


Introduction
In their comprehensive meta-analysis of the twin and adoption literature on antisocial behavior, Rhee & Waldman (Reference Rhee and Waldman2002) noted that the two most common methods for assessment were by self-report using one of a wide variety of questionnaires and through objective measures of criminal behavior (CB) from official records of arrests or convictions. They concluded that individual differences in antisocial behavior are substantially influenced by genetic, shared environmental and individual-specific environmental factors, which account for 41, 19 and 43% respectively of the variance in liability. They also reported higher estimates of heritability and lower estimates of shared environment in studies where antisocial behavior was assessed by official records as compared to self-report. However, Rhee & Waldman also noted a lack of studies using multiple assessment methods and called for more studies of this kind.
The current analysis of data from twin participants extends prior work on the etiology of antisocial behavior in two crucial ways. First, addressing the need identified by Rhee & Waldman (Reference Rhee and Waldman2002), our analysis incorporated both self-report and criminal record data. Second, linking to recent work on antisocial behavior as part of an extended disinhibitory spectrum, our biometric–etiologic analysis focused on a broader construct of externalizing proneness encompassing antisocial/delinquent behavior along with affiliated personality traits (Sher & Trull, Reference Sher and Trull1994; Krueger et al. Reference Krueger, Hicks, Patrick, Carlson, Iacono and McGue2002, Reference Krueger, Markon, Patrick, Benning and Kramer2007). Specifically, using males from the Swedish Twin study of CHild and Adolescent Development (TCHAD; Lichtenstein et al. Reference Lichtenstein, Tuvblad, Larsson and Carlstrom2007), we examined, using multivariate structural equation modeling, the association between five self-report measures of antisocial behavior assessed at age 16–17 years [general externalizing traits (EXT), self-reported delinquency (SRD), impulsivity (IMP), grandiosity (GRD) and callousness (CLS)], and CB as defined by entry in the official Swedish Register of Persons Suspected of Offenses from the age of 13 to 25 years. The availability of both self-report and arrest registration data provided a basis for clarifying relationships between the underlying risk factors for antisocial traits and behaviors as assessed in these two alternative ways. More specifically, we sought to determine if the sources of the association vary between self-report measures of antisocial traits and behaviors and CB. In particular, would results from multivariate twin modeling demonstrate that some externalizing trait dimensions better reflect genetic risk for CB whereas others are superior at indexing the shared environmental or non-shared environmental risk factors?
Method
Subjects for these analyses came from the TCHAD, which began with all twin pairs born in Sweden between May 1985 and December 1986 where both twins were alive and residing in Sweden in 1994 (Lichtenstein et al. Reference Lichtenstein, Tuvblad, Larsson and Carlstrom2007). Twins returned questionnaires at ages 13–14, 16–17 and 19–20 years. In this report, we focused on response data from the age 16–17 assessment, which were available for 83% of those eligible. We chose this assessment because it was in the middle of the age period over which we had information about their registration in the suspect registry.
Each questionnaire was approved by the Ethics Committee of the Karolinska Institute, Stockholm, Sweden. No informed consent was required because, according to Swedish policy, response to the questionnaire constitutes consent. Zygosity was based on well-validated questions to twins and parents chosen from a discriminant analysis of 106 pairs with zygosity determined by DNA markers (Lichtenstein et al. Reference Lichtenstein, Tuvblad, Larsson and Carlstrom2007). Because levels of criminal registration were too low in females to produce stable model parameters, these analyses only included members of the male–male pairs: 442 complete male–male pairs of whom 259 were monozygotic (MZ) and 183 dizygotic (DZ). Of the 518 MZ twins, 424 had complete data, four were missing one variable, and 90 had suspect registry data only. Of the 366 DZ twins, 309 had complete data, three were missing one item, and 54 had suspect registry data only.
