


Introduction
In the USA, transition to certified organic production requires a 3-year period during which the crop must be managed according to the US Department of Agriculture (USDA) national organic standards (USDA, 2015). To manage pests during the transition and after, organic growers rely on biological processes, such as biological control of pests by natural enemies, and cultural and mechanical practices, such as planting at specific times to avoid pests and using tillage and inter-row cultivation to control weeds. As such, the practices implemented during this transition may differentially affect pest and beneficial organisms, resulting in variable risk for crop losses associated with pest damage (Delate and Cambardella, Reference Delate and Cambardella2004; Lundgren et al., Reference Lundgren, Shaw, Zaborski and Eastman2006; Smith et al., Reference Smith, Barbercheck, Mortensen, Hyde and Hulting2011; USDA, 2015). The choice of specific management practices during the transition can thus inadvertently or intentionally affect key natural enemies, resulting in an increase or decrease in pest suppression.
The transition to organic production and continuing organic management can increase the abundance and diversity of beneficial arthropods in general, and Carabidae (Coleoptera) beetles, specifically (Dritschilo and Wanner, Reference Dritschilo and Wanner1980; Pfiffner and Niggli, Reference Pfiffner and Niggli1996; Bengtsson et al., Reference Bengtsson, Ahnström and Weibull2005; Purtauf et al., Reference Purtauf, Roschewitz, Dauber, Thies, Tscharntke and Wolters2005; Lundgren et al., Reference Lundgren, Shaw, Zaborski and Eastman2006). Carabid beetles are predators of arthropods and weed seeds, and contribute to pest suppression in organically-managed systems, helping to minimize weed and insect pest pressures (Lundgren et al., Reference Lundgren, Shaw, Zaborski and Eastman2006; Menalled et al., Reference Menalled, Smith, Dauer and Fox2007; Ward et al., Reference Ward, Ryan, Curran, Barbercheck and Mortensen2011). To take advantage of the predatory potential of carabids, organic growers may implement certain practices to augment the numbers of this beneficial group, including rotating their cash crop with a winter cover crop (Carmona and Landis, Reference Carmona and Landis1999; Shearin et al., Reference Shearin, Chris Reberg-Horton and Gallandt2008; Ward et al., Reference Ward, Ryan, Curran, Barbercheck and Mortensen2011). However, practices that are frequently used for managing pests in organic systems, including inversion tillage for weed control, may contribute to reductions in carabid abundance and diversity (Holland and Reynolds, Reference Holland and Reynolds2003; Thorbek and Bilde, Reference Thorbek and Bilde2004; Hatten et al., Reference Hatten, Bosque-Pérez, Johnson-Maynard and Eigenbrode2007; Mirsky et al., Reference Mirsky, Ryan, Curran, Teasdale, Maul, Spargo, Moyer, Grantham, Weber, Way and Camargo2012).
Inversion tillage and various forms of primary cultivation, which typically overturn the top 15 cm or more of soil employing such implements as the moldboard plow and different types of inter-row cultivators, are widely used for seed bed preparation, fertility management and weed control in many organic systems (Bond and Grundy, Reference Bond and Grundy2001; Bàrberi, Reference Bàrberi2002; Mirsky et al., Reference Mirsky, Ryan, Curran, Teasdale, Maul, Spargo, Moyer, Grantham, Weber, Way and Camargo2012). Because these implements invert the soil, they can kill or frighten important beneficial soil-dwelling organisms and have been associated with other negative outcomes in agroecosystems, e.g., reduced soil quality, the potential for erosion and higher management costs for labor and fuel (Hatten et al., Reference Hatten, Bosque-Pérez, Johnson-Maynard and Eigenbrode2007; Smith et al., Reference Smith, Barbercheck, Mortensen, Hyde and Hulting2011; Mirsky et al., Reference Mirsky, Ryan, Curran, Teasdale, Maul, Spargo, Moyer, Grantham, Weber, Way and Camargo2012). As such, many organic growers are seeking alternatives to reduce the frequency and intensity of tillage (i.e., depth or area disturbed) in their systems to avoid such detrimental effects, and cover crop-based reduced tillage is emerging as a potential option (Mirsky et al., Reference Mirsky, Ryan, Curran, Teasdale, Maul, Spargo, Moyer, Grantham, Weber, Way and Camargo2012, Reference Mirsky, Ryan, Teasdale, Curran, Reberg-Horton, Spargo, Wells, Keene and Moyer2013; Keene, Reference Keene2015). To employ this technique in an organic cover crop-based reduced tillage system (hereafter referred to as OrgCCRT), a grower would strategically use primary tillage for weed control or fertility management, while relying on other management practices to assist in suppression of weeds and as a replacement for the tillage events removed from the system (Mirsky et al., Reference Mirsky, Ryan, Curran, Teasdale, Maul, Spargo, Moyer, Grantham, Weber, Way and Camargo2012). Specifically, annual cash crops are no-till planted into the residues of a high-biomass cover crop established in tilled ground and terminated by a roller-crimper, which produces a mulch layer capable of suppressing weeds during the cash crop growing season (Davis, Reference Davis2010; Mischler et al., Reference Mischler, Curran, Duiker and Hyde2010a; Ward et al., Reference Ward, Ryan, Curran, Barbercheck and Mortensen2011; Mirsky et al., Reference Mirsky, Ryan, Curran, Teasdale, Maul, Spargo, Moyer, Grantham, Weber, Way and Camargo2012). The number of primary tillage events in OrgCCRT can be reduced by half or more in comparison with those managed with multiple annual tillage and cultivation events, and organic growers may gain soil quality benefits without allowing the weed seedbank to increase (Menalled et al., Reference Menalled, Gross and Hammond2001; Davis et al., Reference Davis, Renner and Gross2005).
