


Introduction
The end-Permian mass extinction was the most severe in geological history and saw the disappearance of the majority of marine organisms (e.g., Wignall, Reference Wignall2015). Ecosystem recovery took several million years and was hindered by the Smithian-Spathian extinction around two million years after the main crisis. Early Triassic sea surface temperatures were especially high and reached their zenith in the late Smithian (Romano et al., Reference Romano, Goudemand, Vennemann, Ware, Hermann, Hochuli, Brühwiler, Brinkmann and Bucher2012; Sun et al., Reference Sun, Joachimski, Wignall, Yan, Chen, Jiang, Wang and Lai2012). Black shales and other anoxic facies were also widespread, especially in the late Smithian, although oxic marine red beds (MRBs) became common in the early Spathian (Sun et al., Reference Sun, Wignall, Joachimski, Bond, Grasby, Sun, Yan, Wang, Chen and Lai2015; Song et al., Reference Song, Jiang, Poulton, Wignall, Tong, Song, An, Chu, Tian, She and Wang2017). Thus, high seawater temperatures and anoxia in the late Smithian may have caused the second order extinction event at this time, as seen among conodonts (Orchard, Reference Orchard2007; Stanley, Reference Stanley2009), ammonoids (Hallam and Wignall, Reference Hallam and Wignall1997; Brayard et al., Reference Brayard, Escarguel, Bucher and Brühwiler2009; Stanley, Reference Stanley2009), and bivalves (Chen, Reference Chen, Rong and Fang2004). The small size of organisms (the Lilliput Effect), such as conodonts (Chen et al., Reference Chen, Twitchett, Jiang, Richoz, Lai, Yan, Sun, Liu and Wang2013; Maekawa and Komatsu, Reference Maekawa, Komatsu, Shigeta, Komatsu, Maekawa and Tran2014) and gastropods (Piestch et al., Reference Piestch, Mata and Bottjer2014), may also be attributable to the high temperatures.
The term ‘gondolellid’ derives from the family Gondolellidae Lindström and is composed of two groups—segminiplanate, platform-bearing genera (e.g., Neogondolella Bender and Stoppel, Scythogondolella) and segminate, platform-less genera (e.g., Neospathodus Mosher, Novispathodus Orchard). The fate of platform-bearing gondolellid conodonts during the vicissitudes of the Early Triassic is unclear. They were globally prosperous prior to the end-Permian mass extinction and once again successful in the Middle Triassic. However, in the Early Triassic, conodonts in low-latitude regions, such as South China, were mostly dominated by blade-shaped (or segminate) instead of platform-bearing gondolellids. By comparison, in northern high-latitude areas and the southern margin of Neo-Tethys, the platform-bearing gondolellids became more common (e.g., Orchard and Zonneveld, Reference Orchard and Zonneveld2009; Konstantinov et al., Reference Konstantinov, Sobolev and Yadrenkin2013; Bondarenko et al., Reference Bondarenko, Zakharov, Guravskaya and Safronov2015). Attempts to elucidate conodont biogeographic realms in the Early Triassic (e.g., Yang et al., Reference Yang, Hao and Jiang2001; Klets, Reference Klets2008) are currently hampered by incomplete data from some regions. In this paper, we report newly discovered and abundant Smithian platform-bearing gondolellid conodonts from the Yiwagou Section of Tewo County, Gansu Province, northwestern China that enable us to advance discussions of the evolution of this important Triassic group. In addition, a taxonomic description is provided for some of the less well-known conodonts (Parafurnishius xuanhanensis Yang et al., Reference Yang, Yuan, Henderson and Shen2014 and several Scythogondolella species).
Geological setting and stratigraphy
The Early Triassic Qinling Basin was a major seaway between the South China and Northern China blocks that narrowed to the east and opened to Paleo-Tethys in the west (Lai et al., Reference Lai, Yin and Yang1992, Reference Lai, Yin and Yang1995; Feng et al., Reference Feng, Du, Zhang and Zeng1994; Yin and Peng, Reference Yin and Peng1995). The studied Yiwagou Section was located in a shallow-water, carbonate platform at the northern margin of South China Block and on the southern side of the Basin (Fig. 1). There has been little research in the region because of its inaccessibility and high altitude (but see Yin et al., Reference Yin, Yang and Lai1988, Reference Yin, Yang, Huang, Yang and Lai1992; Lai et al., Reference Lai, Yin and Yang1992, Reference Lai, Yang and Yin1994, Reference Lai, Yin and Yang1995; Yin and Peng, Reference Yin and Peng1995). The Yiwagou Section lies along a ravine near Zhagana village, Tewo County (start point GPS 34.256N, 103.204E, Fig. 1). The section is ~1500 m long, with the highest point reaching 4060 m above sea level, and consists of continuous strata from upper Permian to Lower Triassic composed of the Changhsing Formation (P3ch), the Zhalishan Formation (T1z), and the Maresongduo Formation (T1m). The Changhsing Formation is mostly gray, thick-bedded limestone and oolitic limestone. The Zhalishan Formation (Griesbachian to Smithian) is 565.5m thick, consisting of gray to dark gray, thin- to medium-bedded micritic and bioclastic limestone with intercalations of calcareous, reddish to purple-red, fine-grained sandy limestone (Fig. 2). The Maresongduo Formation (Spathian) consists of red to purple-red, thick-bedded crystallized dolomite, dolomitic limestone, and micritic limestone. Overall, red beds first appear within the lower Zhalishan Formation and roughly increase in importance upwards. Marine Red Beds (MRBs) are more typically found in deeper offshore environment (Song et al., Reference Song, Jiang, Poulton, Wignall, Tong, Song, An, Chu, Tian, She and Wang2017), but are rare in shallow-water carbonate platform settings such as the Yiwagou Section. The MRB horizons likely record fluctuating oxygen concentration and water depth in the southern Qinling Basin (Lai et al., Reference Lai, Yin and Yang1992).

Figure 1. Geographic position of studied area. (1) Tectonic map of Qinling orogenic belt (modified after Dong et al., Reference Dong, Zhang, Liu, Li, Chen, Zhang, Zhang, Yang, Sun and Zhang2015). Scale bar = 1000 km. (2) Location of the Yiwagou Section, Tewo. Scale bar = 20 km. (3) Paleogeography of Qinling Basin (modified after Lai et al., Reference Lai, Yin and Yang1995). 1. National highway; 2. main road; 3. center of an aulacogen; 4. continental deposits; 5. basinal deposits; 6. slope deposits; 7. old land; 8. offshore deposits.

Figure 2. Photographs of marine red beds in Zhalishan Formation from Dienerian to Smithian, Yiwagou Section; 13–17 = bed numbers. Scale bar = 10 m.
Conodont assemblages in this area are typically high diversity but low abundance (Lai, Reference Lai, Yin, Yang, Huang, Yang and Lai1992). Six Early Triassic conodont zones have been established at the Yiwagou Section; in ascending order, they are: Hindeodus parvus Zone, Neospathodus dieneri Zone, Neospathodus pakistanensis Zone, Pachycladina-Parachirognathus Assemblage Zone, Neospathodus triangularis Zone, and the Neospathodus homeri-Neospathodus hungaricus Assemblage Zone (Lai, Reference Lai, Yin, Yang, Huang, Yang and Lai1992, fig. 3). Biogeographically, the assemblages at Yiwagou have been thought to have characteristics transitional between South China and North China, albeit with a greater affinity to South China faunas (Lai et al., Reference Lai, Yin and Yang1992, 1995). In addition to conodonts, the section also yields many foraminifers and bivalves, such as the foraminifers Colaniella media Miklukho-Maklay, Reference Miklukho-Maklay1954 and Reichelina tenuissima Miklukho-Maklay, Reference Miklukho-Maklay1954 in the Changhsing Formation, the bivalves Claraia concentrica (Yabe, Reference Yabe1928), C. hubeiensis Chen in Gu et al., Reference Gu, Huang, Chen, Wen, Ma, Lan, Xu, Liu, Wang, Wang, Qiu, Huang, Zhang, Chen and Wu1976, C. aurita (Hauer, Reference Hauer1850), Eumorphotis sp., Entolium sp. and Pteria sp. in the Zhalishan Formation and the bivalve Chlamys weiyuanensis (Hsü, Reference Hsü1939) in the Maresongduo Formation (Yang et al., Reference Yang, Yang, Wu, Yin, Yang, Huang, Yang and Lai1992).
