


Introduction
The Middle to Late Ordovician was an important interval in the early evolutionary history of crinoids. Following their earliest known occurrence in the Lower Ordovician (Tremadocian), crinoids underwent a major taxonomic radiation as part of the Great Ordovician Biodiversification Event (GOBE) (Ausich and Deline, Reference Ausich and Deline2012; Wright and Toom, Reference Wright and Toom2017), which has been recognized globally across diverse invertebrate groups (Webby et al., Reference Webby, Paris, Droser and Percival2004). For most crinoid clades, peak diversity was reached during the Sandbian to Katian (Peters and Ausich, Reference Peters and Ausich2008; Cole et al., Reference Cole, Ausich, Colmenar and Zamora2017). As a result, crinoid assemblages from this interval represent the earliest development of complex crinoid communities with high species richness, high morphological disparity, and diverse niche space occupation (Foote, Reference Foote1994, Reference Foote1999; Brower, Reference Brower2007; Deline, Reference Deline2009; Deline et al., Reference Deline, Ausich and Brett2012). Thus, increased documentation of the morphology and taxonomic diversity of crinoids from this interval is important for studies of crinoid paleoecology, phylogeny, and macroevolutionary trends.
Fossil crinoids are well known from the Upper Ordovician (Sandbian–Katian) rocks of southern Ontario, particularly from the Bobcaygeon and Verulam formations in the Lake Simcoe region. Although this crinoid fauna has been recognized for nearly a century and a half (e.g., Billings, Reference Billings1858, Reference Billings1859; Springer, Reference Springer1911), little attention has been paid to the taxonomy of the crinoids within the fauna. Recent extensive collection of echinoderm material from the Bobcaygeon and Verulam formations near Brechin, Ontario has produced extensive collections of complete crinoid specimens, many with arms, stems, and attachment structures intact. The exceptional preservation of the recovered echinoderm material merits the designation of the fauna as a Konservat-Lagerstätte, and it is subsequently referred to herein as the “Brechin Lagerstätte.” This study provides a comprehensive re-evaluation of the dicyclic camerate crinoids from the Brechin Lagerstätte by reviewing taxonomic assignments, revising species descriptions to include morphological features that were previously unknown, and describing new taxa. In this contribution, we discuss all known taxa from the Brechin Lagerstätte that belong to Diplobathrida sensu Cole (Reference Cole2017) as well as the dicyclic stem eucamerates (i.e., reteocrinids). In a series of subsequent papers, we will address the monobathrid, cladid, disparid, flexible, and hybocrinid crinoids from the fauna.
Stratigraphy and geologic setting
The Brechin Lagerstätte crinoids described herein were recovered from multiple horizons of the Bobcaygeon and Verulam formations exposed in the vicinity of Brechin, Ontario (Fig. 1). The Bobcaygeon Formation is stratigraphically older and is overlain by the Verulam Formation. Both formations are part of the middle Simcoe Group, which is equivalent to the Trenton and Ottawa groups (Armstrong, Reference Armstrong2000). The Bobcaygeon and Verulam formations are currently considered Late Ordovician (late Sandbian–Katian) in age (Sproat et al., Reference Sproat, Jin, Zhan and Rudkin2015), with the M4-M5 sequence boundary of eastern Laurentia occurring within the Bobcaygeon Formation (Holland and Patzkowsky, Reference Holland and Patzkowsky1996). Because the fossiliferous horizons of the Bobcaygeon are largely restricted to the upper member of the formation, the crinoid material described here is Katian in age (Liberty, Reference Liberty1969; Brookfield and Brett, Reference Brookfield and Brett1988).

Figure 1 Locality map: (1) location of study area in southern Ontario, indicated by black box; (2) location of collection sites including the classic Kirkfield Quarry locality (stars) and nearby towns (filled circles) in the Lake Simcoe region.
Both the Bobcaygeon and Verulam formations are comprised of bioclastic wackestones, grainstones, and packstones interbedded with calcareous shales and siltstones. The Verulam contains more shale than the underlying Bobcaygeon Formation (Liberty, Reference Liberty1969). Several hardgrounds have been documented in detail from the upper Bobcaygeon and lower Verulam (Brett and Liddell, Reference Brett and Liddell1978; Brett and Brookfield, Reference Brett and Brookfield1984; Brett and Taylor, Reference Brett and Taylor1999). These hardground surfaces are commonly heavily burrowed and represent incipient lithification of the sediment that allowed many organisms to encrust the substrate, including edrioasteroids, bryozoans, and holdfasts of crinoids and other pelmatozoans (Brett and Liddell, Reference Brett and Liddell1978; Brett and Taylor, Reference Brett and Taylor1999; Sumrall and Gahn, Reference Sumrall and Gahn2006). The paleoenvironments in which the Bobcaygeon and Verulam formations were deposited have been interpreted as a proximal carbonate shelf that ranged in depth and proximity from shoal to shallow shelf in the Bobcaygeon and from deep shelf to shoal/shallow shelf in the Verulam (Armstrong, Reference Armstrong2000).
Faunal constituents of the Brechin Lagerstätte have been reported in the literature from other units in southern Ontario, such as the Kirkfield, Hull, and Cobourg formations, many of which are correlative to the Bobcaygeon and/or Verulam. However, the names for many of these units have changed extensively. Here, we use the most recent and widely accepted nomenclature for these units rather than their historical names. The Bobcaygeon Formation has replaced many units mentioned in older stratigraphic literature, including the Coboconk, Kirkfield, Rockland, Leray, and Hull formations (Liberty, Reference Liberty1969; Uyeno, Reference Uyeno1974; Armstrong, Reference Armstrong2000; see Swisher et al., Reference Swisher, Westrop and Amati2015). The Kirkfield Formation of earlier workers is equivalent to the middle–upper Bobcaygeon Formation (Liberty, Reference Liberty1967, Reference Liberty1969; Brookfield and Brett, Reference Brookfield and Brett1988; Brett and Taylor, Reference Brett and Taylor1999; Armstrong, Reference Armstrong2000). The Cobourg Formation, Ottawa Group (Uyeno, Reference Uyeno1974) of earlier workers is equivalent to the lower member of the Lindsay Formation (Liberty, Reference Liberty1967; Williams, Reference Williams1991; Armstrong and Carter, Reference Armstrong and Carter2010; Gabdeyan, Reference Gabdeyan2011), which gradationally overlies the Verulam Formation (Brookfield and Brett, Reference Brookfield and Brett1988; Armstrong, Reference Armstrong2000).
Overview of the Brechin Lagerstätte
Previous work
Crinoids from the Upper Ordovician Trenton Group of Canada were first described by Billings in 1858 and 1859, with preliminary descriptions published in 1856 and 1857. Although the material came from multiple localities, a notable collection was recovered from the vicinity of Kirkfield, Ontario (Fig. 1), but the descriptions of fossil crinoids were cursory. In 1911, Frank Springer produced a more thorough summary of Billings’ Kirkfield collection and documented the taxa recognized from the fauna, but even this treatment was not comprehensive. For example, Springer (Reference Springer1911) listed nine species of camerate crinoids from the Kirkfield collection, but only Reteocrinus alveolatus and Cleiocrinus regius were described or discussed at any length. The remaining seven species recognized were only mentioned in brief comments with regards to their relative degree of commonness or rarity. Since Springer’s (Reference Springer1911) monograph, material from Kirkfield and correlative collections in Ontario has not received a comprehensive taxonomic treatment. As a result, the fauna is widely recognized by professional and amateur paleontologists alike, yet remains poorly understood from a taxonomic perspective.
