Introduction
Yam (Dioscorea spp.) is a staple food for more than 100 million people worldwide and an important crop in some countries located in the humid and sub-humid tropics in Africa, Asia and Latin America (Lebot, Reference Lebot2009). Around 5.0 million ha of yams were planted in 2012, producing 58.7 million tonnes of tubers (FAOSTAT, 2013). Yam is currently the third most important root and tuber crop in Africa, responsible for 96% of world production, exceeded only by cassava (Manihot esculenta Crantz) and sweet potato (Ipomoea batatas L. Lam.) (Dufie et al., Reference Dufie, Oduro, Ellis, Asiedu and Maziya-Dixon2013).
The representation of Brazil in the international yam market is still small (0.4% of world production; FAOSTAT, 2013). The Northeast region of Brazil accounts for about 90% of national production, especially the states of Pernambuco, Paraíba, Alagoas, Bahia and Maranhão. In these states, the predominant species are the Guinea yam species, also referred to as the African yam complex (Dioscorea cayenensis Lam./D. rotundata Poir.), followed by the species D. alata L., known as water yam (Veasey et al., Reference Veasey, Siqueira, Gomes, Nascimento, Ferreira, Silva, Silva, Ming, Peroni, Santos, Ming, Amorozo and Kffuri2012).
It is noteworthy that Brazil is predominantly found in the intertropical zone and displays various kinds of weather, ranging from hot and dry to cold and wet. In the Northeast and Southeast regions, tropical and subtropical climates predominate and the amount of rainfall ranges from 1000 to 1500 mm/year on average (Brasil, 2015). There are also a wide variety of vegetation types in Brazil (Schneeberger and Farago, Reference Schneeberger and Farago2003). This variety of climates and vegetation can be directly related to the genetic diversity distribution of the species complex D. cayenensis/D. rotundata.
There are controversies in the literature concerning the taxonomy of this species complex. Some authors consider them to be two different species (Tostain et al., Reference Tostain, Agbangla, Scarcelli, Mariac, Dainou, Berthaud and Pham2007; Bressan et al., Reference Bressan, Briner Neto, Zucchi, Rabello and Veasey2014), while others consider both as part of a species complex (Dansi et al., Reference Dansi, Mignouna, Pillay and Zok2001, Reference Dansi, Dantsey-Barry, Dossou-Aminon, N'Kpenu, Agré, Sunu, Kombaté, Loko, Dansi, Assogba and Vodouhè2013; Mignouna et al., Reference Mignouna, Dansi and Zok2002; Zannou et al., Reference Zannou, Agbicodo, Zoundjihékpon, Struik, Ahanchédé, Kossou and Sanni2009; Girma et al., Reference Girma, Hyma, Asiedu, Mitchell, Gedil and Spillane2014). Several studies have used morphological markers to measure the genetic diversity of these species (Onyilagha, Reference Onyilagha1986; Dansi et al., Reference Dansi, Mignouna, Zoundjihekpon, Sangare, Asiedu and Quin1999; Melo Filho et al., Reference Melo Filho, Santos, Mafra, Santos and Anunciação Filho2000; Bressan et al., Reference Bressan, Briner Neto, Zucchi, Rabello and Veasey2014; Girma et al., Reference Girma, Hyma, Asiedu, Mitchell, Gedil and Spillane2014; Loko et al., Reference Loko, Adjatin, Dansi, Vodouhe and Sanni2015). Among the molecular markers, microsatellites or SSR (simple sequence repeats) stand out for their efficiency, and for being multi-allelic, co-dominant, and showing a simple Mendelian segregation (Agarwal et al., Reference Agarwal, Shrivastava and Padh2008). These markers have been used in several studies to characterize the genetic diversity and structure of the species complex D. rotundata/D. cayenensis (Mignouna et al., Reference Mignouna, Abang and Fagbemi2003; Scarcelli et al., Reference Scarcelli, Daïnou, Agbangla, Tostain and Pham2005, Reference Scarcelli, Tostain, Vigouroux, Agbangla, Daïnou and Pham2006, Reference Scarcelli, Tostain, Vigouroux, Luong, Baco, Agbangla, Daïnou and Pham2011, Reference Scarcelli, Couderc, Baco, Egah and Vigouroux2013; Tostain et al., Reference Tostain, Agbangla, Scarcelli, Mariac, Dainou, Berthaud and Pham2007; Obidiegwu et al., Reference Obidiegwu, Kolesnikova-Allen, Ene-Obong, Muoneke and Asiedu2009; Sartie et al., Reference Sartie, Asiedu and Franco2012; Mengesha et al., Reference Mengesha, Demissew, Fay, Smith, Nordal and Wilkin2013; Silva et al., Reference Silva, Bajay, Monteiro, Mezette, Nascimento, Zucchi, Pinheiro and Veasey2014).
D. cayenensis and D. rotundata have been little explored in Brazil in basic studies of biology, ecology, evolution, genetics and plant breeding. Therefore, this study aimed to assess the genetic diversity and structure of local yam varieties of D. rotundata and D. cayenensis and their management and use by local farmers in Brazil using morphological and molecular markers.
Materials and methods
Plant material and crop management by farmers
In this study, we used 47 yam accessions of D. cayenensis and D. rotundata, collected from the Northeast, Southeast and South regions of Brazil. In the Northeast region, collections were made in the municipalities of Alhandra, Caaporã, Marcação, Rio Tinto, Santa Rita, Sapé and Sobrado, in the state of Paraíba (PB) and in the municipalities of Aliança, Bonito, Ferreiro, Goiana and São Joaquim do Montes, in the state of Pernambuco (PE). In the Southeast region, yams were collected in the municipalities of Ubatuba, Cananéia, Ilha Comprida, Iguape, Iporanga, Eldorado and Mogi das Cruzes, in the state of São Paulo (SP), and in the South region, collections were made in the municipalities of Joinville and Itajaí, in the state of Santa Catarina (SC) (Table 1; Fig. 1).