Materials and methods
A total of 153 samples (each one weighting ~3–4 kg) were collected from the upper Changhsing, Zhalishan and lower Maresongduo formations at Yiwagou. All samples were crushed into 1–2 cm3 fragments, then dissolved in an 8% solution of acetic acid, followed by heavy liquid separation. A stereoscopic binocular microscope was used to find conodonts in the residues, and a scanning electron microscope (SEM) was used for photography.
Repository and institutional abbreviation
All conodonts in this study are stored in the School of Earth Science, China University of Geosciences (CUG) in Wuhan City, Hubei Province, China.
Systematic paleontology
Class Conodonta Eichenberg, Reference Eichenberg1930
Order Conodontophorida Eichenberg, Reference Eichenberg1930
Family Ellisoniidae Clark, Reference Clark1972
Genus Parafurnishius Yang et al., Reference Yang, Yuan, Henderson and Shen2014
Type species
Parafurnishius xuanhanensis Yang et al., Reference Yang, Yuan, Henderson and Shen2014 from the Feixianguan Formation at the Panlongdong Section in Xuanhan County, northeastern Sichuan Province, southwest China.
Parafurnishius xuanhanensis Yang et al., Reference Yang, Yuan, Henderson and Shen2014
Figure 4.1–4.21
- 2014
Parafurnishius xuanhanensis Yang et al., p. 269, pl. 3, figs. A–J, pl. 7, figs. A–O.
Holotype
NIGP161300, from the Feixianguan Formation at the Panlongdong Section in Xuanhan County, northeastern Sichuan Province, southwest China (Yang et al., Reference Yang, Yuan, Henderson and Shen2014, pl. 3, fig. J).
Occurrence
Zhalishan Formation at the Yiwagou Section in Tewo County, Gansu Province, northwestern China, in Dienerian or Smithian.
Description
The P1 element has a variable shape platform with the width-length ratio about 1:2. In upper view, a relatively small cusp lies in the center, surrounded by 8–10 high and strong irregularly distributed denticles. Generally, it has an anterior process with 1–2 denticles and a roughly triangular posterior process with 3–4 denticles, between which there is a broad platform with 2–7 denticles and a cusp. In lateral view, the basal margin is almost straight, but sometimes upturned posteriorly in juvenile elements. Denticles generally erect or inclined posteriorly in the posterior part. In lower view, a large basal cavity is approximately diamond-shaped with a basal pit located in the center. A basal furrow extends from the basal pit to the anterior end, but always no furrow or a very shallow one to the posterior end.
Materials
71 specimens.
Remarks
The specimens at Yiwagou Section are much smaller than those recorded from the Panlongdong Section, Sichuan Province, southwest China (Yang et al., Reference Yang, Yuan, Henderson and Shen2014). The juvenile individual has only one row of denticles in the anterior part that separates into two rows from middle to posterior part, which indicates that this species may have developed from blade-shaped conodonts.
Another conodont, Platyvillosus corniger Kolar-Jurkovšek and Chen in Chen et al., Reference Chen, Jurkovšek, Jurkovšek, Aljinović and Richoz2016, is very similar to this species, but is of Olenekian age. It is found in Spathian strata at Žiri-sortirnica 28 Section, in the Idrija–Žiri area of Slovenia (Chen et al., Reference Chen, Jurkovšek, Jurkovšek, Aljinović and Richoz2016) and Olenekian strata in Mokrice locality, eastern Slovenia (Kolar-Jurkovšek and Jurkovšek, Reference Kolar-Jurkovšek and Jurkovšek2015; Kolar-Jurkovšek et al., Reference Kolar-Jurkovšek, Chen, Jurkovšek, Poljak, Aljinović and Richoz2017). Although Kolar-Jurkovšek et al. (Reference Kolar-Jurkovšek, Chen, Jurkovšek, Poljak, Aljinović and Richoz2017) have highlighted the subtle morphological differences (Chen et al., Reference Chen, Jurkovšek, Jurkovšek, Aljinović and Richoz2016; Kolar-Jurkovšek et al., Reference Kolar-Jurkovšek, Chen, Jurkovšek, Poljak, Aljinović and Richoz2017), it is still difficult to distinguish these two species with specimens from the Yiwagou Section although they probably have different apparatuses (Yang et al., Reference Yang, Yuan, Henderson and Shen2014; Kolar-Jurkovšek et al., Reference Kolar-Jurkovšek, Chen, Jurkovšek, Poljak, Aljinović and Richoz2017). Here we assign them to Pa. xuanhanensis because they appear in a range of samples and are accompanied by Hindeodus postparvus Kozur, Reference Kozur1989b in the first few samples in Induan (not shown in Fig. 3), but this taxon needs further research.
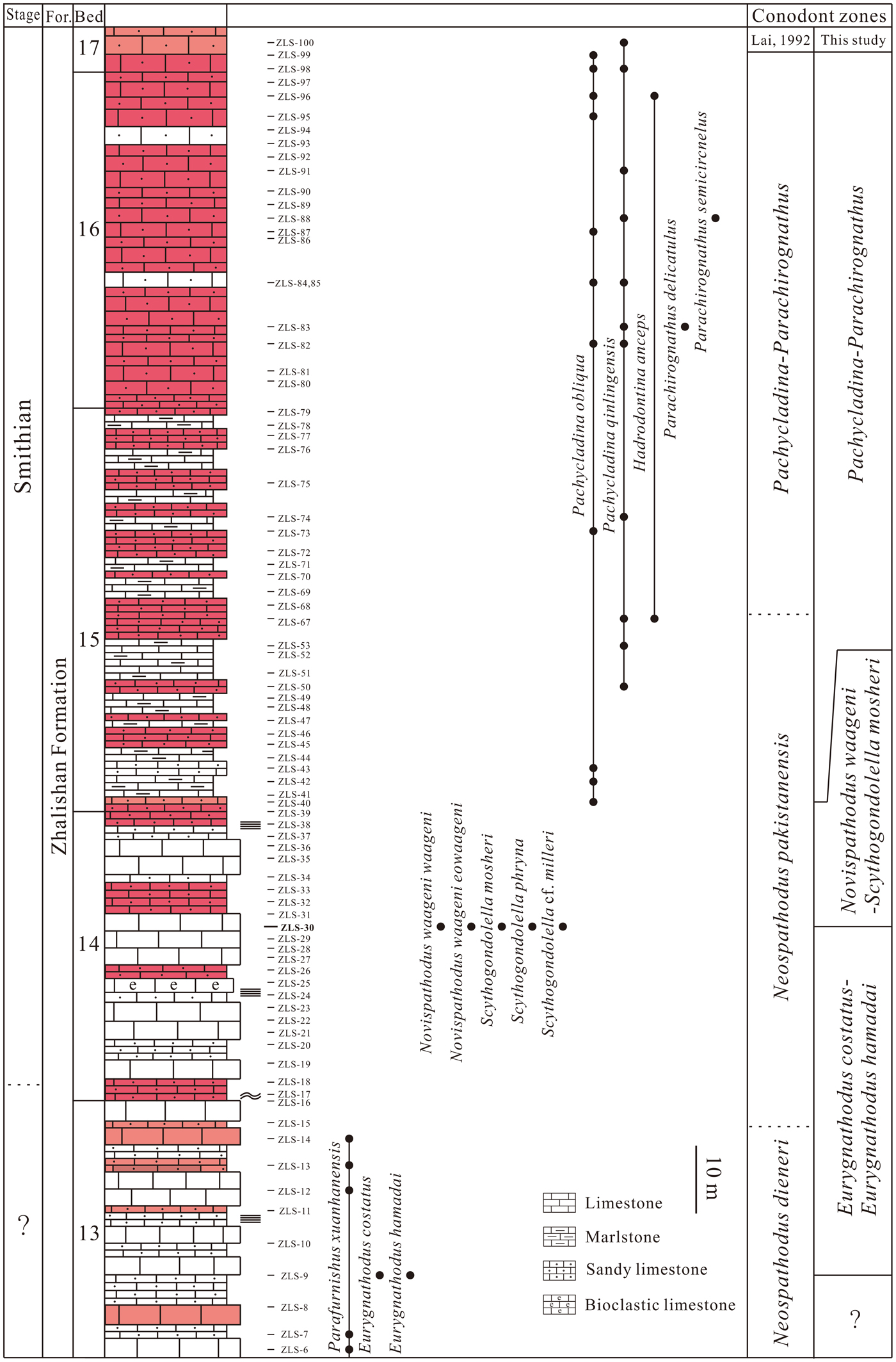
Figure 3. Conodont distribution in the Lower Triassic strata at the Yiwagou Section, Tewo, northwestern China. Scale bar = 10 m.
