


INTRODUCTION
Because conservation on private lands can contribute to achieving national conservation goals (Watkins et al. Reference Watkins, Barret, Smith and Paine1996; Fisher & Dills Reference Fisher and Dills2012), in countries like South Africa, Zimbabwe and Namibia, private landowners have been granted some control over the use of wildlife (Child Reference Child, Prins, Grootenhuis and Dolan2000). In South Africa, private protected areas are more numerous and more extensive than public protected areas (Watkins et al. Reference Watkins, Barret, Smith and Paine1996); over 5000 game ranches and c. 4000 mixed game and livestock ranches cover c. 13% of the country's total land area; officially declared conservation areas cover 5.8% of the land area (Hearne et al. Reference Hearne, Korrûbel and Koch2000; Krug Reference Krug2001; ABSA [Amalgamated Banks of South Africa] 2003). Moreover, private land acquisition for biodiversity protection is often in line with the scientific prioritization of areas for conservation (Fisher & Dills Reference Fisher and Dills2012), although spatial planning using demographic, motivational and management variables can improve achievement of conservation priorities on private lands (Wallace et al. Reference Wallace, Theobald, Ernst and King2008).
Wildlife has multiple uses that generate revenues for the betterment of the landowners. For example, wildlife is used non-consumptively (for example for tourism, photographic safaris, or wildlife viewing) or consumptively (for example for hunting for trophies, and for selling meat and/or hide). Landowners can concentrate on consumptive or non-consumptive use of wildlife, or a mixture of both (Aylward Reference Aylward, Aylward and Lutz2003; de Boer et al. 2007). Game viewing produces a number of revenue sources for the game reserve management, such as entrance fees, accommodation charges, sales from restaurants and shops, trails, rides, tour income, or revenues from permits and fines (Aylward Reference Aylward, Aylward and Lutz2003). Game viewing is the major reason why tourists visit game reserves (see Vial Reference Vial1996; Langholz & Kerley Reference Langholz and Kerley2006), and the number of tourists tends to increase with the probability of seeing the principal species (elephant Loxodonta africana, buffalo Syncerus caffer, rhino Ceratotherium spp., lion Panthera leo, and leopard Panthera pardus; Lindsey et al. Reference Lindsey, Alexander, Mills, Romanach and Woodroffe2007; Okello et al. Reference Okello, Manka and D'Amour2008).
Hunting plays a major role in generating income for landowners or game reserve managers involved in consumptive use (Barnett & Patterson Reference Barnett and Patterson2006). In South Africa, how much hunting landowners can allow to be done on their land is based on quotas set by provincial nature conservation authorities, generally determined by the availability of species in terms of densities (APNR [Associated Private Nature Reserves] 2005; Barnett & Patterson Reference Barnett and Patterson2006). Based on the quotas set, the landowner can sell the hunting licences to hunters, who then come to hunt. Like other forms of wildlife control, hunting is under increasing scrutiny from various interest groups. There has been lobbying against hunting practices by animal-welfare and rights groups, particularly for charismatic species like elephants (Peterson Reference Peterson2004; Burke et al. Reference Burke, Page, van Dyk, Millspaugh and Slotow2008; Lötter et al. Reference Lötter, Henley, Fakir, Pickover, Ramose, Scholes and K.G2008; Kettles & Slotow Reference Kettles and Slotow2009). Negative perceptions of management interventions can also potentially reduce public appeal, and may adversely affect tourism (Burke et al. Reference Burke, Page, van Dyk, Millspaugh and Slotow2008; Lötter et al. Reference Lötter, Henley, Fakir, Pickover, Ramose, Scholes and K.G2008). Hunting not only reduces animal numbers, it likely also negatively affects the visibility or level of habituation of the remaining animals (Leader-Williams & Hutton Reference Leader-Williams, Hutton, Woodroffe, Thirgood and Rabinowitz2005; Reimers et al. Reference Reimers, Loe, Eftestøl, Colman and Dahle2009). This is an important concern where hunting and tourism occurs on shared land (Reimers et al. Reference Reimers, Loe, Eftestøl, Colman and Dahle2009). Non-consumptive uses may also affect wildlife in several ways that might be termed indirect consumptive uses, such as through lowered productivity, indirect mortality, reduced use of refuge, aberrant behaviour, or stress (Pomerantz et al. Reference Pomerantz, Decker, Goff and Purdy1988). However, non-consumptive wildlife uses could be less detrimental if viewing and tourism are undertaken in natural settings, as in parts of southern Africa, through ensuring that there is little disturbance to the natural wildlife habitats and that both wildlife habitat and population are large enough (Novelli et al. Reference Novelli, Barnes and Humavindu2006).
Several models have been used to analyse interactions between ecological and economic factors in wildlife management, mainly dealing with land competition between wildlife and livestock production (see Schulz & Skonhoft Reference Schulz and Skonhoft1996; Kinyua et al. Reference Kinyua, van Kooten and Bulte2000; Fleming & Alexander Reference Fleming and Alexander2003). These studies came up with different strategies on how to optimize land allocation amongst these competing land uses, wildlife and livestock, but there has been no investigation of the competition between wildlife hunting and wildlife tourism, even though it appears to be a common perception that the two activities do not mix (E. Mwakiwa, personal observation 2006; WWF [World Wildlife Fund] 2001; Meletis & Campbell Reference Meletis and Campbell2007; Baldus, et al. Reference Baldus, Damm and Wollscheid2008; Reimers et al. Reference Reimers, Loe, Eftestøl, Colman and Dahle2009).
