



1. Introduction
The Viséan stage was defined in the Dinant Synclinorium, close to the town of Visé. The first subdivision of the Viséan into lower, middle and upper parts, with regional names, was also developed in the same area from the end of the nineteen century and refined in the early twentieth century. These subdivisions were associated with the chronozones V1, V2 and V3 (and subdivided into subchronozones), initially defined on the basis of macrofossils such as brachiopods and rugose corals; goniatites were later added (see historical revision in Demanet, Reference Demanet1958). It was not until the 1960s when Conil & Lys (Reference Conil and Lys1964) and Conil et al. (Reference Conil, Pirlet and Lys1967) used microfossils (foraminifers, conodonts and rare calcareous algae) to more precisely define each chronozone. Within this zonal scheme, V1 was equated to the lower Viséan, V2 to the middle Viséan and V3 to the upper Viséan substages. Refinements of the zonal schemes, and overall increase in the knowledge of the fauna and lithologies in Belgium and around the world, led to some successive modifications to this zonal scheme. The most important change was introduced by Conil et al. (Reference Conil, Groessens and Pirlet1977), who proposed new names for the substages of the Viséan, namely, Moliniacian, Livian and Warnantian, and their associated ‘Cf’ foraminiferal zones, replacing the previous names (Assise de Dinant, Assise de Namèche and Assise de Warnant) and previous chronozones and subchronozones (those of the V1a to V3c superieur). This process was not only a simple replacement of names, but also involved the definition of new stratotypes and the amended foraminiferal zones compared with worldwide stratigraphical scales. Related to the lower and middle Viséan stages (Moliniacian and Livian substages), they correlated the base of the Moliniacian Substage with the base of the Chadian Substage in England (base of their Cf4α = base of the V1a subzone), and considered it as the base of the Viséan Stage. On the other hand, the base of the Livian Substage was correlated with the base of the Holkerian Substage in England (base of Cf5 zone = base of V2b subzone), moving the base of the older middle Viséan substage (or Assise de Namèche) from the base of the V2a to the base of the V2b. Later, Conil et al. (Reference Conil, Groessens, Laloux and Poty1989) proposed that the Tournaisian–Viséan boundary should be situated at the base of the subzone Cf4α2. All these modifications were summarized in the zonal scheme published by Conil et al. (Reference Conil, Groessens, Laloux, Poty and Tourneur1991). A final emendation of the base of the Moliniacian Substage was introduced by Devuyst (in Poty et al. Reference Poty, Devuyst and Hance2006) to equate the base of this substage with the base of the Viséan Stage.
More recent studies have recorded ambiguous foraminiferal assemblages around the world that could be assigned to the middle and lower Viséan Stage, generally considered as the middle Viséan substage (e.g. Vachard et al. Reference Vachard, Termier and Termier1977; Vieslet, Reference Vieslet1983; Vachard & Tahiri, Reference Vachard and Tahiri1991; Fassihi et al. Reference Fassihi, Kulagina, Esfahani and Nurgalilev2018). In the past two decades, some authors have proposed the possible existence of an intermediate foraminiferal assemblage in some countries around the world situated between the lower and middle Viséan Belgian substages (Izart et al. Reference Izart, Tahiri, El Boursoumi, Vachard, Saidi, Chèvremont and Berkhli2001; Devuyst et al. Reference Devuyst, Hance, Hou, Wu, Tian, Coen and Sevastopulo2003; Hance et al. Reference Hance, Hou and Vachard2011; Okuyucu et al. Reference Okuyucu, Vachard and Göncüoğlu2013; Zandkarimi et al. Reference Zandkarimi, Najafian, Vachard, Bahrammanesh and Vaziri2016; Fassihi et al. Reference Fassihi, Kulagina, Esfahani and Nurgalilev2018). However, its occurrence and calibration with the classical outcrops in western Europe has been never undertaken; it is therefore not clear if they have to be considered as regional subdivisions or if there is a biozone with worldwide implication.
The aim of this study is to investigate the possible presence (or absence) of a foraminiferal biozone (or subzone) intermediate in position between the formally recognized foraminiferal biozones (Cf4/Cf5 and MFZ11/MFZ12) in the western Palaeotethys, and validate its potential synchronous dispersal by comparison to basins situated in the eastern Palaeotethys.
2. Lower–middle Viséan boundary interval in western Palaeotethys
This study is limited to the stratotype for the base of the Livian Substage in Belgium, as well as the stratotype for the Holkerian Substage in Cumbria (northern England), other sections in the Askrigg Block, which include a more continuous succession down to the lower Viséan substage, and sections in southern France (Montagne Noire) and the western Meseta of Morocco. Other basins or countries in the western Palaeotethys do not contain good transitions between the lower and middle Viséan carbonates, and they are commonly associated with siliciclastics.
2.a. Foraminifers at the base of the Livian Substage in Belgium
The stratotype for the Livian base was selected in the transition between the carbonates of the Neffe and Lives formations (Conil et al. Reference Conil, Groessens and Pirlet1977), although an argillaceous bentonite of volcanosedimentary origin is located at the boundary, regionally named as the Banc d’Or de Bachant (Fig. 1), considered as the transition between the Cf4 and Cf5 chronozones. More recent studies (Hance et al. Reference Hance, Poty and Devuyst2006; Poty et al. Reference Poty, Devuyst and Hance2006, Reference Poty, Aretz and Hance2014; Poty, Reference Poty2016) have followed the lithostratigraphical subdivision proposed by Conil et al. (Reference Conil, Groessens and Pirlet1977), and new Mississippian foraminiferal zones were defined (MFZ) to standardize more widely distributed foraminiferal guides (Poty et al. Reference Poty, Devuyst and Hance2006). For these authors, some of the guide foraminifers defined for the subchronozones in Conil et al. (Reference Conil, Longerstaey and Ramsbottom1980, Reference Conil, Groessens, Laloux, Poty and Tourneur1991) had too much local character, and could not be identified outside of their influence areas in Belgium.
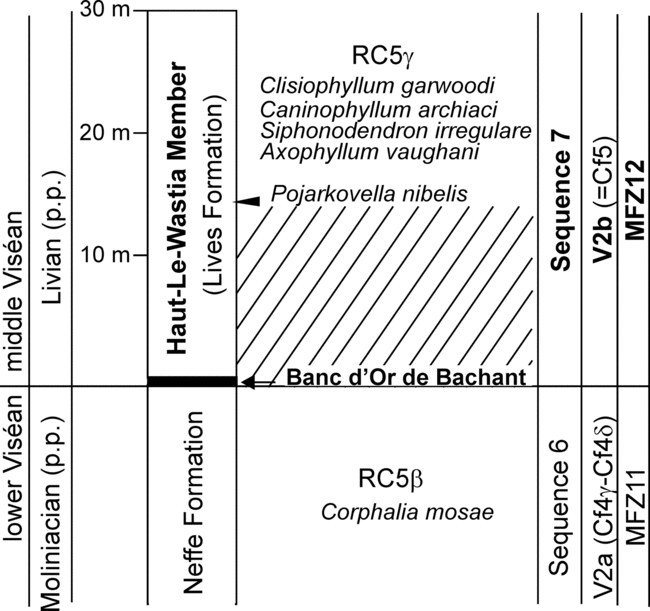
Fig. 1. Biostratigraphic sketch of the Moliniacian–Livian boundary interval stratotype at Lives section in Belgium. Data source: Poty & Hance (2006). Diagonal striped lines have no foraminiferal record because of unfavourable facies or a gap.