Order Ozarkodinida Dzik, Reference Dzik1976
Family Gondolellidae Lindström, Reference Lindström1970
Subfamily Scythogondolellinae Orchard, Reference Orchard and Krystyn2007
Genus Scythogondolella Kozur, Reference Kozur1989a
Type species
Gondolella milleri Müller, Reference Müller1956 from the Lower Triassic Meekoceras bed, Dinner Springs Canyon, northeastern Nevada, USA.
Remarks
Compared with Neogondolella, Scythogondolella is characterized by a very prominent blade-carina, which extends beyond the posterior end to form a free blade in most cases.
Scythogondolella mosheri (Kozur and Mostler, Reference Kozur and Mostler1976)
Figure 5.1–5.21
- 1973
Neogondolella nevadensis Clark; Mosher, p. 169, pl. 19, figs. 17, 18, 24.
- 1973
Neogondolella elongata Sweet; Mosher, p. 166, pl. 19, fig. 19.
- 1976
Gondolella mosheri Kozur and Mostler, p. 8, pl. 1, figs. 9–12.
- 1978
Neogondolella planata Clark; Weitschat and Lehmann, pl. 14, figs. 1–5.
- 1978
Neogondolella nevadensis; Weitschat and Lehmann, pl. 14, figs. 6–10.
- 198
Neogondolella elongatus Sweet; Tian, pl. 1, fig. 11.
- 1983
Neogondolella elongatus; Tian et al., pl. 94. fig. 1.
- 1984
Neogondolella nevadensis; Berry et al., pl. 1, figs. 22–25.
- 1984
Neogondolella nevadensis; Hatleberg and Clark, pl. 2, figs. 14, 15.
- 1991
Neogondolella nevadensis; Zhao and Zhang, pl. 1, figs. 17, 18.
- 2005
Scythogondolella mosheri; Orchard, p. 97, fig. 23A–H.
- 2008
Scythogondolella mosheri; Orchard, p. 410, pl. 5, figs. 1–4.
- 2008
Scythogondolella mosheri; Nakrem et al., pl. 5, figs. 4–6, 12, 13.
- 2014
Scythogondolella mosheri; Golding et al., p. 173, pl. 1, figs. 7–9.
- 2015
Scythogondolella mosheri; Golding et al., p. 167, pl. 12, figs. 19–21.
- 2018
Scythogondolella mosheri; Henderson et al., pl. 1. figs. 35–37.
Holotype
Dolpo, Nepal (Kozur and Mostler, Reference Kozur and Mostler1976, pl. 1, figs. 9–12).
Occurrence
Zhalishan Formation at the Yiwagou Section in Tewo County, Gansu Province, northwestern China, in Smithian.
Description
This species has a wedge-shaped platform, with the maximum width occurring posteriorly. The posterior margin is typically constricted and indented inwards from the large cusp, which projects posteriorly beyond the platform margin. Consequently, some specimens have a heart-shaped plan view. The platform with smooth upturned margins tapers progressively to the anterior end, leaving a free blade of variable length. The blade-carina is typically high with relatively large, discrete denticles, of uniform height in lateral view. In lower view, the rounded expanded basal cavity loop surrounds a small basal pit. Juvenile specimens typically have a more developed carina than platform, with a very prominent cusp projecting posteriorly beyond the platform margin.
Materials
30 specimens.
Remarks
Mosher (Reference Mosher1973) recovered Neogondolella nevadensis (Clark, Reference Clark1959) from the Romunduri and Tardus zones (ammonoid zones of lower and upper Smithian) and Ng. elongata (Sweet, Reference Sweet1970a) from the Tardus Zone in the Canadian Arctic and British Columbia. However, these occurrences are reassigned to Sc. mosheri based on more recent study. Kozur and Mostler (Reference Kozur and Mostler1976) defined Sc. mosheri (= Gondolella mosheri) from the Tardus Zone (upper Jakutian) of Dolpa, Nepal. Some of the elements illustrated as Neogondolella planata (Clark, Reference Clark1959) and Ng. nevadensis (Weitschat and Lehmann, Reference Weitschat and Lehmann1978, pl. 14, figs. 1–10) clearly belong to Sc. mosheri. In addition, Ng. elongatus in Tian (Reference Tian1982, pl. 1, fig. 11) and Tian et al. (Reference Tian1983, pl. 94. fig. 1), Ng. nevadensis in Berry et al. (Reference Berry, Burrett and Banks1984, pl. 1, figs. 23–25), Hatleberg and Clark (Reference Dagis1984, pl. 2, figs. 14, 15), and Zhao and Zhang (Reference Zhao and Zhang1991, pl. 1, figs. 17, 18) can also be considered reassigned to Sc. mosheri.
Scythogondolella mosheri has often been confused with Ng. nevadensis, Ng. planata, and Ng. elongata, but it can be distinguished by its relatively high-bladed carina with large denticles and strong cusp. Both Ng. nevadensis and Ng. planata have lower nodular denticles and a smaller cusp. The name of Ng. elongata (or elongatus) is no longer used, and has been changed to Columbitella elongata (Orchard, Reference Orchard2005), which also has a strong cusp and carina, but a more rounded and expanded loop surrounding its basal pit (Orchard, Reference Orchard2007). Compared with specimens in the Canadian Arctic, Sc. mosheri from Yiwagou are much smaller, with no more than nine denticles, and so are probably juveniles.
Scythogondolella cf. milleri (Müller, Reference Müller1956)
Figure 5.22–5.23
Holotype
Lower Triassic Meekoceras bed, Dinner Springs Canyon, northeastern Nevada, USA (Müller, Reference Müller1956, p. 823, pl. 95, figs. 4–6).
Occurrence
Zhalishan Formation at the Yiwagou Section in Tewo County, Gansu Province, northwestern China, in Smithian.
Description
This large broken specimen has subparallel upturned platform margins ornamented with small nodes. The carina is relatively low with fused denticles. The moderate-sized cusp projects posteriorly beyond the platform margin. In the lower view, the keel with basal groove is broad.
Material
One specimen.
Remarks
This specimen closely resembles Sc. milleri, but it is broken and so we assign it to Sc. cf. milleri. Scythogondolella milleri, defined by Müller (Reference Müller1956) in the Meekoceras Bed of Nevada, is the most widespread and extensively studied species of its genus. Typically, it has uneven upturned platform margins ornamented with nodes or small denticles, which is thought to have evolved from older Sc. mosheri with slightly waved platform margins. Nogami (Reference Nogami1968) identified numerous Sc. milleri ranging from juvenile to gerontic stages in Timor and Japan. The large elements have distinctive, denticulated platform margins, but the juvenile elements closely resembles Sc. mosheri with slightly sculptured platform margins. It seems that juvenile elements of Sc. milleri and Sc. mosheri have very similar early growth stages, but become differentiated in later stages. We also note that one Gondolella milleri illustrated by Nogami (Reference Nogami1968, pl. 10, fig. 11) is Neogondolella nevadensis rather than Sc. milleri because of its low carina. Additionally the specimen illustrated by Liang et al. (Reference Liang, Tong and Zhao2011, pl. 3, fig. 10) from Chaohu, South China should be assigned to Sc. milleri, making it the first report of Smithian platform-bearing gondolellid conodonts in South China.