The efficacy of OrgCCRT as a pest management strategy is a function of cover crop performance and timing of management practices, such as cover crop termination and inversion tillage if implemented (Mischler et al., Reference Mischler, Curran, Duiker and Hyde2010a; Smith et al., Reference Smith, Barbercheck, Mortensen, Hyde and Hulting2011; Ward et al., Reference Ward, Ryan, Curran, Barbercheck and Mortensen2011). Depending on the location, 5000–10,000 kg ha−1 of rolled cover crop dry-matter biomass is necessary to effectively suppress weeds, but biomass production and residue persistence is dependent on cover crop species, planting date and termination date (Mirsky et al., Reference Mirsky, Curran, Mortensen, Ryan and Shumway2009, Reference Mirsky, Curran, Mortensen, Ryan and Shumway2011; Davis, Reference Davis2010; Mischler et al., Reference Mischler, Curran, Duiker and Hyde2010a; Nord et al., Reference Nord, Ryan, Curran, Mortensen and Mirsky2012). Additionally, research has demonstrated that cover crop phenology determines termination success; the cover crop must be mature enough to prevent regrowth following termination by the roller-crimper, but not so mature as to set seed following termination, which would result in volunteer cover crops (Mirsky et al., Reference Mirsky, Curran, Mortensen, Ryan and Shumway2009; Davis, Reference Davis2010; Mischler et al., Reference Mischler, Curran, Duiker and Hyde2010a).
Due to the properties of the rolled cover crop mulch and a reduction in soil disturbance, OrgCCRT may provide diverse resources in space and time for ground dwelling natural enemies, including carabids (Mathews et al., Reference Mathews, Bottrell and Brown2004; Shearin et al., Reference Shearin, Chris Reberg-Horton and Gallandt2008; Blubaugh and Kaplan, Reference Blubaugh and Kaplan2015). The addition of a winter cover crop may ameliorate the microclimate at the soil surface, buffering against extremes in temperature and relative humidity, providing a more favorable microclimate to attract and retain carabids (Shearin et al., Reference Shearin, Chris Reberg-Horton and Gallandt2008; Diehl et al., Reference Diehl, Wolters and Birkhofer2012). This approach does not eliminate tillage, but it does reduce the number of tillage passes and the intensity (depth) of frequent tillage, thus reducing the potential for killing carabids through burial or forcing frequent outmigration (Thorbek and Bilde, Reference Thorbek and Bilde2004; Hatten et al., Reference Hatten, Bosque-Pérez, Johnson-Maynard and Eigenbrode2007). Due to the elimination of chemical insect management, organically managed agroecosystems are also likely to have higher abundance and diversity of many arthropods, some of which may not be pests in agroecosystems but could serve as prey for carnivorous carabids (Döring and Kromp, Reference Döring and Kromp2003; Bengtsson et al., Reference Bengtsson, Ahnström and Weibull2005; Purtauf et al., Reference Purtauf, Roschewitz, Dauber, Thies, Tscharntke and Wolters2005; Menalled et al., Reference Menalled, Smith, Dauer and Fox2007; Birkhofer et al., Reference Birkhofer, Wise and Scheu2008). Additionally, because of the weed management challenges in organic systems, the density and biomass of weeds frequently increases through time as compared with conventionally managed systems, although not always with detrimental effects on yield nor with an increase in the weed seedbank (Menalled et al., Reference Menalled, Gross and Hammond2001; Bàrberi, Reference Bàrberi2002; Ryan et al., Reference Ryan, Mortensen, Bastiaans, Teasdale, Mirsky, Curran, Seidel, Wilson and Hepperly2010). Because many carabids are omnivorous or granivorous with the potential to feed on weed seeds, some species benefit from the additional availability of food resources provided by weeds, and may in fact contribute to maintaining a stable weed seedbank (Menalled et al., Reference Menalled, Gross and Hammond2001, Reference Menalled, Smith, Dauer and Fox2007; Larochelle and Larivière, Reference Larochelle and Larivière2003; Diehl et al., Reference Diehl, Wolters and Birkhofer2012). Thus, employing reduced tillage may contribute to conservation of carabids through the provision of a stable microclimate and ample prey resources.
The OrgCCRT approach to weed management has strong potential, especially for organically-managed agroecosystems, for conserving and augmenting populations of carabid beetles (Döring and Kromp, Reference Döring and Kromp2003; Purtauf et al., Reference Purtauf, Roschewitz, Dauber, Thies, Tscharntke and Wolters2005; Menalled et al., Reference Menalled, Smith, Dauer and Fox2007). However, the crop sequence employed during the transition can have long-term implications for pest control potential, as certain crops may be more efficient in attracting and retaining carabids (Lundgren et al., Reference Lundgren, Shaw, Zaborski and Eastman2006). Likewise, the timing of management practices, including cover crop termination date, can affect the amount and availability of resources in the habitat (Blubaugh and Kaplan, Reference Blubaugh and Kaplan2015). Here we report on a study that examined the interacting effects of initial cash crop and timing of management practices on the carabid community in a feed and forage cropping system employing OrgCCRT in central Pennsylvania, USA. We hypothesized that over the course of the 3-year transition to organic management: (1) activity-density and diversity of carabid beetles will increase in each cover crop with each additional year of organic management; (2) in each cover crop, carabid community composition will shift from a dominance by small, omnivorous species, to larger, granivorous and carnivorous species; and (3) cover crop species, timing of termination and in-crop cultivation strategy will affect carabid community composition.
Materials and Methods
Site
This research was conducted at the Russell E. Larson Agricultural Research Center in Centre County, Pennsylvania (40°43′23″N, 77°55′44″W) at 376 m above sea level (USGS, 2014), with a May–October growing season. During the 3 years of the experiment, average annual precipitation varied, with 1409, 1044 and 1003 mm in years 1–3, respectively (data measured on site) (Keene, Reference Keene2015), with a mean annual precipitation of 1006 mm between 1981 and 2010 (NOAA, 2014). Mean monthly temperatures ranged between −2.7 and 22.3°C (Celsius), with an annual mean of 10.1°C in the years 1981–2010 (NOAA, 2014). The experiment was part of a larger study, the Reduced-Tillage Organic Systems Experiment, hereafter ROSE, which was replicated at three sites in the mid-Atlantic states of Pennsylvania, Maryland and Delaware; however, we only discuss results from the ROSE conducted in Pennsylvania. The total area of the site was approximately 4 ha, and during the experiment, was managed for transition to certified organic production. Soils at the site are representative of the Hagerstown Soil Series according to the USDA, Natural Resources Conservation Service soil classification system, a silt loam classified as prime farmland (Soil Survey Staff, 2014).