Fig. 1. Collection sites for D. cayenensis and D. rotundata accessions in Brazil: 1–8, Iguape/SP; 9–14, Cananéia/SP; 15–17, Iporanga/SP; 18, Eldorado/SP; 19, Mogi Guaçú/SP; 20 and 22, Ubatuba/SP; 21, Ilha Comprida/SP; 23 and 25, Joinville/SC; 24, Itajaí/SC; 26–28, Alhandra/PB; 29, Rio Tinto/PB; 30–32, Marcação/PB; 33–35, Sapé/PB; 36 and 37, Caaporã/PB; 38, Santa Rita/PB; 39, Sobrado/PB; 40, S.J. do Monte/PE; 41 and 42, Bonito/PE; 43 and 44, Goiana/PE; 45 and 46, Aliança/PE; 47, Ferreiro/PE.
Table 1. List of 47 Dioscorea accessions used in this study, including the identification of accessions in the germplasm bank (DGC), municipality and community of origin, folk names and geographic coordinates

D. c., Dioscorea cayenensis; D. r., D. rotundata.
In each municipality and from each farm, a sample of all yam varieties grown by the farmer was requested, in order to obtain as much information on the level of variability within fields. Species identification was carried out after field morphological evaluation, in accordance with the key to the main Dioscorea species provided by IPGRI/IITA (1997) and according to Martin (Reference Martin1978).
Information on the management and use of yam landraces was also recorded during yam collections. This information was obtained through interviews and using standardized semi-structured questionnaires, including questions about the size of the growing area, intercrop with other crops, local origin and names of the varieties, and whether farmers had observed the production of seeds, plus any other characteristics valued by the farmers. This research was approved by the Ethics Committee in Research with Human Beings of Luiz de Queiroz College of Agriculture, University of São Paulo, on 18 December 2007.
Morphological characterization
A whole tuber of each genotype was first planted in 3-litre plastic pots filled with regular soil with organic matter incorporated in a greenhouse, with automatic irrigation. After 3 months, the accessions were transferred to the experimental field, in October 2010, where they were planted in rows with two replications, using a wooden support to guide the growth of the plants. The spacing between rows and between plants was 2.5 and 1.5 m, respectively. A wide spacing was used to avoid the mixing of plants from different accessions.
Morphological characterization was performed on individual plants, using descriptors selected from a list developed by IPGRI and IITA for Dioscorea (IPGRI/IITA, 1997). A total of 18 traits were evaluated: six related to the stems (stem colour, presence or absence of wings and spines, twining direction, stem diameter and stem cross-section shape at the base); six associated with leaf characteristics (position of leaves, leaf shape and width, distance between the petiole and the upper extremity of the adult leaf, petiole colour and length); one related to flowering (presence or absence of flowering); five related to the tubers (presence or absence of underground tubers, number of tubers per plant, tuber shape, skin colour and flesh colour) (Table 2). Skin and flesh tuber colours were assessed visually, according to the colours described in Table 2. Most descriptors (14) were qualitative characters, while only four were quantitative. The quantitative traits were obtained from the average of ten measurements for each plant from each accession. The averages from each accession were placed within a size class (Table 2), to enable their analysis as binary data (presence or absence of a particular class), as performed for the qualitative characters.
Table 2. Traits used for morphological assessment of Dioscorea cayenensis/D. rotundata accessions and percentages found for each class

The evaluations were carried out in two stages: first at 6 months after transplanting in the field, in order to obtain data related to stem and leaf descriptors; and during harvest, in June 2011, when data related to the characterization of tubers were obtained. The character presence or absence of flowering was evaluated throughout the reproductive period.
Microsatellite procedures
DNA extraction followed Doyle and Doyle (Reference Doyle and Doyle1990) methodology with modifications, using lyophilized leaves. Among the modifications was the use of 3% CTAB (cetyl trimethyl ammonium bromide) extraction butter instead of 2%, and 1% polyvinylpyrrolidone (molecular weight 10,000), adding 4% β-mercaptoethanol just before use. After incubation at 65°C for 45 min, the samples were centrifuged for 10 min at 12,000 rpm. Then, the supernatant was transferred to a fresh tube following the addition of chloroform–isoamylalcohol (24:1). This procedure was repeated, adding 500 μl of chloroform–isoamylalcohol (24:1) plus 200 μl of 3% CTAB without β-mercaptoethanol. The next steps were performed according to the original method of Doyle and Doyle (Reference Doyle and Doyle1990). DNA quantification was performed on 0.8% agarose gels stained with blue green loading dye (LGC Biotechnology, São Paulo, Brazil) under UV light.
Ten pairs of primers were used: nine specific for the D. cayenensis/D. rotundata species complex from the enriched library developed by Silva et al. (Reference Silva, Bajay, Monteiro, Mezette, Nascimento, Zucchi, Pinheiro and Veasey2014), and a pair of heterologous primer (Dpr3F12) developed by Tostain et al. (Reference Tostain, Scarcelli, Brottier, Marchand, Pham and Noyer2006) (Supplementary Table S1, available online). PCR analyses were performed in a BioRad MyCycler thermal cycler using the touchdown programme. For DCA2, Dca10 and Dca11 primer pairs, the annealing temperature ranged from 60 to 50°C, while for Dca3, DCA4, Dca5, Dca6, Dca8, Dca9 and Dpr3F12 primer pairs, the annealing temperature ranged from 55 to 45°C. PCRs were carried out in the final volume of 16 μl per reaction, containing 20 ng of genomic DNA, Taq-Polymerase (5 U/μl), 5 × PCR buffer, MgCl2 at 2.5 mM, forward and reverse primers at 10 pmol/μl and each dNTP at 0.2 mM, using the following steps: an initial denaturation step of 5 min at 94°C, followed by ten pre-amplification cycles (30 s at 95°C, 30 s at the initial annealing temperature of 60 or 55°C, decreasing 1°C at each cycle, 50 s at 72°C), then 30 cycles of denaturation (30 s at 95°C, 30 s at 50°C, 50 s at 72°C) and a final extension step of 5 min at 72°C.
The amplified material was subjected to electrophoresis using denaturing 7% polyacrylamide gel, carried out at an initial power of 50 W for 10 min and, thereafter, at 60 W for 3 h. Molecular weight markers of 10 and 100 bp DNA ladders (Invitrogen™, São Paulo, Brazil) were used. Gels were stained with silver nitrate (Creste et al., Reference Creste, Tulmann Neto and Figueira2001), photographed with a digital camera and evaluated using a transilluminator.