Scythogondolella milleri is a cosmopolitan species; it has been found in Siberia (Dagis, Reference Dagis1984; Konstantinov et al., Reference Konstantinov, Sobolev and Yadrenkin2013), Nevada (Müller, Reference Müller1956; Clark and Mosher, Reference Clark and Mosher1966), Utah (Clark et al., Reference Clark, Paul, Solien, Morgan, Sandberg and Clark1979), Canadian Arctic (Orchard, Reference Orchard2008), British Columbia (Mosher, Reference Mosher1973; Solien, Reference Solien1979; Orchard and Tozer, Reference Orchard, Tozer, Moslow and Wittenberg1997), Dolpo, Nepal (Kozur and Mostler, Reference Kozur and Mostler1976), Spiti, India (Orchard, Reference Orchard2007), Timor Island (Berry et al., Reference Berry, Burrett and Banks1984), Japan (Nogami, Reference Nogami1968), Spitzbergen (Nakrem et al., Reference Nakrem, Orchard, Weitschat, Hounslow, Beatty and Mørk2008), and Southern Primorye (Bondarenko et al., Reference Bondarenko, Zakharov, Guravskaya and Safronov2015). In China, it has been reported from Tibet (Tian, Reference Tian1982; Tian et al., Reference Tian, Dai and Tian1983; Wang and Wang, Reference Wang and Wang1995; Zou et al., Reference Zou, Mao, Chen and Rao2006), Qinghai Province (Fang et al., Reference Fang, Zhang, Wang and Wang2013), and, as noted above, Chaohu, South China (Liang et al., Reference Liang, Tong and Zhao2011).
Scythogondolella phryna Orchard and Zonneveld, Reference Orchard and Zonneveld2009
Figure 5.24–5.26
- 2009
Scythogondolella phryna Orchard and Zonneveld, p. 786, pl. 16, figs. 10–16, 20–24.
- 2010
Scythogondolella phryna; Beranek et al., pl. 6, figs. 37–39.
- 2014
Scythogondolella phryna; Golding et al., p. 173, pl. 1, figs. 4–5.
Holotype
GSC 132549 from GSC loc. no. C-103866 (213E), in the Toad Formation on Toad River, northeast British Columbia (Orchard and Zonneveld, Reference Orchard and Zonneveld2009, pl. 16, figs. 16, 23, 24).
Occurrence
Zhalishan Formation at the Yiwagou Section in Tewo County, Gansu Province, northwestern China, in Smithian.
Description
This specimen has a narrow biconvex platform, tapering anteriorly and posteriorly uniformly, with a short free blade occurring in the anterior end. The platform is roughly symmetrical and flat, broadest around the middle. The large cusp and high blade-carina with 10 large discrete denticles that all lean posteriorly. Although they are all broken, the height still apparently surpasses the platform width.
Material
One specimen.
Remarks
This species was originally described from the Wapiti Lake and Toad River areas in northeast British Columbia (Orchard and Zonneveld, Reference Orchard and Zonneveld2009). Scythogondolella phryna seems like an intermediate form between segminiplanate and segminate conodonts with an atrophic platform. It may be a cosmopolitan species, but needs more research.
Results
A total of 2100 conodont specimens (221 P1 elements) were obtained from the Yiwagou samples so far, and this study focuses on those obtained from the upper Zhalishan Formation (Dienerian and Smithian, Figs. 4–7). Most samples were productive, including sample no. ZLS-30 from the Zhalishan Formation, which yielded abundant Scythogondolella (38 P1 elements in total). This conodont fauna has rarely been found in the Qinling Basin or in South China before. Sample no. ZLS-30 included 30 Scythogondolella mosheri, one Sc. Phryna, and one Sc. cf. milleri. The remaining specimens are difficult to identify because of poor preservation. Only one segminiplanate conodont with a deformed platform, assigned to Neogondolella cf. milleri, was found in Zhalishan Formation before this study (Lai, Reference Lai, Yin, Yang, Huang, Yang and Lai1992). Other than Scythogondolella, 13 Novispathodus and some ramiform elements were also recovered from this sample, including six Novispathodus waageni waageni (Sweet, Reference Sweet1970a) and three Novispathodus waageni eowaageni (Zhao and Orchard in Zhao et al., Reference Zhao, Orchard, Tong, Sun, Zuo, Zhang and Yun2007). All specimens in this sample have a Conodont Alteration Index (CAI) around five (i.e., black in color).

Figure 4. SEM photos of conodonts obtained from the Yiwagou Section. (1–21) Parafurnishius xuanhanensis Yang et al., Reference Yang, Yuan, Henderson and Shen2014: (1–6) from sample ZLS-6, registration nos. ZLS17006001, 17006002; (7–12) from sample ZLS-7, registration nos. ZLS17007001, 17007002; (13–21) from sample ZLS-13, registration nos. ZLS17013001–17013003. (22–36) Eurygnathodus costatus Staesche, Reference Staesche1964, from sample ZLS-9, registration nos. ZLS17009001–17009005. (37–45) Eurygnathodus hamadai (Koike, Reference Koike1982), from sample ZLS-9, registration nos. ZLS17009006–17009008. Scale bar = 100 µm.
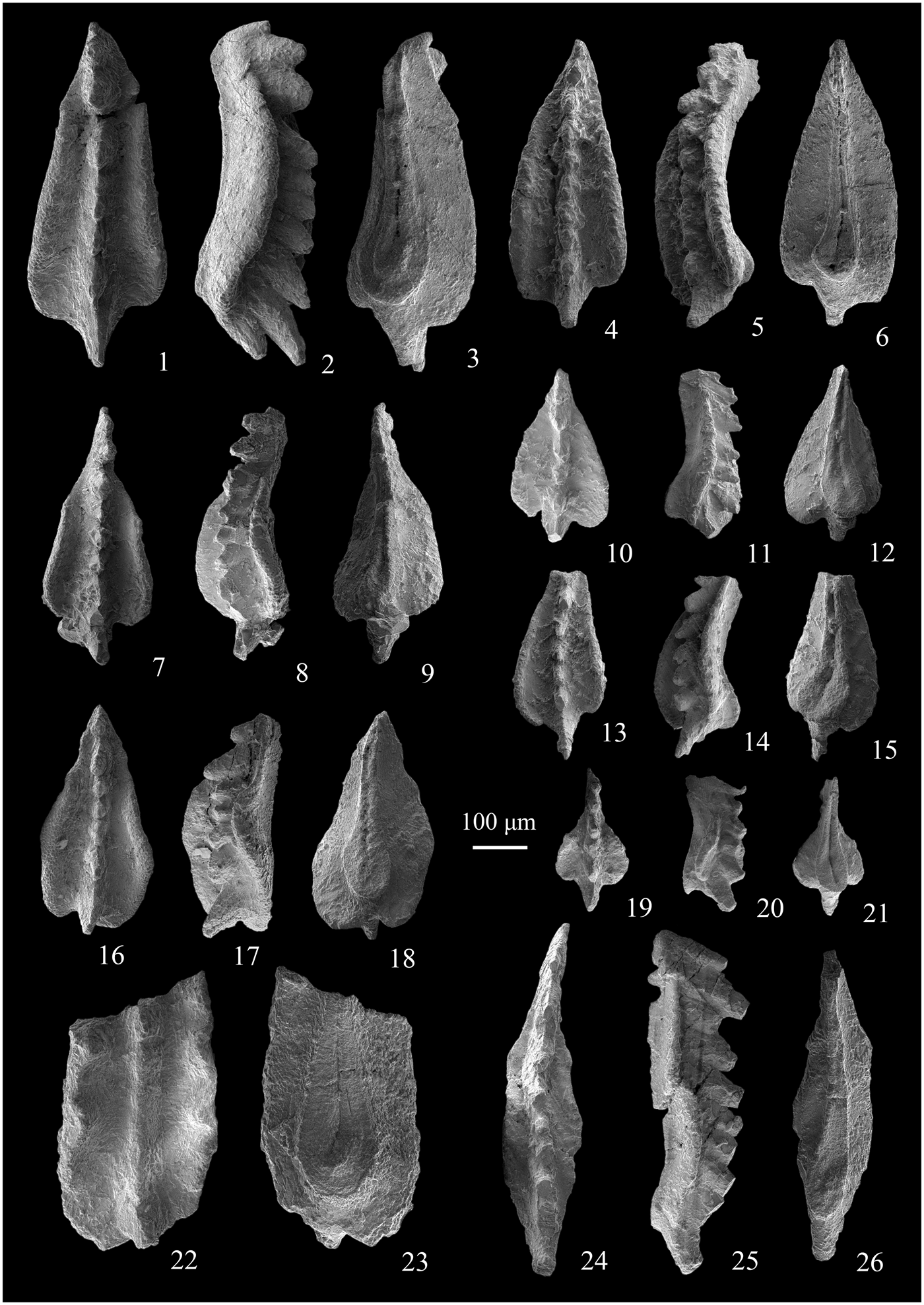
Figure 5. SEM photos of conodonts obtained from the Yiwagou Section. (1–21) Scythogondolella mosheri (Kozur and Mostler, Reference Kozur and Mostler1976), registration nos. ZLS17030001–17030007. (22, 23) Scythogondolella cf. milleri (Müller, Reference Müller1956), registration no. ZLS17030008. (24–26) Scythogondolella phryna Orchard and Zonneveld, Reference Orchard and Zonneveld2009, registration no. ZLS17030009. All come from sample ZLS-30. Scale bar = 100 µm.

