I. INTRODUCTION
The bone tissue of vertebrates is considered to be a composite material formed by inorganic apatite crystallites embedded in an organic matrix, having a hierarchical structure composed of different structural units at different size scales (Landis, Reference Landis1995). It is generally recognized that the microstructure of fossil bones is quite well preserved (e.g. Plet et al., Reference Plet, Grice, Pagès, Verrall, Coolen, Ruebsam, Rickard and Schwark2017).
The Kotelnich vertebrate fossil location (the district of Kotelnich, Russia), characterized by the excellent preservation of fossil remains, is one of the richest sites of the Permian period. The unique skeletal integrity of Permian tetrapods preserved here is explained by the conditions of their burial in a silty anaerobic environment similar to modern bogs. The Kotelnich vertebrate site extends for more than 30 km along the eastern bank of the Vyatka River (from the city of Kotelnich to the village of Vishkil, Kirov region, Russia, Fig. 1). The sediments containing the burials of large parareptiles and theromorph reptiles belong to the upper part of the Tatarian Stage comprising the most recent Permian subdivision of the Russian plate. According to the generally accepted geological timescale, the age of these deposits is estimated at around 250 million years.
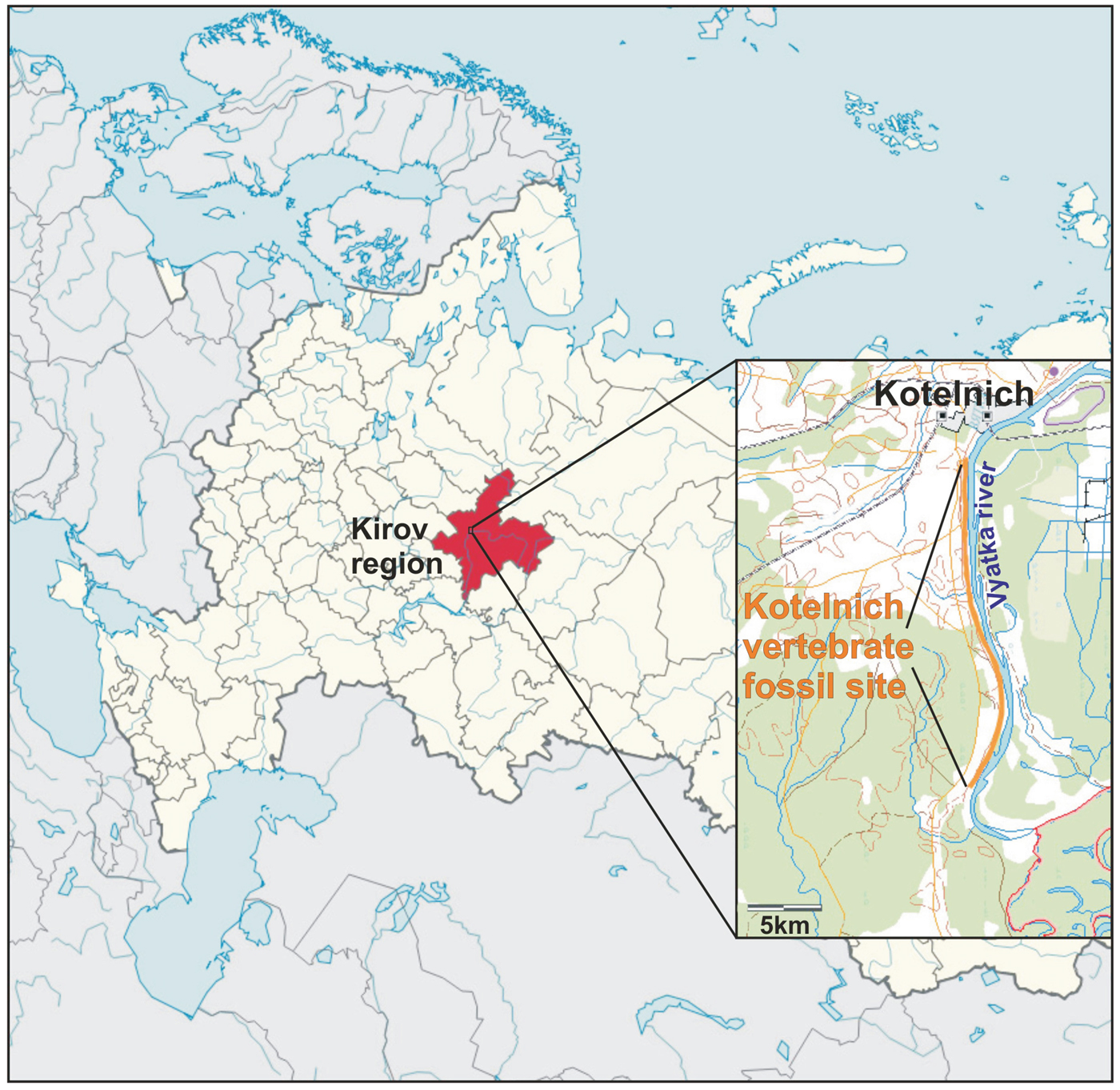
Figure 1. (Color online) Geographic location of the Kotelnich vertebrate fossil site.