There are three main types of private landownership involved in wildlife management in southern Africa, namely game ranching (raising of animals for a variety of products, including meat, hides, feathers and antlers), private conservancy (wildlife landowners pooling natural and financial resources to conserve wildlife, usually operating through a joint management committee), and private nature reserves, also known as game reserves (Krug Reference Krug2001). The forming of groups by landowners (Pasquini et al. 2009) or multilateral governance initiatives (Paloniemi & Tikka Reference Paloniemi and Tikka2008) can contribute to reaching national conservation goals and solve conflicts of interest. Management objectives of game reserves vary from preservation to sustainable use of wildlife. Wildlife use involves obtaining profits from both consumptive and non-consumptive uses (Huffaker Reference Huffaker1993; Hearne & McKenzie Reference Hearne, McKenzie, Prins, Grootenhuis and Dolan2000; ABSA 2003).
To investigate the economic effects of consumptive and non-consumptive use of wildlife, we formulated a mathematical model, partly based on South Africa's Associated Private Nature Reserves (1850 km2), which consist of Timbavati, Klaserie, Umbabat and Balule Private Nature Reserves. Each reserve comprises land belonging to a number of owners, and has its own management structure in place. Landowners in each reserve contractually devote their land to wildlife conservation, so they cannot convert to land-use practices other than wildlife management. Landowners in a game reserve can generally be categorized as commercial or non-commercial. Commercial landowners are involved in tourism and have lodges on their properties; they provide accommodation and associated tourism services (Eagles Reference Eagles2001). Non-commercial landowners have their land for their own recreation, its aesthetic appeal, a place of retirement, an investment, or to contribute to wildlife conservation; they usually have other sources of income, and this and other circumstances influence their willingness to participate in conservation activities (Knight et al. Reference Knight, Grantham, Smith, McGregor, Possingham and Cowling2011).
We investigated how the game reserve management decided on how much land to devote to hunting and how much to tourism or a combination of both. To do this, we employed three scenarios: (1) the hunting and tourism areas are separate but wildlife is shared (partially separated); (2) hunting and tourism co-exist in the same area (not separated); and (3) the hunting and tourism areas are separated by a fence (completely separated).
METHODS
Models
We modelled the management for a game reserve that would like to decide how much land is to be used for wildlife non-consumptive and/or consumptive uses. The game reserve management makes profit from the number of hunters and/or tourists they attract to their game reserve. Therefore, the objective of the management is to maximize the net revenue from a combination of both wildlife consumptive and non-consumptive uses.
Model for scenario 1
Scenario 1, where there is no barrier between the two areas and wildlife roams freely between them, is common in many game reserves in South Africa. The proportion of land under hunting is denoted by A and that under tourism is denoted by L. Land base is fixed, so the relationship between hunting and tourism land-uses is as follows:
 $$\begin{equation}
L + A = 1
\end{equation}$$
$$\begin{equation}
L + A = 1
\end{equation}$$
We assume that there is one wildlife species. The dynamics for this species is given by:
 $$\begin{equation}
\frac{{dX}}{{dt}} = g\left( X \right) - qEX
\end{equation}$$\vskip-2pt
$$\begin{equation}
\frac{{dX}}{{dt}} = g\left( X \right) - qEX
\end{equation}$$\vskip-2pt
where: X denotes the wildlife stock at time t (for notational convenience, we suppress the time notation, but time should be understood to be implicit in all variables); g(X) is the growth function of wildlife, which is dependent on wildlife stock; g(X) is a logistic function with the formulationg(X) = τX[1 − X/K], where τ is the wildlife's intrinsic growth rate and K is the carrying capacity. We adopt the notation where the subscript denotes the derivative with respect to that variable. From the formulation: g(X) > 0, gX
(X) > 0 for
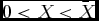 $0 < X < \bar X$
, and gX
(X) < 0 for
$0 < X < \bar X$
, and gX
(X) < 0 for
 $X > \bar X$
(
$X > \bar X$
(
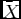 $\bar X$
is the value of X when gX
(X) = 0); gXX
(X) < 0. qEX is the standard Schaefer catch term. In this case, q, the ‘catchability’ coefficient, is the fraction of the stock (namely X) that is ‘caught’ (shot or hunted) by one hunter per unit time. By definition q is less than 1. E is the number of hunters at any time t. For simplicity and traceability of the model, we ignore age and sex structure, although we understand that this could have some bearing on the results.
$\bar X$
is the value of X when gX
(X) = 0); gXX
(X) < 0. qEX is the standard Schaefer catch term. In this case, q, the ‘catchability’ coefficient, is the fraction of the stock (namely X) that is ‘caught’ (shot or hunted) by one hunter per unit time. By definition q is less than 1. E is the number of hunters at any time t. For simplicity and traceability of the model, we ignore age and sex structure, although we understand that this could have some bearing on the results.