Experimental design and crop management
The complete cash crop and cover crop rotation during the 3-year experiment proceeded in the following sequence: a cereal rye cover crop, soybean, wheat, a hairy vetch and triticale cover crop and a cash crop of corn for silage (Supplementary Table 1). We employed four repetitions (blocks) in the experiment (6020 m2 per block), and each of the four blocks contained three crop strips with an area of 2006 m2 each, to include one crop in the 3-year rotation to allow for a full entry design, with each phase of the rotation present in every year of the experiment (Supplementary Fig. 1). In each cropping strip, field operations would proceed as follows, depending on the phase of the rotation (with some variations depending on year, Supplementary Table 1): Seedbed preparation for cover crops and winter wheat included inversion tillage with a four-bottom moldboard plow, disking and two cultimulch passes. Cereal rye was seeded into the tilled seedbed following corn in mid- to late-October at 189 kg seed ha−1, then terminated with two roller-crimper passes spaced approximately 7 days apart in late-May and early-June. Soybean was no-till planted immediately following the second roller-crimper pass of cereal rye, at a rate of 556,000 seeds ha−1. After soybean harvest in mid- to late-October, winter wheat was seeded at a rate of 163 kg seed ha−1 into the tilled soil. Wheat was harvested as a cash crop each year in mid-July, and a mixture of hairy vetch and triticale was seeded into the tilled seedbed at a rate of 34 kg seed ha−1 each in late-August. The hairy vetch and triticale were managed with two passes of the roller-crimper approximately 7 days apart in late-May through late-July, and corn was planted after the first roller crimper pass at a rate of 84,000 seeds ha−1. Corn was harvested in October, and the soil tilled in anticipation of planting cereal rye.
Within each cropping strip, treatments were imposed using a split-split plot design to determine the effect on the carabid community of cover crop termination timing and in-crop cultivation strategy in the corn and soybean phases of the rotation. At the main plot level in these two crops, treatments consisted of three different dates of cover crop rolling and termination, (Early, Middle and Late) relative to a typical corn and soybean planting date used by organic growers in the region, and based on cover crop phenology (Mischler et al., Reference Mischler, Duiker, Curran and Wilson2010b; Nord et al., Reference Nord, Ryan, Curran, Mortensen and Mirsky2012). These termination dates effectively control the amount and quality of cover crop biomass on the soil surface, as the cover crop was terminated prior to or at the time of cash crop planting, regardless of cover crop stage. As such, cover crops managed later may have amassed more biomass at the soil surface, or in the case of hairy vetch, which flowers in the spring, a later termination date may result in more flowers, which could attract alternative prey items for predatory carabids (Mischler et al., Reference Mischler, Duiker, Curran and Wilson2010b; Mirsky et al., Reference Mirsky, Curran, Mortensen, Ryan and Shumway2011; Eisenhauer and Reich, Reference Eisenhauer and Reich2012; Heil, Reference Heil2015; Keene, Reference Keene2015). In the first year of the experiment, the cover crops were only managed once with the roller-crimper, which did not result in adequate control and a second rolling was added in years 2 and 3. In the early termination dates in all 3 years of the experiment, cover crop regrowth was common due to a lack of complete control by the roller-crimper (Keene, Reference Keene2015).
At the split-plot level, the use of high residue inter-row cultivation was compared with an alternative in-crop weed management strategy in corn and soybean (with plots of 167 m2 per cultivation treatment). The high residue cultivator is equipped with a no-till coulter to cut the residue, which is followed by a single 50 cm wide sweep to sever emerged weeds while leaving the surface residue intact. In the corn phase of the rotation, high residue inter-row cultivation was compared with a control treatment, which relied solely on the rolled cover crop mulch for within-season weed control. We planted all corn cultivation treatments in 76 cm rows. In the soybean phase, high residue cultivation was utilized in 76 cm row soybean and was compared with soybean planted in 38 cm rows that did not utilize high residue cultivation, but relied on row spacing and cereal rye residue for within-season weed control. Depending on the crop and year, high residue cultivation occurred approximately 4–6 weeks after planting and utilized two passes spaced approximately 2–7 days apart (Supplementary Table 1).
In the first year of the experiment, we employed an additional split to divide each cropping strip to compare an expressive weed management strategy, using a tine weeder, to a control treatment, dividing the final plots into 9.1 × 9.1 m2. After the first year, this treatment was dropped for lack of efficacy, and preliminary analyses indicated no lasting effect of this treatment, so we considered the 9.1 × 9.1 m2 plots independent due to the cultivation employed in the first year.
Arthropod sampling
To characterize the carabid community, we deployed one pitfall trap simultaneously in each 9.1 × 9.1 m2 plot, timed to occur approximately 1 week after the emergence of corn and soybean. This resulted in different pitfall sampling dates for each cover crop termination date treatment, with sampling occurring approximately 2–3 weeks after the second rolling of each cover crop (Supplementary Table 1). At the time of trapping in the rolled cover crops, the soil was completely covered by a dense mat of cover crop residue created by the roller-crimper. We moved the mat to expose the soil to place the pitfall trap, and then returned the mat to natural conditions after the trap was placed. In the treatments planted with hairy vetch and triticale (hereafter abbreviated as HVT), hairy vetch represented the bulk of the mulch biomass, with the hairy vetch stems creating a tangled mat of dense residue, which could be easily lifted off the soil surface as a single piece. In the cereal rye treatments, the cereal rye stems were rolled parallel to the crop rows, with each stem aligned in the same direction relative to the others. Both corn and soybean were at growth stage V1 at the time of pitfall sampling (Nordby, Reference Nordby2004; Nafziger, Reference Nafziger2009), with average heights of 13 cm in soybean and 21 cm in corn. We conducted a single pitfall-trapping event approximately 1 month prior to wheat harvest, regardless of the termination date treatments of the crops preceding wheat. The standing wheat crop was at Feekes growth stage 11 (Nafziger, Reference Nafziger2009), with an average height of 68 cm at the time of pitfall sampling.