Since the studies of Conil et al. (Reference Conil, Pirlet and Lys1967, Reference Conil, Groessens and Pirlet1977), the guides for the base of the Livian, V2b or Cf5 zones were considered to be Pojarkovella nibelis (Durkina, Reference Durkina1959) and Koskinotextularia (named as Cribrostomum in 1967 or cribrate palaeotextulariids in 1977). Later, Archaediscus at concavus stage was also recognized as an auxiliary marker (Conil et al. Reference Conil, Longerstaey and Ramsbottom1980, Reference Conil, Groessens, Laloux, Poty and Tourneur1991), as well as more questionable or regional taxa such as primitive ‘Millerella’, Rhodesinella, Lituotubella, Nodosarchaediscus, Omphalotis minima (Rauzer-Chernousova & Reitlinger in Rauzer-Chernousova et al. Reference Rauzer-Chernousova, Belyaev and Reitlinger1936), Consobrinellopsis consobrina (Lipina, Reference Lipina1948) and Endothyranopsis (e.g. Conil, Reference Conil, Wagner and Holub1977; Conil et al. Reference Conil, Longerstaey and Ramsbottom1980, Reference Conil, Groessens, Laloux, Poty and Tourneur1991; Poty et al. Reference Poty, Devuyst and Hance2006). The evolutionary stages of the genus Archaediscus involving the shape of the sutures between the whorls, involutus, concavus, angulatus and tenuis were defined by Pirlet & Conil (Reference Pirlet and Conil1974) and later modified by Conil et al. (Reference Conil, Longerstaey and Ramsbottom1980) and Vachard (Reference Vachard1988a). From a taxonomical point of view they have no species value, but the presence of this character in a group of species of this genus has been used as a biostratigraphic tool in numerous studies in Europe (Conil et al. Reference Conil, Groessens and Pirlet1977, Reference Conil, Longerstaey and Ramsbottom1980; A.R.E. Strank, unpub. PhD thesis, University of Manchester,1981; Somerville, Reference Somerville2008) and in Asia (e.g. Hance et al. Reference Hance, Hou and Vachard2011; Zandkarimi et al. Reference Zandkarimi, Vachard, Najafian, Hamdi and Mosaddegh2017, Reference Zandkarimi, Vachard, Najafian, Mosaddegh and Ehteshami-Moinabadi2018). Another important character within the Archaediscus is the progressive reduction of the microgranular layer during its evolution, which led to Brenckle et al. (Reference Brenckle, Ramsbottom and Marchant1987) to consider the distinction between the genus Paraarchaediscus Orlova, Reference Orlova1955 (for those forms with well-developed microgranular layer) from Archaediscus Brady, Reference Brady1873 (for those forms with reduced microgranular layer) valid. This is a continuum process where the end-members are easily distinguished, whereas there are a multitude of intermediate forms in which it is difficult to decide if they belong to one or the other genus due to the ambiguity in the boundary between both genera. All the species recorded in the lower part of the middle Viséan substage would correspond to the genus Paraarchaediscus but, as a result of uncertainty and questionable taxonomic value of the genus Paraarchaediscus, ‘Archaediscus’ is used here. However, when data are based on previous studies by other authors, we have preserved the original notation as Archaediscus or Paraarchaediscus. The first occurrence of Pojarkovella nibelis is some distance above the base of the Lives Formation (at a level 14.3 m above the base, in the 30-m-thick basal Haut-le-Wastia Member in the Livian stratotype section; Fig. 1); in other regions of Belgium, this species first occurs much higher compared with the base of this substage (Poty & Hance, Reference Poty and Hance2006; Poty et al. Reference Poty, Aretz and Hance2014). Koskinotextularia occurs later than P. nibelis (Poty et al. Reference Poty, Devuyst and Hance2006). These authors also defined the rugose coral assemblage (RC5γ) for the Haut-Le-Wastian Member, characterized by the first occurrence of Siphonodendron irregular (Phillips, 1936), Axophyllum vaughani (Salée, Reference Salée1913), Caninophyllum archiaci (Milne-Edwards & Haime, Reference Milne-Edwards and Haime1852) and Clisiophyllum garwoodi (Salée, Reference Salée1913) (Fig. 1). Unfortunately, the first occurrence of this assemblage was situated ‘in the first open-marine bioclastic limestone’ and not from the base of the unit, which is mostly composed of massive limestones, intraclastic limestones or evaporitic limestones with stromatolites (Hance, Reference Hance1979).
Some of the previously described auxiliary foraminiferal guides for the Livian Substage are now confidently recognized as first occurring in the lower Viséan (Moliniacian) substage, such as Nodosarchaediscus, Omphalotis minima and Consobrinellopsis consobrina; in the most recent revision, they are considered as abundant species in the Livian assemblages (Poty et al. Reference Poty, Devuyst and Hance2006).
The Banc d’Or de Bachant is a volcanosedimentary bentonite, 10–30 cm thick with local pedogenic features, and is recognized as representing a gap in limestone deposition (erosion and/or hiatus) (Poty & Hance, Reference Poty and Hance2006). This Banc d’Or de Bachant is recognized in all the regions in Belgium and northern France, and is considered as the base of the sequence 7 (Hance et al. Reference Hance, Poty and Devuyst2001). Other authors have correlated this gap of the Banc d’Or de Bachant with similar rocks in Germany and England (A. Huyghe-Buissart, unpub. PhD thesis, University of Lille, 1993), although the stratigraphic position for this gap in England, at least, does not seem to correspond exactly to the position in Belgium. The presence of this hiatus has been criticized by some authors (e.g. Kalvoda, Reference Kalvoda2002; Hance et al. Reference Hance, Poty and Devuyst2006; Poty, Reference Poty2016), who have recommended the search for a new stratotype where there is a record of continuous sedimentation. Taking into consideration the studies in Britain (Conil et al. Reference Conil, Longerstaey and Ramsbottom1980), the Belgian subdivision can also be applied there. The hiatus between the Cf4 and Cf5 zones has been traditionally considered as negligible and, consequently, the possible influence and/or implication of the Banc d’Or de Bachant overlooked. Above the bentonite horizon, the Haut-le-Wastia Member is composed predominantly of lime mudstones, with stromatolites and local breccias due to evaporite dissolution. The lower part of the Lives Formation and the Banc d’Or are therefore not the most favourable facies for the recovery of foraminifers (Fig. 1). There is a 30-m-thick thick interval where the knowledge of typical lower and middle Viséan microfossils is poorly known, and even the positioning of the base of the middle Viséan substage at the base of the Banc d’Or de Bachant is not supported by any fossil guide.
2.b. The Arundian–Holkerian boundary interval in northern England
The stratotype for the base of the Holkerian Substage is situated at Barker Scar near Holker Hall in south Cumbria, northern England. The boundary was located at the contact between the Dalton Beds and the overlying Park Limestone, as originally defined by George et al. (Reference George, Johnson, Mitchell, Prentice, Ramsbottom, Sevastopulo and Wilson1976). The base of the Holkerian Substage was based on macrofossils, such as brachiopods (Davidsonina carbonaria (McCoy, Reference McCoy1852), Composita ficoidea (Vaughan, Reference Vaughan1903) and Linoprotonia corrugatohemispherica (Vaughan, Reference Vaughan1903)) and rugose corals (Axophyllum vaughani and Lithostrotion minus (McCoy, Reference McCoy1849)), as well as microfossils, the foraminifers Archaediscus at concavus stage and double-walled palaeotextulariids. However, in Barker Scar only Linoprotonia corrugatohemispherica occurs at the base of the Park Limestone (Rose & Dunham, Reference Rose and Dunham1977). These authors also considered Axophyllum cf. vaughani as typical of the lower beds in the Park Limestone, whereas they considered that Davidsonina carbonaria, absent from Barker Scar, occurs at equivalent levels elsewhere in England. In contrast, Composita ficoidea is commonly recorded in the upper part of the Dalton Beds (Fig. 2; beds e and i in Rose & Dunham, Reference Rose and Dunham1977). The double-walled palaeotextulariids is readily a misidentification, because they have never been recorded below the upper Viséan strata anywhere within the Palaeotethys; most probably, they were referring to the single-layered palaeotextulariids.