Figure 6. SEM photos of conodonts obtained from the Yiwagou Section. (1–6) Novispathodus waageni waageni (Sweet, Reference Sweet1970a), from sample ZLS-30, registration nos. ZLS17030039–17030041. (7–10) Novispathodus waageni eowaageni (Zhao and Orchard in Zhao et al., Reference Zhao, Orchard, Tong, Sun, Zuo, Zhang and Yun2007), from sample ZLS-30, registration nos. ZLS17030042, 17030043. (11, 12) Parachirognathus n. sp. A, from sample ZLS-82, registration nos. ZLS17082001, 17082002. (13) Pachycladina obliqua Staesche, Reference Staesche1964, S2 element, from sample ZLS-42, registration no. ZLS17042001. (14–19) Pachycladina qinlingensis Lai, Reference Lai, Yin, Yang, Huang, Yang and Lai1992: (14) P2 element, from sample ZLS-88, registration no. ZLS17088001; (15) P2 element, from sample ZLS-85, registration no. ZLS17085001; (16) P2 element, from sample ZLS-74, registration no. ZLS17074001; (17) P2 element, from sample ZLS-53, registration no. ZLS17053001; (18) S1 element, from sample ZLS-98, registration no. ZLS17098001; (19) S2 element, from sample ZLS-67, registration no. ZLS17067001. (20–22) Parachirognathus semicircnelus Tian and Dai in Tian et al., Reference Tian, Dai and Tian1983: (20) S2 element, from sample ZLS-88, registration no. ZLS17088002; (21, 22) S1 elements, from sample ZLS-88, registration nos. ZLS17088003, 17088004. Scale bar = 100 µm.

Figure 7. SEM photos of conodonts obtained from the Yiwagou Section. (1–7) Pachycladina qinlingensis: (1) M element, from sample ZLS-83, registration no. ZLS17083001; (2) M element, from sample ZLS-91, registration no. ZLS17091001; (3) S3–4 elements, from sample ZLS-98, registration no. ZLS17098002; (4) S3 element, from sample ZLS-50, registration no. ZLS17050001; (5) S1 element, from sample ZLS-50, registration no. ZLS17050002; (6) S3 element, from sample ZLS-67, registration no. ZLS17067002; (7) S2 element, from sample ZLS-82, registration no. ZLS17082003. (8–10) Pachycladina obliqua: (8) M element, from sample ZLS-95, registration no. ZLS17095001; (9) S3–4 elements, from sample ZLS-96, registration no. ZLS17096001; (10) S1 element, from sample ZLS-98, registration no. ZLS17098003. (11) Parachirognathus delicatulus Wang and Cao, Reference Wang and Cao1981, S1 element, from sample ZLS-83, registration no. ZLS17083002. (12, 13) Hadrodontina anceps Staesche, Reference Staesche1964: (12) P1 element, from sample ZLS-96, registration no. ZLS17096002; (13) P1 element, from sample ZLS-67, registration no. ZLS17067003. Scale bar = 200 µm.
Conodont zones at Yiwagou Section
In addition to Scythogondolella mosheri, Sc. phryna, Sc. cf. milleri and Novispathodus waageni (Sweet, Reference Sweet1970a) mentioned above, Eurygnathodus costatus Staesche, Reference Staesche1964, E. hamadai (Koike, Reference Koike1982), and Parafurnishius xuanhanensis were also discovered for the first time from the Yiwagou Section (Fig. 3). Three conodont zones are established in the Dienerian to Smithian interval, in ascending order they are: Eurygnathodus costatus-E. hamadai Assemblage Zone, Novispathodus waageni-Scythogondolella mosheri, Assemblage Zone, and the Pachycladina-Parachirognathus Assemblage Zone.
Eurygnathodus costatus-Eurygnathodus hamadai Assemblage Zone
Lower limit: first occurrence of E. costatus and E. hamadai. Upper limit: first occurrence of Novispathodus waageni. Associated conodonts include Pa. xuanhanensis and some ramiform elements. At the Yiwagou Section, 31 Eurygnathodus costatus and 29 E. hamadai specimens were recovered from sample no. ZLS-9. Both E. costatus and E. hamadai are widely distributed globally, and often co-occur in the late Dienerian or Smithian (e.g., Chen et al., Reference Chen, Jurkovšek, Jurkovšek, Aljinović and Richoz2016). Parafurnishius xuanhanensis was found in Induan strata from the Panlongdong Section, Sichuan Province, southwest China (Yang et al., Reference Yang, Yuan, Henderson and Shen2014). This is the second discovery of this species, which indicates a connection between Yiwagou and northeastern Sichuan.
Novispathodus waageni-Scythogondolella mosheri Assemblage Zone
Lower limit: first occurrence of Nv. waageni and Sc. mosheri. Upper limit: first occurrence of Pc. obliqua Staesche, Reference Staesche1964. Associated conodonts include Sc. phryna, Sc. cf. milleri, and ramiform elements. Both Nv. w. waageni and Nv. w. eowaageni occur in sample no. ZLS-30. Novispathodus waageni is a cosmopolitan species found in diverse facies (Tong et al., Reference Tong, Zakharov, Orchard, Yin and Hansen2003), although it is not known from southern European sections (Chen et al., Reference Chen, Jurkovšek, Jurkovšek, Aljinović and Richoz2016), and its first appearance indicates the beginning of the Smithian. Goudemand et al. (Reference Goudemand, Orchard, Tafforeau, Urdy, Brühwiler, Brayard, Galfetti and Bucher2012) re-allocated this species from Neospathodus to Novispathodus based on its multi-element apparatus. Zhao et al. (Reference Zhao, Orchard, Tong, Sun, Zuo, Zhang and Yun2007) differentiated Nv. w. eowaageni from Nv. w. waageni by its more upright denticles and subdivided the original Nv. waageni Zone into two subzones, a lower Nv. w. eowaageni and an upper Nv. w. waageni subzone. The Nv. w. waageni and Nv. w. eowaageni from Yiwagou Section have a length to width ratio of ~3:1, and their basal cavities are not developed, a typical feature of small elements (Sweet, Reference Sweet1970a). Novispathodus w. eowaageni has been found in South China, Malaysia, Kashmir, Spiti, Canada, and Western Australia, and ranges from the Smithian to the early Spathian (e.g., Zhao et al., Reference Zhao, Chen, Chen and Cao2013; Chen et al., Reference Chen, Jiang, Lai, Yan, Richoz, Liu and Wang2015).
Scythogondolella is a cosmopolitan Smithian conodont that lived in relatively deep-water environments. Kozur (Reference Kozur1989a) named the genus, later Orchard (Reference Orchard2005, Reference Orchard2007, Reference Orchard2008) and Orchard and Zonneveld (Reference Orchard and Zonneveld2009) reconstructed its apparatus and added some new species. Several Smithian conodont zones were established by Orchard and Zonneveld (Reference Orchard and Zonneveld2009) in the Wapiti Lake area of western Canada, including the Sc. lachrymiformis Zone, and the Sc. mosheri Zone, which could be subdivided into the Sc. phryna and Sc. milleri subzones. Compared with segminate conodont zones (such as Novispathodus and Neospathodus zones), platform-bearing gondolellid zones could be more refined, and have great potential to improve the precision of correlation of Smithian strata worldwide. In conclusion, the Novispathodus waageni-Scythogondolella mosheri Assemblage Zone at Yiwagou is considered to be of Smithian age.