Measures
We used five self-report measures of externalizing traits and behaviors all completed by the twin at ages 16–17. The measures, along with three representative items and their internal consistency as assessed by Cronbach's α (Tavakol & Dennick, Reference Tavakol and Dennick2011), were:
(1) EXT: Externalizing items from the Child Behavior Checklist (Achenbach, Reference Achenbach1991): 19 items from the aggressiveness scale and six items from the delinquency scale, each scored 0–2: ‘I am louder than other kids’. ‘I lie or cheat’. ‘I threaten to attack others’. (Cronbach's α + 0.82.)
(2) SRD: The self-report delinquency scale (Junger-Tas et al. Reference Junger-Tas, Terlouw and Klein1994) consisting of 29 items, each beginning with ‘How many times in the past year have you …’ and scored 0–5: ‘How many times in the past year have you used fake ID?’ ‘How many times in the past year have you stolen something from school?’ ‘How many times in the past year have you beaten someone that didn't belong to your family so that this person had to seek medical care?’ (+0.92.)
(3) CLS: Youth Psychopathic Traits Inventory (YPI; Andershed et al. Reference Andershed, Kerr, Stattin, Levander, Blaauw and Sheridan2002) callous-unemotional scale consisting of 15 items (of which five assess remorselessness, five unemotionality, and five callousness). Each item is scored 0–3: ‘To be nervous or worried is a sign of weakness’. ‘When other people have problems, it is often their own fault, therefore, one should not help them’. ‘To feel guilty and remorseful about things you have done that have hurt other people is a sign of weakness’. (+0.89.)
(4) GRD: The YPI (Andershed et al. Reference Andershed, Kerr, Stattin, Levander, Blaauw and Sheridan2002) grandiosity-charm scale consisting of 20 items (of which five assess dishonest charm, five grandiosity, five lying, and five manipulation). Each item is scored 0–3: ‘I am better than anyone on almost anything’. ‘I am good at making people believe me when I make something up’. ‘It's easy for me to charm and seduce someone to get what I want from them’. (+0.78.)
(5) IMP: The YPI (Andershed et al. Reference Andershed, Kerr, Stattin, Levander, Blaauw and Sheridan2002) impulsivity-irresponsibility scale consisting of 15 items (of which five assess thrill seeking, five impulsiveness, and five irresponsibility). Each item is scored 0–3: ‘I like to do exciting or dangerous things even if it is forbidden or illegal’. ‘It often happens that I do things without thinking ahead’. ‘I have probably skipped school or work more than most other people’. (+0.84.)
To receive a score on a scale, twins had to answer at least 50% of the items. For those who did not answer all items, the mean response to the items answered was rescaled to the same range of values for those who answered all items. For statistical analysis, the five continuous variables were trichotomized into approximately equal-sized categories.
The TCHAD data were linked to the National Swedish Criminal Statistics reports on ‘persons suspected of offenses’ using the personal identification number unique to each citizen. We here defined CB as registration in this ‘Suspect Registry’. We used the number of dates on which the individual had entries between 6 December 1999 and 10 June 2011. Of the 884 male twins from this sample, 215 (24.3%) appeared in the suspect registry. Of these, 95 (44.2%) had their first appearance prior to 1 December 2002 (the approximate time the questionnaires were completed). The criminal suspect variable was trichotomized for analysis with 0 = no appearances (75.7%), 1 = one appearance (9.8%) and 2 = two or more appearances (14.5%).
To give a sense of the kinds of crimes in the suspect registry, we consulted the 2010 official crime report for Sweden. In that year, 126000 persons suspected of offenses were registered in Sweden in the following major categories: theft 17%, drugs 17%, violation of the person 16% (which includes assault and sexual offenses), traffic offenses 16% (most commonly driving under the influence), fraud 10%, vandalism 3%, and other crimes 20% (Ernbo & Petersson, Reference Ernbo and Petersson2011). Although it is not possible to link suspicions for crimes to actual convictions, approximately 63% of individuals in the suspect registry are eventually convicted for a crime within a 4-year period. The suspect registry does not include minor offenses such as parking violations or truancy.