In each plot, the pitfall traps were placed approximately 6 m from the outer, drive-row edge, of the plot and 3 m from the southern edge of the plot (Supplementary Fig. 1), and between crop rows. Each trap consisted of a 1-L plastic deli container buried level with the soil surface into which we placed a 50 mL plastic specimen cup, filled with 30 mL ethylene glycol as a killing agent and preservative. Inside the opening of the deli container, we placed a funnel (114 mm in diameter) to facilitate insect movement into the specimen cup and to exclude larger animals from entering the trap (Weeks and McIntyre, Reference Weeks and McIntyre1997). Traps remained open for 72 h, after which the samples were removed from the field and brought to a laboratory for processing and identification.
In the laboratory, we removed all Carabidae adults from the trap, preserved them in 80% ethanol, and counted and identified them to species using identification keys in Bosquet (Reference Bosquet2010) and local reference collections. Two species within the genera Amara, Amara impuncticollis Say and Amara littoralis Mannerheim, are difficult to identify to species without dissections, and we included these two species at the higher taxonomic level of Amara impuncticollis group, with the group considered as a single species due to similarities in morpholgical and behavioral characteristics (Larochelle and Larivière, Reference Larochelle and Larivière2003; Bosquet, Reference Bosquet2010). Information regarding the ecology, behavior, phenology and size of the adults of each species was collected from various sources (Larochelle and Larivière, Reference Larochelle and Larivière2003; Lundgren, Reference Lundgren2009; Bosquet, Reference Bosquet2010; Bohan et al., Reference Bohan, Caron-Lormier, Muggleton, Raybould and Tamaddoni-Nezhad2011; Dearborn et al., Reference Dearborn, Nelson, Donahue, Bell and Webster2014). We classified adult carabids into trophic groups according to their predominant feeding preferences described in the literature, by the following designations: carnivorous, feeding primarily on animal tissues; omnivorous, feeding on both animal and plant tissues; and herbivorous, feeding primarily on plant materials, including seeds (Lundgren, Reference Lundgren2009). Size classes were assigned as follows: small, <5 mm; medium, between 5 and 10 mm; and large, >10 mm (Eyre et al., Reference Eyre, Luff, Atlihan and Leifert2012).
Statistical analyses
Mean activity-density (the relative measure of presence and movement of arthropods captured by pitfall traps), species richness, Smith-Wilson evenness, and proportions of each trophic group and size class of adult carabids were analyzed using a general linear mixed effects two-way analysis of variance (ANOVA) model using PROC GLIMMIX in SAS version 9.3 (Lang, Reference Lang2000; SAS Institute, 2014). All activity-densities were log10 (x + 1) and all proportions were arcsine square root transformed to meet assumptions of normality and homogeneity of variances prior to use in all analyses conducted in PROC GLIMMIX, but we present untransformed data throughout this paper (Gotelli and Ellison, Reference Gotelli and Ellison2004; Kutner et al., Reference Kutner, Nachtsheim, Neter and Li2005; Ives, Reference Ives2015). We examined nine combinations of cover crop treatment and year (crop-year, e.g., wheat 2011) and the six combinations of the termination date treatment and high residue cultivation treatment (e.g., early-no cultivation) as single fixed effects. Random effects were specified as an overall residual effect, a block by year variance component and a block by initial crop variance component; we specified a Poisson distribution for count data, and a normal distribution for proportions, according to the recommended standards for use in mixed models (Gotelli and Ellison, Reference Gotelli and Ellison2004; Kutner et al., Reference Kutner, Nachtsheim, Neter and Li2005; Bolker et al., Reference Bolker, Brooks, Clark, Geange, Poulsen, Stevens and White2009; Ives, Reference Ives2015). We used lsmeans statements using the Tukey method to conduct post hoc tests of means to isolate differences within year, and within the crop, cover crop termination date and high residue cultivation treatments.
To describe patterns within the entire carabid community, we compared expected species richness by crop and pooled across the 3 years using rarefaction in the BiodiversityR package in R (Kindt and Coe, Reference Kindt and Coe2005; Kutner et al., Reference Kutner, Nachtsheim, Neter and Li2005; R Core Team, 2013). Using the random method, by which sites are added in a random order, the smoothed rarefaction curves allow for comparison of the number of species expected at a given site for each crop. This allows for comparisons between crops based on the number of plots (sites) sampled, and significance was determined by non-overlapping confidence intervals (Gotelli and Colwell, Reference Gotelli and Colwell2001).
We used principal response curves (PRC) using the prc function in the vegan package of R (a package designed specifically for analyzing ecological communities) to determine community responses through time during the experiment (van den Brink and ter Braak, Reference van den Brink and ter Braak1999; R Core Team, 2013; Oksanen et al., Reference Oksanen, Blanchet, Kindt, Legendre, Minchin, O'Hara, Simpson, Solymos, Stevens and Wagner2015). As a form of constrained redundancy analysis, PRCs allow for the graphical representation of changes in a community through time, in response to specific treatments as compared with a control (van den Brink and ter Braak, Reference van den Brink and ter Braak1999; Borcard et al., Reference Borcard, Gillet and Legendre2011). PRCs calculate individual weights for each species, with a positive or negative weight indicating the response of the species to the treatment or control, with higher absolute values of weights indicating a stronger response to the pattern shown in the PRC (van den Brink and ter Braak, Reference van den Brink and ter Braak1999). To compare changes in the carabid community through the 3 years of the experiment, we conducted PRCs on the main effect of crop treatment, with wheat set as control to characterize differences between the standing crop and the two rolled cover crops. Separately, we analyzed each of the crop treatments individually through the 3-year experiment, by the main effects of high residue cultivation, with no cultivation set as the control, and termination date of the cover crop, with early termination date set as the control. Species were only included in the analyses if they had an activity-density >1% of the total for the subset of the data in each analysis. We selected this threshold to exclude disproportionate effects of rare species (McCune and Grace, Reference McCune and Grace2002). We transformed the species activity-densities using the Hellinger transformation prior to conducting the PRCs (van den Brink and ter Braak, Reference van den Brink and ter Braak1999; Legendre and Gallagher, Reference Legendre and Gallagher2001), and we used a Monte Carlo simulation with 4999 permutations to test the overall significance of each PRC. We present only significant PRCs showing only species with weights greater than or equal to an absolute value of 0.5, which indicates that the pattern of those species through time matches the pattern shown in the PRC. Species with weights close to zero may either have no response to the treatments specified in the PRC, or may have a response that does not match that shown in the PRC (van den Brink and ter Braak, Reference van den Brink and ter Braak1999).