Taphonomy
The crinoid skeleton is composed of numerous, discrete plates that may be weakly cemented, as in the case of camerate crinoids, or uncemented, as in the case of cladids and disparids. In particular, the calyx and arms typically disarticulate quickly after death unless burial and preservation of the specimen occurs rapidly (Brett and Baird, Reference Brett and Baird1986; Brett et al., Reference Brett, Moffat and Taylor1997). Although columnals from the stem are typically robust and may be preserved as isolated elements, it is rare for the entire stem and holdfast to be found in association with the calyx. As a result, fully articulated crinoid calyces with complete arms are uncommon, and intact stems and holdfasts in association with calyces are exceedingly rare. Likewise, other echinoderms are also susceptible to rapid disarticulation because of their multi-elemental skeleton. Only exceptional environmental conditions will result in preservation of complete echinoderm specimens (Brett et al., Reference Brett, Moffat and Taylor1997). As a result, the exceptional preservation of echinoderm material from the Lake Simcoe region merits designation of the fauna as a Konservat-Lagerstätte.
Echinoderms from the Brechin Lagerstätte most commonly occur either on hardground surfaces from the uppermost 3–4 m of the Bobcaygeon Formation or in high-density clusters on non-hardground bedding planes, which are most common in the lower 2–3 m of the Verulam Formation, but can also be found in the upper Bobcaygeon (Brett and Taylor, Reference Brett and Taylor1999). Although echinoderms are exceptionally preserved in both types of assemblages, those from hardground surfaces are more likely to retain holdfast structures. This in situ preservation indicates that rapid, episodic events repeatedly buried the community, resulting in preservation of intact arms, stems, and attachment structures (Brett and Baird, Reference Brett and Baird1986). Specimens preserved in high-density assemblages are usually found in shale-filled topographic lows and may be associated with skeletal debris (Brett and Taylor, Reference Brett and Taylor1999); Cupulocrinus and Pleurocystites are particularly common in these assemblages. Although crinoid stems and holdfasts are less commonly preserved in these horizons, calyces and feeding structures typically remain articulated. In order for delicate structures like these to have remained intact, only limited disturbances of the sediment could have occurred after burial. Thus, rock slabs from the Brechin Lagerstätte preserving multiple specimens, whether on hardground surfaces or in high-density concentrations on non-hardground surfaces, can be taken to represent ecological snapshots with little to no time averaging.
Faunal constituents
In addition to more than 20 crinoid genera, the Brechin Lagerstätte also preserves a diverse echinoderm fauna representing at least eight other major echinoderm groups, including asteroids, ophiuroids, edrioasteroids, cystoids, cyclocystoids, homalozoans, edrioblastoids, and paracrinoids. The cystoid Pleurocystites squamosus Billings, Reference Billings1874 is particularly abundant, often occurring in dense associations on bedding planes in the upper Bobcaygeon Formation. Although the non-crinoid echinoderms have not been critically evaluated from a taxonomic perspective, preliminary investigation indicates at least 19 non-crinoid genera occur in the Brechin Lagerstätte. Previous work on the hardgrounds and paleoecology of the Bobcaygeon and Verulam formations has paid particular attention to hardground-encrusting echinoderms such as edrioasteroids and paracrinoids (e.g., Brett and Liddell, Reference Brett and Liddell1978; Sumrall and Gahn, Reference Sumrall and Gahn2006).
Materials and methods
Localities and specimen preparation
Although echinoderm fossils are known from several horizons in the Bobcaygeon and Verulam formations, the majority of the crinoid material described here was collected near the Bobcaygeon–Verulam contact in the Carden Quarry (44°34'335''N, 79°06'095''W), located 6 km east of Brechin, and the LaFarge Quarry (44°31'559''N, 79°09'478''W), located 2 km southeast of the town of Brechin. In both of these quarries, specimens were collected from blast piles sourced from stratigraphic intervals containing approximately 15 m of upper Bobcaygeon and 5 m of lower Verulam, referred to hereafter as the “Bobcaygeon-Verulam contact zone.” Material is predominately from the collection of J.M. Koniecki. Additional material examined is from the James Dick Quarry (44°29’937”N, 79°09’616”W, collection of K. Brett), which is lower Verulam, and the classic Kirkfield Quarry (44°35'6.32''N, 78°58'8.16''W), which is middle–lower Bobcaygeon. The classic Kirkfield Quarry locality is now flooded and known as Kirkfield Lake.
Specimens of Cleiocrinus lepidotus n. sp. from the Cincinnati Museum Center Invertebrate Paleontology Collections were prepared and/or reconstructed from disarticulated material by S.R. Cole; Archaeocrinus maraensis n. sp. UMMP 74688 was prepared by K. Brett; all other specimens were prepared by J. Koniecki. Unless otherwise noted, specimens were photographed after coating with ammonium chloride.
Phylogenetic analysis
Phylogenetic analysis can play a useful role in alpha taxonomy as a quantitative method for identifying closely related taxa, thereby allowing new taxa to be classified and diagnosed in evolutionarily meaningful ways. This approach complements traditional taxonomy by allowing quantitatively generated hypotheses of evolutionary history to be combined with assessment of homologous morphologic features. Here, a phylogenetic analysis was conducted to examine relationships among dicyclic camerates from the Brechin Lagerstätte. Three of the four genera identified from the Brechin Lagerstätte (Cleiocrinus, Archaeocrinus, and Reteocrinus) were included in a recent phylogenetic analysis that inferred relationships among all Ordovician camerate crinoids (Cole, Reference Cole2017). As a result, the primary focus of the analysis conducted here was to examine the evolutionary relationships of Priscillacrinus elegans n. gen. n. sp., described herein. Priscillacrinus n. gen. was coded for the same 112 discrete morphological characters described by Cole (Reference Cole2017). Using the phylogeny of Ordovician camerate genera recovered by Cole (Reference Cole2017) as a constraint, the placement of Priscillacrinus n. gen. within the tree was analyzed in PAUP* using a heuristic parsimony search with tree bisection reconnection and 1000 random addition sequences (Swofford, Reference Swofford2003).
The phylogenetic analysis recovered a single most-parsimonious tree (Fig. 2). The placement of Priscillacrinus n. gen. relative to Cleiocrinus, Archaeocrinus, and Reteocrinus reveals that the assemblage of dicyclic taxa in the Brechin Lagerstätte has high phylogenetic diversity, meaning that diversity is distributed across the tree rather than being restricted to a particular clade of closely related taxa. The recovered tree identifies Cotylacrinna, which is from the Sandbian of Iowa, as the genus most closely related to Priscillacrinus n. gen. These two genera form a clade with Goyacrinus, Anthracocrinus, and Rheocrinus, all of which are assigned to the family Anthracocrinidae. Although Cotylacrinna is currently assigned to Rhodocrinitidae rather than Anthracocrinidae, it has been advocated that this genus be reassigned to Anthracocrinidae because it shares the features that are diagnostic of the family (Cole, Reference Cole2017). As a result, the position of Priscillacrinus n. gen. within this clade suggests it belongs within the Anthracocrinidae. Further support for this conclusion on the basis of skeletal morphology is given below following the systematic description of Priscillacrinus n. gen.

Figure 2 Phylogenetic placement of Priscillacrinus n. gen. (indicated by star) relative to other Ordovician camerate genera.
Repositories and institutional abbreviations
New material described here is reposited in the University of Michigan Museum of Paleontology (UMMP). Additional material examined is from the Cincinnati Museum Center Invertebrate Paleontology Collection (CMCIP) and the Smithsonian National Museum of Natural History (NMNH). Type material not examined is from the Geological Survey of Canada (GSC) and the Field Museum of Natural History (FMNH).
Systematic paleontology
Classification and terminology
The classification of crinoid higher taxa used here follows Ausich et al. (Reference Ausich, Kammer, Rhenberg and Wright2015), Cole (Reference Cole2017), and Wright et al. (Reference Wright, Ausich, Cole, Rhenberg and Peter2017). Other aspects of suprageneric classification used herein follow Moore and Teichert (Reference Moore and Teichert1978) and Brower and Veinus (Reference Brower and Veinus1974).
Morphologic terminology follows Ubaghs (Reference Ubaghs1978a). The plating of interrays is given in the standard abbreviated form as the number of plates in each range from the proximal-most plate to the last range before the tegmen. In the posterior interray, the primanal is indicated by “P” and the first interradial in regular interrays is indicated by “1.” The shorthand notation used to describe the nodal-internodal structure of heteromorphic columns is from Webster (Reference Webster1974).