Data analysis
The twin models used in these analyses decompose the sources of individual differences in liability to antisocial traits and behaviors into three components: additive genetic effects (A), shared environment (C) and non-shared environment (E) (Guerrini et al. Reference Guerrini, Cook, Kest, Devitgh, McQuillin, Curtis and Gurling2005). Shared environment reflects family, community and school experiences that increase similarity in twins raised together. Non-shared environment includes both environmental experiences not shared by twins and measurement error.
Our multivariate twin models estimated the degree to which genetic and environmental influences are shared across the five self-report measures of antisocial behavior and CB versus those influences that are specific to each individual variable. This was achieved by including in the model genetic and environmental common factors that influence risk for more than one variable in addition to variable specific influences. Independent pathway structural equation twin models were fitted using the full information maximum likelihood method in Mx (Neale et al. Reference Neale, Boker, Xie and Maes2003) because we were particularly interested in examining how genetic and shared environmental risk factors contributed differentially to patterns of covariance among our variables.
We began our model fitting with a basic 1-1-1 model, where the first, second and third numbers reflect the number of common factors genetic, shared environmental and non-shared environmental, and with genetic and environmental factors specific to each variable. We then sought to simplify the resulting model by deleting one by one all of the common factors, and then the variable specific genetic and shared environmental effects. (We did not attempt to eliminate the variable specific non-shared environmental effects as these include errors of measurement that could not plausibly be set to zero.) Next, we made the model progressively more complex while systematically searching at each step to find a ‘best-fitting’ model.
Our aim was to find the model that reflected the optimal balance between parsimony and explanatory power. This aim was operationalized by Akaike's Information Criterion (AIC; Akaike, Reference Akaike1987; Williams & Holahan, Reference Williams and Holahan1994), which equals χ2 − 2df, where df is the difference in the degrees of freedom of the two models. We sought to minimize the AIC value. If the best-fit model contained multiple common factors for A, C or E, the resultant factor loadings estimated by Mx were extracted and then rotated in SAS (SAS Institute, 2005) using a Varimax rotation to improve interpretability.
Results
Phenotypic associations
Using both logistic regression and polychoric correlation, we examined the association between our five self-report measures and CB as defined as presence in the Swedish Suspect Registry. The most strongly associated measure was SRD [odds ratio (OR) 2.35, p < 0.0001, r = + 0.42], followed by IMP (OR 2.29, p < 0.0001, r = + 0.34), EXT (OR 2.01, p < 0.0001, r = + 0.36), GRD (OR 1.61, p < 0.0001, r = + 0.25) and CLS (OR 1.47, p = 0.002, r= + 0.19).
Twin model fitting
Our first, or baseline, model (111_111) contained one genetic, one shared environmental and one non-shared environmental common factor and variable specific genetic, shared and non-shared environmental effects (Table 1). In models 2–4, we tried to eliminate the genetic, shared environmental and non-shared environmental common factors respectively. All of these models fitted more poorly than model 1. In models 5–7, we tried to eliminate the variable specific shared environmental, the variable specific genetic and both the genetic and shared environmental variable specific factors respectively. Of these, model 6 produced the lowest AIC value.
Table 1. Model fitting results for six self-report measures of externalizing traits and behaviors at ages 16–17 and registration in the Criminal Suspect Registry up to age 25
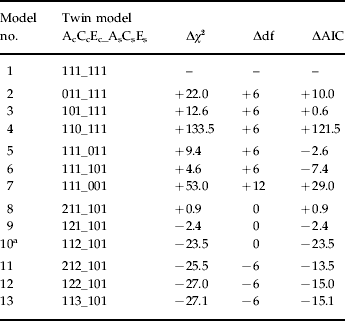
Δχ2, change in χ2 from model 1; Δdf, change in degrees of freedom from model 1; ΔAIC, change in Akaike's Information Criterion freedom from model 1.
Model 1: χ2 = 8113.47, df = 4535, AIC = −956.53.
a Best-fit model.