Pachycladina-Parachirognathus Assemblage Zone
Lower limit: First occurrence of Pc. obliqua. Upper limit: undefined. Hundreds of ramiform elements occur in this zone, and their sizes vary substantially from bed to bed (and will be subject to future study). This zone is characterized by the bloom of Pachycladina and Parachirognathus, including Pachycladina obliqua, Pc. qinlingensis Lai, Reference Lai, Yin and Yang1992, Pc. sp., Parachirognathus delicatulus Wang and Cao, Reference Wang and Cao1981, Pa. semicircnelus Tian and Dai in Tian et al., Reference Tian, Dai and Tian1983, and Pa. sp.; the conodont Hadrodontina anceps Staesche, Reference Staesche1964 also occurs. Pachycladina and Parachirognathus are widely distributed in South China (Jiang et al., Reference Jiang, Luo, Lu and Tian2000; Wang et al., Reference Wang, Wang, Li and Wie2005; Yan et al., Reference Yan, Wang, Jiang, Wignall, Sun, Chen and Lai2013; Chen et al., Reference Chen, Jiang, Lai, Yan, Richoz, Liu and Wang2015), Qinling areas (Lai, Reference Lai, Yin, Yang, Huang, Yang and Lai1992), Tibet (Xia and Zhang, Reference Xia, Zhang, Sha, Wang and Lu2005), and western USA (Clark et al., Reference Clark, Paul, Solien, Morgan, Sandberg and Clark1979; Solien, Reference Solien1979). Pachycladina and Hadrodontina are common in the Dinarides (Perri, Reference Perri1991; Kolar-Jurkovšek and Jurkovšek, Reference Kolar-Jurkovšek and Jurkovšek2015; Kolar-Jurkovšek et al., Reference Kolar-Jurkovšek, Chen, Jurkovšek, Poljak, Aljinović and Richoz2017), whereas Parachirognathus is not very frequent. This zone corresponds to the Pachycladina-Parachirognathus Assemblage Zone in the Beibei Area, Chongqing (Jiang, Reference Jiang1982) and the Bianyang Section, Guizhou (Yan et al., Reference Yan, Wang, Jiang, Wignall, Sun, Chen and Lai2013), South China. It is also equivalent to the Parachirognathus-Fumishius Assemblage Zone and upper Parachirognathus ethingtoni Zone in the Great Basin, western USA (Clark et al., Reference Clark, Paul, Solien, Morgan, Sandberg and Clark1979; Solien, Reference Solien1979), where they were all assigned to the Smithian Substage.
Parafurnishius xuanhanensis also occurs below the Eurygnathodus costatus-Eurygnathodus hamadai Assemblage Zone (Fig. 3), but its age could not be well constrained because of the few reports about this species. Our conodont study has not allowed us to discern the level of the Dienerian/Smithian boundary at Yiwagou Section, although it is probably below sample no. ZLS-30. Further study, including a C isotope analysis, may help locate this level.
Discussion
Global geographical distribution of Early Triassic platform-bearing gondolellid conodonts
Ten platform-bearing gondolellid conodont genera (Clarkina, Neoclarkina Henderson, Borinella Budurov and Sudar, Scythogondolella, Neogondolella, Gladigondolella Müller, Paullella Orchard, Columbitella Orchard, Magnigondolella Golding and Orchard, and Spathogondolella Jiang) occur in the Early Triassic. The evolution of platform-bearing gondolellids in the Early Triassic remains controversial. Klets and Kopylova (Reference Klets, Kopylova, Lucas and Spielmann2007) suggested an evolutionary lineage consisting of: Mesogondolella—>Clarkina—>Neospathodus—>Neogondolella—>Scythogondolella. However, we consider that Neogondolella is unlikely to have evolved from Neospathodus. The opinion we tentatively support is as follows: Clarkina survived the end-Permian mass extinction in low abundance until disappearing in the late Griesbachian. A few Neogondolella, Neoclarkina, and Borinella species evolved from Clarkina during this time. Clarkina, Neogondolella, and Neoclarkina have very similar P1 elements, but the latter two can be distinguished from Clarkina with their different S0 elements (Henderson and Mei, Reference Henderson and Mei2007). In this paper, Clarkina is used for those conodonts that originated in the Permian and extended into the Early Triassic. As for Scythogondolella, Orchard (Reference Orchard and Krystyn2007) proposed an evolutionary lineage consisting of: Cl. nassichuki (Orchard in Orchard and Krystyn, Reference Orchard and Krystyn1998)—>Cl. krystyni (Orchard in Orchard and Krystyn, Reference Orchard and Krystyn1998)—>Cl. discreta (Orchard and Krystyn, Reference Orchard and Krystyn1998)—>Sc.? sp. A—>Smithian Scythogondolella. Scythogondolella? sp. A in the early Dienerian (Orchard, Reference Orchard2007, Orchard and Zonneveld, Reference Orchard and Zonneveld2009) and Ng. sp. B in the late Dienerian (Hatleberg and Clark, Reference Hatleberg and Clark1984, pl. 1, fig. 3) might be the ancestor of Smithian Scythogondolella. Its multielement apparatus of 15 elements differs from Neogondolella in both the P and S elements (Orchard, Reference Orchard2005, Reference Orchard2007; Goudemand et al., Reference Goudemand, Orchard, Tafforeau, Urdy, Brühwiler, Brayard, Galfetti and Bucher2012). However, because the multielement apparatuses have not been completely established, the genera of some species might be corrected in the future.
The geographical distribution of platform-bearing gondolellids (Fig. 8), based on published records and our new findings, are discussed below.

Figure 8. Global distribution of platform-bearing gondolellid conodonts in the Early Triassic (paleogeographic map is modified after Muttoni et al., Reference Muttoni, Gaetani, Kent, Sciunnach, Angiolini, Berra, Garzanti, Mattei and Zanchi2009 and Sun et al., Reference Sun, Joachimski, Wignall, Yan, Chen, Jiang, Wang and Lai2012; occurrences are given in the text).
Griesbachian
Some Clarkina species, including Cl. carinata (Clark, Reference Clark1959), Cl. planata (Clark, Reference Clark1959), Cl. orchardi (Mei, Reference Mei, Wang and Wang1996), Cl. kazi (Orchard in Orchard and Krystyn, Reference Orchard and Krystyn1998), Cl. meishanensis Zhang et al. in Zhang et al., Reference Zhang, Lai, Ding, Wu and Liu1995, Cl. zhejiangensis (Mei, Reference Mei, Wang and Wang1996), Cl. nassichuki (Orchard), Cl. taylorae (Orchard in Orchard et al., Reference Orchard, Nassichuk and Rui1994), Cl. tulongensis (Tian, Reference Tian1982), Cl. deflecta (Wang and Wang, Reference Wang and Wang1981), Cl. changxingensis (Wang and Wang, Reference Wang and Wang1981), Cl. prediscreta Wu et al., Reference Wu, Ji, Trotter, Yao and Zhou2014, Cl. postwangi (Tian, Reference Tian1993a, Reference Tianb), and Cl. hauschkei Kozur, Reference Kozur2004, survived the end-Permian mass extinction and lived alongside newly evolved species—Neogondolella griesbachensis Orchard, Reference Orchard2007, Ng. lehrmanni (Chen et al., Reference Chen, Beatty, Henderson and Rowe2009), Neoclarkina krystyni (Orchard in Orchard and Krystyn, Reference Orchard and Krystyn1998), and Nc. discreta (Orchard and Krystyn, Reference Orchard and Krystyn1998)—in the Griesbachian. They have been reported from northern low-latitude regions: South China (e.g., Jiang et al., Reference Jiang, Lai, Luo, Aldridge, Zhang and Wignall2007, Reference Jiang, Lai, Yan, Aldridge, Wignall and Sun2011; Zhang et al., Reference Zhang, Tong, Shi, Lai, Yu, He, Peng and Jin2007; Chen et al., Reference Chen, Beatty, Henderson and Rowe2009; Zhao et al., Reference Zhao, Chen, Chen and Cao2013), western USA (Clark et al., Reference Clark, Paul, Solien, Morgan, Sandberg and Clark1979); northern high-latitude areas: British Columbia (Mosher, Reference Mosher1973; Henderson, Reference Henderson1997; Orchard and Zonneveld, Reference Orchard and Zonneveld2009; Golding et al., Reference Golding, Orchard, Zonneveld, Henderson and Dunn2014), Canadian Arctic (Mosher, Reference Mosher1973; Henderson, Reference Henderson1997; Orchard, Reference Orchard2007, Reference Orchard2008), Svalbard (Nakrem et al., Reference Nakrem, Orchard, Weitschat, Hounslow, Beatty and Mørk2008), Greenland (Teichert and Kummel, Reference Teichert and Kummel1976); south margin of Neo-Tethys: Oman (Krystyn et al., Reference Krystyn, Richoz, Baud and Twitchett2003), West Pakistan (Sweet, Reference Sweet1970a), Kashmir (Sweet, Reference Sweet1970b), India (Goel, Reference Goel1977; Krystyn and Orchard, Reference Krystyn and Orchard1996; Orchard and Krystyn, Reference Orchard and Krystyn1998; Krystyn et al., Reference Krystyn, Balini and Nicora2004), Nepal (Hatleberg and Clark, Reference Hatleberg and Clark1984), Tibet (Tian, Reference Tian1982; Orchard et al., Reference Orchard, Nassichuk and Rui1994; Wang and Wang, Reference Wang and Wang1995; Orchard and Krystyn, Reference Orchard and Krystyn1998; Wu et al., Reference Wu, Ji, Trotter, Yao and Zhou2014); and Panthalassa: Japan (Igo, Reference Igo1989; Koike, Reference Koike1996). Borinella megacuspa Orchard, Reference Orchard2007, which originated in the late Griesbachian, has been reported from the Canadian Arctic (Orchard, Reference Orchard2008).