Fig. 2. Biostratigraphic sketch of the Arundian–Holkerian boundary interval stratotype at Barker Scar (Cumbria) and lower part of the Silverdale Borehole (Askrigg) in northern England. Bed thickness of the Barker Scar section according to Rose & Dunham (Reference Rose and Dunham1977) and Silverdale Borehole sensu Waters et al. (Reference Waters, Cózar, Somerville, Haslam, Millward and Woods2017). Base of the Park Formation according to Johnson et al. (Reference Johnson, Soper and Burgess2001). Diagonal striped lines have no foraminifers because of gaps.
Ramsbottom (Reference Ramsbottom1981) considered the same macrofossils as markers for the base of the Holkerian Substage and specified the foraminiferal list of potential markers. He considered the main guides as Koskinotextularia and Pojarkovella nibelis, but also mentioned Archaediscus at concavus stage and transitional forms to the angulatus stage, as well as the regional markers Holkeria and Dainella holkeriana Conil & Longerstaey in Conil et al. (Reference Conil, Longerstaey and Ramsbottom1980).
However, the conclusions in A.R.E. Strank (unpub. PhD thesis, University of Manchester, 1981), who also contributed in Ramsbottom (Reference Ramsbottom1981), were not exactly the same. She considered that the Archaediscus at concavus stage are not well-developed in England as in other basins, and that these forms appeared inconsistently from the Arundian to the Asbian substages. In her study, it is possible to find Omphalotis minima and Lituotubella from the base of the Barker Scar section (> 29 m below the boundary between the Dalton Beds and Park Limestone) and Archaediscus at concavus stage (A. varsanofievae Grozdilova & Lebedeva, Reference Grozdilova and Lebedeva1954 and A. stilus amplus Conil & Lys, Reference Conil and Lys1964) nearly from the base of the section (> 23 m below the boundary), both from bed a (Fig. 2). On the other hand, she recorded Consobrinellopsis (as Palaeotextularia sp.) in bed k (2.5 m above the boundary), and also Koskinotextularia and Pojarkovella nibelis in bed k (3.5 m above the boundary; Fig. 2). It can therefore be concluded that Omphalotis minima, Lituotubella and ‘Archaediscus’ at concavus stage first occur much earlier than Koskinotextularia and P. nibelis. Unfortunately, the Barker Scar section does not extend down far enough into the Arundian Substage as to allow more detailed first occurrences of the foraminifers to be recorded during the early Viséan Age; the Dalton Beds (Formation) reaches up to 255 m thick, but only the upper c. 30 m is exposed at Barker Scar.
Riley (Reference Riley1993) questioned the validity of the Barker Scar section as a stratotype because lateral sections with palaeosols (2 km apart) were correlated with bed i in Barker Scar where some sandy strata are recorded, and compared with the Davidsonina carbonaria beds of Garwood (Reference Garwood1913). He assumed a non-sequence zone in the lower part of the succession, also supported by the absence of the transition from fasciculate (Siphonodendron) to cerioid (Lithostrotion) rugose corals, as recorded in Ireland and Wales.
Johnson et al. (Reference Johnson, Soper and Burgess2001) revised the stratigraphy in the region and, at Plumpton quarries, they described the topmost 1.8 m of the Dalton Formation (lithological units were described for first time as formations in this work) as being composed of compact, pale brown, fine-grained dolomites and sandstones with traces of carbonaceous rootlets, interpreted as palaeosols. This confirms the non-sequence in the Dalton Formation, but not in the stratotype section at the base of the Park Limestone Formation. In Barker Scar, they considered that beds e–i contained similar sandy and partly dolomitized characteristics of the upper part at Plumpton, including the non-sequence interval (Fig. 2). Furthermore, they described bed j as a grey, dolomitized, bioclastic packstone with quartz grains, and compared it with the Park Limestone Formation, but less darker. They recognized this bed as a mappable horizon and, although no diagnostic fossils were recorded, they considered bed j as the base for the Park Limestone Formation (Fig. 2).
This lithological change at the base of the Park Limestone Formation was adopted by Waters et al. (Reference Waters, Dean, Jones, Somerville, Waters, Somerville and Jones2011, Reference Waters, Cózar, Somerville, Haslam, Millward and Woods2017), although the stratigraphic level for the base of the Holkerian Substage (with diagnostic fossils) was certainly not modified by Johnson et al. (Reference Johnson, Soper and Burgess2001) from the original position at the base of bed k (Fig. 2).
In the Askrigg Block, the lateral equivalent to the succession in South Cumbria is composed of the Chapel House Limestone and Kilnsey formations (Arthurton et al. Reference Arthurton, Johnson and Mundy1988). The Kilnsey Formation is subdivided into the Scaleber Force Limestone and Scaleber Quarry Limestone members, and the Arundian–Holkerian boundary is supposedly located at the boundary between both members. Foraminifers in the Scaleber Force Limestone Member are typically Conilidiscus bucculentus (Conil & Lys, Reference Conil and Lys1964), C. settlensis Conil in Conil et al. (Reference Conil, Longerstaey and Ramsbottom1980), Uralodiscus sp., Planoarchaediscus and Glomodiscus miloni (Pelhâte, Reference Pelhâte1967), an assemblage assigned to the Arundian (Strank in Arthurton et al. Reference Arthurton, Johnson and Mundy1988). Important macrofauna in the Scaleber Quarry Limestone Member are Lithostrotion vorticale (Parkinson, Reference Parkinson1808), Composita ficoidea, Davidsonina carbonaria, Linoprotonia corrugatohemispherica associated with small ‘concavus stage and partial angulatus stage Archaediscus and Nodosarchaediscus’, as well as Koskinotextularia cribriformis and Pojarkovella nibelis (Arthurton et al. Reference Arthurton, Johnson and Mundy1988). Fossil assemblages can be readily compared with that of the Barker Scar section as well as some of the macrofauna, and the base of the Scaleber Quarry Limestone Member can therefore be correlated with the base of the Park Limestone Formation.