Statistical analyses
Morphological data were evaluated as binary data (presence or absence of a phenotypic class) for both qualitative and quantitative characters. Due to the polyploid nature of D. cayenensis and D. rotundata (Dansi et al., Reference Dansi, Mignouna, Pillay and Zok2001; Girma et al., Reference Girma, Hyma, Asiedu, Mitchell, Gedil and Spillane2014), microsatellite bands were evaluated as presence or absence of bands, according to the procedures adopted for dominant markers. Thus, the number of bands for each pair of primers and also the polymorphism percentage (number of polymorphic bands/total number of bands) were obtained. The polymorphism information content (PIC), proposed by Botstein et al. (Reference Botstein, White, Skolnick and Davis1980), was calculated by the following formula:
 $$\hbox{PIC} = 1 - { \sum _{ i - 1}^{ n } }\, p _{ i }^{2},$$
$$\hbox{PIC} = 1 - { \sum _{ i - 1}^{ n } }\, p _{ i }^{2},$$
where p i is the allele (band) frequency at each locus and n is the number of observed alleles.
The Shannon index (H′) (also called the Shannon–Weaver index or Shannon–Wiener index), one of the diversity indices used to quantify the diversity in categorical data (Yeh et al., Reference Yeh, Yang, Boyle, Ye and Mao1997), was obtained in two separate analyses. The first was made considering the two species (23 D. cayenensis accessions and 24 D. rotundata accessions) and the second separating the accessions by their region of origin (Southeast, Northeast and South). To calculate this index, the following formula was used:
 $$H^{\prime} = { \sum _{ i = 1}^{ s } }\,( p _{ i }\,\hbox{log}\, p _{ i }),$$
$$H^{\prime} = { \sum _{ i = 1}^{ s } }\,( p _{ i }\,\hbox{log}\, p _{ i }),$$
where p i is the frequency of each variety, considering i ranging from 1 to S (richness).
Both molecular and morphological data were used to obtain Jaccard's similarity coefficients. The similarity matrices were subjected to Darwin software v.5.0 (Perrier and Jacquemoud-Collet, Reference Perrier and Jacquemoud-Collet2006) for the cluster analyses using the unweighted neighbour-joining algorithm. The correlation between the genetic and geographic distance matrices as well as between morphological and molecular genetic distances were obtained by Pearson's correlation (r), whose significance was evaluated by the Mantel test (Mantel, Reference Mantel1967), using the NTSYS-pc software (Rohlf, Reference Rohlf1992).
To verify the genetic structure of the sampled accessions, we conducted Bayesian analysis using the software Structure v.2.3.1 (Pritchard et al., Reference Pritchard, Stephens and Donnelly2000; Falush et al., Reference Falush, Stephens and Pritchard2007). The Structure software was run using the admixture model, correlated allele frequencies and repeated ten times for each K (number of clusters assumed) with a burn-in of 500,000 iteraction followed by 500,000 Markov chain Monte Carlo iteraction. The most likely number of clusters was chosen using the ΔK method (Evanno et al., Reference Evanno, Regnaut and Goudet2005).
The analysis of molecular variance (AMOVA), using Arlequin software (Schneider et al., Reference Schneider, Roessli and Excoffier2000), was obtained for the discrimination of the partition of genetic variability observed in the following sampled levels: within and among geographic regions, and between and within species, considering D. cayenensis and D. rotundata as two different species.
Results
Crop management
Yam cultivation (D. cayenensis/D. rotundata) in the studied areas was mostly performed in less than 2 ha fields. Only 11% of the respondents used a cultivation area of over 2 ha, with large-scale marketing purposes, as in the case of some farmers in the Northeast region of the following municipalities: Camucim de São Félix/PE (5 ha); Bonito/PE (10 ha); Alhanbra/PB (20 ha); São Joaquim do Montes/PE (30 ha). Also, 52% of the interviewed farmers grew yam intercropped with other crops such as cassava, sweet potato, beans (Phaseolus vulgaris L.), corn (Zea mays L.), coconut (Cocos nucifera L.) and sugarcane (Saccharum spp.). The fields were of fundamental importance for the conservation of biodiversity, especially for the local communities in this study, where 61% of the farmers cultivated yams exclusively in the fields, 21% only in home gardens and 18% both in the swidden fields and home gardens. This information was provided by farmers, mostly men (89%), with a mean age of 53.3 years (23 ≤ n≤ 83). When asked for how long have they lived in the studied area, most farmers (78%) replied that they lived in the same area for more than 20 years, while 45% stated that they lived in the same place where they were born.
Of the interviewed farmers, 48% grew more than one yam variety, but not necessarily belonging to the species complex D. rotundata/D. cayenensis, as farmers also grew other yam species, such as D. alata, D. trifida and D. bulbifera. Regarding the species D. rotundata and D. cayenensis, the most cited variety in the Northeast region of Brazil, in the states of Pernambuco and Paraíba, was the variety inhame da Costa (coast yam, 45.5%), followed by inhame de Pernambuco (Pernambuco yam, 22.7%), inhame original (original yam), inhame (yam) (9.1% each), and the yam varieties japecanga, inhame branco (white yam) and inhame da folha larga (large leaf yam) (4.5% each). In the states of São Paulo and Santa Catarina, in the Southeast and South regions of Brazil, the most cited folk name was cará de espinho (spine yams, 70.8%), followed by cará d'Angola (yam from Angola, 12.5%), cará guaçu (guaçu yam), cará São João roxo (Saint John's purple yam) and cará espinhento (spiny yam) (4.2% each), plus inhame da Costa for an accession obtained in Mogi Guaçú/SP (4.2%), but this accession was originated from the Northeast region, which explains the name, typical from this region (Table 1).