To further isolate associations between carabid species and treatment, we conducted an indicator analysis using the multipatt function in the indicspecies package (De Cáceres and Legendre, Reference De Cáceres and Legendre2009). Multipatt calculates an indicator value (IndVal) for each species, which is the product of the specificity (S) value for each species, the probability that a site with a specific species belongs to the treatment specified, and the fidelity (F) of a species, the probability that a species will be found in sites belonging to a treatment. Associations with a specific treatment are reported based on the highest IndVal for each species; we restricted the analyses to only allow associations with one cover crop by termination and planting date treatment (De Cáceres, Reference De Cáceres2013).
Results
Time in organic management
A total of 1786 individual carabids in 47 species were collected during the 3 years of the experiment and across the experimental site (Table 1), with more than half (56%) of the carabid beetles captured in the final year of the experiment. Bembidion quadrimaculatum oppositum Say, a small, endemic and cosmopolitan beetle, was the most abundant species, accounting for 49% of the total individuals captured at the experimental site (Table 1). The main effect of crop-year was significant for total activity-density (F 8,50 = 29.41, P < 0.0001) and the number of species captured (richness) (F 8,51 = 27.42, P < 0.0001). Both total activity-density and richness consistently increased during the 3 years of the experiment according to post hoc tests of means (P < 0.01, Table 2). The main effect of crop-year was also significant for Smith-Wilson evenness (F 8,66 = 5.15, P < 0.0001), with post hoc tests of means indicating a higher mean evenness-density in the third year as compared with years one (t = 4.66, P = 0.00) and two (t = 2.56, P = 0.03) of the experiment. The mean size of carabid beetles shifted over time (Table 2), with the proportion of medium (5–10 mm) carabids significant at the main effect of crop-year (F 8,69 = 2.58, P = 0.02). A higher proportion of medium carabids were captured in the third year compared with the first year (t = 9.416, P = 0.02). There were no significant changes in the proportion of carabid beetles by trophic group during the three years of the experiment.
Table 1. Total activity-densities by year of ground beetle species representing greater than 1% of total captures, during the 3-year experiment.
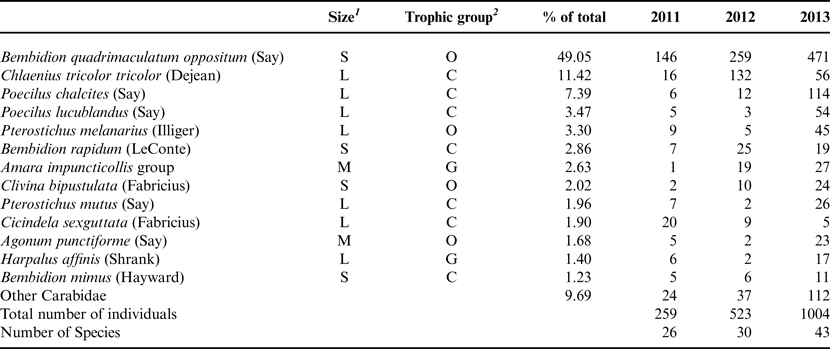
1 Size classes: S = small (0–5 mm); M = medium (5–10 mm); L = large (>10 mm).
2 Trophic groups: C = mostly carnivorous; G = mostly granivorous; O = mostly omnivorous.
Data were pooled across treatments for each year (n = 144).
Table 2. Annual mean of assessments related to the Carabidae community (± SEM) across treatments.
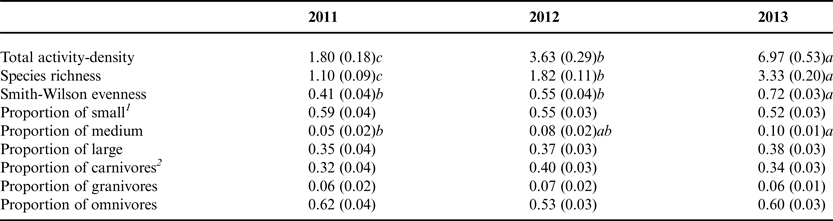
1 Size classes: S = small (0–5 mm); M = medium (5–10 mm); L = large (>10 mm).
2 Trophic groups: Carnivores = mostly carnivorous; Omnivores = mostly omnivorous; Granivores = mostly granivorous.
Response variables were tested by mixed effects models, and values with different letters within the same row are significantly different at P < 0.05. Data were pooled across treatments for each year (n = 144), and we present untransformed means.
According to principal response curves, the composition of the carabid community responded to crop treatment across the 3 years of the experiment (Fig. 1, F 1,423 = 27.63, P < 0.001). The compositional changes through time were due to the differences in the principle responses of Bembidion quadrimaculatum oppositum Say (species weight of −1.75) to crop, with this species also having the highest activity-density at the site (Table 1). Time in organic management (year) accounted for 11% and crop treatment accounted for 5% of the variance within the carabid community, respectively. In years 1 and 2 of the experiment, the coefficients for the rolled cover crops were closer to each other than to wheat, indicating that these treatments may be more comparable than either treatment is to wheat (Fig. 1). However, by the third year of the experiment, the rolled cereal rye treatment had diverged from the other two crop treatments, with fewer B. quadrimaculatum oppositum captured in rolled cereal rye in 2013 compared with the other two crop treatments (38, as compared with 224 in HVT, and 209 in wheat).

Figure 1. Principal response curve (PRC) by crop across the 3 years of the experiment, with wheat set as the control treatment (a). Note that axes are on different scales, with the first y-axis representing the canonical coefficient (effect) of the overall PRC, and the second y-axis representing species weights. Only one species had a significant relationship to the principal response (species score ≥ 0.5). HVT, rolled hairy vetch and triticale; Rye, cereal rye.
