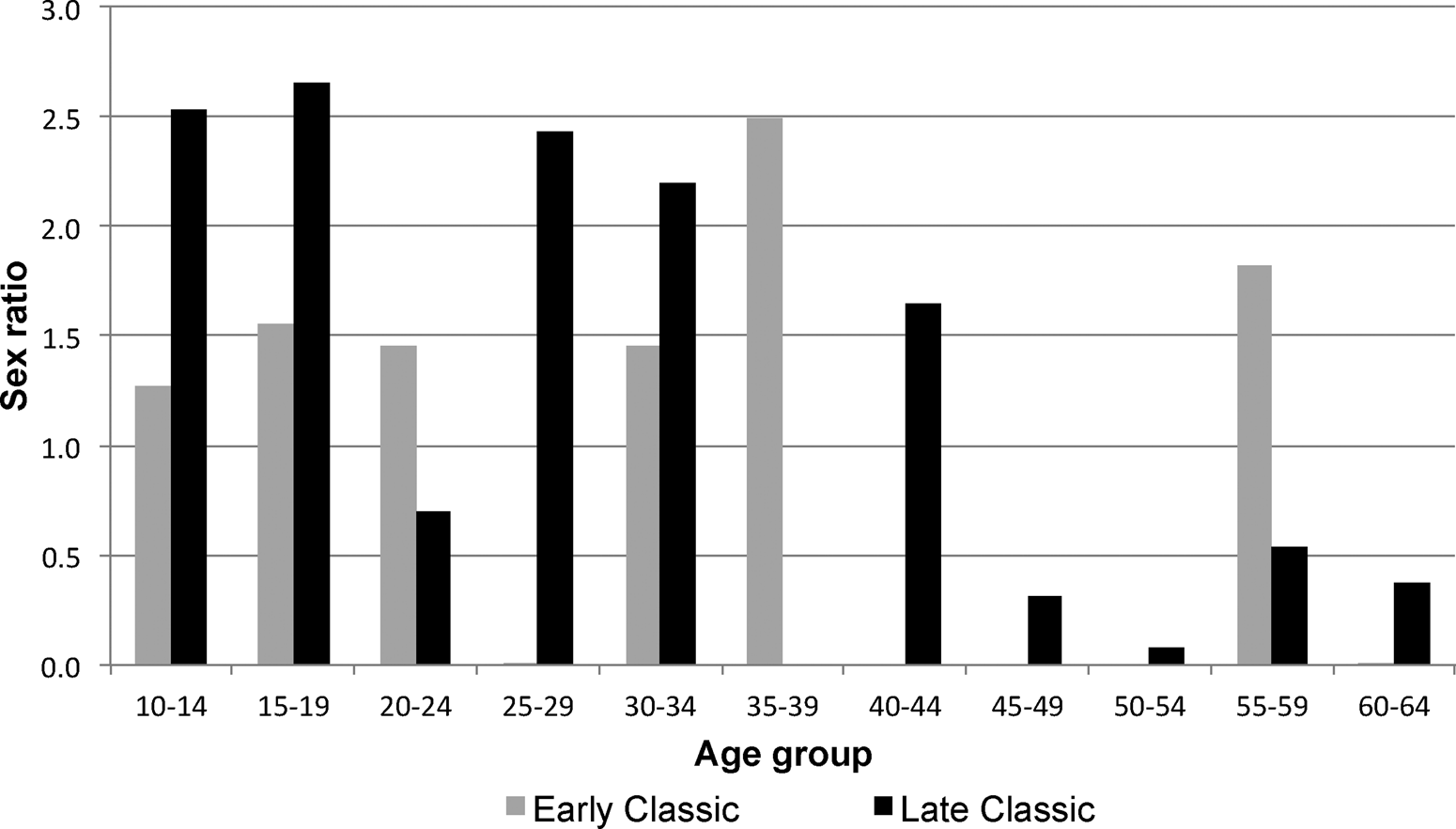
Despite an overall balanced sex ratio, however, demographic profiles among males do differ from those of females. Both Early and Late Classic profiles show a rather regular normal distribution, with modals between 30 to 44 years (kurtosis = −1.053 for Early Classic; −0.511 for Late Classic). Like females, Late Classic males’ age-at-death peaks at 50–59 years of age, although the mean age-at-death between periods increased only in females. What is striking is the high sex ratio favoring males, in particular between ages 10 and 34 during the later phase of occupation (Figure 8). Eshed and coauthors (Reference Eshed, Avi Gopher, Gage and Hershkovitz2004) suggest that this sex bias may not mirror a real situation, with taphonomic, mortuary practices, or methodological problems in sex determination generating the bias in the skeletal record, or that bias is the product of selected infanticide (against baby girls). Although some potential methodological issues may create some error in sexing skeletons, we can rule out taphonomic or mortuary practices, because large segments of the site's subsoil were excavated, yielding no evidence for differences in the way males and females were interred (Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014).