Most farmers (58%) acquired the tubers for free among friends, neighbours, relatives or bought them in fairs of the same municipality, while 20% acquired them in another municipality in the same state, and only 12% cultivated yams originated from other Brazilian states. The reasons mentioned were to increase the number of tubers of an existing variety and also to try new varieties. Moreover, only 10% never bought tubers, always planting the tubers derived from materials obtained for a long time, passed from generation to generation. According to the survey, 42% of the farmers reduced their yam cultivation area due to increased production costs, lack of information regarding the proper crop management, and especially due to a decrease in labour work of younger people, and 42% of the farmers planted yam varieties in the past, which they do not plant any more.
Regarding the sexual reproductive process, 65% of the respondents observed the occurrence of flowering, where after a certain time the inflorescences dried up. This moment indicates to the farmers the ideal time to harvest the tubers. The fruiting process was not observed by any of the respondents, and when asked about the way of propagating the crop, all farmers said it was through vegetative propagation, using the tuber.
Morphological characterization
The morphological characterization showed that 100% of the local varieties had green and round stem with cross-section shape at the base, no wings on the stem, presence of spines on the stem, growth direction in the anticlockwise direction and green petiole. Furthermore, 98% had stem diameter at 15 cm from the base of the plant >0.6 cm, while 2% had diameters ranging from 0.4 to 0.6 cm. One of the most important characters in the visual separation of the two species was leaf shape; we found 52% of sagittate leaves, typical of D. rotundata (Supplementary Fig. S1, available online), and 48% of cordate leaves, typical of D. cayenensis (Supplementary Fig. S2, available online), according to Martin (Reference Martin1978). As for the position of the leaves, 67% had opposite leaves while 33% showed alternate leaves. All of the accessions classified as D. rotundata showed opposite leaves, except for one accession, while D. cayenensis accessions showed both opposite and alternate leaves. The leaf width varied from < 0–6 cm (40%), 6–10 cm (42%) to >10 cm (18%). Eighty one percent showed the distance between the petiole and the upper extremity of the adult leaf to be < 10 cm, 8% showed it to be from 10 to 15 cm, while 11% showed a distance of >15 cm. As for the petiole length, 56% showed it to be < 5 cm while 44% showed it to be from 5 to 10 cm.
All of the accessions (100%) had underground tubers, varying from one tuber (58%) to a few (two to five) tubers (42%). Tuber shape varied from elongated (54%), irregular (38%) to round (8%) shape, while tuber skin colour (beneath the bark) was shown to be brown (77%) or yellow (23%). Most of the D. cayenensis accessions showed an irregular tuber shape, while most of D. rotundata accessions displayed an elongated tuber shape. The two species were not discriminated with respect to tuber skin colour. However, flesh colour was an important trait for the separation of the accessions, considering that the yellow colour is typical of D. cayenensis and the white colour typical of D. rotundata, according to Martin (Reference Martin1978) and according to the observations made in this study. Flesh colour varied in this study from white (52%), yellow (46%) and yellow and purple (2%), with only one individual in this last category.
In regards to flowering, only 17% of the varieties flowered, but no seed was observed for any of them. Among the 17% (eight accessions), all but one were classified as D. rotundata according to the flesh tuber colour and leaf shape.
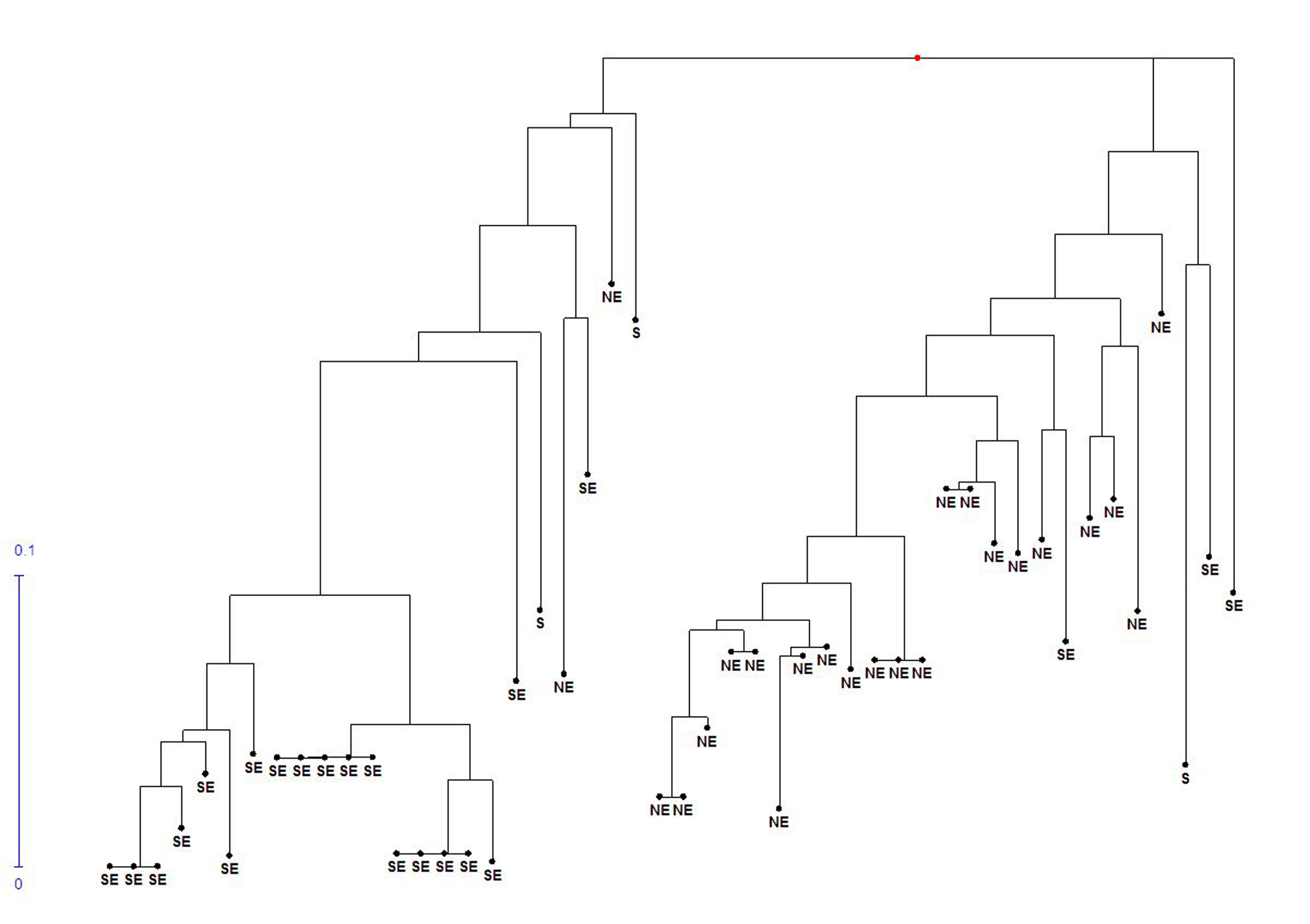
In the cluster analysis for morphological characters (Supplementary Fig. S3, available online), the accessions were divided into two groups, with a clear separation of the accessions collected in the Southeast and Northeast regions of Brazil in two isolated groups. Among the accessions from the South region, one of them (accession no. 23) was situated in an isolated group, while the other two (accession no. 22 and 25) were grouped together in the Northeast accessions. We observed that the Southeast accessions were composed of the morphological types equivalent to D. cayenensis (Supplementary Fig. S2, available online), while the Northeast accessions were constituted of the morphological types equivalent to D. rotundata (Supplementary Fig. S1, available online).