Assuming the game reserve management derives its income from selling hunting licences and tourism, its profit function (ε) is given by:
 $$\begin{equation}
\varepsilon = \eta {A^\alpha }{X^\beta }E + T{L^\alpha }{X^\beta }
\end{equation}$$
$$\begin{equation}
\varepsilon = \eta {A^\alpha }{X^\beta }E + T{L^\alpha }{X^\beta }
\end{equation}$$
For the first term on the right hand side of Eq. (3): ηA α X β Eis the game reserve management unit net profit per hunter (US$ hunter–1), which is a Cobb-Douglas function. The unit net profit per hunter depends on: the charge per hunter (η measured in US$); the wildlife stock X, the land area under hunting activity A; and number of hunters, E. As landowners, the game reserve management make a profit on how long a hunter spends hunting on their land. The number of wildlife the hunters shoot will be determined by the Schaefer catch model (the last term in Eq. 2). We assume that hunters will be prepared to pay more to hunt in a bigger area (A) and also to hunt where there are more animals (namely where X is larger); then the unit profit per hunter is an increasing function of X and A. As for the other parameters for the unit net profit function from tourism: α is the parameter for land; β is the parameter for number for wildlife. One of the specifications is that α + β < 1, thus representing decreasing marginal ‘productivity’ as land and animals increase.
The second term on the right hand side of Eq. (3) represents the unit net profit per tourist visiting to view wildlife. The unit net profit per tourist function is also given by a Cobb-Douglas function. This profit depends on the net charge per tourist, T, paid by the tourist to view wildlife, a constant (measured in US$), size of land under tourism (L) and number of wildlife (X). The specification of the unit net profit per tourist function meets the following assumptions: with increasing numbers of wildlife, the profit from tourism will rise as more tourists are attracted (Lindsey et al. Reference Lindsey, Alexander, Mills, Romanach and Woodroffe2007; Okello et al. Reference Okello, Manka and D'Amour2008); larger areas under tourism would attract more tourists, since it would give them a larger area for wildlife viewing (Hearne & McKenzie Reference Hearne, McKenzie, Prins, Grootenhuis and Dolan2000) based on the assumption that the larger the land property, the higher the diversity of wildlife species, and the higher the landscape diversity for tourists (Aylward Reference Aylward, Aylward and Lutz2003; H. Killian, personal communication 2006). For model simplicity we assume that there is uniform mixing of wildlife between the hunting and tourism areas (Sanchirico & Wilen Reference Sanchirico and Wilen1999).
We assume that the objective of the game reserve management is to maximize the present value of the unit net profits from hunting and tourism, as given by:
 $$\begin{equation}
\mathop {Max}\limits_{E,A} PV = \int\limits_0^\infty {\left[ {\eta {A^\alpha }{X^\beta }E + T{L^\alpha }{X^\beta }} \right]} {e^{ - rt}}dt
\end{equation}$$
$$\begin{equation}
\mathop {Max}\limits_{E,A} PV = \int\limits_0^\infty {\left[ {\eta {A^\alpha }{X^\beta }E + T{L^\alpha }{X^\beta }} \right]} {e^{ - rt}}dt
\end{equation}$$
where r is the economic discount rate. We assume thatr > 0.
Maximization takes place subject to Eqs (1) and (2), so that the current value Hamiltonian H (Clark Reference Clark2005) for this problem is as follows:
 $$\begin{equation}
H = \eta {A^\alpha }{X^\beta }E + T{L^\alpha }{X^\beta } + \mu \left[ {g\left( X \right) - qEX} \right]
\end{equation}$$
$$\begin{equation}
H = \eta {A^\alpha }{X^\beta }E + T{L^\alpha }{X^\beta } + \mu \left[ {g\left( X \right) - qEX} \right]
\end{equation}$$
with X as the state variable, A and E as the control variables, and μ the costate variable for the wildlife stock.
Model for scenario 2
In this scenario, we explore the case where the game reserve management is undertaking both hunting and tourism on the same area. The Hamiltonian function in this case would be:
 $$\begin{equation}
H = \eta {\Pi ^\alpha }{X^\beta }E + T{\Pi ^\alpha }{X^\beta } - ES + \mu \left[ {g\left( X \right) - qEX} \right]
\end{equation}$$
$$\begin{equation}
H = \eta {\Pi ^\alpha }{X^\beta }E + T{\Pi ^\alpha }{X^\beta } - ES + \mu \left[ {g\left( X \right) - qEX} \right]
\end{equation}$$
where S is the unit damage cost caused by hunting on tourism. Although it could be argued that both hunting and tourism cause some disturbance to wildlife, we assume here that hunting causes direct loss of wildlife (Pomerantz et al. Reference Pomerantz, Decker, Goff and Purdy1988). However, we assume that tourism is being done in natural settings, and as such is causing little disturbance to the natural wildlife habitats (Novelli et al. Reference Novelli, Barnes and Humavindu2006). Therefore for simplicity we assume that hunting is the only land use coming with an ecological cost. We assume hunting is perceived negatively by tourists, and therefore reduces tourism due to its negative effects on the number of game and its visibility (Hearne & McKenzie Reference Hearne, McKenzie, Prins, Grootenhuis and Dolan2000; Burke et al. Reference Burke, Page, van Dyk, Millspaugh and Slotow2008; Lötter et al. Reference Lötter, Henley, Fakir, Pickover, Ramose, Scholes and K.G2008; Reimers et al. Reference Reimers, Loe, Eftestøl, Colman and Dahle2009). In addition, Π is the total area of the game reserve and as such Π = 1.