Class Crinoidea Miller, Reference Miller1821
Subclass Camerata Wachsmuth and Springer, Reference Wachsmuth and Springer1885
Family Reteocrinidae Wachsmuth and Springer, Reference Wachsmuth and Springer1885
Included genera
Reteocrinus Billings, Reference Billings1859 and Quechuacrinus Guensburg and Waisfeld, Reference Guensburg and Waisfeld2015.
Diagnosis
Dicyclic camerates with prominent ray ridges; depressions or gaps between basal and infrabasal plates; radial circlet interrupted in all rays; posterior with ray-like anitaxial series; interrays depressed, presumably not ankylosed or only weakly cemented, comprised of small and irregularly arranged plates; position of primaxil highly variable; arms uniserial, apinnulate; stem pentameric in at least distal portion.
Remarks
In addition to Reteocrinus, the genera Gaurocrinus, Cnemecrinus, and Quechuacrinus were previously assigned to family Reteocrinidae (Guensburg and Waisfeld, Reference Guensburg and Waisfeld2015). Phylogenetic evaluation of all Ordovician camerate crinoids by Cole (Reference Cole2017) did not recover Gaurocrinus or Cnemecrinus as members of Reteocrinidae, although Cnemecrinus is closely related to the family. The diagnosis for Reteocrinidae given above is emended from recent diagnoses (Guensburg, Reference Guensburg1984; Guensburg and Waisfeld, Reference Guensburg and Waisfeld2015) to reflect exclusion of Gaurocrinus and Cnemecrinus from the family (Cole, Reference Cole2017).
Genus Reteocrinus Billings, Reference Billings1859
Type species
Reteocrinus stellaris Billings, Reference Billings1859.
Diagnosis
Reteocrinids with bilateral symmetry through A ray and CD interray; interrays depressed, not ankylosed, comprised of numerous small, irregularly arranged plates; median ray ridges prominent; infrabasals upright, visible in side view; depressions between basal plates and variably between infrabasal plates, in some cases filled with small irregular plates; posterior interray wide with prominent anitaxial ridge originating from CD basal; first arm branching varies in position from radial plate to sixth primibrachial; fixed brachials bifurcating several times within the calyx; free arms uniserial, apinnulate, branched; stem circular, pentagonal, or pentalobate, commonly pentameric in dististele.
Occurrence
Upper Ordovician (Sandbian–Katian); United States (Oklahoma, Tennessee, Ohio, Illinois) and Canada (Ontario).
Remarks
Species-level characters in Reteocrinus include shape of the calyx, plate ornamentation, condition of the interinfrabasal areas, shape of the infrabasals, number of primibrachials, and stem characteristics (Kolata, Reference Kolata1982; Guensburg, Reference Guensburg1984). One of the defining characteristics of species of Reteocrinus is the presence of spaces between basal plates (interbasal gaps) and indentations between infrabasal plates (interinfrabasal depressions). The suturing of cup plates around these spaces gives the cup a reticulated appearance. In some species (e.g., R. stellaris), the interbasal gaps are filled with small plates.
Reteocrinus stellaris Billings, Reference Billings1859

Figure 3 Reteocrinus specimens from the Bobcaygeon-Verulam contact zone, southern Ontario. (1–3) Reteocrinus alveolatus UMMP 74675, (Carden Quarry), distal crown showing terminus of anal tube above arms; (1) lateral view of crown; (2) lateral view of crown, opposite side; (3) adoral view of anal tube and terminal arm tips; (4–6) Reteocrinus alveolatus UMMP 74673 (LaFarge Quarry); (4) lateral view of complete specimen with stem and holdfast; (5) lateral view of crown; (6) close-up of holdfasts attached to hardground surface; (7) Reteocrinus alveolatus UMMP 74674 (Carden Quarry); (8) Reteocrinus alveolatus UMMP 74672 (LaFarge Quarry); (9, 10) Reteocrinus stellaris, UMMP 74671 (Carden Quarry); (9) lateral view of crown and proximal stem; (10) close-up of calyx showing interbasal gap plates, photographed under alcohol. All scale bars 5 mm.
Holotype
GSC 1525 (lectotype).
1859 Reteocrinus stellaris Billings, p. 64, pl. 9, figs. 4a–e.
1868 Reteocrinus stellaris; Reference ShumardShumard, p. 394.
1868 Retiocrinus stellaris [sic]; Reference BigsbyBigsby, p. 23.
1883a Reteocrinus stellaris; Reference Wachsmuth and SpringerWachsmuth and Springer, p. 261, figs. 1–3.
1889 Reteocrinus stellaris; Reference MillerMiller, p. 277, fig. 416.
1897 Reteocrinus stellaris; Reference Wachsmuth and SpringerWachsmuth and Springer, p. 178, pl. 9, figs. 3a–c.
1910 Reteocrinus stellaris; Reference Grabau and ShimerGrabau and Shimer, p. 547.
1911 Reteocrinus stellaris; Springer, p. 10, pl. 1, figs. 6, 7.
1915 Reteocrinus stellaris; Reference BasslerBassler, p. 1100.
1938 Reteocrinus stellaris; Reference BasslerBassler, p. 163.
1943 Reteocrinus stellaris; Reference Bassler and MoodeyBassler and Moodey, p. 660.
1946 Reteocrinus stellaris; Reference WilsonWilson, p. 25.
1973 Reteocrinus stellaris; Reference WebsterWebster, p. 230.
1978b Reteocrinus stellaris; Reference UbaghsUbaghs in Moore and Teichert, p. T414, figs. 223.1a, b.
1984 Reteocrinus stellaris; Guensburg, p. 28, pl. 4, figs. 1–3.
1985 Reteocrinus stellaris; Reference SmithSmith, p. 168, pl. 7.4.10.
1986 Reteocrinus stellaris; Reference WebsterWebster, p. 274.
1988 Reteocrinus stellaris; Reference WebsterWebster, p. 145.
Diagnosis
Reteocrinus with three primibrachials; radials, basals, and infrabasals ornamented with angular keeled ridges that surround interbasal depressions; fixed brachials lacking lateral projections; interbasal depressions filled with small, irregular plates; interinfrabasal depressions shallow or indistinct; stem circular.
Occurrence
Previously, R. stellaris had been reported from Upper Ordovician (Katian) occurrences from the Hull and Cobourg formations, Ottawa Group, Ontario. The occurrence reported here is from the Bobcaygeon–Verulam contact zone, Carden Quarry, Upper Ordovician (lower Katian).
Materials
UMMP 74671 (Bobcaygeon–Verulam contact zone, Carden Quarry).
Remarks
Thorough description of R. stellaris is provided in Guensburg (Reference Guensburg1984), so a redescription is not necessary. Relative to R. alveolatus, R. stellaris is uncommon in the collection described here from the Brechin region. Unlike R. alveolatus, specimens of this species display small, polygonal plates filling the interbasal gaps (Fig. 3.10)
Reteocrinus alveolatus (Miller and Gurley, Reference Miller and Gurley1894)
Holotype
FMNH 6045.
1894 Retiocrinus alveolatus Miller and Gurley [sic], p. 26, pl. 2, fig. 22.
1897 Retiocrinus alveolatus [sic]; Reference MillerMiller, p. 752, fig. 1390.
1911 Reteocrinus alveolatus; Springer, p. 10, pl. 1, figs. 1–5.
1915 Reteocrinus alveolatus; Reference BasslerBassler, p. 1100.
1943 Reteocrinus alveolatus; Reference Bassler and MoodeyBassler and Moodey, p. 659.
1978b Reteocrinus alveolatus; Reference UbaghsUbaghs in Moore and Teichert, p. T414, fig. 223.1c, d.