The Bayesian analysis, conducted using the Structure software, supported the cluster analysis, as it also grouped the accessions into two groups (Fig. 2), as suggested by the method of Evanno et al. (Reference Evanno, Regnaut and Goudet2005). Also, in this case, one of the groups classified all the Southeast accessions (dark grey colour), while accession no. 19, originated from Mogi Guaçú/SP, showed about 40% characters of the other group. The second group (light grey colour) classified all the Northeast accessions and two accessions from the South, while accession no. 23 also from the South (from Joinville, SC) showed about 40–45% characters of the first group. Therefore, these two analyses show clear morphological differences between the accessions from the Southeast and Northeast regions of Brazil, which seems to be reflecting the difference between the species D. cayenensis (collected in the Southeast region) and D. rotundata (collected mostly in the Northeast region).

Fig. 2. Bayesian approach for the analysis of 47 D. cayenensis/D. rotundata accessions (a) and 24 D. rotundata accessions (b) based on 18 morphological traits. Each accession is represented by a vertical bar. Accessions 1 to 22 originated from the Southeast region, 23 to 25 from the South region, and 26 to 47 from the Northeast region in Brazil. Details of each accession are given in Table 1.
The AMOVA performed by region showed that most of the variability (50.9%; P< 0.01) was found between regions, although a significant percentage of the total variation was also attributed to the variation within regions (49.1%) (Table 3). To confirm this hypothesis, a second AMOVA was performed considering the two species separately (Table 3). This new analysis showed that most of the variation was attributed to the variation between species (60.2%; P< 0.01). However, a still significant proportion (39.8%) was attributed to the variation within species, showing that despite the differences between the two species, there is also within-species diversity.
Table 3. AMOVA for 47 yam accessions, considering the three regions in Brazil [South (Santa Catarina state), Southeast (São Paulo state) and Northeast (Pernambuco and Paraíba states)] and the two species [D. cayenensis (23 accessions) and D. rotundata (24 accessions)], evaluated with 18 morphological traits and ten microsatellite primers

DF, degree of freedom; SS, sum of squares.
a Number of permutations = 1000.
In order to analyse more deeply into the intra-specific diversity, new analyses were performed using the Structure software, but this time for each species separately. For these analyses, D. cayenensis accessions were not separated into groups, showing, therefore, a morphological uniformity among them. On the other hand, D. rotundata accessions were divided into two groups, with accessions no. 22 (Ubatuba/SP), no. 31 (Marcação/PB), no. 33 (Sapé/PB), no. 40 (S.J. do Monte/PE) and no. 41 (Bonito/PE) classified into one group and all the others into another group (Fig. 2), showing a morphological distinction between these groups. The five accessions cited above differed from the others because all of them showed flowering, along with two other accessions (no. 32 and 34), in addition to having a greater distance between the petiole and the upper extremity of the adult leaf (>15 cm), and higher leaf width (>10 cm).
Microsatellite characterization
A total of 32 bands (alleles), ranging from two to four bands per primer, with an average of 3.2 bands per primer pair, were obtained for 47 yam accessions of D. rotundata and D. cayenensis (Supplementary Table S1, available online). The percentage of polymorphic bands ranged from 33 to 100%, with an average of 56.8%. Based on the PIC value, varying from 0.34 to 0.70, all primer pairs were classified as very informative (PIC>0.5), according to the classification of Botstein et al. (Reference Botstein, White, Skolnick and Davis1980), except for the primer pair Dca8, with a PIC value of 0.34.
Genetic diversity, assessed by the Shannon index, showed a very similar index for the accessions of D. cayenensis (H′ = 0.27) and D. rotundata (H′ = 0.23). By keeping the accessions of the two species together, but separated by region, the value of the Shannon index was higher for the three accessions of the South region (H′ = 0.31), intermediate for the Southeast accessions (H′ = 0.27) and lower for the Northeast accessions (H′ = 0.21). Although the accessions of the South region had the highest diversity, this result should be viewed with reservations due to the low number of accessions, certainly not representative of the region.
In the cluster analysis, the accessions were classified into two groups, and again with a clear separation of the Southeast and Northeast accessions (Supplementary Fig. S4, available online). The three accessions of the South region were classified within the Southeast region group for the SSR markers. This result corroborates the data found for the morphological characterization, which also showed a clear separation of the accessions of the Northeast and Southeast regions.
The result of the Bayesian analysis using the Structure software for microsatellite data is in accordance with the morphological data, classifying the accessions into two groups (Fig. 3). Also in this case, one of the groups included most accessions (no. 1 to 18) from the Southeast region (light grey colour), plus an accession from the South region (no. 25). The Northeast accessions were classified into a second group (dark grey colour), along with accessions no. 19, 20, 21 and 22 from the Southeast region and accessions no. 23 and 24 from the South region. Therefore, there were also clear genetic differences, evaluated by molecular markers, between local varieties from the Southeast and Northeast regions of Brazil, which again seems to be reflecting the differences between the species D. cayenensis (collected in the Southeast region) and D. rotundata (collected in the Northeast region).