The occurrence of the non-sequence interval in the upper part of the Chapel House Limestone Formation and Archaediscus at concavus stage from the base of the Kilnsey Formation led Waters et al. (Reference Waters, Cózar, Somerville, Haslam, Millward and Woods2017) to correlate the base of the latter with the base of the Park Limestone Formation. At the preserved base of the Chapel House Limestone Formation in the Silverdale Borehole (this formation has a maximum thickness of 12 m; Murray, Reference Murray1983), the assemblage is composed of numerous typical upper Tournaisian and lower Viséan forms (Caligella, Dainella, Eosinopsis, Septabrunsiina, Eoparastaffella, Septatournayella, Eblanaia, Spinobrunsiina, Tournayellina, and Spinochernella, as well as Glomodiscus sp., G. miloni, Uralodiscus adindanii Brenckle & Marchant (Reference Brenckle and Marchant1987), U. elongatus (Conil & Lys, Reference Conil and Lys1964), U. rotundus (Chernysheva, Reference Chernysheva1948), and Conilidiscus settlensis) which, in addition to Latiendothyranopsis menneri solida (Conil & Lys, Reference Conil and Lys1964), Nodosarchaediscus, Plectogyranopsis moraviae (Conil & Longerstaey in Conil et al. Reference Conil, Longerstaey and Ramsbottom1980) and P. settlensis (Conil & Longerstaey in Conil et al. Reference Conil, Longerstaey and Ramsbottom1980)), allow the assemblage to be assigned to the Cf4δ subzone (Figs 2, 3). The upper part of the formation is dolomitized and does not contain foraminifers, and there is a non-deposition and development of a palaeosol (Fig. 2). At the base of the overlying Kilnsey Formation, Archaediscus at concavus stage, Omphalotis minima and Pojarkovella sp. were recorded. Specimens of Pojarkovella sp., Holkeria? and Ninella are recorded 7 m above the base, and typical Pojarkovella nibelis, Koskinotextularia, Archaediscus concavus trans. angulatus stage and Endothyranopsis compressa (Rauzer-Chernousova & Reitlinger in Rauzer-Chernousova et al. Reference Rauzer-Chernousova, Belyaev and Reitlinger1936) occur 2 m above (Fig. 2). In higher levels of the formation, and occurring in several steps, are Archaediscus at angulatus stage, Pseudoendothyra and Endothyranopsis crassa (Brady, Reference Brady1876) as well as Koskinobigenerina, which suggest a relatively condensed succession in Arundian – lower Asbian strata compared with other regions in northern England, but including all the foraminiferal subzones defined by Conil et al. (Reference Conil, Longerstaey and Ramsbottom1980).

Fig. 3. Selected foraminiferal key species from the Askrigg Block in northern England and in the Montagne Noire in southern France. Scale bar: (a–g) 200 μm and (h–t) 400 μm. (a) ‘Archaediscus’ at concavus stage, level Pc5339, Valuzière section. (b) ‘Archaediscus’ at concavus stage, level EWJ1565, Silverdale Borehole. (c) ‘Archaediscus’ at concavus stage transitional to the angulatus stage, level EWJ1565, Silverdale Borehole. (d) Nodosarchaediscus cornua (Conil & Lys, Reference Conil and Lys1964), level Pc5339, Valuzière section. (e) Nodosarchaediscus cf. saleei (Conil & Lys, Reference Conil and Lys1964), level Pc4660, Puech de la Suque section. (f) Nodosarchaediscus sp., level Pc5148, Puech de la Suque Hill 248 section. (g) ‘Archaediscus’ at concavus stage transitional to the angulatus stage, level Pc5337, Valuzière section. (h) Pojarkovella sp., level EWJ1563, Silverdale Borehole. (i) Pojarkovella sp., level EWJ1566, Silverdale Borehole. (j) Pojarkovella sp., level EWJ1562, Silverdale Borehole. (k) Pojarkovella nibelis, level EWJ1561, Silverdale Borehole. (l) Endothyranopsis aff. compressa, level EWJ1573, Silverdale Borehole. (m) Latiendothyranopsis menneri solida, level EWJ1573, Silverdale Borehole. (n) Plectogyranopsis moraviae, level EWJ1573, Silverdale Borehole. (o) Plectogyranopsis settlensis, level Pc4900, Pic de Vissou section. (p) Ugurus intermedius Vachard et al. (Reference Vachard, Coronado, Izart and Cózar2018), level Pc5537, Valuzière section. (q) Endothyranopsis compressa, level EWJ1562, Silverdale Borehole. (r) Omphalotis minima, level Pc4983, Puech de la Suque section. (s) ‘Millerella’ aff. infulaeformis Ganelina (Reference Ganelina1951), level Pc4896, Pic de Vissou section. (t) Koskinotextularia sp., level Pc4660, Puech de la Suque section.
It is necessary to highlight the interval from the first occurrence of Archaediscus at concavus stage and Pojarkovella sp. up to the first occurrence of Koskinotextularia and P. nibelis. The latter taxa allow the correlation with bed k of the Park Limestone of the Barker Scar stratotype; this interval is therefore correlated with the upper part of the Dalton Beds and assigned to the Arundian Substage. The level with Koskinotextularia/P. nibelis in other outcrops where the Scaleber Force Limestone and Scaleber Quarry Limestone members are well differentiated is coincident with the boundary between both members, as well as the abovementioned macrofauna (Dunham & Wilson, Reference Dunham and Wilson1985; Arthurton et al. Reference Arthurton, Johnson and Mundy1988). The boundary between Scaleber Force Limestone and Scaleber Quarry Limestone members mimics the fossil record of that in the boundary between Dalton Formation and Park Limestone Formation.
In the Kilnsey Crag, the type locality for the Kilnsey Limestone Formation, the younger member is well-exposed and only the top 1–2 m of the Scaleber Force Limestone Member is seen (see Waters et al. Reference Waters, Cózar, Somerville, Haslam, Millward and Woods2017, fig. 6). The occurrence of Endothyranopsis compressa is recorded at the top of the lower member.
2.c. Lower–middle Viséan boundary interval in southern France
In the Montagne Noire, the transition between the lower and middle Viséan substages is rarely recognized in sections because of the poor exposure and commonly hostile facies for the foraminifers (Cózar et al. Reference Cózar, Izart, Vachard and Coronado2017; Vachard et al. Reference Vachard, Coronado, Izart and Cózar2018). Archaediscus at concavus stage has only been recorded at Valuzières section, at the same level as Pojarkovella sp. and Endothyranopsis compressa (Figs 3, 4). However, Archaediscus at concavus stage transitional to the angulatus stage and Pojarkovella nibelis, as well as the regional markers such as Ugurus, primitive ‘Millerella’ and Cribrospira, occur only 0.8 m above this level (Fig. 3p, s). This small difference in the Valuzières section between the first appearances of the taxa does not allow a clear conclusion if this intermediate zone at the top of the lower Viséan strata is present. It could be poorly developed, but additional sections should confirm this short interval. Nevertheless, the succession in shallow-water platforms of the Montagne Noire can also be considered as relatively condensed, with a limited thickness. Moreover, below the Valuzières Formation a typical griotte limestone occurs, the Combe Roland Formation, which is rather poor in foraminifers; in Tourière (Fig. 4) and in the type sections, it also contains Archaediscus at concavus stage as well as transitional forms to the angulatus stage (Cózar et al. Reference Cózar, Izart, Vachard and Coronado2017; Vachard et al. Reference Vachard, Coronado, Izart and Cózar2018). Calciturbidites of the slope facies do not contain the transition to the lower Viséan substage; however, the lower part of the middle Viséan Colonnes Formation also shows the same first occurrences of the taxa (Fig. 5). The stratigraphic records of the taxa are rather similar to those in northern England.

Fig. 4. Biostratigraphic sketch of the Valuzières and Tourière sections, Montagne Noire (southern France). CB – Combe Rolland Formation; Tou – Tourière Formation.

Fig. 5. Biostratigraphic sketch of the Puech de la Suque-Hill 248 and Pic de Vissou sections, Montagne Noire (southern France).