1984 Reteocrinus alveolatus; Guensburg, p. 29, pl. 4, figs. 5–7, 9, 14, 15.
Diagnosis
Reteocrinus with two to four primibrachials; cup plates with rounded keeled ridges; cup plates and fixed brachials smooth or ornamented with faint, irregular, discontinuous ridges; fixed brachials with spine-like processes projecting laterally into interradial areas; interinfrabasal gaps deep, small, diamond-shaped to semicircular; interbasal gaps deep, filled with small, irregular plates; stem pentagonal to pentastellate, distally pentameric.
Occurrence
Reteocrinus alveolatus was previously known from Upper Ordovician (Katian) occurrences in the Curdsville Member of the Lexington Formation, Kentucky and from the Hull Formation, Ottawa Group, Ontario. The occurrences reported here are from the Bobcaygeon-Verulam contact zone, LaFarge and Carden Quarries, Upper Ordovician (lower Katian).
Description
Calyx straight-sided cone to height of radials, cylindrical above radials with straight or slightly convex sides, higher than wide, lobate; crown about twice as high as maximum calyx width. Cup plates deeply cleft, smooth, with multiple rounded limbs; fixed brachial plates smooth or ornamented with faint, irregular, discontinuous ridges; brachials thick, raised from surface of calyx, forming prominent rounded median ray ridges; plate sutures flush with surface of plates.
Infrabasal circlet upright, entirely visible in side view; infrabasal plates five, wider than high, nearly as large as basals. Interinfrabasal gaps small and deep, not in contact with stem margin, shape variably triangular, subrounded, or diamond-shaped.
Basal circlet entirely visible in side view; basal plates x- or y-shaped, approximately as high as wide, largest plates in calyx; CD basal plate pentaxial (stellate with five rays), usually higher than wide, supporting primanal. Interbasal gaps deeply depressed, diamond-shaped to semi-circular, filled with small, irregularly arranged stellate plates similar to plating of interrays.
Radial circlet interrupted in all interrays; radial plates five, inverted y-shape, higher than wide, smaller than basals.
Regular interrays narrow, depressed, in contact with tegmen, maintaining equal width or constricted only in distal-most portion. Interray plating irregular, consisting of many small polygonal plates; interray plates with coarse stellate ornamentation, presumably not rigidly cemented.
Posterior interray wider, in contact with tegmen; anal series conspicuous, ray-like, comprised of a single column of 10 or more narrow plates originating from the CD basal and continuing onto the tegmen; primanal rectangular, about three times higher than wide, higher plates in series decreasing slightly in size; anal series flanked by narrow, depressed areas with plating identical to that of regular interrays.
Primibrachials two to five, variable within specimens; first primibrachial quadrangular, slightly wider than high. Secundibrachials typically three, less commonly two or four, variable within specimens, approximately as high as wide. Fixed intrabrachial plates within rays, plating similar to that of regular interrays.
Arm openings probably 20, ungrouped, position at which arms become free indistinct. Free arms apinnulate, poorly isotomously branched, up to six bifurcations in fixed and free brachials combined; brachials rectilinear uniserial. Anal opening unknown.
Tegmen contiguous with interrays, sac-like, reaching to the height of the arms, terminating in a rounded tip; tegmen plates small, nodose to coarsely stellate, irregularly arranged, similar to interray plates.
Stem pentameric, heteromorphic N333233313332333 where fully differentiated, stem meres thin; proximal stem pentastellate to pentalobate, gradually expanding near the base of the calyx, commonly with sharp ridges running lengthwise along axes of pentastellate stem; medial stem grading into pentagonal shape; distal stem circular to obscurely pentagonal. Lumen small, circular, surrounded by five small canals, unconnected to the lumen, positioned in the stem lobes. Holdfast circular to subcircular, composed of many small, smooth, irregular plates, commonly with a central depression where the stem attaches, cemented to the seafloor, diameter approximately equal to that of the calyx.
Materials
UMMP 74672 and 74673 (Bobcaygeon-Verulam contact zone, LaFarge Quarry); UMMP 74671, 74674, 74675, and 74677.1 (Bobcaygeon-Verulam contact zone, Carden Quarry).
Remarks
Although R. alveolatus has been previously described from abundant material, the tegmen and anal tube was not previously known. The description given here includes details of these structures, which are preserved in UMMP 74675.
Infraclass Eucamerata Cole, Reference Cole2017
Order Diplobathrida Moore and Laudon, Reference Moore and Laudon1943
Superfamily Rhodocrinitoidea Roemer, Reference Roemer1855
Family Anthracocrinidae Strimple and Watkins, Reference Strimple and Watkins1955
Genus Priscillacrinus new genus
Type species
Priscillacrinus elegans new species, by monotypy.
Diagnosis
As for species, by monotypy.
Etymology
The genus name Priscillacrinus is in honor of Priscilla Wright, mother of D.F. Wright.
Remarks
Superficially, Priscillacrinus n. gen. appears similar to the rhodocrinitid Diabolocrinus, but is differentiated by its additional bifurcation within the fixed brachials. Priscillacrinus n. gen. is assigned to Anthracocrinidae on the basis of its basal concavity involving infrabasals and partial basals, prominent median ray ridges, fixed brachials bifurcating twice within the calyx, regular interray plating 1–2 with the first interray plate enlarged, 20 free arm openings, and fixed pinnules. Additionally, phylogenetic analysis recovered Priscillacrinus n. gen. within the anthracocrinid clade (Fig. 2). In the recovered tree, Priscillacrinus n. gen. is most closely related to Cotylacrinna, a diplobathrid from the Middle Ordovician of Iowa (Brower, Reference Brower1994). Morphologically, Priscillacrinus n. gen. and Cotylacrinna share many features including a low, lobate calyx; thick calyx plates; interrays with proximal plating 1-2-3; at least two brachial bifurcations with the second branching occurring where arms become free; fixed intrabrachials between half rays but not quarter rays; fixed pinnules; and flat chisel biserial brachials. Priscillacrinus n. gen. differs from Cotylacrinna in that it possesses heavily ornamented cup plates, no ray lobes built from fixed brachials, multiple fixed pinnules in each ray, two secundibrachials in each quarter ray, and four free arms per ray. In contrast, Cotylacrinna has no plate ornamentation and only weak median ray ridges, distinctive ray lobes built from fixed brachials, fixed pinnules occurring only occasionally in rays, usually four secundibrachials per ray, and four to six free arms per ray.
Priscillacrinus elegans new species

Figure 4 Camerate crinoids from the Bobcaygeon-Verulam contact zone, LaFarge Quarry, southern Ontario. (1–4) Priscillacrinus elegans n. gen. n. sp.; (1) UMMP 74678.1, holotype, close-up of complete crown; (2) UMMP 74678.1 and 74678.2, holotype and paratype, two crowns with partial stems; (3) UMMP 74679, paratype, specimen with distal calyx and arms preserved; (4) UMMP 74680, paratype, calyx with proximal arms and partially disarticulated tegmen; (5) Cleiocrinus regius UMMP 74681.1; (6) Cleiocrinus sp. UMMP 74678.3, holdfast (left) and associated distal stem (right). All scale bars 5 mm unless otherwise noted.