Fig. 3. Bayesian approach for the analysis of 47 D. cayenensis/D. rotundata accessions based on 10 SSR markers (a) and 23 D. cayenensis accessions based on SSR markers, for K= 2 groups (b) and K= 3 groups (c). Each accession is represented by a vertical bar. Accessions 1 to 22 originated from the Southeast region, 23 to 25 from the South region, and 26 to 47 from the Northeast region of Brazil. Details of each accession are given in Table 1.
The AMOVA performed by region indicated that most of the variability (51.9%; P< 0.01) was concentrated between regions, although a significant percentage of total variation was also attributable to the variation within regions (48.1%) (Table 3). In the variance analysis considering the two species separately, genetic variation was in the same proportion between species (49.4%; P< 0.01) and within species (50.6%; P< 0.01). These data suggest that the species complex can indeed be two different species, but to reach this conclusion, further studies should be conducted, including ancestral species.
To test the hypothesis of within-species variability, new Bayesian analyses were conducted for each species separately, similarly to the morphological data. However, the results were the opposite from the morphological data, since this analysis showed no difference among the D. rotundata accessions for the SSR markers, with all of the accessions being classified into one group. However, D. cayenensis accessions were grouped into two (K= 2) or three (K= 3) groups, according to the method of Evanno et al. (Reference Evanno, Regnaut and Goudet2005). The analysis of D. cayenensis accessions divided into two groups (Fig. 3), separated accessions no. 8 (Iguape/SP), no. 17 (Iporanga/SP), no. 19 (Mogi Guaçu/SP), no. 20 (Ubatuba/SP), no. 21 (Ilha Comprida/SP), no. 22 (Ubatuba/SP) and no. 23 (Joinville/SC) from the remaining accessions of Iguape/SP, Cananéia/SP, Iporanga/SP and Eldorado/SP. Examining the map (Fig. 1), we observed that these accessions were geographically more distant from the others, except for accessions no. 8 and 17, as this group includes two accessions from the South region, in the state of Santa Catarina, and accessions from more distant municipalities, such as Mogi Guaçú, Ubatuba and Ilha Comprida. The analysis of the accessions that divided into three groups (Fig. 3) proved to be interesting, because besides the separation of the accessions cited above, no. 8, 19, 20, 21, 22 and 23, it separated in a third group all accessions from Iporanga/SP (no. 15–17) and Eldorado/SP (no. 18), which were located more into the interior of the state of São Paulo, being farther away from most of the coastal accessions (Fig. 1), such as Iguape and Cananéia accessions. This third group also included accessions no. 11 and 13 from Cananéia.
All the genetic structure and population dissimilarity analyses suggest that there is a geographic isolation between accessions. This finding is supported by the high and significant correlation between geographic and genetic distances for morphological (r= 0.68; P< 0.01) and molecular markers (r= 0.64; P< 0.01), and also by the high and significant correlation between morphological and molecular genetic distances (r= 0.56; P< 0.01), according to the Mantel test.
Discussion
This study showed that, in Brazil, yam (D. cayenensis/D. rotundata) cultivation occurs mainly by farmers practising family farming, usually using the swidden fields as a planting unit, similar to what happens in some countries in Africa, where the centre of origin and diversity of these species is located (Mahalakshmi et al., Reference Mahalakshmi, Ng, Atalobhor, Ogunsola, Lawson and Ortiz2007). The swidden fields are of fundamental importance for the conservation of biodiversity, since several multi-purpose species are usually cultivated and a high diversity is observed (Scarcelli et al., Reference Scarcelli, Tostain, Vigouroux, Agbangla, Daïnou and Pham2006; Peroni et al., Reference Peroni, Kageyama and Begossi2007; Amorozo, Reference Amorozo2008). Traditional small-scale farming plays a key role in maintaining the diversity of cultivated plants, representing an important form of in situ conservation of genetic resources for future generations (Bressan et al., Reference Bressan, Veasey, Peroni, Felipim and Santos2005; Zizumbo-Villarreal et al., Reference Zizumbo-Villarreal, Colunga-Garciamarín, Cruz, Delgado-Valerio and Gepts2005; Salako et al., Reference Salako, Fandohan, Kassa, Assogbadjo, Idohou, Gbedomon, Chakeredza, Dulloo and Kakaı2014).
Considering that the vast majority of farmers lived on the site for more than 20 years, with 45% living in the same place where they were born, we found low geographic mobility in this study, which may have influenced the origin of the accessions, the trade relationship of the varieties among farmers and the modes of knowledge transmission. We also found that most of the varieties were originated in the study area. These results reinforce the occurrence of low mobility among local people and show that when trade occurs, it is on a regional basis, with limited range. When asked about where the yam varieties originated, 68% of the farmers stated they did not know, with answers such as this one: ’when I was born it already existed in the family…; it was here already when I arrived…; it has been here for a long time; it is local, it always existed here’. Similar data were obtained by Tamiru et al. (Reference Tamiru, Becker and Maass2008), who found that there was no formal system for supplying yam seeds in Ethiopia, as well as specialized farmers in the production of planting materials.
Another issue that showed a low geographic mobility is the regionalization of vernacular names assigned to this crop (Veasey et al., Reference Veasey, Siqueira, Gomes, Nascimento, Ferreira, Silva, Silva, Ming, Peroni, Santos, Ming, Amorozo and Kffuri2012). In the Northeast states, it is known as inhame (yam), with variations within this general denomination, such as inhame da Costa (coast yam), inhame de Pernambuco (yam from Pernambuco), inhame original (original yam), among others. However, in other regions of the country, as in the South and Southeast, it is known by farmers by the general denomination of cará (yam), such as cará de espinho (spine yams), which is the most common name in these regions. According to Loko et al. (Reference Loko, Adjatin, Dansi, Vodouhe and Sanni2015) and other studies in Africa (Dansi et al., Reference Dansi, Mignouna, Zoundjihekpon, Sangare, Asiedu and Quin1999; Tamiru et al., Reference Tamiru, Becker and Maass2008), vernacular names given to yam cultivars often vary across ethnic zones and even between villages within ethnic areas.