The unique richness of the location is attested in the discovery and scientific description of 18 new species of animals during the period from 1933 to the present (Ivakhnenko, Reference Ivakhnenko2001). The main representatives of the Kotelnich fauna are herbivorous pareiasaurs (Deltavjatia vjatkensis).
Despite the large number of publications devoted to the studies of bone and dental remains of Cretaceous and Jurassic dinosaurs (e.g. Chipera and Bish, Reference Chipera and Bish1990; Elorza et al., Reference Elorza, Astibia, Murelaga and Pereda-Suberbiola1999; Piga et al., Reference Piga, Santos-Cubedo, Brunetti, Piccinini, Malgosa, Napolitano and Enzo2011; Kowal-Linka et al., Reference Kowal-Linka, Jochum and Surmik2014), there has been very little work carried out on the study of fossil reptiles of the Permian period. The aim of the present work is to carry out an X-ray diffraction (XRD) study of bone phase composition and structural features of D. vjatkensis pareiasaurus, a Permian parareptile.
II. EXPERIMENTAL
Two D. vjatkensis pareiasaur rib fragments (1.5 × 2.5 cm) were studied. The samples were collected during the excavations by Natural History Museum of Tatarstan and Institute of Geology and Geochemistry, UB RAS during the joint field trip to the “Kotelnich location of pareiasaurs” palaeontological reserve.
The pareiasaur rib fragments (sample mass 500 mg) were powdered manually in a jasper mortar and analyzed by XRD using a Shimadzu XRD-7000 powder X-ray diffractometer with СuKα radiation (λ = 1.5406 Å) operating at 40 kV and of 30.0 mA. XRD patterns were collected during 8 h with 0.1° min−1 step across the angular range of 20–70°. The preliminary qualitative phase analysis of the bones was conducted using the ICDD Powder Diffraction File database PDF-2 (ICDD, 2008). To perform the quantitative full profile analysis and unit-cell lattice parameters determination of the bone, digitized X-ray powder profiles were analyzed according to the Rietveld method (Rietveld, Reference Rietveld1967) using the Sietronics SiroQuant software (Taylor, Reference Taylor2004).
III. RESULTS AND DISCUSSION
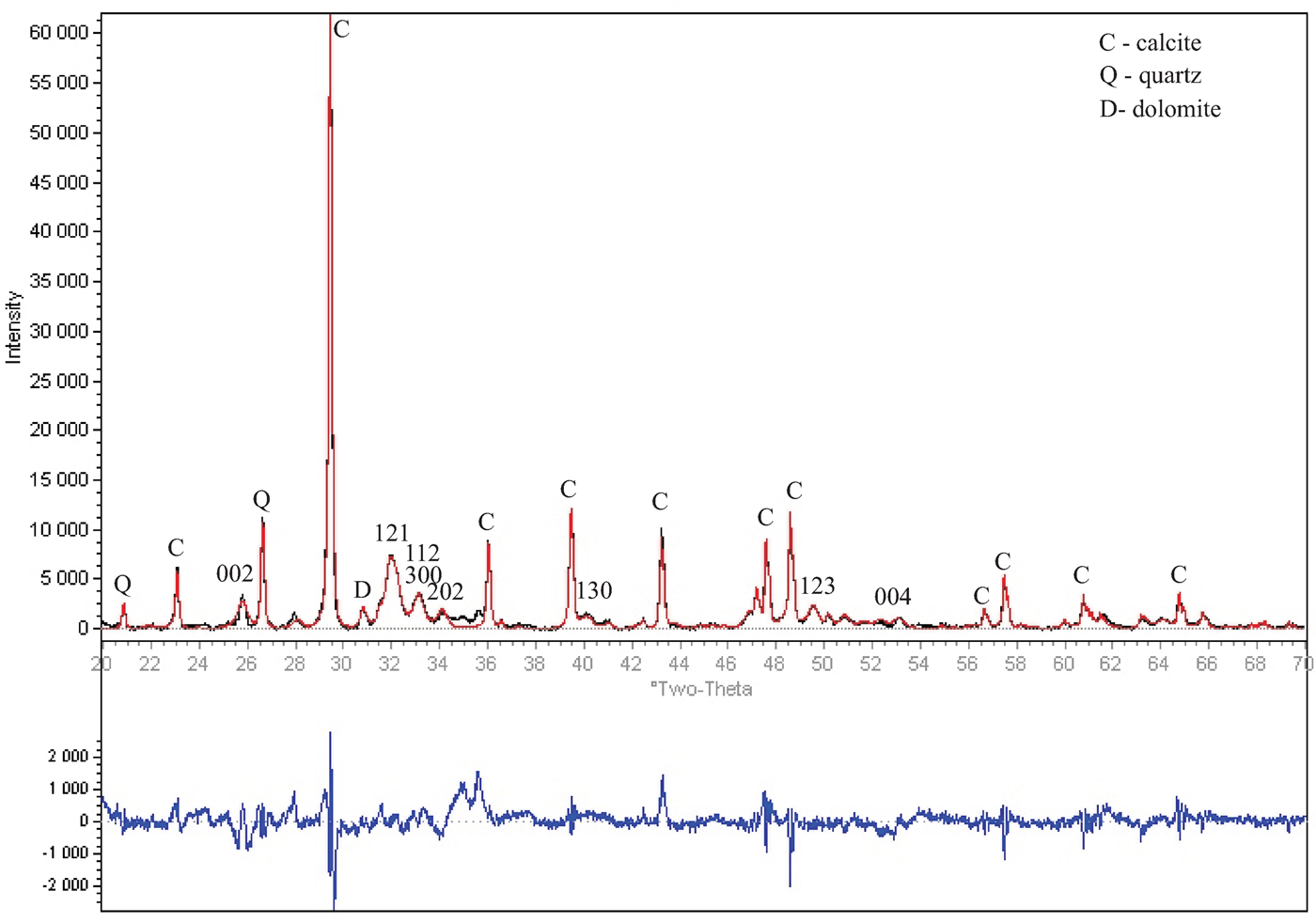