Model for scenario 3
In this scenario, the hunting area is completely separated from the tourism area by a fence, so there are two separate wildlife populations that do not mix. The relationship between hunting area, A, and tourism area, L, is still as defined by Eq. (1). The wildlife dynamics for the area under hunting are given by:
 $$\begin{equation}
\frac{{d\Phi }}{{dt}} = g\left( \Phi \right) - qE\Phi
\end{equation}$$\vskip 4pt
$$\begin{equation}
\frac{{d\Phi }}{{dt}} = g\left( \Phi \right) - qE\Phi
\end{equation}$$\vskip 4pt
where Φ is the wildlife stock in the game reserve section where hunting is practised.
The wildlife dynamics for the area under tourism is given by:
 $$\begin{equation}
\frac{{d\Omega }}{{dt}} = g\left( \Omega \right)
\end{equation}$$
$$\begin{equation}
\frac{{d\Omega }}{{dt}} = g\left( \Omega \right)
\end{equation}$$
where Ω is the wildlife stock in the game reserve section where tourism is being undertaken.
The profit function from tourism is now given by TL αΩβ. However, the profit function from hunting is given by ηA αΦβ E.
The Hamiltonian in this case is given by:
 $$\begin{equation}
H = \eta {A^\alpha }{\Phi ^\beta }E + \mu \left[ {g\left( \Phi \right) - qE\Phi } \right] + T{L^\alpha }{\Omega ^\beta }
\end{equation}$$
$$\begin{equation}
H = \eta {A^\alpha }{\Phi ^\beta }E + \mu \left[ {g\left( \Phi \right) - qE\Phi } \right] + T{L^\alpha }{\Omega ^\beta }
\end{equation}$$
Numerical analyses
To substantiate some of the analytical model results and further explorations, especially for scenarios 1 and 2 described above, we undertook some simple numerical exercises. Our concern in the numerical analyses was not to model the precise tourism and hunting functions but to explore the general implications of interactions between these two land uses when selected parameter values change. Hence the parameters of the model (Table 1) were not estimated. In one case, we made some slight modifications to three parameters (τ = 0.005; q = 0.8; α = 0.001) for clarity (see later).
Table 1 Parameters and their values.

RESULTS
Analytical results
Hunting-tourism: partially separated
Based on Eq. (5), it can be shown (Clark & Munro Reference Clark and Munro1975; Conrad & Clark Reference Conrad and Clark1987; Clark Reference Clark2005; see Supplementary material) that the optimal harvest strategy will be the one that brings wildlife stock (X), to the steady state level in this scenario, X*1, given by the modified golden rules (Eqs 10 and 11).
 $$\begin{equation}
r = {g_X}\left( X \right) + \frac{{g\left( X \right)\beta }}{X} + \frac{{q\beta T{L^\alpha }}}{{\eta {A^\alpha }}} - \frac{{g\left( X \right)}}{X}
\end{equation}$$
$$\begin{equation}
r = {g_X}\left( X \right) + \frac{{g\left( X \right)\beta }}{X} + \frac{{q\beta T{L^\alpha }}}{{\eta {A^\alpha }}} - \frac{{g\left( X \right)}}{X}
\end{equation}$$
 $$\begin{equation}
T{L^{\alpha - 1}} = \eta {A^{\alpha - 1}}E
\end{equation}$$
$$\begin{equation}
T{L^{\alpha - 1}} = \eta {A^{\alpha - 1}}E
\end{equation}$$
On the left hand side of Eq. (10), r represents the economic discount rate prevailing on the market. The argument is that for wildlife to be competitive, what we here term the economic return on wildlife (represented by the right hand side of Eq. 10) must equal the discount rate. If wildlife is not competitive, then management would decide to invest elsewhere where they can get higher economic returns. In most cases wildlife investments are insufficiently economically competitive, which explains why wildlife species in protected areas might be endangered (Barnes Reference Barnes1996; Bulte & van Kooten Reference Bulte and van Kooten1999; Fleming & Alexander Reference Fleming and Alexander2003). The issue of importance here is therefore to find ways of making wildlife investments competitive economically so as to promote wildlife conservation.
The terms appearing on the right hand side of Eq. (10), which we have already referred to as the economic return on wildlife, comprise elements that enhance or lower the wildlife economic competitiveness for the game reserve management. The economic return on wildlife comprises the following terms: (1) marginal wildlife productivity (gX
(X)), (2) the ‘growth rate & wildlife parameter product: population’ ratio (
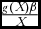 $\frac{{g( X )\beta }}{X}$
), (3) ‘hunting and tourism effects’ (
$\frac{{g( X )\beta }}{X}$
), (3) ‘hunting and tourism effects’ (
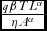 $\frac{{q\beta T{L^\alpha }}}{{\eta {A^\alpha }}}$
), and (4) the ‘growth rate: population’ ratio (
$\frac{{q\beta T{L^\alpha }}}{{\eta {A^\alpha }}}$
), and (4) the ‘growth rate: population’ ratio (
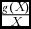 $\frac{{g( X )}}{X}$
).
$\frac{{g( X )}}{X}$
).
Implications of scenario 1
From Eq. (10), hunting and tourism have a positive effect on the economic return on wildlife through the term ‘hunting and tourism effects’, (
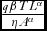 $\frac{{q\beta T{L^\alpha }}}{{\eta {A^\alpha }}}$
). For wildlife to be competitive, the following components of the ‘hunting and tourism effects’ should be relatively high: catchability coefficient, wildlife parameter, charge per tourist and proportion of land allocated to tourism, which are all components of the numerator. Conversely, the components of the denominator, namely charge per hunter and proportion of land under hunting should be relatively low. This implies that although hunting area should be kept relatively low (because hunting is consumptive), the management should attract hunters through low charges and hunting should be made relatively easy for the hunter through a high catchability coefficient. This means that both tourism and hunting land uses might add to the enhancement of viability of wildlife stock, unlike in a situation where one of the land uses is undertaken on its own.