We then turned to adding new common factors to model 6. Models 8–10 added an additional genetic, shared environmental and non-shared environmental common factor. The AIC for model 10 was substantially superior to models 6, 8 or 9. In models 11–13, we then added to model 10 an additional genetic, shared environmental and non-shared environmental factor. None of these improved the model fit or even came very close to that observed from model 10, which therefore was our best-fit model.
Best-fit model
The best-fit model, the results of which are given in Table 2, has six noteworthy features. First, loadings of all the variables on the single genetic common factor were relatively homogeneous (in the range 0.35–0.55) with the exception of grandiosity, which loaded +0.71. This common factor explained 16% of the variance in risk for CB.
Table 2. Parameter estimates of the best-fit model for five self-report measures of externalizing traits and behaviors at ages 16–17 and registration in the Criminal Suspect Registry up to age 25
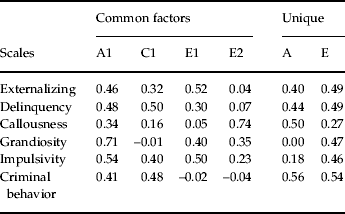
A1, first (and only) genetic common factor; C1, first (and only) shared environmental common factor; E1, first individual-specific environmental common factor; E2, second individual-specific environmental common factor. A and E refer to genetic and individual-specific environmental factors unique to the individual scales.
Second, loadings on the single shared environmental common factor could be divided into two groups. EXT, SRD, IMP and CB all loaded in the range 0.3–0.5. By contrast, CLS had a loading of only +0.16 and GRD a loading of zero. This common factor explained 23% of the variance in risk for CB.
Third, neither of the two non-shared environmental common factors loaded appreciably on registration in CB. That is, any environmental experiences that differed between these twins in mid-adolescence and impacted on their self-reported externalizing traits and behaviors had little or no effect on their risk for CB.
Fourth, there was, nonetheless, a coherent structure to these two non-shared environmental common factors. The two variables with highest loadings on the first factor were EXT and IMP. Some set of environmental experiences unique to each twin seemed to impact on this disinhibitory dimension of externalizing behavior. By far the highest loading on the second non-shared environmental common factor was for the CLS scale, reflecting an independent set of environmental experiences that influence the callousness dimension of externalizing behavior.
Fifth, variable specific unique genetic factors were important for all the variables in the model with the exception of grandiosity. For this variable only, all of its genetic variance was captured by the genetic common factor. Also of note, the variable with the strongest unique genetic loading was CB registration, where genetic influence accounted for 31% of the variance in risk.
Finally, the non-shared environmental variable specific loadings (which also reflect measurement error) were broadly similar (∼0.50) for five of six variables in the model and appreciably lower only for CLS.
Table 3 presents the results for the best-fit model in a complementary way. It can be seen that the estimated heritabilities for the self-report externalizing measures in mid-adolescence and criminal registration were all in a relatively narrow band from 32% to 50%. However, the extent to which these genetic risk factors arose from the common factor versus variable specific factors varied widely. Of note, two-thirds of the genetic risk for CB arose from unique genetic risk factors not shared with the self-report measures.
Table 3. The proportion of variation of each index of externalizing behavior due to genetic, shared environmental and individual-specific environmental factors in our best-fit model and the proportion of the variation within each category due to the effects of common factors versus index-specific influences
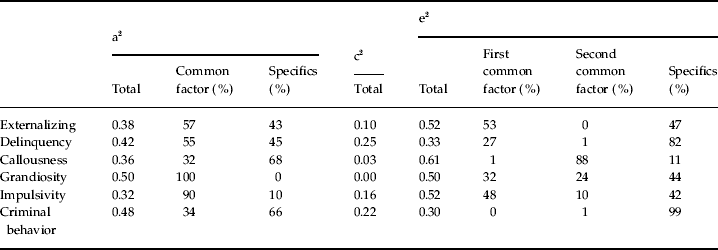
The contribution of shared environmental factors differed widely across variables and was highest for CB and SRD. The proportion of variance contributed by unique environmental effects also varied substantially and was lowest for CB. That is, familial factors (both genetic and shared–environmental) contributed 70% of the variance in risk for registration in the suspect registry.