2.d. Lower–middle Viséan boundary interval in the western Meseta of Morocco
This study is limited to the western Meseta because the lower and middle Viséan substages are not documented in the eastern Meseta (M. Berkhli, unpub. MSc thesis, University of Lille, 1993). The occurrence of the lower Viséan substage in Morocco has been a much debated point since early studies in the region, because assemblages older than the V2a are rarely documented (Huvelin & Mamet, Reference Huvelin and Mamet1997). Subsequent to the chronostratigraphical changes introduced by Conil et al. (Reference Conil, Groessens and Pirlet1977), many sections were considered from their bases to be middle Viséan (V2a) in nature; problematic lower Viséan strata were later documented (e.g. Verset, Reference Verset1983, Reference Verset1988), although it is not clear whether the successions based on those assemblages compared with the V2a were considered as lower or middle Viséan (e.g. Vachard et al. Reference Vachard, Termier and Termier1977). To avoid problems with the comparison with the European zonal scales, these authors proposed a regional zonal scheme for Morocco using foraminifers and calcareous algae (Vachard, Reference Vachard1988b; Vachard & Tahiri, Reference Vachard and Tahiri1991); this has been updated (in stratigraphic ranges and guides) in subsequent studies (Vachard & Berkhli, Reference Vachard and Berkhli1992; Berkhli et al. Reference Berkhli, Vachard and Paicheler2000; Izart et al. Reference Izart, Tahiri, El Boursoumi, Vachard, Saidi, Chèvremont and Berkhli2001, Reference Izart, Vachard and Berkhli2017).
The basal zone Cfm1 is characterized by the primitive forms of Uralodiscus and Glomodiscus, with U. rotundus as a marker and Archaediscus convexus Grozdilova & Lebedeva in Dain & Grozdilova (Reference Dain and Grozdilova1953) in its upper part. This zone is typically represented in the Sidi Sebaa Formation (Fig. 6), and was correlated with the upper part of the Moliniacian Substage and lower part of the Livian, upper Arundian and lower Holkerian, upper MFZ11 and lower MFZ12, upper V2a and lower V2b, upper Cf4δ and lower Cf5 substages (Izart et al. Reference Izart, Vachard and Berkhli2017). The base of the succession in the western Meseta seems to be only present at the Sidi Sebaa section where, in the lithological unit E2 (Fig. 6), Uralodiscus rotundus, Glomodiscus oblongus, Lapparentidiscus nanus (Reitlinger, Reference Reitlinger1969) and Archaediscus at involutus stage (Vachard & Fadli, Reference Vachard and Fadli1991) occur. This assemblage can be compared with the Cf4γ-Cf4δ zones of Conil et al. (Reference Conil, Longerstaey and Ramsbottom1980, Reference Conil, Groessens, Laloux, Poty and Tourneur1991), most of the MFZ11 zone of Poty et al. (Reference Poty, Devuyst and Hance2006), and the upper Moliniacian Substage (Devuyst et al. Reference Devuyst, Hance and Poty2006). Above this level the first change in the assemblages is characterized by the appearance of Archaediscus convexus (at concavus stage), which, in addition to Sidi Sebaa, it is also recognized at north Oulmès (Vachard & Tahiri, Reference Vachard and Tahiri1991; Izart et al. Reference Izart, Tahiri, El Boursoumi, Vachard, Saidi, Chèvremont and Berkhli2001), Agouraï (M. Ben Abbou, unpub. MSc thesis, Univiversity of Sidi Mohamed ben Abdellah, Fès, 1990) and Skoura boutonnière (Izart et al. Reference Izart, Poty and Vieslet1989; unpublished data). In this interval, Endothyranopsis compressa has been documented, although generally as ex group species or in open nomenclature (E. cf. compressa and E. aff. compressa) (Vachard & Tahiri, Reference Vachard and Tahiri1991; Izart et al. Reference Izart, Tahiri, El Boursoumi, Vachard, Saidi, Chèvremont and Berkhli2001).

Fig. 6. Biostratigraphic sketch of the Sidi Sebaa section, Morocco. Bed thickness based on D. Fadli (unpub. PhD thesis, University of Rabat, 1990).
In an early attempt, Izart et al. (Reference Izart, Tahiri, El Boursoumi, Vachard, Saidi, Chèvremont and Berkhli2001) subdivided the Cfm1 zone into the Cfm1a subzone characterized by Uralodiscus rotundus and a younger Cfm1b subzone characterized by Archaediscus convexus (Fig. 7), although these subzones were not used in later studies.

Fig. 7. Correlation of the different proposals for the MFZ11B subzone in eastern Palaeotethys basins and the Cfm1b subzone from Morocco, according to their authors.
Assemblages composed of Uralodiscus/Glomodiscus/‘Archaediscus’ at concavus stage are predominant in this upper interval, which can be correlated with the unnamed zone recorded in England at the top of the Arundian Substage.
The Cfm2 zone is currently characterized by Uralodiscus and Conilidiscus as markers (e.g. Berkhli et al. Reference Berkhli, Vachard and Paicheler2000), but also by Endothyranopsis compressa and Forschia parvula Rauzer-Chernousova (Reference Rauzer-Chernousova1948b) in older works (Vachard, Reference Vachard1988b). The zone was correlated with parts of the Cf5, MFZ12, Livian and Holkerian substages and upper part of the V2b zone (Izart et al. Reference Izart, Vachard and Berkhli2017). Its type locality is Tizi ben Zizouit (Termier et al. Reference Termier, Termier and Vachard1975). In this case, the foraminiferal stratigraphic record in different sections is not as consistent as for the previous stratigraphic records. In the Mdakra Massif and El Hamman Ridge, Conilidiscus appears much earlier than Endothyranopsis compressa (Vachard & Fadli, Reference Vachard and Fadli1991; Izart et al. Reference Izart, Tahiri, El Boursoumi, Vachard, Saidi, Chèvremont and Berkhli2001), whereas in Oulmès, E. compressa occurs much earlier (Vachard & Tahiri, Reference Vachard and Tahiri1991). In Agouraï, both taxa occur from the base of the succession, with E. compressa in the basal conglomeratic/sandy unit (Berkhli et al. Reference Berkhli, Vachard and Paicheler2000). In Skoura, only Conilidiscus occurs (Izart et al. Reference Izart, Poty and Vieslet1989, as Tubispirodiscus). The subdivision of the Cfm1b and Cfm2 zones therefore does not seem useful, because the late occurrence of Conilidiscus compared with taxa such as ‘Archaediscus’ at concavus stage and E. compressa is not clear enough and the succession is in need of revision. The suite of foraminifers recorded in this interval might also be compared with those of the unnamed interval at the top of the Arundian Substage.
The Cfm3 zone is characterized by Koskinotextularia bradyi (von Möller, Reference von1879) as a guide. It is correlated with the upper part of the Cf5, Holkerian, Livian and MFZ12 and with the V3a zones. The second marker, Pojarkovella nibelis, is only known in one section close to Tiflet town (Vieslet, Reference Vieslet1983), whereas the taxon is mostly recorded in the upper Viséan limestones of Morocco (Vachard & Tahiri, Reference Vachard and Tahiri1991). The type locality of this zone is at Irhzer Aoujgal (Termier et al. Reference Termier, Termier and Vachard1975). Unquestionable middle Viséan markers are only recognized from this Cfm3 zone. The virtual lack of Pojarkovella nibelis is a handicap for comparison with Europe because Koskinotextularia is a comparatively rare taxon; on contrast, P. nibelis from its basal range is commonly recorded and therefore more useful for the recognition of middle Viséan carbonates.
These chronozones are temporally slightly out of phase compared with European zones, and some of the European markers arrived later to the Meseta (Vachard & Tahiri, Reference Vachard and Tahiri1991). The occurrences of typical taxa of the Cfm1 and Cfm2 zones in sections assigned to the Cfm3 zone (e.g. Vieslet, Reference Vieslet1983) significantly complicate the recognition of the lower and middle Viséan substages in Morocco.