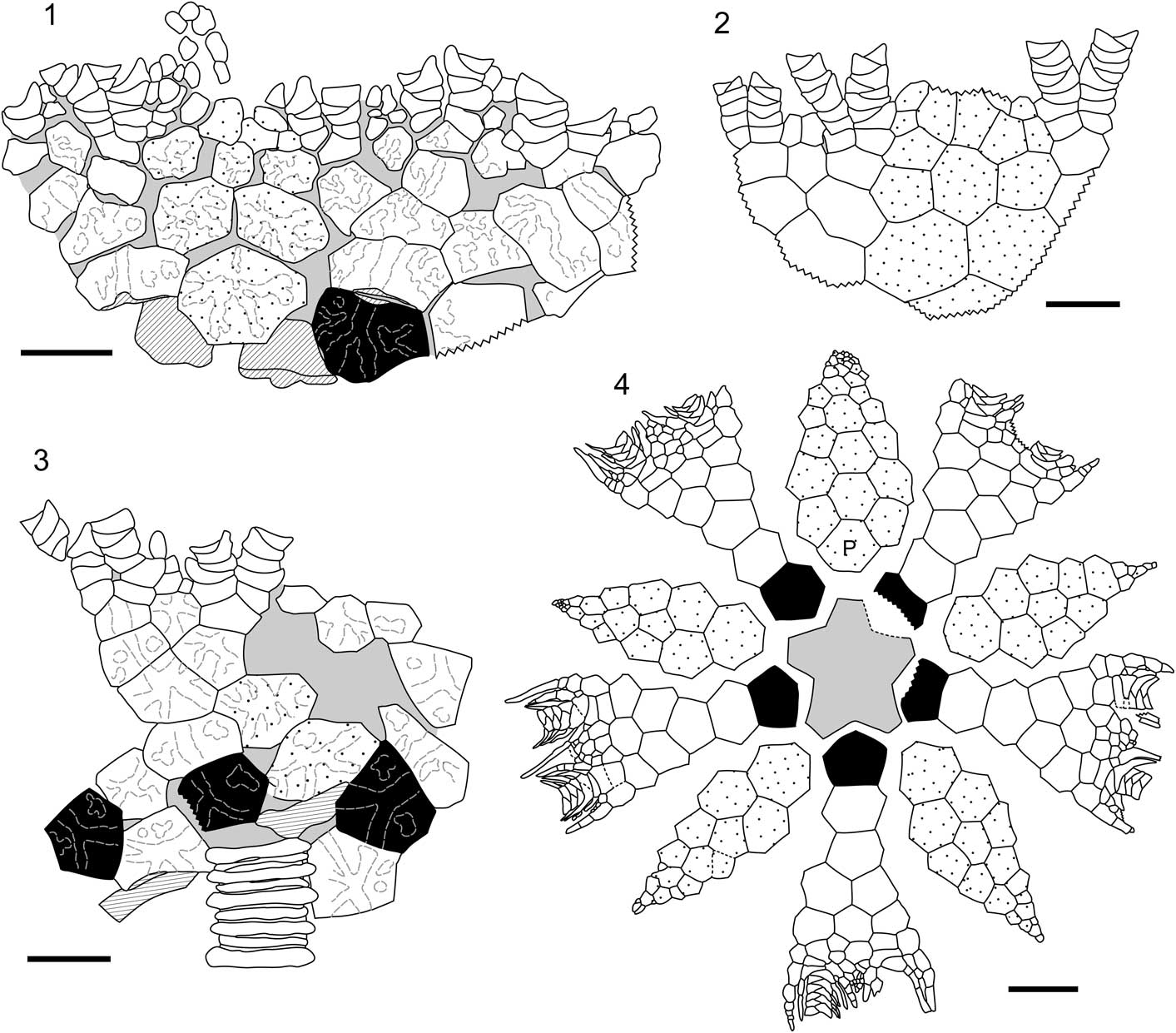
Figure 5 Camera lucida drawings of new taxa from the Bobcaygeon-Verulam contact zone, southern Ontario. (1–3) Priscillacrinus elegans n. gen. n. sp. (LaFarge Quarry); (1) UMMP 74678.1, holotype; (2) UMMP 74679, paratype; (3) UMMP 74678.2, paratype; (4) Archaeocrinus sundayae n. sp. UMMP 74682, holotype (Carden Quarry). All scale bars 5 mm; black fill = radials, gray fill = matrix/plating unknown, ruled fill = plate edges/undersides, stipple = interray plates, solid black lines = plate boundaries, dashed black lines = inferred plate boundaries, dashed gray lines = ray ridges/plate ornamentation, jagged lines = broken plate margins, P = primanal.
Holotype
UMMP 74678.1, holotype; 74678.2, paratype; 74679, paratype; 74680, paratype.
Diagnosis
Anthracocrinid with low bowl-shaped calyx; calyx plates thick; infrabasals and proximal basals involved in basal concavity; first interray plate largest in calyx; interrays wide with many large plates (~25), in contact with tegmen, proximal plating 1-2-3, higher rows irregular; fixed pinnules present; free arms 20, unbranched, wedge biserial.
Occurrence
The specimens documented here are from the Bobcaygeon-Verulam contact zone, LaFarge Quarry, Upper Ordovician (early Katian).
Description
Calyx low bowl shape, twice as wide as high; plates thick, ornamented with heavy irregular ridges; plate sutures flush with plate surface; arm rays ornamented with discontinuous ridges.
Infrabasals not observed, confined to the basal concavity. Basal circlet entirely visible in side view; basal plates five, hexagonal, wider than high, narrowing distally.
Radial circlet interrupted in all rays by contact of basals with primanal and first interradial plates; radial plates pentagonal, wider than high; approximately equal in size to basal plates.
Regular interrays in contact with tegmen, not depressed, interrupting the radial circlet in all rays. First interradial plate heptagonal, wider than high, slightly larger than radials and basals. Second range with two plates; proximal plating 1-2-3, followed by four to six higher rows each with three to six irregularly organized plates that decrease in size distally and grade into the tegmen.
Posterior interray undifferentiated or not observed.
First primibrachial fixed, hexagonal, more than twice as wide as high; second primibrachial axillary, pentagonal. Secundibrachials two, fixed, second secundibrachial axillary. Fixed tertibrachials usually giving rise abaxially to one fixed pinnule in each quarter ray, fixed pinnule placement variable, arms typically free above fourth tertibrachial. Fixed intrabrachials between secundibrachials and tertibrachials of each half ray, proximal plating 1-2-2, no fixed intrabrachials between quarter rays.
Arm openings 20, grouped by half rays, with robust arm bases protruding from the calyx. Free arms unbranched, tapering gradually, pinnulate; brachials wedge biserial throughout free arms.
Tegmen conical, nearly as high as calyx; tegmen plates thick, polygonal, similar in size and ornamentation to distal interray plates. Anal opening unknown.
Stem circular, holomeric; heteromorphic N212 in proximal and medial stem. Distal stem, holdfast, and lumen unknown.
Etymology
The specific name is after the Latin elegans, meaning elegant or graceful.
Materials
UMMP 74678.1, holotype; 74678.2, paratype; 74679, paratype; 74680, paratype (Bobcaygeon-Verulam contact zone, LaFarge Quarry).
Remarks
It does not appear that this or any other species of Priscillacrinus n. gen. was previously recognized from material collected from the Kirkfield region. It is one of the rarest species in the collection described herein, yet is distinctive because of its heavy ornamentation on the calyx plates.
Family Cleiocrinidae Miller, Reference Miller1890
Genus Cleiocrinus Billings, Reference Billings1857
Type species
Cleiocrinus regius Billings, Reference Billings1857.
Diagnosis
Calyx high conical or pyriform; calyx plates perforated by many pores along plate sutures; infrabasals in deeply invaginated basal concavity; basals and radials five each, in a single circlet of 10 plates that overlaps the stem and basal concavity; fixed brachials bifurcating asymmetrically numerous times within the calyx, giving rise to 40 or more arms and fixed ramules; interray plates absent, brachial plates sutured together; CD interray comprised of a single column of plates.
Occurrence
Middle–Upper Ordovician (Darriwilian–Katian); United States (Oklahoma, New York, Iowa, Tennessee, Kentucky) and Canada (Ontario).
Remarks
After contemplating the phylogenetic position of Cleiocrinus, Frank Springer (Reference Springer1911, p. 44) concluded, “…until better proofs of its relations are furnished it will have to remain in a sort of palaeontological no-man’s land.” Indeed, Cleiocrinus is a bizarre taxon unlike any other crinoid known, and its affinities with other taxa remain obscure. Originally, the genus was thought to have diverged from other diplobathrids early in the evolutionary history of camerates, and Cleiocrinus was assigned along with Spyridocrinus to suborder Zygodiplobathrina, whereas all other diplobathrids were assigned to Eudiplobathrina (Ubaghs, Reference Ubaghs1953, Reference Ubaghs1978). However, this superordinal division has not been supported by phylogenetic analysis (Cole, Reference Cole2017). Despite its many unusual apomorphic features, Cleiocrinus is well known and moderately diverse (see Springer, Reference Springer1905; Guensburg, Reference Guensburg1984).