Overall, the distribution of platform-bearing gondolellids was still global following the end-Permian extinction, but they were only common in high-latitude regions, such as Svalbard and the Canadian Arctic (Klets, Reference Klets2008), and they were no longer flourishing in the low latitudes. However, Clarkina became extinct at the end of Griesbachian.
Dienerian
The diversity of platform-bearing gondolellids fell to a minimum in the Dienerian and they disappeared from equatorial regions. The last appearance of Neoclarkina discreta, Neogondolella griesbachensis, and Borinella megacuspa was in the Canadian Arctic (Orchard, Reference Orchard2008), while several new species of Borinella—B. nepalensis (Kozur and Mostler, Reference Kozur and Mostler1976), B. chowadensis (Orchard, Reference Orchard2007), B. sweeti (Kozur and Mostler, Reference Kozur and Mostler1976)—have been reported from British Columbia, West Pakistan, India (Orchard, Reference Orchard2007; Orchard and Krystyn, Reference Orchard and Krystyn2007) and Svalbard (Hatleberg and Clark, Reference Hatleberg and Clark1984, orignally assigned to Ng. elongata). And the new species Neogondolella mongeri (Orchard, Reference Orchard2007) was reported from British Columbia (Orchard, Reference Orchard2007; Orchard and Zonneveld, Reference Orchard and Zonneveld2009).
Smithian
Orchard et al. (Reference Orchard, Grădinaru, Nicora, Lucas and Spielman2007) considered the middle Smithian to be the heyday of Early Triassic conodonts. In additon to Borinella, three new genera (Scythogondolella, Neogondolella, and Paullella) that probably evolved in the late Dienerian or early Smithian have been found in Smithian. Among these, Scythogondolella was a dominant and cosmopolitan genus (Klets, Reference Klets2008; Orchard, Reference Orchard2008). In contrast, other platform-bearing gondolellids were rarer and less widespread. Scythogondolella is represented by seven species—Sc. mosheri, Sc. milleri, Sc. phryna, Sc. lachrymiformis Orchard, Sc. rhomboidea Orchard and Zonneveld, Sc. ellesmerensis Orchard, and Sc. crenulata (Mosher)—and five other species in open nomenclature (Sc.? sp. A Orchard and Sc. B–E Orchard), distributed across northern high-latitude regions: Siberia (Dagis, Reference Dagis1984; Klets and Yadrenkin, Reference Klets and Yadrenkin2001; Konstantinov et al., Reference Konstantinov, Sobolev and Yadrenkin2013), Canadian Arctic (Mosher, Reference Mosher1973; Kozur and Mostler, Reference Kozur and Mostler1976; Orchard, Reference Orchard2007, Reference Orchard2008; Orchard and Zonneveld, Reference Orchard and Zonneveld2009; Beranek et al., Reference Beranek, Mortensen, Orchard and Ullrich2010), British Columbia (Mosher, Reference Mosher1973; Orchard, Reference Orchard2008; Orchard and Zonneveld, Reference Orchard and Zonneveld2009; Golding et al., Reference Golding, Orchard, Zonneveld, Henderson and Dunn2014, 2015; Henderson et al., Reference Henderson, Golding and Orchard2018), Svalbard (Weitschat and Lehmann, Reference Weitschat and Lehmann1978; Hatleberg and Clark, Reference Hatleberg and Clark1984; Nakrem et al., Reference Nakrem, Orchard, Weitschat, Hounslow, Beatty and Mørk2008), South Primorye (Kozur and Mostler, Reference Kozur and Mostler1976; Buryi, Reference Buryi1979; Bondarenko et al., Reference Bondarenko, Zakharov, Guravskaya and Safronov2015), Qinghai, China (part of the Tarim Plate, Fang et al., Reference Fang, Zhang, Wang and Wang2013); the south margin of Neo-Tethys: Tibet (Tian, Reference Tian1982; Tian et al., Reference Tian, Dai and Tian1983; Zhao and Zhang, Reference Zhao and Zhang1991; Wang and Wang, Reference Wang and Wang1995; Zou et al., Reference Zou, Mao, Chen and Rao2006), India (Matsuda, Reference Matsuda1984), Timor Island (Nogami, Reference Nogami1968; Berry et al., Reference Berry, Burrett and Banks1984), West Pakistan and Nepal (Kozur and Mostler, Reference Kozur and Mostler1976; Hatleberg and Clark, Reference Hatleberg and Clark1984). Some also occurred in relatively low-latitude (~10°N) regions: western USA (Müller, Reference Müller1956; Clark and Mosher, Reference Clark and Mosher1966; Clark et al., Reference Clark, Paul, Solien, Morgan, Sandberg and Clark1979; Solien, Reference Solien1979; Orchard, Reference Orchard2008), Chaohu, South China (Liang et al., Reference Liang, Tong and Zhao2011), Qinling Basin, northwestern China (this paper), and Panthalassa: Japan (Nogami, Reference Nogami1968).
Newly evolved Neogondolella (Ng. altera Klets and Yadrenkin, Reference Klets and Yadrenkin2001, Ng. composita Dagis, Reference Dagis1984, Ng. jakutensis Dagis, Reference Dagis1984, Ng. sibirica Dagis, Reference Dagis1984) occurred in high-latitude Siberia (Dagis, Reference Dagis1984; Klets and Yadrenkin, Reference Klets and Yadrenkin2001) and Svalbard (Dagis and Korchinslaya, Reference Dagis, Korchinskaya, Dagis and Dubatolov1989). Borinella species (B. buurensis Dagis, Reference Dagis1984, B. chowadensis, B. nepalensis) and Paullella meeki (Paull, Reference Paull1983) are known from high northern latitudes regions: Siberia (Dagis, Reference Dagis1984), Canadian Arctic (Beranek et al., Reference Beranek, Mortensen, Orchard and Ullrich2010), British Columbia and western USA (Orchard, Reference Orchard2007, Reference Orchard2008, Orchard and Zonneveld, Reference Orchard and Zonneveld2009; Golding et al., Reference Golding, Orchard, Zonneveld, Henderson and Dunn2014, Reference Golding, Orchard, Zonneveld and Wilson2015; Henderson et al., Reference Henderson, Golding and Orchard2018), Svalbard (Hatleberg and Clark, Reference Hatleberg and Clark1984, originally assigned to Ng. elongata; Nakrem et al., Reference Nakrem, Orchard, Weitschat, Hounslow, Beatty and Mørk2008); and southern Neo-Tethys: India (Orchard and Krystyn, Reference Orchard and Krystyn2007; Orchard, Reference Orchard2010) and Nepal (Dagis, Reference Dagis1984).