3. Lower–middle Viséan boundary interval in eastern Palaeotethys
Unusual foraminiferal assemblages have been described from China, Iran and Turkey, whereas they are unknown from other countries, such as Australia, in this palaeobiogeographic region.
3.a. Lower–middle Viséan boundary interval in China
Devuyst et al. (Reference Devuyst, Hance, Hou, Wu, Tian, Coen and Sevastopulo2003) and Devuyst (unpub. PhD thesis, Université catholique de Louvain, 2006) suggested the presence of an interval zone at the top of the Pengchong section (Tournaisian–Viséan boundary global boundary stratotype section and point), unrecognized in Belgium. This interval zone was based on the presence of primitive archaediscids (Uralodiscus, Glomodiscus and Conilidiscus) after the first occurrence of Pojarkovella nibelis. This zone was named as zone G in Devuyst et al. (Reference Devuyst, Sevastopulo, Hance, Hou, Kalvoda and Wu2004) following the zones defined previously in the section by Hance et al. (Reference Hance, Muchez, Hou and Wu1997). However, the Pengchong section is characterized, overall, by poor archaediscid assemblages, attributed by the authors to a lack of deep-water foraminifers in turbidites, due to little downslope mixing of the fauna (Devuyst et al. Reference Devuyst, Sevastopulo, Hance, Hou, Kalvoda and Wu2004). The occurrence of primitive assemblages of archaediscids at the base of inferred middle Viséan rocks is not sufficient to justify the occurrence of this new zone, because this fact is also recognized in Morocco, Moravia, Russia and Kazakhstan (e.g. Reitlinger et al. Reference Reitlinger, Vdovenko, Gubareva, Shcherbakov, Wagner, Winkler Prins and Granados1996; Kalvoda, Reference Kalvoda2002; Brenckle & Milkina, Reference Brenckle and Milkina2003; Kulagina et al. Reference Kulagina, Gibshman and Pazukhin2003); this can only be a matter of longer persistence of the taxa for regional ecological factors.
Hance et al. (Reference Hance, Hou and Vachard2011) subdivided the MFZ11 – Uralodiscus rotundus Zone into a lower subzone (MFZ11A), containing mostly the classical markers defined in Poty et al. (Reference Poty, Devuyst and Hance2006), and an upper subzone (MFZ11B), characterized by the first occurrence of Pojarkovella (Fig. 7); the specimens first occurring at this level are more primitive than P. nibelis, a marker for the MFZ12 Zone. This MFZ11B subzone was only recognized in two sections, Mopanshan and Jiusi, but neither contains the transition into the MFZ12, meaning that the transition into the middle Viséan substage cannot be studied in detail. Other sections in the region contain poorer assemblages in the upper part of the lower Viséan substage, and the MFZ11B subzone could not be distinguished and, contrary to the previously studied Pengchong and Yajiao sections by Devuyst et al. (Reference Devuyst, Hance, Hou, Wu, Tian, Coen and Sevastopulo2003, Reference Devuyst, Sevastopulo, Hance, Hou, Kalvoda and Wu2004) and Devuyst (unpub. PhD thesis, Université catholique de Louvain, 2006), Hance et al. (Reference Hance, Hou and Vachard2011) only recognized the MFZ11 and MFZ12 zones in the same sections. It has to be highlighted that the latter authors considered Endothyranopsis ex gr. compressa as typical of the lower Viséan MFZ11 zone, but was only illustrated from the MFZ11B subzone. It must also be noted that the first occurrence of Conilidiscus is located by those authors in the MFZ11A subzone.
The unusual record of the primitive archaediscids in China represents a significant complication when trying to calibrate the first occurrences of other taxa, as well as establish a precise definition of the foraminiferal zones. But it also has to be added that the first occurrence of Archaediscus at concavus stage and the transitional forms to the angulatus stage are both recorded from the MFZ12 zone (e.g. Hance et al. Reference Hance, Hou and Vachard2011, pl. 51, fig. 14) and only primitive Pojarkovella first occurs below this substage.
3. Lower–middle Viséan boundary interval in Turkey
Okuyucu et al. (Reference Okuyucu, Vachard and Göncüoğlu2013) studied material assigned to the MFZ11B subzone in the Istanbul terrane. These authors considered the first appearances of Uralodiscus rotundus (and other species of the genus), Paraarchaediscus, Cribranopsis and Conilidiscus and the disappearance of Eoendothyranopsis, Eoparastaffella, Pseudolituotubella, Condrustella and Eotextularia characteristic of the MFZ11 zone. They interpreted that Paraarchaediscus (‘Archaediscus’ herein) and Conilidiscus might be guides for the MFZ11B subzone. However, these taxa are recorded from near the base of the MFZ11 zone in Belgium (Poty et al. Reference Poty, Devuyst and Hance2006), as well as from the MFZ11A subzone in China (Hance et al. Reference Hance, Hou and Vachard2011); they might therefore be used as regional markers, but not to characterize a global MFZ11B subzone.
Compared with the European zones, this modification into the MFZ11B should be correlated with the Cf4γ-δ; clearly, it cannot be compared with the MFZ11B in China (Fig. 7). There is no mention of other important taxa in Turkey such as primitive Pojarkovella, the first Endothyranopsis, and therefore no evidence of any unusual foraminiferal assemblage below the typical middle Viséan strata.
3.c. Lower–middle Viséan boundary interval in Iran
Zandkarimi et al. (Reference Zandkarimi, Najafian, Vachard, Bahrammanesh and Vaziri2016) identified the MFZ11B subzone on the basis of advanced Glomodiscus and Archaediscus in NW Alborz. This definition is rather similar to that in Turkey (Fig. 7). The rest of the assemblage contains Archaediscus/Paraarchaediscus at involutus stage, and Ammarchaediscus, Planoarchaediscus and Eotextularia; the assemblage should therefore also be correlated with the Cf4γ-δ subzones of Conil et al. (Reference Conil, Longerstaey and Ramsbottom1980, Reference Conil, Groessens, Laloux, Poty and Tourneur1991), and with the classical concept of the MFZ11 in Poty et al. (Reference Poty, Devuyst and Hance2006). The first occurrence of primitive Pojarkovella is at the same horizon as P. nibelis, and no difference exists.
Zandkarimi et al (Reference Zandkarimi, Vachard, Najafian, Hamdi and Mosaddegh2017, Reference Zandkarimi, Vachard, Najafian, Mosaddegh and Ehteshami-Moinabadi2018) only recognized the MFZ11 and MFZ12 zones in northern Alborz, and they did not subdivide the former zone. The occurrence of Pojarkovella spp. is synchronous in the main stratigraphic sections at Naserabad and Dozdehban, and also coincident with the first appearance of Koskinotextularia. Zandkarimi et al. (Reference Zandkarimi, Vachard, Najafian, Mosaddegh and Ehteshami-Moinabadi2018) described the occurrence of Endothyranopsis compressa from the MFZ11 zone, although this species (generally very rare in the region) was used for the recognition of the base of the middle Viséan substage at the Naserabad section (at sample Ns102) whereas P. nibelis occurs 1 m above (at Ns103). Other specimens recorded in older rocks are identified with a question mark, and its presence in the lower Viséan deposits of Iran is therefore questioned.