Cleiocrinus regius Billings, Reference Billings1857
Holotype
GSC 1443a.
1857 Cleiocrinus regius Billings, p. 276.
1859 Cleiocrinus regius; Billings, p. 53, pl. 5, figs. 1a–g.
1868 Cleiocrinus regius; Reference ShumardShumard, p. 359.
1868 Cleiocrinus regius; Reference BigsbyBigsby, p. 18.
1889 Cleiocrinus regius; Reference MillerMiller, p. 232, fig. 264.
1905 Cleiocrinus regius; Springer, p. 110, pl. 1, figs. 1–10.
1910 Cleiocrinus regius; Reference Grabau and ShimerGrabau and Shimer, p. 562.
1911 Cleiocrinus regius; Springer, p. 41, pl. 5, figs. 7–9.
1915 Cleiocrinus regius; Reference BasslerBassler, p. 230.
1938 Cleiocrinus regius; Reference BasslerBassler, p. 67.
1943 Cleiocrinus regius; Reference Bassler and MoodeyBassler and Moodey, p. 365.
1946 Cleiocrinus regius; Reference WilsonWilson, p. 23, pl. 5, fig. 2.
1950 Cleiocrinus regius; Reference UbaghsUbaghs, p. 119, fig. 7.
1953 Cleiocrinus regius; Ubaghs, figs. 33d–f.
1973 Cleiocrinus regius; Reference WebsterWebster, p. 81.
1978a Cleiocrinus regius; Ubaghs in Moore and Teichert, p. T102, figs. 76.1, 174.1–3, 221.1a–d.
1986 Cleiocrinus regius; Reference WebsterWebster, p. 97.
1994 Cleiocrinus regius; Brower, p. 570, fig. 1.
1999 Cleiocrinus regius; Reference SimmsSimms in Hess et al., p. 32, figs. 54a, 54b.
Diagnosis
Cleiocrinus with an elongate calyx; distinct, narrow median ray ridges; plates ornamented with fine ridges crossing plate sutures; pores positioned along plate sutures, ~18 pores per 5 mm; two primibrachials, three to seven secundibrachials, four to twelve tertibrachials.
Occurrence
Cleiocrinus regius was previously known from the Upper Ordovician (Katian) Hull and Cobourg formations, Ottawa Group, Ontario and from the Upper Ordovician (Katian) Rivoli Member of the Dunleith Formation, Galena Group, Iowa. The occurrence documented here is from the Bobcaygeon-Verulam contact zone, LaFarge Quarry, Upper Ordovician (lower Katian).
Materials
UMMP 74681.1 (Bobcaygeon-Verulam contact zone, LaFarge Quarry); USNM S7137 (middle–upper Bobcaygeon Formation, Kirkfield Quarry).
Remarks
Extensive material is known for C. regius, and the species was described at length by Springer (Reference Springer1905).
Cleiocrinus lepidotus new species

Figure 6 Cleiocrinus lepidotus n. sp. from southern Ontario. (1, 5–8) CMCIP 76789, holotype (middle–upper Bobcaygeon Fm., Kirkfield Quarry); (1) lateral view of partial crown and stem, posterior interray indicated by arrow; (5) lateral view of partial crown, opposite side; (6) radial/basal circlet showing radial plate (center), wide fixed primibrachials, and adjacent basals; (7) proximal (upper) stem articular facet, photographed under alcohol; (8) distal (lower) stem articular facet, photographed under alcohol; (2, 3, 9–11) CMCIP 76790, paratype (middle–upper Bobcaygeon Fm., Kirkfield Quarry); (2) lateral view of distal calyx and arm bases; (3) posterior view of partial calyx; (9) close-up of fixed brachial plates showing pitted ornamentation; (10) closeup of fixed brachial plates showing sutural pores adjacent to plate corners, photographed under alcohol; (11) close-up of posterior interray; (4) UMMP 74681.2, paratype (Bobcaygeon-Verulam contact zone, LaFarge Quarry), lateral view of small partial calyx. All scale bars 5 mm unless otherwise indicated.
Holotype
CMCIP 76789, holotype; CMCIP 76790, paratype; UMMP 74681.2, paratype.
Diagnosis
Cleiocrinus with medium conical to bowl-shaped calyx; sutural pores few, positioned along plate sutures to either side of each plate corner; plates ornamented with fine pitting; median ray ridges narrow, rounded; 20 free arms; abundant fixed pinnules.
Occurrence
The occurrences documented here are from the middle–upper Bobcaygeon Formation, Kirkfield Quarry, Upper Ordovician (lower Katian) and from the Bobcaygeon–Verulam contact zone, LaFarge Quarry, Upper Ordovician (lower Katian).
Description
Calyx medium cone with bowl-shaped base, as high as wide; plates ornamented with fine pitting; median ray ridges narrow, rounded. Upper and lower margins of fixed brachial plates undulatory, medial portion of distal margin deflected downward; distal plate margins slightly elevated from the calyx surface and appearing to overlap the plates above, giving the appearance of weak imbrication.
Sutural pores positioned along plate sutures, relatively large in size compared to other species, typically eight per plate, positioned to either side of each plate corner; pores more closely spaced in more distal plates.
Infrabasals not observed; infrabasal circlet probably confined to basal concavity and overlapped by higher circlet as is typical in other Cleiocrinus species.
Basals and radials arranged in a single circlet of 10 plates. Basals small, pentagonal, higher than wide, with angled surface directed upwards. Radials slightly smaller than basals, rectangular as known, wider than high.
Rays comprised of numerous fixed brachials that are fused laterally; interray plates absent. Posterior interray imperfectly preserved, articulation with CD basal not observed; middle to higher portion of posterior interray forming an anal series comprised of a single column of wide, low plates; anal series plates four to five times wider than high, upper and lower margins strongly undulatory and similar in shape to the patelloid process of flexible crinoids. Anal series ornamented by a broad, protruding anitaxial ridge.
Primibrachials two; first primibrachial roughly hexagonal, largest plate in calyx, more than twice as wide as high, resting on underlying radial and sloping upper margins of adjacent basals; second primibrachial axillary, wider than high. Secundibrachials three in preserved rays, third axillary, all secundibrachials approximately twice as wide as high. Tertibrachials four to six, bifurcating at different positions within rays, dimensions becoming more equant distally, but still wider than high throughout. Quartibrachials variable in number, giving rise to one arm and two large fixed pinnules per quarter ray; 20 free arms and 40 large fixed pinnules in total; higher quartibrachials giving rise to additional, smaller fixed pinnules that grade into free pinnules. Undulatory pattern of plate margins becoming increasingly exaggerated in higher fixed brachials.
Arm openings 20; two large fixed pinnules positioned between each arm, 40 large fixed pinnules in total. Brachials weakly cuneate uniserial, pinnulate.
Stem obtusely pentagonal with rounded margins; lumen large, diameter ~60% stem diameter. Proximal to medial stem heteromorphic, N212; nudinodals moderately thick with smooth, rounded latera, priminternodals slightly thinner and with smaller diameter than the first; internodals very thin, not visible in side view, entirely overlapped by nudinodals, comprised of five narrow petal-like arcuate segments that are only marginally in lateral contact.
Distal column, holdfast, distal arms, and tegmen unknown.
Etymology
The specific name lepidotus is derived from the Greek lepidotós, meaning scaly, in reference to the scale-like appearance of the calyx plates.
Materials
CMCIP 76789, holotype (middle–upper Bobcaygeon Formation, Kirkfield Quarry); CMCIP 76790, paratype (middle–upper Bobcaygeon Formation, Kirkfield Quarry); UMMP 74681.2, paratype (Bobcaygeon-Verulam contact zone, LaFarge Quarry).