Finally, we examined the percentage of familial covariance between our self-report measures and CB attributable to genetic versus shared environmental factors. (We focused on the familial covariance because of the trivial contribution of unique environmental factors to covariance between our self-report measures and CB.) This varied widely. The covariance between EXT and CB was 55% genetic and 45% shared environment. The corresponding figures for our other self-report measures were SRD 45%/55%, CLS 64%/36%, GRD 99%/1% and IMP 53%/47%.
Discussion
Using multivariate twin modeling, we examined genetic and environmental contributions to relationships between self-report measures of externalizing traits and behaviors assessed at age 16–17 and registration as a suspect for CB from the ages of 13 to 25. Our primary aim was to clarify, in a representative sample of Swedish male–male twin pairs, the relationships between the risk factors for antisocial traits and behaviors as assessed by self-report and for CB as assessed by registration in the Swedish suspect registry. Would some of the questionnaire measures provide better indices of the genetic risk for CB while others would prove more effective for indexing environmental contributions to risk?
Five major results from this investigation warrant particular emphasis. First, in accord with the meta-analytic results from Rhee & Waldman (Reference Rhee and Waldman2002) and with other reports examining sources of influence on these measures from the TCHAD sample (Eley et al. Reference Eley, Lichtenstein and Moffitt2003; Tuvblad et al. Reference Tuvblad, Eley and Lichtenstein2005; Larsson et al. Reference Larsson, Andershed and Lichtenstein2006; Forsman et al. Reference Forsman, Lichtenstein, Andershed and Larsson2008, Reference Forsman, Lichtenstein, Andershed and Larsson2010), we found all of the measures of antisocial traits and behaviors, including registration for arrests, to be moderately heritable. Indeed, the contribution of genetic influence to one of the self-report measures, GRD, slightly exceeded that for CB. This finding is important because it indicates a prominent role for constitutionally based individual differences in influencing self-report in addition to more objective indices of externalizing deviancy (Frisell et al. Reference Frisell, Pawitan, Langstrom and Lichtenstein2012).
Second, contrary to the trends reported in the meta-analysis of Rhee & Waldman (Reference Rhee and Waldman2002), we found evidence of shared environmental effects to be as strong or stronger for CB as for the self-report measures. This finding in our dataset may in part arise because CB for the current study sample was assessed from adolescence and young adulthood (as opposed to later adulthood), a time period when shared environmental influences on criminal behavior may be especially potent (Rhee & Waldman, Reference Rhee and Waldman2002). If so, the implication would be that the strength of shared environmental contributions to externalizing deviancy (including CB and impulsive antisocial tendencies more broadly) may be more related to age of assessment than to mode of measurement.
Third, in aggregate, our self-report measures reflected about a third of the genetic risk toward CB. Genetic influences accounted for a substantial proportion of the familial covariance of CB with each of the self-report measures, with the highest proportion observed for the GRD subscale of the YPI and the lowest evident for the SRD measure. The very strong contribution of genetic influence to the covariance between the GRD scale measure and CB is of interest in light of the prior findings of Larsson et al. (Reference Larsson, Andershed and Lichtenstein2006), who reported evidence, from this same sample, for a predominantly (>60%) genetic factor underlying the subscales of the YPI, on which GRD loaded most highly (0.75 versus 0.49 and 0.33 for IMP and CLS respectively). In conjunction with the current findings, the implication is that the well-documented association between psychopathic traits and criminal deviancy (Hare, Reference Hare2003; Cooke et al. Reference Cooke, Michie, Hart and Clark2004) is mediated in part by genes, and that the grandiose-manipulative facet of psychopathy as indexed by the YPI serves as a particularly effective index of the genetic component of this association. Nonetheless, it is noteworthy that two-thirds of the genetic variance in CB was not tapped by any of our self-report measures. This could reflect some inadequacy of the scales that we used or perhaps limitations inherent to self-report more generally, especially for the assessment of externalizing psychopathology. Alternatively, our results could reflect in part changes in the genetic substrate for CB from the time of mid-adolescence (when we obtained our self-report) to that of early adulthood, the period over which we assessed CB.