The loss of genetic resources was noticed when farmers mentioned that they had planted yam varieties in the past that currently are no longer planted, and the main reasons for this, according to the respondents, are related to changes in physical and chemical soil conditions, reduced labour and unequal distribution of land. In Togo, Africa, yam production has been found to be hampered by many constraints, such as pests and diseases, poor soils, drought, flooding or high soil moisture, causing severe yield losses and genetic erosion (Dansi et al., Reference Dansi, Dantsey-Barry, Dossou-Aminon, N'Kpenu, Agré, Sunu, Kombaté, Loko, Dansi, Assogba and Vodouhè2013). Several factors have challenged the continuity of agriculture practised by indigenous peoples and small-family producers. Among them, the expansion of modern agriculture in large regions, increased land concentration, and the expansion of non-agricultural job opportunities (industry and services) are important factors to be considered. In view of this, a significant portion of new rural generations have emigrated or engaged in activities of secondary and tertiary sectors in the rural area (Amorozo, Reference Amorozo2008). In a 20-year comparison on the maintenance of cassava diversity in the state of Mato Grosso, Brazil, Marchetti et al. (Reference Marchetti, Massaro-Junior, Amorozo and Butturi-Gomes2013) found similar diversity indices in 1992 and 2011, but the mean number of varieties per farmer decreased from 9.4 to 5.4. The main reasons for this have been reported as the reduction by two-thirds in the size of cultivated areas, and the fact that the majority of young people are not engaged in agriculture activities any more or are engaged only part-time.
Regarding the morphological characterization, various characters are used for the classification of yam landraces. In the present study, the most important traits that allowed the discrimination of the two species under study were the following: cordate leaf shape and yellow tuber flesh colour for D. cayenensis and sagittate leaf shape and white tuber flesh colour for D. rotundata. Also, tuber shape was mostly elongated or round for D. rotundata and irregular for D. cayenensis, and these differences may be related to the fact that D. rotundata was mostly found in the Northeast region, where this species is widely used as their main commercial yam crop, while in the Southeast region, the main commercial yam crop is D. alata (Veasey et al., Reference Veasey, Siqueira, Gomes, Nascimento, Ferreira, Silva, Silva, Ming, Peroni, Santos, Ming, Amorozo and Kffuri2012). Considering that the Northeast region is where these species are mostly used in a commercial scale, the white colour and the elongated or round tuber shape may be ideal traits for the commercialization of these species.
Dansi et al. (Reference Dansi, Mignouna, Zoundjihekpon, Sangare, Asiedu and Quin1999) established an identification key for the differentiation of cultivars of the complex D. cayenensis/D. rotundata, and also observed traits related to leaf shape and colour of the tubers as important descriptors to differentiate between these species. A recent study (Girma et al., Reference Girma, Hyma, Asiedu, Mitchell, Gedil and Spillane2014) has concluded that only tuber flesh colour could distinguish the two species, with yellow flesh colour being attributed to D. cayenensis, although it showed some ambiguity in some accessions.
Onyilagha (Reference Onyilagha1986) evaluated 14 D. rotundata accessions using morphological characters, and concluded that the characters with higher discriminatory power were leaf size, tuber size, size of flower spikelets per node and spikelet length. Melo Filho et al. (Reference Melo Filho, Santos, Mafra, Santos and Anunciação Filho2000) characterized morphologically 11 accessions of Dioscorea spp. from the yam collection of the Federal Rural University of Pernambuco, and observed that among the analysed traits, the length and width of the leaf had the highest contribution in discriminating the accessions. In our study, leaf size was important in differentiating D. rotundata accessions, with five accessions showing a greater distance (>15 cm) between the petiole and the upper extremity of the adult leaf and higher leaf width. Bressan et al. (Reference Bressan, Briner Neto, Zucchi, Rabello and Veasey2014) used 24 morphological traits to assess 21 local varieties of D. cayenensis originating from the Vale do Ribeira, in the state of São Paulo, and two D. rotundata accessions. The accessions were classified into two groups, according to their municipality of origin (Iguape and Cananéia) in the cluster analysis, which may indicate the influence of cultural units in the morphological variation.
In the analysis performed with ten SSR loci, we found moderate diversity among the accessions, with 3.2 alleles per locus and a PIC average of 0.57. Obidiegwu et al. (Reference Obidiegwu, Kolesnikova-Allen, Ene-Obong, Muoneke and Asiedu2009) observed a higher number of alleles per locus, with an average of 8.06, in the analysis of 219 D. cayenensis/D. rotundata accessions from various countries in Africa, using 15 SSR loci, and found a slightly higher PIC average value of 0.65. Sartie et al. (Reference Sartie, Asiedu and Franco2012) observed two to seven alleles per locus in a total of 32 SSR loci, in the assessment of four species of Dioscorea, including the two species in this study, and D. alata and D. dumetorum. Mengesha et al. (Reference Mengesha, Demissew, Fay, Smith, Nordal and Wilkin2013) estimated an average of 8.6 alleles per locus in the assessment of 58 accessions (21 wild and 37 cultivated) of three species (D. abyssinica Hochst. ex Knuth, D. praehensilis Benth. and D. cayenensis/D. rotundata).
In the cluster and Bayesian analyses performed in this study, for both morphological and molecular markers, consistent groups were obtained according to the region of origin of the accessions. Besides, high and significant genetic correlations were found between geographic and genetic distances and also between morphological and molecular markers. Sartie et al. (Reference Sartie, Asiedu and Franco2012) also observed a high and significant correlation coefficient (r= 0.67) between genetic (SSR) and morphological markers. It was interesting to note, in our study, that the identified accessions, according to the morphological analysis, as belonging to D. cayenensis, i.e. those with yellow flesh tubers and cordate leaves, originated from the Southeast region. On the other hand, the accessions identified as D. rotundata, with white flesh tubers and sagittate leaves, originated from the Northeast region. The climates in these two regions are different, with the Northeast showing a tropical climate, as well as dryer areas as in the Sertão, where rains are irregular with long periods of drought and high temperatures, while the Southeast is characterized by a subtropical climate (Brasil, 2015). Vegetation can also be quite different in the two regions, and this variety of climates and vegetation could be related to the distribution of the species D. cayenensis and D. rotundata in Brazil.