Cover crop identity and management
More than half of the individual carabid beetles (54%) were collected in the rolled HVT, with 29% and 16% collected in the standing wheat and rolled cereal rye, respectively, during the 3 years of the experiment (Supplementary Table 2). The rarefaction curves for the three crop treatments indicate that we approached an adequate sampling effort with regard to the number of species collected for each of the crop treatments (Supplementary Fig. 2), but that in rolled HVT, fewer sites (traps) would be necessary to obtain any given number of species compared with rolled cereal rye and wheat. Based on overlapping 95% confidence intervals in the rarefaction curves (not shown, Supplementary Fig. 2), the total number of species captured in each crop treatment across the 3 years of the experiment were not significantly different, with 33 in rolled cereal rye, and 34 in both rolled HVT and wheat.
The 3-year mean activity-density and species richness differed significantly among each of the three crop treatments (Table 3), with activity-density significantly higher in rolled HVT than in rolled cereal rye (t = −11.33, P < 0.0001) and wheat (t = −7.25, P < 0.0001) and significantly higher in wheat than rolled cereal rye (t = 4.08, P < 0.0001). The crop treatments also differed in community composition (Supplementary Table 2), as indicated by a significantly higher Smith-Wilson evenness and proportion of medium carabids in rolled HVT as compared with both rolled cereal rye (t = −2.21, P = 0.03) and wheat (t = −3.35, P = 0.00, Table 3), and a significantly higher proportion of carnivores in rolled cereal rye compared with wheat (t = −2.03, P = 0.04, Table 3). This difference in community composition also explains why the total numbers of carabid species captured in each cover crop treatment were comparable (Supplementary Fig. 2), but the mean numbers of species captured in each cover crop treatment were significantly different (Table 3).
Table 3. Plot mean (± SEM) across treatments and years by crop of response variables related to the Carabidae community, and tested by mixed effects models.

1 Size classes: S = small (0–5 mm); M = medium (5–10 mm); L = large (>10 mm).
2 Trophic groups: Carnivores = mostly carnivorous; Omnivores = mostly omnivorous; Granivores = mostly granivorous.
Data were pooled across treatments for each crop, and we show untransformed means (n = 144). Values with different letters in the same row are significantly different at P < 0.05.
HVT, rolled hairy vetch and triticale.
This strong response of the carabid community to crop treatment was also apparent in the differential effect of the cover crop termination date on the accumulated 3-year means within the cover crop treatments (Table 4). In rolled HVT, the 3-year mean activity-density significantly increased through time with later cover crop termination dates, with higher activity densities in the late termination date compared with the middle termination date (t = −3.02, P = 0.00) and early termination date (t = −5.55, P < 0.0001), and middle compared with early (t = −2.53, P = 0.01). However, the proportion of small carabids was significantly lower in the late termination date compared with the middle (t = 2.70, P = 0.01) and early (t = 2.78, P = 0.01), while the proportion of large carabids was significantly higher in the late compared with the middle (t = −3.04, P = 0.00) and early (t = −2.70, P = 0.01) termination dates (Table 4). The proportion of carnivorous carabids was significantly higher in the late compared with the middle termination date in rolled HVT (t = −2.15, P = 0.03).
Table 4. Means (± SEM) for the significant response variables reflecting assessments of the Carabidae community within each crop, tested by mixed effects models.
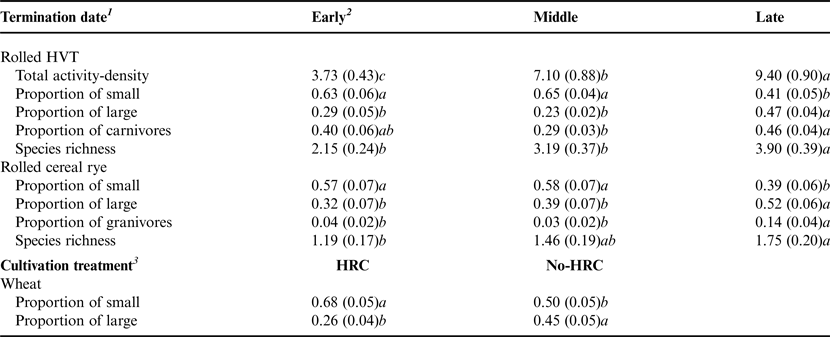
1 n = 48.
2 Early, middle and late reflect cover crop termination dates in anticipation of cash crop planting at standard dates for each crop in central Pennsylvania.
3 n = 72.
Data were pooled across year, and we show untransformed means. Means with different letters in the same row are significantly different at P < 0.05.
HRC, high residue cultivation.
The termination date affected the carabids in rolled cereal rye (Table 4), with a decrease in the proportion of small carabids in the late compared with the middle (t = 2.56, P = 0.01) and early termination dates (t = 2.63, P = 0.01). The proportion of large carabids increased from the early to late termination date (t = −3.00, P = 0.00), and middle to late (t = −2.11, P = 0.04), as did the proportion of granivorous carabids from the early to late (t = −2.31, P = 0.02) and middle to late (t = −2.84, P = 0.01). Species richness increased with the late compared with the early termination date in rolled cereal rye (t = −4.22, P < 0.0001). Only in wheat was the effect of high residue cultivation significant; because of the experimental design, wheat, by the third year of the experiment, was the only crop to have followed two crops in which high residue cultivation had occurred. In wheat, the proportion of small carabids was significantly higher (t = −2.80, P = 0.01), and the proportion of large carabids was significantly lower (t = 2.97, P = 0.00) in treatments which had previously received cultivation compared with treatments which had not previously received cultivation.