Fourth, in our best-fitting model, the self-report measures were much better at tapping the shared environmental influences on CB. Here, the scales that performed best were the SRD and IMP. The finding for SRD, which among the self-report measures is most ostensibly similar to that for CB, suggests that the actual performance of antisocial acts, whether measured objectively by police records or by self-report, is strongly influenced by family, peer and/or school effects. Alternatively, the MZ and DZ twin brothers could be frequently committing the antisocial acts together. The finding for the IMP subscale of the YPI, which reflects impulsivity, irresponsibility and thrill seeking, suggests likewise that the impulsive behavioral style component of psychopathy (cf. Cooke & Michie, Reference Cooke and Michie2001) as assessed by self-report is particularly susceptible to shared environmental influence.
Fifth, our self-report variables failed to index the non-shared environmental effects that predispose to CB. Some proportion of any estimate of non-shared environment in a cross-sectional twin study reflects errors of measurement. If, in fact, error accounted for a very high proportion of the observed individual specific environment for arrest registration, this might explain these results. However, CB was assessed over a 12-year period whereas our self-report measures were obtained at a single point in time (at age 16–17). If individual specific environmental effects on externalizing behaviors are time limited in their effect, it might be expected that we would find little correspondence in their impact on CB relative to self-report.
In sum, we found strong evidence for variation in the degree to which the five self-report measures of externalizing traits and behavior were able to index the genetic versus the shared environmental risk factors for CB. In particular, we found that GRD and to a somewhat lesser extent CLS tapped the genetic vulnerability to CB. By contrast, SRD, which reflected genetic risk more weakly, emerged as the best measure for assessing the shared environmental risk to CB. EXT and IMP were the most balanced indicators of etiologic risk, with nearly equal proportions of the observed covariation for these measures reflecting genetic and shared environmental influences. However, it is also important to note that two-thirds of the genetic variance for CB was not indexed by any of the self-report measures used.
Limitations
These results should be interpreted in the context of five potential methodological limitations. First, our sample was restricted to twins born in Sweden. The degree to which these results would extrapolate to other populations is uncertain. Second, we focused exclusively on male participants. Females had to be excluded from the analysis because rates of CB among female participants in the current sample were too low to produce stable model estimates. Nonetheless, it will be important in future research to make efforts to surmount this limitation to better understand the etiologic determinants of criminal/antisocial behavior and affiliated personality traits in women at differing ages.
A third limitation of the current study is that a single aggregate measure of CB was examined rather than indices of more specific crime types. This approach was used because of the modest base rates of CB in the current epidemiological sample and the requirements of power for biometric analyses. In view of evidence for distinct etiologic contributions to crimes of differing types in Sweden (Frisell et al. Reference Frisell, Lichtenstein and Langstrom2011), it will be valuable to also address this limitation in future work involving larger samples.
Fourth, although we obtained cooperation from 83% of all subjects, the sample was not entirely representative. Not surprisingly, those who did not cooperate were nearly twice as likely to have a criminal registration as those who did (34.7% v. 18.7%; OR 0.54, p = 0.002). However, the Mx modeling used full information maximum likelihood with a ‘missing at random’ assumption, which would operate to minimize the bias introduced.
Fifth, we examined registration for criminal suspects rather than convictions because it provided higher base rates and greater statistical power. However, a limitation to this approach is that it entails the likelihood of some false positives that could conceivably have biased parameter estimates in our model.
Acknowledgments
This research was supported by the Swedish Council for Working Life and Social Research (project 2004-0383), the Swedish Research Council (2004-1415), the Ellison Medical Foundation, and the National Institute of Mental Health (MH089727).
Declaration of Interest
None.