Other studies, such as Ramser et al. (Reference Ramser, Weising, Lopez-Peralta, Terhalle, Terauchi and Kahl1997), Dansi et al. (Reference Dansi, Mignouna, Zoundjihékpon, Sangare, Asiedu and Ahoussou2000), Mignouna et al. (Reference Mignouna, Dansi and Zok2002) and Bressan et al. (Reference Bressan, Briner Neto, Zucchi, Rabello and Veasey2014), also found a clear separation between these two species. Ramser et al. (Reference Ramser, Weising, Lopez-Peralta, Terhalle, Terauchi and Kahl1997) studied the taxonomy and phylogeny of the species D. cayenensis/D. rotundata through molecular markers, such as RAPD, microsatellite and chloroplast DNA, and observed a clear separation of these two species and the formation of two large groups. They also observed, using AMOVA, that most of the variability was found among the groups of accessions (89.2%), i.e. between species, a result similar to our study, in which most of the morphological variability (60%) was observed due to the variation between species. Dansi et al. (Reference Dansi, Mignouna, Zoundjihékpon, Sangare, Asiedu and Ahoussou2000), who analysed 465 accessions from the D. cayenensis/D. rotundata complex collected in different localities of the Republic of Benin in traditional farming systems, using isozymes, observed a clear separation of the two species in the cluster analysis. Mignouna et al. (Reference Mignouna, Dansi and Zok2002) and Bressan et al. (Reference Bressan, Briner Neto, Zucchi, Rabello and Veasey2014), analysing accessions from this species complex using morphological and isozyme markers, also observed the formation of two significant groups, observing a clear separation between D. rotundata and D. cayenensis accessions. Sartie et al. (Reference Sartie, Asiedu and Franco2012) considered that D. rotundata and D. cayenensis may have co-evolved from the same ancestor, and may have become separated as a result of the domestication process, selection or due to the genotype × environment interaction. The results obtained by Girma et al. (Reference Girma, Hyma, Asiedu, Mitchell, Gedil and Spillane2014) with next-generation sequencing-based genotyping, cytometry and morphological evaluations, showing increased ploidy levels and heterozygosity in D. cayenensis, and allele sharing between the two species, indicate that D. cayenensis arose from D. rotundata but not vice versa, and also that D. cayenensis could be considered as a subspecies of D. rotundata.
Considering the genetic structure among the accessions evaluated for both types of markers (morphological and microsatellites), we found that most of the variability occurred between regions. However, high variability was also found within regions, probably due to the fact that these are vegetatively propagating species with frequent exchange of materials among farmers, increasing variability within regions. Also, the fact that the farmers did not notice any fruiting, the same happening in our field experiment, makes the occurrence of sexual reproduction, and thus genetic variability arising through recombination, unlikely in our conditions. However, the increase in variability through the occurrence of mutation is possible (Scarcelli et al., Reference Scarcelli, Tostain, Vigouroux, Luong, Baco, Agbangla, Daïnou and Pham2011, Reference Scarcelli, Couderc, Baco, Egah and Vigouroux2013). According to Scarcelli et al. (Reference Scarcelli, Tostain, Vigouroux, Luong, Baco, Agbangla, Daïnou and Pham2011), each variety could be interpreted as being the product of sexual reproduction having evolved by mutation. Our study did not show any evidence of sexual reproduction, although further studies more directed to this objective might provide a different view.
The high and significant correlation between genetic and geographic distances obtained in this study for both markers shows the spatial structure of genetic variability. Tostain et al. (Reference Tostain, Agbangla, Scarcelli, Mariac, Dainou, Berthaud and Pham2007) conducted a study with 146 accessions of D. rotundata using ten pairs of SSR heterologous primers, reporting a high genetic diversity and also a low but significant correlation between genetic diversity and geographic distances, showing a slight spatial structure. Other studies have found a low correlation between genetic and geographic distances, but found high diversity among yam cultivars of the D. cayenensis/D. rotundata complex (Tostain et al., Reference Tostain, Agbangla, Scarcelli, Mariac, Dainou, Berthaud and Pham2007; Zannou et al., Reference Zannou, Agbicodo, Zoundjihékpon, Struik, Ahanchédé, Kossou and Sanni2009). Significant but low correlations [r= 0.08; P= 0.0007 for SSR and r= 0.16; P= 0.0002 for ISSR (inter-simple sequence repeats)] were found between genetic and geographic distances among 53 accessions of D. trifida, originated from several regions in Brazil (Nascimento et al., Reference Nascimento, Rodrigues, Koehler, Gepts and Veasey2013), showing a slight spatially structured genetic diversity for this species in Brazil. On the contrary, Siqueira et al. (Reference Siqueira, Bonatelli, Günther, Gawenda, Schmid, Pavinato and Veasey2014) found a low correlation (r= 0.05; P= 0.016) between genetic and geographic distances among 72 local varieties of water yam (D. alata) originated from several regions in Brazil. They attributed this result to a continuous exchange of accessions of this species by farmers throughout the country.
The results in our study indicate a separation between D. cayenensis and D. rotundata accessions in Brazil, with D. cayenensis predominantly occurring in the Southeast and D. rotundata in the Northeast of the country. Therefore, it is concluded that, in Brazil, most of the genetic variability of these species is structured between regions and between species. It also suggests that these species were probably introduced in Brazil in different areas and currently there is a certain geographic isolation keeping them separated from each other. However, we recommend that further studies be carried out using larger sample sizes in other states and municipalities of the country to confirm this finding. A considerable intra-specific variability was also found in this study. Finally, it can be concluded that the genetic data obtained in this study will be useful for yam breeding programmes in Brazil and can also be used aiming at in situ/on-farm conservation purposes.
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S1479262115000647
Acknowledgements
The authors would like to thank the researchers Antônio Henrique dos Santos, Lin Chau Ming, Nivaldo Peroni and Marcos V.B.M. Siqueira for their assistance in this research and the agriculturists for their contributions in the field collecting and interviews. The authors also wish to thank the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP) (Process no. 2007/04805-2) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the financial support and scholarships provided for this study.