$\frac{{q\beta T{L^\alpha }}}{{\eta {A^\alpha }}}$
). For wildlife to be competitive, the following components of the ‘hunting and tourism effects’ should be relatively high: catchability coefficient, wildlife parameter, charge per tourist and proportion of land allocated to tourism, which are all components of the numerator. Conversely, the components of the denominator, namely charge per hunter and proportion of land under hunting should be relatively low. This implies that although hunting area should be kept relatively low (because hunting is consumptive), the management should attract hunters through low charges and hunting should be made relatively easy for the hunter through a high catchability coefficient. This means that both tourism and hunting land uses might add to the enhancement of viability of wildlife stock, unlike in a situation where one of the land uses is undertaken on its own.
The component ‘growth rate & wildlife parameter product:population’ ratio (
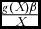 $\frac{{g( X )\beta }}{X}$
), has an additive effect on the economic return on wildlife, whilst the component ‘growth rate: population’ ratio (
$\frac{{g( X )\beta }}{X}$
), has an additive effect on the economic return on wildlife, whilst the component ‘growth rate: population’ ratio (
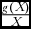 $\frac{{g( X )}}{X}$
) has a negative effect on the economic return on wildlife. This implies that, to make wildlife competitive, the marginal returns from wildlife, through the wildlife parameter β, should be as high as possible.
$\frac{{g( X )}}{X}$
) has a negative effect on the economic return on wildlife. This implies that, to make wildlife competitive, the marginal returns from wildlife, through the wildlife parameter β, should be as high as possible.
Equation (11) gives the equilibrium condition between demand for the two land uses, namely tourism and hunting. The function TL α − 1 on the left hand side of Eq. (11) represents the demand of land for tourism. Furthermore ηA α − 1 E on the right hand side of Eq. (11) is the demand function of land for hunting. The intersection of these two functions gives the equilibrium amount of land that should be allocated to hunting, A, (hence, the remainder would be for tourism, L). This intersection would also give the optimum opportunity cost of land, OP*, in other words, the optimum wildlife stock (see later).
Hunting-tourism: not separated
For the scenario where the game reserve management is undertaking hunting and tourism on the same area, the optimal harvest strategy will be the one that brings wildlife stock (X), to the steady state level in this scenario, X*2, given by the modified golden rules (Eqs 12 and 13).
 $$\begin{equation}
r = {g_X}\left( X \right) + \frac{{g\left( X \right)\eta \beta }}{{{X^{1 - \beta }}\left( {\eta {X^\beta } - S} \right)}} + \frac{{q\beta T{X^\beta }}}{{\left( {\eta {X^\beta } - S} \right)}} - \frac{{g\left( X \right)}}{X}
\end{equation}$$
$$\begin{equation}
r = {g_X}\left( X \right) + \frac{{g\left( X \right)\eta \beta }}{{{X^{1 - \beta }}\left( {\eta {X^\beta } - S} \right)}} + \frac{{q\beta T{X^\beta }}}{{\left( {\eta {X^\beta } - S} \right)}} - \frac{{g\left( X \right)}}{X}
\end{equation}$$
 $$\begin{equation}
0 = \alpha {X^\beta }\left( {T + \eta E} \right)
\end{equation}$$
$$\begin{equation}
0 = \alpha {X^\beta }\left( {T + \eta E} \right)
\end{equation}$$
In Eqs (12) and (13), we have taken into consideration that the proportion of land for both hunting and tourism is equal to 1 (Π = 1) in this scenario. If we compare the right hand side of Eq. (10) in scenario 1, the economic return on wildlife for scenario 1 (ERW1 in Fig. 1 a), with that of Eq. (12), the economic return on wildlife in scenario 2 (ERW2 in Fig. 1 a), an interesting observation can be made. The steady state of wildlife biomass in scenario 1 (X*1) is smaller than the optimum biomass in scenario 2 (X*2) (Fig. 1 a). The undertaking of hunting and tourism in the same area comes with a cost, S, the unit damage cost caused by hunting on tourism.

Figure 1 (a) The optimum wildlife stock when the effect of the unit damage cost caused by hunting on tourism (S) is zero and greater than zero; (b) Comparison of the optimum wildlife numbers for scenarios where hunting and tourism areas are partially separated (X*1) not separated (X*2) completely separated (Φ*3) and their respective economic return on wildlife (ERW) curves; (c) The effect of shift by the same magnitude of the demand curves for hunting (HD1) and for tourism (TD1) when the curves have the same slope; (d) The effect of shift by the same magnitude of the demand curves for hunting (HD1) and for tourism (TD1) when the curve for hunting (HD1) is steeper than that for tourism (TD1).