Remarks
Cleiocrinus lepidotus n. sp. is most similar to C. perforatus but differs from this and all other species of Cleiocrinus on the basis of its plate ornamentation, the number and arrangement of sutural pores, and the condition of the posterior interray. Whereas C. lepidotus n. sp. is ornamented with fine pitting, other Cleiocrinus species are unornamented (C. perforatus) or have a variety of ridges arranged in radiating, concentric, and/or rhombic patterns (all other species). Among species of Cleiocrinus, sutural pore number and arrangement ranges from several large pores along each plate margin (C. perforatus) to small elongated slits (C. laevis) to many small, closely spaced circular pores (all other species). In contrast, C. lepidotus n. sp. has few, relatively large sutural pores (typically two per plate margin) that are positioned to either side of each plate corner. In addition, in many species of Cleiocrinus, the external plate sutures converge into larger pores before reaching the inner surface of the plate (e.g., C. regius, C. bromidensis). The large pores of C. lepidotus n. sp. appear to penetrate the calyx directly and do not converge, a character apparently shared with C. perforatus. Finally, all species of Cleiocrinus for which the posterior is known have a posterior interray that is not in contact with the tegmen, whereas the posterior of C. lepidotus n. sp. is in contact with the tegmen.
Cleiocrinus sp.
Materials
UMMP 74678.3 (Bobcaygeon-Verulam contact zone, LaFarge Quarry).
Remarks
Cleiocrinus holdfasts are morphologically distinctive and can be assigned to the genus with confidence. The holdfast structure is a high inverted cone comprised of small, intergrown plates that encrust the underlying surface. The center of the cone is deeply depressed, and the distal stem rests within the depression. It is probable that the specimen examined here belongs to C. regius, but a specific assignment cannot be made with certainty.
Family Rhodocrinitidae Roemer, Reference Roemer1855
Remarks
Recent phylogenetic analyses of Ordovician camerates (Cole, Reference Cole2017) reinforce the long-held suspicion that Rhodocrinitidae is a polyphyletic group. The family is in need of substantial systematic revision, but such an undertaking is beyond the scope of this manuscript because it requires a comprehensive phylogenetic analysis of Diplobathrida. Despite this issue, the name Rhodocrinitidae is maintained herein because there is not yet an alternative classification that reflects the evolutionary relationships of diplobathrids at the family level. It is possible that some of the taxa assigned herein to Rhodocrinitidae will require reassignment to a different family once a revised, phylogenetically informed classification is completed.
Genus Archaeocrinus Wachsmuth and Springer, Reference Wachsmuth and Springer1881
Type species
Archaeocrinus lacunosus (Billings, Reference Billings1857).
Diagnosis
Rhodocrinitid with a medium to high conical or globose calyx; infrabasals small, restricted to basal concavity; basals large; regular interrays in contact with tegmen, proximal plating 1-2; CD interray wider, proximal plating 1-3, forming an anal series; arms 10, uniserial or biserial, branching isotomously.
Occurrence
Middle–Upper Ordovician (Darriwilian–Katian); United States (Oklahoma, New York, Kentucky, Virginia, Tennessee) and Canada (Ontario, Quebec).
Remarks
Springer (Reference Springer1911) reported both A. microbasalis and A. lacunosus from the region of Kirkfield. However, all specimens examined from the Brechin region differ significantly from previously described species. This suggests previous specific identifications of Archaeocrinus from Kirkfield may have been in error.
Archaeocrinus sundayae new species

Figure 7 Archaeocrinus sundayae n. sp. from the Bobcaygeon-Verulam contact zone, Carden Quarry, southern Ontario. (1, 4, 7, 8) UMMP 74682, holotype; (1) CD interray view; (4) E ray view; (7) adoral view of calyx base, CD interray oriented down (8) aboral view of tegmen and anal tube, CD interray oriented down; (2, 3, 5) UMMP 74683, paratype; (2) close-up of crown; (3) lateral view of complete specimen; (5) close-up of holdfast; (8) juvenile specimen UMMP 74684, paratype, lateral view of complete crown and proximal stem. All scale bars 5 mm.
Holotype
UMMP 74682, holotype; UMMP 74683, paratype; UMMP 74684, paratype.
Diagnosis
Archaeocrinus with lobate globose to bowl-shaped calyx; weakly defined median ray ridges; lack of plate ornamentation; fixed pinnules numerous, preventing contact of interrays with tegmen; regular interrays with few (<10) plates; posterior interray with additional plates and weak anitaxial ridge; fixed brachials bifurcating once within the calyx, giving rise to 10 arm openings; arms biserial, pinnulate, poorly isotomously branched.
Occurrence
Material described and figured here is from the Upper Ordovician (lower Katian) Bobcaygeon-Verulam contact zone, Carden Quarry. Two additional specimens (CMCIP 54029) examined are referred to A. sundayae n. sp., which are from the Upper Ordovician (Katian) Curdsville Formation, Garrard Co., Kentucky, on highway KY 34, just east of the bridge crossing Herrington Lake.
Description
Calyx bowl-shaped to globose, slightly wider than high, lobate where arms become free; plates lacking ornamentation; plate sutures clearly visible, flush with plate surface; base of calyx concave without any surrounding ornamentation; median ray ridges faintly convex.
Infrabasals not observed, missing (UMMP 74682) or completely hidden by column (UMMP 74683). Basal circlet truncate proximally, forming rim of basal concavity; basal plates five, hexagonal, almost entirely visible in side view, slightly wider than high.
Radial circlet interrupted in all rays by contact of basals and primanal and with first interradial plates; radial plates five, hexagonal in C and D rays, pentagonal in all other rays, slightly wider than high, equal in size to basal plates.
Regular interrays not in contact with tegmen, not depressed, interrupting the radial circlet in all rays. First interradial plate heptagonal, as wide as high, slightly larger than radials and basals. Second range with two plates; proximal plating 1-2-3-2-2 or 1-2-2-2 followed by higher rows of very small plates and fixed pinnules.
Posterior interray about as wide as regular interrays, not in contact with tegmen; primanal heptagonal, wider than high, smaller than first interray plates; proximal plating P-3-3-3-5-3-4, followed by higher rows of fixed pinnules; faint anitaxial ridge along medial column of plates.
First primibrachial fixed, hexagonal, wider than high; second primibrachial axillary, pentagonal. Secundibrachials fully fixed into calyx to about fifth secundibrachial, proximal brachials above fifth secundibrachial becoming biserial and forming lobate arm bases attached to the calyx by fixed pinnules; second, fourth and fifth secundibrachials giving rise to fixed pinnules on alternating sides of each half ray, higher secundibrachials all giving rise to fixed pinnules grading into free pinnules in higher brachials. Fixed intrabrachials between secundibrachials of each half ray comprised of a single plate or followed by up to two rows of one or two plates each; higher rows composed of fixed pinnules preventing contact with tegmen.
Tegmen low, comprised of numerous small plates; tegmen plates undifferentiated, ornamented with small nodes. Plates covering ambulacral areas convex, showing branching pattern on tegmen; interambulacral areas depressed, interambulacral area opposite the posterior interray with a deep depression. Anal tube cylindrical, positioned subcentrally toward the posterior interray; remnants of small spines projecting from some anal tube plates, presumably originally present on all anal tube plates; total height of anal tube unknown; anal opening presumably from anal tube terminus.
Arm openings 10, ungrouped; numerous fixed pinnules positioned between all arm openings. Free arms poorly isotomously branched, bifurcating twice; first branching typically on the 13th secundibrachial, second branching irregular in height. Brachials wedge biserial throughout free arms. Pinnules on every brachial except axillary arm plates; fixed pinnules stout, robust; distal pinnules longer and thinner.
Stem circular, holomeric; heteromorphic in proximal and medial stem, N3231323, becoming homeomorphic in distal stem. Holdfast composed of ~15 terminal rhizoids, clustered on one side of column. Lumen unknown.
Etymology
The species name sundayae is for Steffanie Sunday, wife of J.M. Koniecki.
Materials
UMMP 74682, holotype; 74683, paratype (Bobcaygeon-Verulam contact zone, Carden Quarry); and 74684, paratype (Bobcaygeon-Verulam contact zone, Carden Quarry); CMCIP 54029 (two specimens, Curdsville Formation, Garrard Co., Kentucky).