Within each termination date treatment, only the principal response of rolled HVT was significant (Fig. 2, F 1,135 = 10.09, P < 0.001), indicating that termination date of the rolled HVT cover crop was a significant driver in the composition of the carabid community within this cover crop. Time in organic management accounted for 12% and termination date treatment accounted for 10% of the variance in the community, respectively. Two species had strong responses that corresponded to the PRC: B. quadrimaculatum oppositum (weight = −0.74) and Chlaenius tricolor tricolor Dejean (weight = 1.08). In indicator analysis, both of these species were associated with rolled HVT. B. quadrimaculatum oppositum was associated with the middle termination date in HVT, and C. tricolor tricolor was associated with the late termination date (Table 5). Of the remaining 13 species of carabid beetles showing a significant relationship to a crop and termination date, 10 of those species were associated with rolled HVT (Table 5). While not all the species associated with HVT had high values for specificity (probability that the species was captured in a treatment) to cover crop and termination date, both B. quadrimaculatum oppositum and C. tricolor tricolor had high values for fidelity (probability that a treatment contains a species), e.g., 54% of C. tricolor tricolor were captured in late HVT (specificity), and 67% of the traps included that species (fidelity).

Figure 2. Principal response curve (PRC) for each year in rolled hairy vetch and triticale (HVT) by termination date treatment (early, middle or late relative to standard cover crop termination dates in central Pennsylvania), with the early termination date set as control. Note that axes are on different scales, with the first y-axis representing the canonical coefficient (effect) of the overall PRC, and the second y-axis representing species weights. Only two species had a response that matches the PRC (species score ≥ 0.5).
Table 5. Significant indicator species for each crop and termination date, pooled across years, according to the multipatt function in indicspecies package of R, with the method restricted to selecting only one treatment per species.

1 Size classes: S = small (0–5 mm); M = medium (5–10 mm); L = large (>10 mm).
2 Trophic groups: Carnivores = mostly carnivorous; Omnivores = mostly omnivorous; Granivores = mostly granivorous.
3 Specificity (S) is the probability that a site with a specific species belongs to the treatment specified; fidelity (F) is the probability that a species will be found in sites belonging to that treatment (e.g., 100% of Cicindela punctulata were found in late cereal rye, but only 6.25% of the sites contained the species).
HVT = rolled hairy vetch and triticale; Rye, rolled cereal rye; Ind. Val statistic, indicator value, the product of specificity and fidelity.
Discussion
During the transition to certified organic production, cultural practices may determine the potential for successful pest management, as certain practices may conserve and augment populations of carabid beetles (Lundgren et al., Reference Lundgren, Shaw, Zaborski and Eastman2006). We hypothesized that carabid activity-density and diversity would increase during a 3-year transition to organic management at a site at which insect management previously included insecticides. At our site, carabid beetle activity-density, species richness, and community evenness increased during the transition period. We observed significant increases in the proportion of medium-sized beetles, but no significant changes in the proportion of each trophic group of the beetles. As expected, management practices during the organic transition influenced carabid communities. The cover crop species used preceding cash crops and cover crop termination timing influenced carabid community composition. We also observed legacy effects of high residue cultivation on carabid community composition in wheat. High residue cultivation occurred in the corn and soybean preceding wheat, and resulted in a significantly higher proportion of smaller carabids in the cultivated treatments as compared with uncultivated, likely due to the potential for small organisms to better tolerate soil disturbances (Szysko et al., Reference Szysko, Vermeulen, Klimaszewski, Abs, Schwerk, Brandmayr, Lövei, Brandmayr, Casele and Taglianti2000; Holland, Reference Holland2004). Our results confirm those observed in other studies, in that specific management factors, including the addition of cover crops in a rotation, the use of mulch and the intermittent use of tillage, and the timing and type of management factors, can affect carabid activity-density and diversity (Thorbek and Bilde, Reference Thorbek and Bilde2004; Clark et al., Reference Clark, Szlavecz, Cavigelli and Purrington2006; Shearin et al., Reference Shearin, Chris Reberg-Horton and Gallandt2008; Renkema et al., Reference Renkema, Lynch, Cutler, Mackenzie and Walde2012; Russon and Woltz, Reference Russon and Woltz2014). Minimal research exists, however, on the effect of this high residue environment within the organic transition on carabid beetles in cropping systems employing organic cover crop-based reduced tillage (OrgCCRT), and on the timing of management practices during the transition; the results we present here are novel in that regard.
We identified a significant trend of increased activity-density and species richness in each additional year of organic management. Other researchers have suggested that at least 4 years are necessary for arthropod populations to stabilize to any changes in environmental conditions (Sabais et al., Reference Sabais, Scheu and Eisenhauer2011). Increases in species richness and activity-densities of generalist predators, and carabids specifically, in organic agriculture have been attributed to fewer in-field activities, such as cessation of application of synthetic agrochemicals, higher soil quality and resultant soil biodiversity, and increases in weed abundance leading to greater diversity of microclimate, habitat, seed and arthropod prey resources (Kielty et al., Reference Kielty, Allen-Williams, Underwood and Eastwood1996; Birkhofer et al., Reference Birkhofer, Wise and Scheu2008; Guy et al., Reference Guy, Bohan, Powers and Reynolds2008; Frank et al., Reference Frank, Shrewsbury and Denno2011; Diehl et al., Reference Diehl, Wolters and Birkhofer2012). However, carabids in our experiment responded differently to each crop treatment during the transition, as indicated by the principal response curves, and the type and timing of management activities preceding pitfall trapping in each of the crops may play a significant role in our results (Cole et al., Reference Cole, McCracken, Dennis, Downie, Griffin, Foster, Murphy and Waterhouse2002; Lundgren et al., Reference Lundgren, Shaw, Zaborski and Eastman2006; Lundgren and Fergen, Reference Lundgren and Fergen2011; Blubaugh and Kaplan, Reference Blubaugh and Kaplan2015).