Undertaking hunting and tourism in the same area has two effects on the economic return on wildlife (compare Eqs 10 and 12). The first effect is that it increases the ‘growth rate & wildlife parameter product:population’ ratio as compared to the situation in scenario 1, provided S is small enough to keep the denominator positive. The second effect is that it increases the ‘hunting and tourism effects’ provided S is very small, otherwise ‘hunting and tourism effects’ becomes negative. S has a geometric effect on the economic return on wildlife curve. We let ERW(S = 0) andX* S = 0, to denote the economic return on wildlife and optimum wildlife stock, respectively, whenS = 0, and ERW(S > 0) and X* S > 0 to denote the economic return on wildlife and optimum wildlife stock, respectively, whenS > 0. S causes the economic return on wildlife curve to shift outwards from ERW(S = 0) to ERW(S > 0), and the wildlife stock increases from X* S = 0 to X* S > 0 (Fig. 1 a). Hence, S causes an increase in optimal wildlife stock as the landowner tries to maintain both hunting and tourism in the same area (Fig. 1 a, b).
Hunting-tourism: completely separated
In this scenario, the hunting area is completely separated from the tourism area by a fence. The optimal harvest strategy that will result in the wildlife stock steady state level,Φ* in the hunting section is given by the modified golden rules (Eqs 14 and 15). However, the optimum wildlife stock in the tourism section is given by Ω*, which we do not model explicitly here. However, because there is no hunting, the size of the wildlife stock here is assumed to be at its carrying capacity.
 $$\begin{equation}
r = {g_\Phi }\left( \Phi \right) + \frac{{\beta g\left( \Phi \right)}}{\Phi } - \frac{{g\left( \Phi \right)}}{\Phi }
\end{equation}$$
$$\begin{equation}
r = {g_\Phi }\left( \Phi \right) + \frac{{\beta g\left( \Phi \right)}}{\Phi } - \frac{{g\left( \Phi \right)}}{\Phi }
\end{equation}$$
 $$\begin{equation}
T{L^{\alpha - 1}}{\Omega ^\beta } = \eta {A^{\alpha - 1}}{\Phi ^\beta }E
\end{equation}$$
$$\begin{equation}
T{L^{\alpha - 1}}{\Omega ^\beta } = \eta {A^{\alpha - 1}}{\Phi ^\beta }E
\end{equation}$$
Comparison of the economic return on wildlife, in scenario 3 (Eq. 14), depicted by ERW3 curve (Fig. 1 b), to scenario 1 (Eq. 10), depicted by ERW1 (Fig. 1 b), shows that the economic return on wildlife curve would shift downwards, implying that the wildlife stocks per unit area in the hunting section of the game reserve would drop (Fig. 1 b). However, in the tourism section, the wildlife per unit area would be higher than in the hunting area. The wildlife stock in the tourism area would be kept at carrying capacity.
The right hand side of Eq. (15) shows the demand function for land for hunting. If a comparison of the demand for land for hunting is made between scenario 1 (right hand side of Eq. 11) and scenario 3 (right hand side of Eq. 15), then the following can be deduced: the demand for land for hunting in the former scenario is higher than in the latter scenario, provided hunting dominates tourism.
Scenarios 1 and 3 have differing effects on the demand schedules for land for hunting versus tourism depending on which use is dominating (Fig. 1 c, d). To clearly illustrate the point, we assumed that the curves for both the demand for land for tourism and hunting were straight lines: for scenario 1 represented by TD1 and HD1, respectively, and for scenario 3, TD3 and HD3, respectively. We analysed the effects when the demand schedule for hunting was relatively steeper and flatter on opportunity cost of land and the area of land allocated to hunting (conversely to tourism), as we moved from scenario 3 (full separation of hunting and tourism areas) to scenario 1 (partial separation where only wildlife is shared).
When the demand schedule for hunting land is steeper (implying that hunting is dominating; Fig. 1 d), then a shift in both of the demand curves would favour allocation of more land to hunting at the expense of tourism. However, if the demand curves have the same absolute slope, then if there is a shift in the demand curves by the same magnitude as in the earlier case, there would be no change in allocation of land between hunting and tourism, but the opportunity cost of land would shift upwards (Fig. 1 c) implying increased wildlife stock. Thus, as we shift from scenario 3 (complete separation of hunting from tourism), to scenario 1 (hunting and tourism occurring in separate areas, but sharing wildlife stocks), the opportunity cost of land increases, but whether tourism or hunting is allocated more land depends on which activity dominates.
Numerical results
Scenario 1: hunting area and tourism areas separated, but wildlife shared
The demand for land for tourism (TD) and the demand for land for hunting (HD) derived from the left hand side and right hand side of Eq. (11), respectively, intersect to give the amount of land that should be devoted to hunting, A, (and hence, the remainder is assumed to be for tourism, L), given the optimum opportunity cost of land, OP* measured in terms of wildlife stock (Fig. 2 a).

Figure 2 Scenario where hunting and tourism are separate, but wildlife is shared. (a) The intersection of the demand curves for: land for hunting and land for tourism; (b) the relationship between the number of hunters and wildlife numbers; (c) comparison of the economic return on wildlife when the proportion of land allocated to hunting is 0.1 and 0.8.
There is a negative relationship between the number of hunters and equilibrium wildlife numbers (Fig. 2 b). Thus, as more hunters are attracted by the game reserve, wildlife stocks decrease. In our model, 50 hunters or more would result in the local extinction of the wildlife in the game reserve.