Figure 8. Sex ratio at Xcambó by chronological periods.
Considered jointly, all the archaeological, skeletal, and demographic forms of evidence indicate that the socioeconomic changes and the site's predominant role as a relatively autonomous salt production port and administrative center within short- and long-distance trade routes triggered noticeable shifts from the Early to the Late Classic. Although Xcambó cannot be considered an “urbe” sensu stricto (to the same extent as large and important centers like Teotihuacan), Cucina and coauthors (Reference Cucina, Sierra-Sosa, Tiesler, Charlotte Arnauld, Pereira and Beekman2021), based on a series of skeletal and dental indicators, suggest that Xcambó's social and contextual change mirrored the process of urbanization. The results from the demographic simulations performed in the present study are consistent with this hypothesis and provide further indicators (like IMR, and TFR) that Tiesler and colleagues (Reference Tiesler, Cucina, Sierra-Sosa, Falla and Meindl2005) and Cucina and colleagues (Reference Cucina, Sierra-Sosa, Tiesler, Charlotte Arnauld, Pereira and Beekman2021) do not take into account.
Although the rural/urban threshold is hard to define, Champion and Hugo (Reference Champion, Hugo, Champion and Hugo2004:8, Table 1.2) listed a series of indicators that distinguish an urban context from a rural one. In urban contexts, fertility decreases in parallel with a decrease in infant mortality, a pattern that is evident at Xcambó (although IMR did not drop to the level found in modern urban contexts). Subsistence in urban environments is based on (among other factors) administrative tasks, in contrast with more agriculturally oriented roles in rural areas. At Xcambó, the new administrative activities performed by the male segment of the population clearly contrast with the more intense physical activities (Wanner et al. Reference Wanner, Sierra-Sosa, Alt and Tiesler2007; Maggiano et al. Reference Maggiano, Schultz, Kierdof, Sierra-Sosa, Maggiano and Tiesler2008) previously performed by their Early Classic peers. The population of Xcambó seemed to be more diverse in the later phase of occupation (Cucina et al. Reference Cucina, Sierra-Sosa, Tiesler, Charlotte Arnauld, Pereira and Beekman2021; Tiesler Reference Tiesler2014), which echoes Champion and Hugo's (Reference Champion, Hugo, Champion and Hugo2004) claim that ethnicity is more varied in urban areas and more homogeneous in rural ones.
Finally, immigration levels are higher in urban contexts, whereas net outmigration prevails in rural areas (Champion and Hugo Reference Champion, Hugo, Champion and Hugo2004). This last factor provides valuable hints as to why, despite the decrease of fertility rates during the Late Classic as indicated by the demographic indicators obtained from both methodological approaches, the archaeological and skeletal evidence shows increases in growth rates (Ortega-Muñoz et al. Reference Ortega-Muñoz, Andrea Cucina and Sierra-Sosa2018; Sierra-Sosa Reference Sierra-Sosa2004). Sierra-Sosa and coauthors (Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014; see also Cucina, Tiesler, et al. Reference Cucina, Tiesler, Sierra-Sosa and Neff2011; Cucina et al. Reference Cucina, Sierra-Sosa, Tiesler, Charlotte Arnauld, Pereira and Beekman2021) have provided isotopic evidence of considerable foreign immigration at the site only during the Late Classic. Of the 131 individuals analyzed for strontium isotopes, 20 presented a 87Sr/86Sr ratio below the local signature (Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014). The more detailed analysis of Cucina and coauthors (Reference Cucina, Sierra-Sosa, Tiesler, Charlotte Arnauld, Pereira and Beekman2021) indicated that only one person (4%) belonged to the Early Classic period, whereas 19 (18.1%) could be assigned to the Late Classic. That Early Classic foreigner came from an area adjoining the peninsula's northern coast (where Xcambó is located), whereas the Late Classic foreigners likely came from places as far south as the Petén or the Usumacinta River basin (Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014). Among the Late Classic immigrants are one juvenile and two subadults younger than 10 years of age. The remainder are all adults, distributed roughly equally between the sexes. The presence of infants, although rare, suggests that not only single adult individuals but also entire families with their infant offspring may have moved into the site. This conclusion strengthens inferences of prompt assimilation, drawn from the differences between the head shapes of infants and adults of Xcambó's Late Classic settlers (Tiesler Reference Tiesler and Cucina2015). Although in- and out-migration may have effects on a population's demographic stationarity, they do not affect the interpretation of paleodemographic parameters based on living conditions, which are indicative mainly of mortality and fertility. Nonetheless, a more thorough discussion on the extent and effects of immigration will be possible only when a larger 87Sr/86Sr isotope database is available, which will be the topic of a future article.