Spathian
The bloom of Scythogondolella species was terminated in the late Smithian extinction along with Borinella and Paullella, leaving Neogondolella as the sole platform-bearing gondolellid survivor (although no species of this genus are known to have survived). The Spathian saw several new Neogondolella species appear—Ng. jubata Sweet, Reference Sweet1970a, Ng. amica Klets, Reference Klets1998, Ng. captica Klets, Reference Klets1998, Ng. taimyrensis Dagis, Reference Dagis1984, Ng. paragondolellaeformis Dagis, Reference Dagis1984, Ng. shevyrevi (Kozur and Mostler, Reference Kozur and Mostler1976), Ng. dolpanae Balini, Gavrilova, and Nicora, Reference Balini, Gavrilova and Nicora2000. They were joined by another four genera that originated in the late Spathian and ranged into Middle Triassic: Gladigondolella (Gl. malayensis Nogami, Reference Nogami1968, Gl. carinata Bender, Reference Bender1970), Columbitella elongata (Sweet, Reference Sweet1970a), Magnigondolella regalis (Mosher, Reference Mosher1970), and Spathogondolella jiarongensis Jiang and Chen in Chen et al., Reference Chen, Jiang, Lai, Yan, Richoz, Liu and Wang2015.
By the late Spathian platform-bearing gondolellids had returned to equatorial regions and thus they re-attained a global distribution. They occurred in equatorial regions: South China (Jiang, Reference Jiang1982; Wang, Reference Wang1982; Wang et al., Reference Wang, Wang, Li and Wie2005; Chen et al., Reference Chen, Jiang, Lai, Yan, Richoz, Liu and Wang2015) and Transcaucasia (Orchard, Reference Orchard2007); in the northern hemisphere: Siberia (Dagis, Reference Dagis1984; Klets, Reference Klets1998), Canadian Arctic (Mosher, Reference Mosher1973), Svalbard (Weitschat and Lehmann, Reference Weitschat and Lehmann1978; Hatleberg and Clark, Reference Hatleberg and Clark1984), British Columbia (Mosher, Reference Mosher1973; Orchard and Tozer, Reference Orchard, Tozer, Moslow and Wittenberg1997; Orchard, Reference Orchard2008), Western USA (Clark et al., Reference Clark, Paul, Solien, Morgan, Sandberg and Clark1979; Solien, Reference Solien1979; Carey, Reference Carey and Clark1984; Orchard and Tozer, Reference Orchard, Tozer, Moslow and Wittenberg1997; Orchard, Reference Orchard2005), Greece (Gaetani et al., Reference Gaetani, Jacobshagen, Nicora, Kauffmann, Tselepidis, Sestini, Mertmann and Coroneou1992), South Primorye (Buryi, Reference Buryi1979), and Romania (Orchard et al., Reference Orchard, Grădinaru, Nicora, Lucas and Spielman2007); in the southern hemisphere: Oman (Orchard, Reference Orchard1994), West Pakistan (Sweet, Reference Sweet1970a), India (Matsuda, Reference Matsuda1984), Tibet (Tian, Reference Tian1982, Reference Tian1983; Zhao and Zhang, Reference Zhao and Zhang1991; Wang and Wang, Reference Wang and Wang1995), and Nepal (Hatleberg and Clark, Reference Hatleberg and Clark1984).
Controls on the evolution of Early Triassic platform-bearing gondolellid conodonts
Some platform-bearing gondolellids were deep-water, nektobenthic groups, such as Clarkina and Neogondolella (Lai et al., Reference Lai, Wignall and Zhang2001), but others possibly prefer pelagic environments, such as Borinella and Gladigondolella (Orchard, Reference Orchard2007; Kozur et al., Reference Kozur, Moix and Ozsvart2009; Zhang et al., Reference Zhang, Sun, Lai, Joachimski and Wignall2017). The former are likely to have been affected by the substantial temperature and seafloor oxygenation fluctuations known to have affected Early Triassic marine habitats, and the latter have probably been mainly affected by the seawater temperature. In particular, the diversity and evolution of platform-bearing gondolellids appear to have closely followed the temperature record of the time (Sun et al., Reference Sun, Joachimski, Wignall, Yan, Chen, Jiang, Wang and Lai2012). The disappearance in late Griesbachian of Clarkina, which was dominant in end-Permian strata, coincides with a temperature rise that began at the Permo-Triassic boundary and peaked at the late Griesbachian. The rise saw the platform-bearing gondolellids of low diversity and abundance contract to relatively high-latitude regions in the Dienerian (Fig. 8). The subsequent cooling trend in the Dienerian, which peaked at the Dienerian/Smithian boundary, coincided with the appearance of new taxa, including Scythogondolella, Paullella, and several Neogondolella species. Temperature then rose again through the Smithian, peaking late in the substage. It appears that many conodonts could not cope with these high temperatures, especially in tropical latitudes, causing genera such as Neogondolella and Borinella to become restricted to high-latitude regions (Fig. 8). In contrast, Scythogondolella was tolerant of a broad range of temperatures and was found in all latitudes. However, the latest Smithian temperature peak coincides with the extinction of all platform-bearing gondollelids, except for Neogondolella, which saw a turnover among its constituent species. Finally, platform-bearing gondolellids returned to the equatorial regions and achieved global distribution again during late Spathian cooling.
Early Triassic marine oxygenation tracks the temperature oscillations reasonably closely, with peak intensity of anoxia during the warmest intervals (Griesbachian/earliest Dienerian and late Smithian) while the anoxicity of the Spathian appears to have been more regionally variable (Song et al., Reference Song, Wignall, Tong, Bond, Song, Lai, Zhang, Wang and Chen2012; Sun et al., Reference Sun, Wignall, Joachimski, Bond, Grasby, Sun, Yan, Wang, Chen and Lai2015; Wignall et al., Reference Wignall, Bond, Sun, Grasby, Beauchamp, Joachimski and Blomeier2016; Huang et al., Reference Huang, Chen, Wignall and Shao2017). Thus, the crises in platform-bearing gondolellid fortunes correspond with widespread anoxic episodes, although their subsequent Spathian radiation was at a time when anoxia was reasonably widespread. However, the extent of anoxia in Spathian may not have been sufficient to hinder radiation, and it is noteworthy at Yiwagou that this interval is represented by marine red beds.
Conclusions
Smithian platform-bearing gondolellid conodonts Scythogondolella mosheri, Sc. phryna, and Sc. cf. milleri have been discovered at Yiwagou, northwestern China, together with Novispathodus waageni waageni and Nv. w. eowaageni in the same sample. This is the first report of Smithian platform-bearing gondolellid conodonts in Paleo-Tethys. Additionally, Eurygnathodus costatus, E. hamadai, and Parafurnishius xuanhanensis are also here reported from the Dienerian to Smithian interval at the Yiwagou Section. Three conodont zones have been established from Dienerian to Smithian, they are in ascending order: Eurygnathodus costatus-E. hamadai Assemblage Zone, Novispathodus waageni-Scythogondolella mosheri Assemblage Zone, and Pachycladina-Parachirognathus Assemblage Zone.
The fluctuating fortunes of the platform-bearing gondolellids closely follow the seawater temperature record with low diversity related to peaks of temperature in Early Triassic seas. Compilation of global platform-bearing gondolellid occurrences shows that high temperatures in the late Griesbachian–early Dienerian and in the Smithian saw their retreat to higher, cooler latitudes. Scythogondolella is shown to be both a thermophilic and eurythermic genus, which was successful during the high temperatures of the Smithian, when other platform-bearing gondolellids disappeared from tropical latitudes. However, the peak temperatures late in the Smithian may have been too much for even Scythogondolella to survive.
Closely linked oxygenation trends may have also played a role in controlling diversity. However, the occurrence of marine red beds in the Smithian suggests oxygenation levels were frequently extremely good in the shallow-water carbonate platform at Yiwagou (e.g., Lai and Xu, Reference Lai, Xu, Yin, Yang, Huang, Yang and Lai1992).
Acknowledgments
This work was supported by the Natural Sciences Foundation (grant nos. 41572002, 41661134047). We thank H.X.S. Dong, Y.C. Li, X.H. Niu, H. Song, P.F. He, L.F. Li, H.Y. Ma, Z.J. Ma, L. Kang, and M.Y. Wang for their great help in field assistance. Special thanks to Z.Y. Sun and Y. Chen for their personal communication about this paper. We thank two anonymous reviewers for their helpful comments and constructive suggestions.