We modelled how many hunters would be required for the game reserve management to realize a positive return from wildlife given different proportions of land allocated to hunting. In our model, when the land allocated to hunting was 0.1, it would require at least 22 hunters to achieve a positive economic return from wildlife (Fig. 2 c). However, if the proportion of land allocated to hunting is increased to 0.8, then at least 40 hunters are required for the game reserve to realize a positive economic return from wildlife (Fig. 2 c). This means that, if the game reserve management decided to allocate extra land to hunting, then they would need to attract more hunters to achieve positive economic returns from their wildlife investment.
We can also depict the different combinations of the proportion under hunting and the number of hunters that would give the same return to wildlife investment for the game reserve management through a contour and three dimensional plot (Fig. 3 a, b). For instance if the game reserve management allocates 0.4 of their land to hunting, then they would need to attract at least 30 hunters for them to realise positive returns from wildlife investment (Fig. 3 a). The contour plot and the three dimension plot show that for a given level of wildlife investment return, there is a positive relationship between land allocated to hunting and number of hunters.

Figure 3 Scenario where hunting and tourism are separate but wildlife is shared. (a) Contour plot: levels of the economic return on wildlife obtained from different combinations of the proportion of land under hunting and the number of hunters; (b) three-dimensional plot of the levels of the economic return on wildlife obtained from different combinations of the proportion of land under hunting and the number of hunters.
Scenario 2: Hunting and tourism on the same area
We now examine the relationship between the economic return on wildlife and the number of hunters (Fig. 4 a), where at first we assume that S, the unit damage cost caused by hunting on tourism is 0. There is a positive relationship between the economic return on wildlife and number of hunters (Fig. 4 a). To realize a positive economic return from wildlife investment, the number of hunters in the model should be at least 14, as compared to scenario 1 where at least 22 hunters were required when the land allocated to hunting was 0.1, and at least 40 hunters were required when land allocated to hunting was 0.8 (Fig. 2 c). The economic return on wildlife increases as more hunters are attracted by the game reserve. This shows that undertaking both tourism and hunting can result in high returns to wildlife investment for the game reserve management, if the management attracts sufficient numbers of hunters.

Figure 4 (a) Relationship between the economic return on wildlife and the number of hunters for scenario 2 assuming S is 0. (b) Scenario 2, comparing the relationship between the economic return on wildlife and the number of hunters when the unit damage cost caused by hunting on tourism (S) is 0 and when it is 12. (c) Comparing the relationship between the economic return on wildlife and the number of hunters for scenarios 1 and 2.
We have assumed in the model that when hunting and tourism are performed in the same area, hunting has some negative effects on tourism. We represented this effect through the parameter, S, which denotes a unit damage cost on tourism due to hunting. If we compare the cases when S is zero and when S is 12, the economic return on wildlife for the case when S is 0 overlies that when S is 12 (Fig. 4 b, compare with Fig. 1 a). This means that when the unit damage cost on tourism due to hunting increases, it results in an increase in the wildlife economic return to investment. This might seem as a strange result. This could be explained by looking at the findings from theory (see Figs 1 a and b, and Eq. 10). A positive S causes an increase in the economic return on wildlife (Fig. 1 a), partly due to an increase in the wildlife population (Eq. 10). The last term in Eq. (10), which we termed the ‘growth rate:population ratio’, can only enhance wildlife viability if it is kept as small as possible. This is achieved by maintaining the denominator X, which represents wildlife stock, at as large a value as possible. So S results in an increase in the wildlife population, which then enhances the economic return from wildlife through both hunting and tourism (in this case the wildlife effect will have more influence than the negative hunting effect on tourism). This partly explains why we have deduced that there is a larger wildlife population in scenario 2 than in scenario 1 (Fig. 1 b). A point to note here is that the effect of S occurs only in scenario 2, and is absent in other scenarios. Overall this means that undertaking hunting and tourism in the same area is beneficial to the game reserve provided the effect of these two uses is not complex and is marginal. Another point to note is that there is a limit to the number of hunters that can come to the game reserve; in this case the number cannot exceed 44 (Fig. 4 b). If there are more than 44 hunters, then there is a steep drop in the economic return on wildlife.
We next compare the economic return on wildlife for scenarios 1 and 2. The economic return on wildlife curve for scenario 2 is higher than that of scenario 1. For scenario 2, the economic return on wildlife become positive when there are more than six hunters, but, for scenario 1, it becomes positive when there are 40 or more hunters (Fig. 4 c). Scenario 2 always dominates scenario 1. This shows that undertaking both consumptive and non-consumptive uses enhances the overall return from wildlife investment for the management of a large game reserve. Using these findings, we can already deduce that scenario 2 is also superior to scenario 3 (where hunting and tourism are completely separated) in terms of returns to wildlife investment (Fig. 1 b). In the same way, we deduce that scenario 1 has a higher level of economic return on wildlife than scenario 3 (Fig. 1 b).
How to reach the steady state
Equation (10) is the result of a derivation for the economic return on wildlife for steady state control in the case of maximum sustainable yield based on Pontryagin's minimum principle. How is this steady state, given an arbitrary initial state, reached? A natural approach would be to use bang-bang control on the hunting control variable E to swiftly move to an optimal steady state, but since the Hamiltonian is not linear in all control variables, it is not guaranteed the steady state corresponding to a maximum sustainable yield steady state can be reached easily. However, setting the easiest-to-control variable, namely E, to zero allows the population to increase to carrying capacity. Whilst recovering, hunters may be allowed on the land as soon as the population reaches the maximum sustainable yield (MSY) steady state value. The X* with MSY will then, by construction of the optimal solution, maintain this steady state.