Regarding any social shifts and their possible impact on the age-at-death distribution patterns, we did not notice real changes in the Sx and μx during the first 15 years of age, in the IMR, or in the 75% quartile of survivorship between chronological periods. Instead, changes are more noticeable in the juvenile and middle-adult segments of both populations. The data in Figure 4 indicate that the mortality in the 10–40 years age segment is slightly lower in the Late Classic, a phenomenon that has been reported in other studies, where mortality in middle age decreases more rapidly than in immature and senescent ages (Eshed et al. Reference Eshed, Avi Gopher, Gage and Hershkovitz2004; Gage Reference Gage1993: 53–54; Meslé et al. Reference Meslé, Vallin and Andreyev2002; Vallin Reference Vallin, Schofield, Reher and Bideau1991), with the obvious consequence of an increase of life expectancy across the time.
Finally, we may only speculate why the settlement was abandoned at the end of the Classic. However, based on our paleodemographic re-creation, the reasons should relate to larger geopolitical, social or economic factors, rather than local living conditions, as we argued in earlier work (Sierra-Sosa et al. Reference Sierra-Sosa, Cucina, Douglas Price, Burton and Tiesler2014).
Conclusions
So far, few paleodemograhic studies have been applied successfully to the reconstruction of prehispanic Maya populations, and fewer still compare life expectancy at birth between different time periods. These types of efforts have been hampered by poor preservation, reduced skeletal numbers, low representativeness caused by irregular burial placement, and differentiated burial practices. In this study we conducted one of the few feasible paleodemographic studies in the Maya area.
The skeletal series of the Classic period of Xcambó, Yucatan, is not only one of the region's largest skeletal collections available for bioarchaeological analyses but also is one of the most representative of the ancient living population. Its reconstructed demographic profiles, considered jointly with other lifestyle indicators, biodistance data, and the archaeological record, hold promise for analogous future investigations of Classic Maya population structure and social dynamics. They provide fresh answers to questions concerning internal growth mechanisms, living conditions, and social status and contribute to the paleodemographic discussion in the Maya area that suggests a growing population in coastal sites of the peninsula of Yucatan from Classic to Postclassic times, with life expectancy at birth ranging between 15 and 42 years (Cohen et al. Reference Cohen, O'Connor, Danforth, Jacob, Armstrong, Whittington and Reed1997; Hernández-Espinoza and Márquez-Morfín Reference Hernández-Espinoza and Márquez-Morfín2015).
Every scenario, however, must be explained within its own social and ecological context to thoroughly understand the relationship between each society's lifestyle, fertility, illness, and mortality. The pursuit of several academic lines of inquiry and comparisons of different datasets have proven crucial in this endeavor, thus enhancing our knowledge about not only the biosocial continuum of Xcambó's trajectory but also ancient Maya coastal populations in general. These interdisciplinary efforts necessarily presume close communication across disciplines and collaboration between scholars.
Acknowledgements
The skeletal material was studied as part of the project Bioarqueología en Área Maya at the Facultad de Ciencias Antropológicas (UADY) and was excavated by the Proyecto Arqueológico Xcambó (Thelma Sierra-Sosa, INAH Center Yucatán, Mexico). This research stemmed from a postdoctoral research visit by the senior author at the bioarchaeological facilities of UADY and has been funded by CONACYT Grant H-150521. We are grateful to Richard Meindl (Kent State University) for generously sharing ideas and debates on ancient paleodemographical caveats in past years and to Hiram Beltran-Sánchez for his recommendations about demographic phenomena in past societies.
Data Availability Statement
The data presented in this article are available at the Laboratorio de Bioarqueología e Histomorfología of the Facultad de Ciencias Antropológicas, Universidad Autónoma de Yucatán, Mérida, Yucatán, México.