The XRD pattern of a pareiasaur bone fragment is shown in Figure 2. Bone phase composition is defined as calcite (55.1–52.4) %, apatite (44.0–37.2) %, quartz (0.7–6.2) %, and dolomite (0.2–4.2) %. The porous bone can include detrital quartz and dolomite derived from the embedding rock (Silaev et al., Reference Silaev, Ponomarev, Kiseleva, Smoleva, Simakova, Yu, Martirosyan, Vasil'ev, Khazov and Tropnikov2017; Elorza et al., Reference Elorza, Astibia, Murelaga and Pereda-Suberbiola1999). On the contrary, calcite is rather widespread among the fossil apatite deposits as newly-formed mineral (Elorza et al., Reference Elorza, Astibia, Murelaga and Pereda-Suberbiola1999), filling bone pores or depositing on its surface as a thin white crust (Piga et al., Reference Piga, Santos-Cubedo, Brunetti, Piccinini, Malgosa, Napolitano and Enzo2011).

Figure 2. (Color online) Observed (black line), calculated (red line), and difference XRD pattern (blue line, bottom) for pareiasaur bone fragment refined by Rietveld analysis technique [Sietronics SiroQuant (Taylor, Reference Taylor2004)]. Apatite reflections are marked by numbers.
Refined apatite unit-cell parameters of a and b-axis (9.3526 ± 0.0001 and 9.3587 ± 0.0001) Å and c-axis (6.8930 ± 0.0001 and 6.8968 ± 0.0001) Å correspond to carbonate F-apatite (Majid and Hussain, Reference Majid and Hussain1996; Stathopoulou et al., Reference Stathopoulou, Psycharis, Chryssikos, Gionis and Theodorou2008) (Table I).
Table I. Refined unit-cell parameters, FWHM of 002 and 300 apatite reflections, and apparent crystallite dimensions of Pareiasaur bone as compared with literature data.

Crystallinity index (CI) was determined as the full width at half maximum (FWHM) of the 002 reflection in degrees 2θ (Elorza et al., Reference Elorza, Astibia, Murelaga and Pereda-Suberbiola1999) and comprised 0.266–0.250, which was typical for Mesozoic vertebrate bones (0.2–0.3, Elorza et al., Reference Elorza, Astibia, Murelaga and Pereda-Suberbiola1999).
Apatite has a hexagonal crystallographic structure with [001] crystallographic direction aligned along the length of the crystals. Based on this structure, the average size of coherently diffracting domains (otherwise referred to as crystallites) can be determined: the length from the 002 reflection, and the width from the size corresponding to the 300 reflection.