However, although not acknowledged by the authors, ‘Archaediscus’ at concavus stage are recorded from the early Viséan Age (Zandkarimi et al. Reference Zandkarimi, Najafian, Vachard, Bahrammanesh and Vaziri2016, figs 7.35, 7.38; Zandkarimi et al. Reference Zandkarimi, Vachard, Najafian, Hamdi and Mosaddegh2017, figs 7.29, 7.30). Furthermore, although Zandkarimi et al. (Reference Zandkarimi, Vachard, Najafian, Hamdi and Mosaddegh2017) recognized the occurrence of Paraarchaediscus at concavus transitional to the angulatus stage from the middle part of the middle Viséan strata, specimens are recorded from closer positions to the base of the middle Viséan Stage (Zandkarimi et al. Reference Zandkarimi, Najafian, Vachard, Bahrammanesh and Vaziri2016, fig. 9.12; Zandkarimi et al. Reference Zandkarimi, Vachard, Najafian, Hamdi and Mosaddegh2017, figs 7.37, 7.38, 7.40).
Fassihi et al. (Reference Fassihi, Kulagina, Esfahani and Nurgalilev2018) subdivided the succession at the Sanandaj-Sirjan region in Central Iran into two local zones. The older zone was correlated with the MFZ11B subzone due to the occurrence of Glomodiscus, Uralodiscus and Paraarchaediscus and assigned a latest early Viséan age (Fig. 7). The youngest local zone was characterized by the appearance of Lapparentidiscus bokanensis Vachard (Reference Vachard1980) and the disappearance of dainellids. These authors correlated this zone with the lowest ‘upper’ Viséan or MFZ12 strata of Belgium. (It must be noted that, in the Russian scale, the Viséan Stage is only divided into lower and upper, and the Tulian substage, more or less equivalent to the middle Viséan substage in western Europe, is the lower part of the upper Viséan substage.) However, there is no foraminifer which suggests a middle Viséan age within this assemblage, and the entire succession should be assigned to the Cf4γ-δ subzones of early Viséan age.
4. Biozone proposal and correlation
The biostratigraphic records of the analysed basins and/or sections confirm the existence of an intermediate interval (biozone) between the classical well-known chronozones (V2a and V2b) defined in western Europe (Fig. 8). Some authors have interpreted this as an interval zone (Devuyst et al. Reference Devuyst, Hance, Hou, Wu, Tian, Coen and Sevastopulo2003) that should be characterized by the coexistence of typical early and middle Viséan faunas. However, this suggestion is rather problematical because it is difficult to establish precisely the last occurrence (disappearance) of the key taxa, as well as ecological factors which allowed the longer survival of fauna, which is not acknowledged. These problems are exemplified with the succession in the western Meseta of Morocco, where some taxa arrived later to the region and others survived longer (i.e. had extended stratigraphic ranges) than in the neighbouring European basins (Vieslet, Reference Vieslet1983; Vachard & Tahiri, Reference Vachard and Tahiri1991; Cózar et al. Reference Cózar, Vachard, Izart and Coronado2019).

Fig. 8. Stratigraphic range of the main foraminiferal markers.
It is considered here more appropriate to precisely establish the first occurrences of taxa rather than the last occurrences to characterize this biozone. The main guides for the zone are considered to be the first occurrence of ‘Archaediscus’ at concavus stage and primitive Pojarkovella (Fig. 3a, b, h–j). Furthermore, Endothyranopsis compressa first occurs slightly higher in the zone (Fig. 3q), whereas the first Endothyranopsis s.s. (named in the literature as E. ex gr. compressa or aff. compressa or cf. compressa) first occur from the base of the zone (Fig. 3l). Indirectly, this zone usually contains a mixture of preserved early Viséan genera (Uralodiscus, Glomodiscus, Conilidiscus), although this fact cannot be considered as a definitive feature.
It must be highlighted that in the younger zone (Cf5, MFZ12, V2b) with the same main guides as classically known, Pojarkovella nibelis and Koskinotextularia, the ‘Archaediscus’ at concavus transitional forms to the angulatus stage are recorded (Fig. 3c, g, k, t) nearly from the base (Fig. 8).
In terms of nomenclature, this new zone is a problem. It roughly corresponds to the MFZ11B subzone defined in China by Hance et al. (Reference Hance, Hou and Vachard2011), as well as the MFZ11B subzone in Iran (sensu Zandkarimi et al. Reference Zandkarimi, Najafian, Vachard, Bahrammanesh and Vaziri2016), but differs significantly from the MFZ11B subzone in Turkey (sensu Okuyucu et al. Reference Okuyucu, Vachard and Göncüoğlu2013). Furthermore, the notation MFZ11B suggests intuitively that it is part of the lower Viséan substage. However, compared with the stratotype of the Livian Substage in Belgium, it is considered as the base of the middle Viséan substage; where the MFZ or Cf zones were defined, the zone is considered here as part of the middle Viséan substage. A subdivision into MFZ12α (with the newly proposed guides) and MFZ12β (with the classical taxa described by Poty et al. Reference Poty, Devuyst and Hance2006) would therefore be more appropriate (Figs 8, 9). Compared with the classical chronozones, the Cf5 zone was defined on the basis of Koskinotextularia, Pojarkovella nibelis and ‘Archaediscus’ at concavus stage (Conil et al. Reference Conil, Longerstaey and Ramsbottom1980, Reference Conil, Groessens, Laloux, Poty and Tourneur1991); subdivision into a Cf5α (this new one) and Cf5β (typical Cf5) is therefore the most logical subdivision because it is correlated with the non-sequence interval in the Lives section of Belgium and considered as part of the middle Viséan substage (Fig. 8).

Fig. 9. The new biostratigraphic zones proposed here and correlation with previous biozones. Boxes with diagonal lines indicate hiatus or absence of recognized faunas.
5. Discussion
The occurrence of this MFZ12α or Cf5α subzone from the western extreme of the Palaeotethys across to the eastern basins of this ocean confirms the validity of this subzone, and that it should be considered as a potential global event; it is certainly useful for foraminiferal correlations throughout Palaeotethys. However, its occurrence has not been demonstrated in geographically intermediate regions of the Palaeotethys, such as Russia. It is difficult to validate the stratigraphic record of the involved species in this region because the Tulian Substage (or its lateral equivalents) are commonly of little thickness or composed mostly of siliciclastics. Biozones described there usually list numerous foraminiferal guides to characterize this substage. However, they do not record the first occurrence of individual taxa or the presence of any type of successive steps in assemblages. This list of markers includes Endothyranopsis s.s., Globoendothyra s.s., Lituotubella, Mstinia (as Haplophragmella), Urbanella miranda (Rauzer-Chernousova, Reference Rauzer-Chernousova1948a), Eostaffella, Pseudoendothyra (as Parastaffella), Vissariotaxis exilis (Vissarionova, Reference Vissarionova1948), Cribrospira, Omphalotis omphalota (Rauzer-Chernousova & Reitlinger in Rauzer-Chernousova et al. Reference Rauzer-Chernousova, Belyaev and Reitlinger1936), O. minima, Archaediscus krestovnikovi (Rauzer-Chernousova, Reference Rauzer-Chernousova1948d) and A. moelleri (Rauzer-Chernousova, Reference Rauzer-Chernousova1948a), and also Koskinotextularia, Pojarkovella nibelis, Consobrinellopsis, Cribrostomum and Palaeotextularia (e.g. Rauzer-Chernousova, Reference Rauzer-Chernousova1948c; Lipina & Reitlinger, Reference Lipina and Reitlinger1970; Gibshman, Reference Gibshman, Ross, Ross and Brenckle1997; Kulagina et al. Reference Kulagina, Gibshman and Pazukhin2003). In most zonal schemes, the main guides of the Tulian are restricted to Endothyranopsis compressa and ‘Archaediscus’ krestovnikovi.