Summary of model scenarios
Under the assumption that there is more wildlife biomass per unit area if there is only non-consumptive tourism, rather than when there is only hunting (Munro Reference Munro1990; Schulz & Skonhoft Reference Schulz and Skonhoft1996), combining tourism and hunting creates a potential loss of wildlife biomass per unit area (Table 2). As we move from scenarios where hunting and tourism areas are completely separated (scenario 3), through where they are partially separated (scenario 1), to where they are co-existing (scenario 2), the wildlife biomass per hectare decreases (this is what we term the biomass effect) but the land area increases (the land effect), and the wildlife offtake increases (the wildlife offtake effect represented here indirectly by number of hunters) (Table 2; Fig. 2 a). So, if management was interested in maintaining wildlife biomass, they would completely separate hunting and tourism sections of the game reserve, provided hunting did not completely dominate tourism. If management was interested in the maximization of profit, then scenario 2 provides the optimal scenario, combining hunting and tourism areas, provided tourism does not completely dominate hunting and the land effect overpowers the biomass effect. If the biomass effect overpowers the land effect, then the landowners would be better off choosing scenario 1. Further, if tourism dominates, then scenario 1 would be the best option when wildlife biomass is highest. If management were interested in satisfying their hunting clientele, they would choose scenario 2 provided the negative effect of tourism was not high.
Table 2 Comparisons of optimum biomass and profit for the three scenarios: from a wildlife point, scenario 1, where hunting does not dominate, appears to be optimal, and also appears to be the most profitable scenario. Wildlife offtake is represented indirectly in the models by the number of hunters.

DISCUSSION
We examined from a theoretical point of view, and also with use of a numerical example, the conditions imposed on a landowner when allocating land between consumptive and/or non-consumptive land uses, which are potentially conflicting. Our work uses a framework similar to that of Schulz and Skonhoft (Reference Schulz and Skonhoft1996), as both models assume that the amount of land is fixed. However, Schulz and Skonhoft (Reference Schulz and Skonhoft1996) analysed two competing land uses, wildlife and livestock production, where wildlife in the parkland generated externalities outside the park where livestock was produced. In our case, we analysed two activities belonging to the wildlife land-use category that were potentially conflicting. We also substantiated some of our theoretical analytical findings using a simple numerical analysis.
In South Africa, there are numerous game reserves and ranches where hunting and tourism occurs together or, in other words, wildlife is shared between consumptive and non-consumptives uses, as evidenced by the removal of fences separating private properties and even the removal of fences that are separating private and public reserves. (Lindberg et al. Reference Lindberg, James, Goodman, Aylward and Lutz2003; APNR 2005; Patterson & Khosa Reference Patterson and Khosa2005). However, some lobbyists and animal rights group advocate that hunting and tourism should not co-exist (Hearne & McKenzie Reference Hearne, McKenzie, Prins, Grootenhuis and Dolan2000; ABSA 2003), which would be analogous to attaching an extremely high value on the negative effect of hunting on tourism. If their assessment is correct, then the best strategy would be a complete separation of the two land-use activities, using fences to separate hunting from the tourism areas. Although this is not part of our model, another way to look at this would be to hold the land-use activities at different times; hunting during the hunting season, whilst engaging in tourism the remainder of the time (Hearne & McKenzie Reference Hearne, McKenzie, Prins, Grootenhuis and Dolan2000; ABSA 2003). This would ensure that tourism and hunting benefited from each other through a larger optimal wildlife stock than if only hunting was practised. In this case, then the model would need to accommodate a time variable, since tourism and hunting land uses would be sharing wildlife and area, but time would be separating the two activities.
Our model allows some insight into wildlife management decisions by a game reserve management or a landowner with a very large property, who may need to consider how to combine wildlife consumptive and non-consumptive uses. Our framework might be useful for analysing wildlife management cases where land size is fixed and land-use activities are both consumptive and non-consumptive. Areas that could be further explored include models for more than one species, or inclusion of spatial heterogeneity in the model, as species are generally not homogenously distributed over the landscape with regard to sex or age, influencing the economic returns per unit area. However, we believe that our framework could assist in analysing a significant number of real world wildlife management cases. The key aspect to emerge from our work is that consumptive and non-consumptive uses are not mutually exclusive, but that careful planning is needed to ensure that multiple reserve objectives can be met. Further, our results indicate that the two uses can be undertaken in the same contiguous area. Whether they are spatially (or temporally) separated depends on the magnitude of the consumptive use. Where the consumptive use is not dominant, the two could be compatible in the same shared area provided the wildlife population is sufficiently numerous. Transparency in the economic consequences of these two potential competing land uses can convince landowners to change their management policies (Paloniemi & Tikka Reference Paloniemi and Tikka2008; Knight et al. Reference Knight, Grantham, Smith, McGregor, Possingham and Cowling2011), and thereby enhance biodiversity conservation (Fisher & Dills Reference Fisher and Dills2012).
ACKNOWLEDGEMENTS
This study is part of an integrated programme, the Elephant Movements and Bio-economic Optimality Program (TEMBO), financially supported by the Netherlands Foundation for the Advancement of Tropical Research (WOTRO), the Dr Marie Luttig Trust, and Shell South Africa. We thank the Associated Private Nature Reserves (APNR) for providing assistance. We also thank anonymous reviewers for their valuable comments on earlier versions of the paper.
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0376892915000296.