In a study to evaluate weed management tactics during the organic transition in fresh-market tomatoes, Blubaugh and Kaplan (Reference Blubaugh and Kaplan2015) identified a significantly higher seasonal activity-density of Harpalus pensylvanicus Dejean adults, an important weed-seed predator, in no-till treatments with a cereal rye cover crop managed by a roller-crimper than in treatments with tillage or with tillage and a living mulch of crimson clover (Trifolium incarnatum L.). The authors suggest that adult female Harpalus spp., generally fall-breeders, prefer to oviposit in sites that have not been tilled for at least one growing season and with abundant plant cover. This suggestion was supported by significantly higher captures of Harpalini larvae where weed cover, and thus potential food resources, was abundant (Blubaugh and Kaplan, Reference Blubaugh and Kaplan2015). Cover crops were planted into tilled ground in our experiment, but among our treatments, the HVT treatment in our system is planted earliest in the fall (early September, Supplementary Table 1), after which no disturbances occurred in the field until spring. Cereal rye and wheat are both planted later than HVT, in mid- to late October. Considering that many fall-breeding carabids are active as early as August and carabid activity has been known to decrease by October in some regions (Carmona and Landis, Reference Carmona and Landis1999; Shearin et al., Reference Shearin, Chris Reberg-Horton and Gallandt2008), this later disturbance and shorter period for crop growth prior to winter may limit the amount of ground cover available as a resource to carabids in the fall (Blubaugh and Kaplan, Reference Blubaugh and Kaplan2015). While the growth habits of cereal rye and wheat are likely to be similar in the fall, cereal rye was subjected to several more field disturbances in the spring prior to pitfall trapping, including two rolling events. Because the HVT treatment is also subject to rolling in the spring, the earlier establishment of the HVT in the fall may play a significant role in attracting and supporting carabids prior to winter.
By the third year of our experiment, the composition of the carabid community was more similar between the rolled HVT treatments and wheat than between the two rolled cover crop treatments (Fig. 1). This indicates that the differences, or similarities, within the carabid communities between crops may be a function of the individual carabid species themselves, rather than characteristics of the habitat. Döring and Kromp (Reference Döring and Kromp2003) make a similar suggestion in central European cropping systems, and propose that breeding season (e.g., spring or fall), wing dimophism and humidity preference will in part determine a species' association with either an organic or conventional system. These authors propose, in part, that macropterous species with high dispersal power and spring breeders with an adult autumn population benefit from organic management. Two of the species for which we observed increases in activity-density from year 1–3 of our experiment, Poecilus chalcites (Say) and Poecilus lucublandus (Say) (Table 1), meet these criteria, in that both are spring breeders and macropterous and submacropterous, respectively (Larochelle and Larivière, Reference Larochelle and Larivière2003). Pitfall traps are somewhat biased toward species that are mobile during the time trapping takes places, which in our case, is spring breeders (Lang, Reference Lang2000).
We conducted pitfall trapping on different calendar dates in order to draw inferences regarding which carabids may be present and able to contribute to biological control when the cash crop was in an early, and vulnerable, vegetative state. That we only conducted one pitfall trapping event per treatment per year is a limitation of our study, but our results are still indicative of strong associations between certain carabid species and specific cover crops and cover crop management date. Because we did not capture many larval carabids, we did not identify these to species, and thus we cannot exclude from our results that particular species might have been present in juvenile stages at different time points in our system or that adults might have been present in specific treatments at different times (e.g., B. quadrimaculatum oppositum could have also been abundant in the early and late treatments on the same date as the middle, if we sampled these all at the same time). However, individual carabid species will respond differently to various stimuli in the environment, as well as to disturbances such as the rolling of the cover crops, which is likely why we observed such strong associations between certain species and specific termination dates of our cover crops (Kielty et al., Reference Kielty, Allen-Williams, Underwood and Eastwood1996; Döring and Kromp, Reference Döring and Kromp2003; Larochelle and Larivière, Reference Larochelle and Larivière2003; Thorbek and Bilde, Reference Thorbek and Bilde2004; Shearin et al., Reference Shearin, Chris Reberg-Horton and Gallandt2008; Eyre et al., Reference Eyre, Luff, Atlihan and Leifert2012). Additionally, OrgCCRT is specifically designed to meet the pest management needs of organic growers, and our results can help support pest management decisions (Mirsky et al., Reference Mirsky, Ryan, Teasdale, Curran, Reberg-Horton, Spargo, Wells, Keene and Moyer2013). The late termination dates of our cover crops (specifically HVT), are strongly associated with a higher activity-density and diversity of beetles, as well as a higher proportion of carnivorous beetles. Assuming a more abundant and diverse carabid community will be more resilient and able to respond to immediate pest pressures in the system, this termination date may provide growers with the opportunity to maximize their pest control potential while promoting the conservation of these important predators (Hatten et al., Reference Hatten, Bosque-Pérez, Johnson-Maynard and Eigenbrode2007; Letourneau and Bothwell, Reference Letourneau and Bothwell2008; Crowder and Jabbour, Reference Crowder and Jabbour2014; Lundgren and Fausti, Reference Lundgren and Fausti2015).
Balancing the risks and benefits associated with cropping system and pest management choices during the transition to organic management is essential for growers. A low intensity system, which may require fewer inputs in time, fuel and labor, while maximizing the long-term biological control potential of the system, could be beneficial for growers during the transition (Delate and Cambardella, Reference Delate and Cambardella2004; Smith et al., Reference Smith, Barbercheck, Mortensen, Hyde and Hulting2011). Employing OrgCCRT in our system, we identified significant differences associated with cover crop species and management during the 3-year transition to organic certification, but to fully understand the benefits of cover crops to carabid conservation and pest management, it would be worthwhile to lengthen the experimental time evaluating OrgCCRT. Thus, continuing the rotation beyond 3 years could provide additional information regarding the key factors influencing the carabid associations with each cover crop (e.g., fall planting date). Additionally, understanding the significant characteristics of each cover crop as it relates to the carabid community (e.g., amount of biomass in the fall, differences in microhabitat complexity, availability of prey associated with each crop, etc.) could provide additional information regarding the ways in which OrgCCRT may be manipulated to further conserve carabids for pest management.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S1742170516000466.
Acknowledgements
This research was supported by a grant through the United States Department of Agriculture, Organic Research and Education Initiative. A. Rivers also received a graduate student grant through the Northeast Sustainable Agriculture Research and Education program to supplement the main research conducted in the experimental site. The authors wish to thank Robert Davidson, Carnegie Museum of Natural History, Pittsburgh, Pennsylvania for confirming Carabidae identifications, Bryan Vinyard, USDA-ARS, for development of the statistical model in SAS, Mark Dempsey and Clair Keene for support with data aggregation and technical support, and Drs John Tooker, Bill Curran, Ed Rajotte and Ebony Murrell for suggestions on an earlier version of the manuscript.