This foraminiferal list suggests that the Tulian Substage (and equivalent) contains amalgamated foraminiferal markers for the middle Viséan substage and lower part of the upper Viséan substage in western Europe (most likely the result of poor carbonate precipitation in this Russian stage), from the Cf5 to the Cf6β zones and subzones of Conil et al. (Reference Conil, Groessens, Laloux, Poty and Tourneur1991). However, the main guides are generally similar to those of the Cf5α and MFZ12α, in that they first occur together at the base of this substage in some published sections (Rauzer-Chernousova, Reference Rauzer-Chernousova1948c; Postoyalko, Reference Postoyalko, Malakhova and Chuvashov1975; Kulagina et al. Reference Kulagina, Gibshman and Pazukhin2003; Kulagina, Reference Kulagina2011), whereas Pojarkovella nibelis first appears in younger beds as well as some of the above-listed Tulian markers (Kulagina & Klimenko, Reference Kulagina and Klimenko2014; Klimenko et al. Reference Klimenko, Kulagina, Gorozhanina, Kochetova, Zhernovkova and Puchkov2018). It must be noted that under the name of ‘Archaediscus’ krestovnikovi both concavus and angulatus stages are indistinctly identified, but those specimens illustrated from the base of the substage seem to correspond to the concavus stage. Additionally, the co-occurrence of the primitive archaediscids at the base of the Tulian Substage is a common feature (Bensh et al. Reference Bensh, Rumyantseva, Sergunkova, Wagner, Winkler Prins and Granados1996; Reitlinger et al. Reference Reitlinger, Vdovenko, Gubareva, Shcherbakov, Wagner, Winkler Prins and Granados1996; Brenckle & Milkina, Reference Brenckle and Milkina2003; Kulagina et al. Reference Kulagina, Gibshman and Pazukhin2003).
It can therefore be interpreted that this Cf5α or MFZ12α subzone also occurs in these intermediate (central) positions of the Palaeotethys, but that more detailed biostratigraphic ranges of the foraminifers in the Tulian Substage (Fig. 7) are necessary to clarify if the markers are exactly the same or if they present some peculiarities.
On the other hand, the validity of the MFZ11B subzone cannot be corroborated; although it marks the first occurrence of the ‘Archaediscus’ at involutus stage as well as Conilidiscus, the original definition in China also includes the first occurrence of the primitive Pojarkovella, which is situated in the middle Viséan strata (Fig. 8). Although the occurrence of ‘Archaediscus’ and Conilidiscus would correspond to the Cf4γ subzone of Conil et al. (Reference Conil, Groessens, Laloux, Poty and Tourneur1991), because of the scarcity of archaediscids in China (Devuyst et al. Reference Devuyst, Hance, Hou, Wu, Tian, Coen and Sevastopulo2003; Hance et al. Reference Hance, Hou and Vachard2011) it is not always recognizable in some sections. The guides for the recognition of the Cf4δ subzone of Conil et al. (Reference Conil, Longerstaey and Ramsbottom1980) were relatively limited, which makes its recognition in other basins more difficult (as previously criticized by Poty et al. Reference Poty, Devuyst and Hance2006). However, the first occurrences of Latiendothyranopsis menneri solida (Fig. 3m), Cribranopsis, Nodosarchaediscus (Fig. 3d–f), Consobrinellopsis, Lituotubella, Omphalotis minima (Fig. 3r), Plectogyranopsis settlensis (Fig. 3o) and Plectogyranopis moraviae (Fig. 3n) are recognized in this interval ; since these taxa have a wider or even worldwide distribution, they are more useful taxa than the original guides. This suite of foraminifers would allow the subdivision of the MFZ11 into three subzones: MFZ11α, MFZ11β and MFZ11γ (Fig. 9). The MFZ11α subzone is characterized by the first occurrence of Uralodiscus rotundus, as well as common species of the Glomodiscus (e.g. G. miloni) and Uralodiscus (U. elongatus). The MFZ11β subzone is characterized by the first occurrence of primitive ‘Archaediscus’ at involutus stage with well-developed microgranular layer (Paraarchaediscus auct.), and Conilidiscus. The MFZ11γ subzone is characterized by the first occurrence of Latiendothyranopsis menneri solida, Cribranopsis, Nodosarchaediscus, Consobrinellopsis, Lituotubella, Omphalotis minima, the first Endothyranopsis s.s. and large species of Plectogyranopsis (Plectogyranopsis settlensis and P. moraviae). There seem to be small differences in first occurrences of these species between Belgium and northern France (e.g. Conil et al. Reference Conil, Longerstaey and Ramsbottom1980, Reference Conil, Groessens, Laloux, Poty and Tourneur1991; Poty et al. Reference Poty, Devuyst and Hance2006), although the species described have a limited geographic dispersion; in other nearby regions (southern and northern France and England), they seem to occur more or less synchronously. Nevertheless, the precise first occurrences of each individual taxon need further investigation.
6. Conclusions
The revision of selected sections spanning the lower–middle Viséan boundary interval in the western and eastern Palaeotethys basins, including also the Livian (Belgium) and Holkerian (England) stratotypes in England, suggests the occurrence of a new foraminiferal zone in between the classical chronozones defined in those countries. This new subzone (Cf5α or MFZ12α) is characterized by the first occurrences of ‘Archaediscus’ at concavus stage, primitive species of Pojarkovella and the first representatives of the genus Endothyranopsis, including the first occurrence of Endothyranopsis compressa (at least from the middle part of this zone). The zone is intercalated within the classical V2a and V2b, Cf4δ and Cf5 and the MFZ11 and MFZ12 zones. It is partly similar to the MFZ11B zone proposed in China by previous authors. However, the zone is assigned to the middle Viséan substage due to its position compared with the stratotype section for the lower–middle Viséan boundary interval in Belgium, and new nomenclature is proposed to avoid the ambiguity of the original MFZ11B assignation to the lower Viséan substage.
The zone allows a precise correlation between the middle Viséan rocks or regional stages between the base of the Livian Substage in Belgium and the upper Arundian Substage in England, as well as the clear definition of which western Meseta limestones and equivalents in the eastern Palaeotethys (China and Iran) should be considered as being of early or middle Viséan age (a matter of discussion for a long time). The succession in the more central positions of the Palaeotethys (such as Russia) seems to have this biozone present, although further analyses are necessary to subdivide the Tulian Substage with greater precision.
We propose the subdivision of the classical Cf5 zone into a lower Cf5α subzone (for this new zone) and an upper Cf5β subzone, characterized by the classical guides Pojarkovella nibelis and Koskinotextularia, or, in the zonal scheme of Poty et al. (Reference Poty, Devuyst and Hance2006), to a MFZ12α and MFZ12β subzones, respectively. Revision of the markers for the ‘MZF11’ also suggest that the MFZ11 zone, as defined by Poty et al. (Reference Poty, Devuyst and Hance2006), can be subdivided in most basins into three subzones: a lower MFZ11α subzone (characterized by the first occurrence of Uralodiscus rotundus, as well as most species of Glomodiscus), a middle MFZ11β subzone (characterized by the first occurrence of ‘Archaediscus’ at involutus stage and Conilidiscus) and an upper MFZ11γ subzone (characterized by the first occurrence of Nodosarchaediscus, Consobrinellopsis, Lituotubella, Omphalotis minima and some species of Latiendothyranopsis, Endothyranopsis and Plectogyranopsis).
Acknowledgements
Fieldwork for PC and IDS was funded by the Spanish Ministry of Ciencia, Innovación y Universidades (project no. CGL2016-78738BTE). Revision by two anonymous reviewers is acknowledged.