FWHM of 002 and 300 principal reflections corresponding to apatite crystallites were measured after background subtraction and correction for the instrument function. Considering that strain broadening is negligible (Dumont et al., Reference Dumont, Kostka, Sander, Borbely and Kaysser-Pyzalla2011), true crystallite sizes were determined from the Scherrer equation with the Scherrer's constant K = 2 for the (002) reflection and K = 1 for the (300) reflection (Dumont et al., Reference Dumont, Kostka, Sander, Borbely and Kaysser-Pyzalla2011):
 $$t{\rm} = {\rm} K\lambda /({\rm FWHM}\,\cdot {\rm cos}\theta ),$$
$$t{\rm} = {\rm} K\lambda /({\rm FWHM}\,\cdot {\rm cos}\theta ),$$where t is the true crystallite size, K is the Scherrer's constant, λ is the wavelength of the radiation used, Å (1.5406 Å for СuKα), FWHM is the full width at half maximum for the reflection measured, radians, and θ is half the diffraction angle for the reflection measured, degrees.
The error of crystallite sizes was obtained from the error of the FWHM provided by the Peakfit curve fitting program (Peakfit v. 4.11, Systat Software, Inc, San Jose, CA, USA).
Obtained crystallite dimensions (Table I) are in a good agreement with the values for Seismosaurus bone (Chipera and Bish, Reference Chipera and Bish1990), and are much greater than crystallite sizes for recent (10–15 nm) and for subfossil (20–25 nm) bone (Dumont et al., Reference Dumont, Kostka, Sander, Borbely and Kaysser-Pyzalla2011). Regarding the limited number of samples studied, this fact might indicate that crystallite size could have been increasing with geologic age, as well as with the degree of fossilization, i.e. the recrystallization of hydroxyapatite to F-apatite. Dumont et al. (Reference Dumont, Kostka, Sander, Borbely and Kaysser-Pyzalla2011) have shown that crystallite size in sauropod dinosaur bones does not increase with age or bone length. Similar values of bone crystallite sizes for Seismosaurus [which was the largest known dinosaur having the length of 40–50 m and weight of 140 tons (Gillette, Reference Gillette1991)] and for two rib fragments of D. vjatkensis pareiasaur (length up to 1.5–2.0 m, weight up to 150–300 kg, Ivakhnenko, Reference Ivakhnenko2001) might support this conclusion.
The geometrical parameter describing the shape of apatite crystallites is the aspect ratio defined as the ratio of crystallite length and width. Crystallites in fossil pareiasaur bones have the aspect ratio of 2.3–2.5, which are greater as compared with the aspect ratio of crystallites in recent mammals (1.7–2.0, Trueman et al., Reference Trueman, Behrensmeyer, Tuross and Weiner2004) and Seismosaurus bone (1.8, Chipera and Bish, Reference Chipera and Bish1990) and smaller than in fossil sauropod bones (3–5, Dumont et al., Reference Dumont, Kostka, Sander, Borbely and Kaysser-Pyzalla2011). Contrary to the results of Dumont et al. (Reference Dumont, Kostka, Sander, Borbely and Kaysser-Pyzalla2011), the aspect ratio appeared to be independent of bone length.
IV. CONCLUSION
The Kotelnich vertebrate fossil site, one of the richest of the Permian period, is characterized by the excellent preservation of fossil remains because of their burial in a silty anaerobic environment similar to modern bogs. Two bone fragments of Permian parareptile D. vjatkensis from this location are investigated using XRD. The bone is well-preserved and consists of apatite and secondary calcite, as well as quartz and dolomite derived from the embedding rock. From the refined crystal lattice parameters of the apatite phase, it can be assumed that these fossil bones underwent a post-mortem transformation from hydroxyapatite to authigenic fluorapatite structure.
Apatite crystallite size is larger in fossil pareiasaur bone than in subfossil and recent mammal bone and is in a good agreement with the values for Seismosaurus bone. Both crystallite size and aspect ratio obtained for the fossil pareiasaur bones support the conclusion about their independence of an individual's bone length. In other words, crystallite size and aspect ratio in fossil bioapatites are characteristic of authigenic fluorapatite that has been formed from pristine hydroxyapatite crystallites during fossilization processes. However, these conclusions are based on a small data set and require further confirmation.
ACKNOWLEDGEMENT
The reported study is carried out at the Geoanalyst Center for Collective Use and funded by RFBR according to the research project No. 18-35-00462_mol_a.