Remarks
Of the 11 species of Archaeocrinus previously described, several are known from Ontario, including A. marginatus, A. lacunosus, and A. microbasalis. Archaeocrinus sundayae n. sp. is differentiated from other species of Archaeocrinus on the basis of its numerous fixed pinnules, very narrow contact between the interrays and the tegmen, relatively few interray plates (≤10 large plates) in four or five rows, lack of plate ornamentation, faint median ray ridges, and up to two bifurcations in free arms. In contrast, A. lacunosus has no fixed pinnules, interrays in contact with the tegmen, numerous interray plates (>20 large plates), and irregular stellate ridges; A. microbasalis has few fixed pinnules, interrays in contact with the tegmen, numerous interray plates in 10 or more rows, striated stellate ornamentation, distinct median ray ridges, and three bifurcations in free arms. Although A. marginatus is poorly known, it differs from A. sundayae n. sp. in that the interrays are comprised of many large plates (>20) in at least seven rows. Archaeocrinus sundayae n. sp. is also very similar to A. snyderi Guensburg, Reference Guensburg1984, but the latter differs in that it lacks lobate arm bases, has interrays in contact with the tegmen, has no anitaxis in the posterior interray, lacks tertinternodals in the stem, and possesses uniserial arms that bifurcate at least three times.
Specimen UMMP 74684 is likely a juvenile form. Although the specimen is much smaller than other specimens of A. sundayae n. sp. examined, the presumed juvenile specimen is remarkably similar to adults in terms of interray plating, overall shape, organization of the calyx, and arm construction. The arms of the juvenile specimen display fully developed biserial brachials, which often do not develop until late in ontogeny. This suggests that this species developed adult plating early in ontogeny and then proceeded to grow isometrically through enlargement of plates, rather than continued insertion of plates and/or changes in plate dimensions.
Archaeocrinus maraensis new species

Figure 8 Archaeocrinus maraensis n. sp. from southern Ontario. (1, 3) UMMP 74685, holotype (lower Verulam Fm., James Dick Quarry); (1) close-up lateral view of crown; (3) view of crown and proximal to medial stem; (2, 4, 5) UMMP 74686.1, paratype, juvenile specimen (Bobcaygeon-Verulam contact zone, Carden Quarry); (2) holdfast; (4) close-up lateral view of crown; (5) lateral view of crown, stem, and holdfast; (6, 7) UMMP 74688, paratype (lower Verulam Fm., James Dick Quarry); (6) close-up of calyx showing calyx plate ornamentation; (7) lateral view of crown. All scale bars 5 mm.
Holotype
UMMP 74685, holotype; UMMP 74686.1, paratype; UMMP 74688, paratype.
Diagnosis
Archaeocrinus with medium globose calyx; plates with delicate, irregular granular and stellate ornamentation; numerous fixed pinnules; interrays not in contact with tegmen; free arm openings 10; free arms bifurcating isotomously three times; brachials flat chisel biserial; stem strongly heteromorphic.
Occurrence
Material described and figured here is from the Upper Ordovician (lower Katian) Bobcaygeon-Verulam contact zone, Carden Quarry; and from the lower Verulam Formation, James Dick Quarry.
Description
Calyx medium bowl-shaped to globose, about as wide as high, arm bases protruding slightly; plates with fine, irregular granular ornamentation and stellate ridges; plate sutures clearly visible, flush with plate surface; base of calyx concave without any surrounding ornamentation; median ray ridges faint, narrow, beginning in center of radial plates.
Infrabasals not observed, entirely in basal concavity. Basal circlet curving slightly at base of calyx but entirely visible in side view, not part of basal concavity; basal plates five, hexagonal, wider than high.
Radial circlet interrupted in all rays by contact of basals and primanal with first interradial plates; radial plates five, pentagonal, slightly wider than high, equal in size to basal plates.
Regular interrays not in contact with tegmen, not depressed, interrupting the radial circlet in all rays. First interradial plate heptagonal, as wide as high, similar in size to radials and basals. Second range with two plates; typically 10 or more rows of plates above first interray plate; proximal plating 1-2-3-3-3-3 or 1-2-3-3-2-2 followed by higher rows of three to five small plates each.
First primibrachial fixed, hexagonal, wider than high; second primibrachial axillary, pentagonal or heptagonal. Secundibrachials fully fixed into calyx to about fifth secundibrachial, proximal brachials above becoming biserial and forming protruding arm bases attached to the calyx by fixed pinnules; second, fourth, and fifth secundibrachials giving rise to fixed pinnules on alternating sides of each half ray. Fixed intrabrachials between secundibrachials of each half ray, proximal plating usually 1-2-2-2; higher rows comprised of fixed pinnules preventing contact with tegmen.
Arm openings 10, ungrouped; fixed pinnules positioned between all arm openings. Free arms isotomously branched, bifurcating three times; first branching typically around the 15th secundibrachial, second branching occurring shortly above the first; third branching somewhat variable in height. Brachials flat chisel biserial throughout free arms. Pinnules on every free brachial, densely spaced.
Stem circular, holomeric; strongly heteromorphic, diameter of largest nodals more than twice that of smallest intervening internodals; proximal stem N212, medial stem N3231323, distal stem becoming homeomorphic. Lumen pentalobate, ~18% of stem diameter; crenularium present around periphery of columnal facet, but details not well preserved. Holdfast comprised of numerous terminal rhizoids at the end of the column.
Tegmen, anal opening, and additional stem features unknown.
Etymology
The species name maraensis is in reference to the Mara Township where the holotype specimen was recovered.
Materials
UMMP 74685, holotype (lower Verulam, James Dick Quarry); UMMP 74688, paratype (lower Verulam, James Dick Quarry); UMMP 74686.1, juvenile specimen, paratype (Bobcaygeon-Verulam contact zone, Carden Quarry).
Remarks
Archaeocrinus maraensis n. sp. is most similar to A. sundayae n. sp., A. microbasalis, and A. snyderi. All four species have fixed pinnules and branched arms, but they differ with regards to the number of fixed pinnules, the condition of the interrays, and features of the free arms. In particular, A. maraensis n. sp. has free arms that bifurcate at least three times, numerous fixed pinnules, interrays not in contact with the tegmen, flat chisel biserial arms, and fine granulose and stellate ornamentation. Although A. microbasalis has arms that branch three times and flat chisel biserial brachials, it differs from A. maraensis n. sp. in that it has few fixed pinnules, interrays in contact with the tegmen, and stellate ornamentation. Similarly, A. snyderi has fixed pinnules and arms that bifurcate three times, but it has interrays that contact the tegmen, uniserial arms, and lacks plate ornamentation. Finally, although A. sundayae n. sp. has numerous fixed pinnules and interrays that do not contact the tegmen, it differs from A. maraensis n. sp. in that it has arms that bifurcate only twice, wedge chisel biserial arms, and lacks plate ornamentation.
UMMP 74686.1 is probably a juvenile specimen of A. maraensis n. sp.; it shows little morphological differentiation from adult specimens of the species other than its much smaller size compared to other specimens of A. maraensis n. sp. UMMP 74686.1 clearly belongs to A. maraensis n. sp. rather than A. sundayae n. sp. because it has similar stem morphology and irregular stellate ridges and granular ornamentation. Similar to A. sundayae n. sp., however, the juvenile specimen of A. maraensis n. sp. has the same proportions as adult specimens, indicating isometric growth may be common to the genus.
Acknowledgments
We thank B. Hunda (Cincinnati Museum Center) and D. Miller (University of Michigan Museum of Paleontology) for access to museum specimens and assistance with specimen curation. K. Brett is thanked for donating UMMP 74688 for study and providing comments on an early draft of this manuscript. We thank G. Sevastopulo and B. Deline for providing thoughtful reviews and C. Sumrall for helpful comments. SRC was supported by a J. Thomas Dutro Jr. Student Award from the Paleontological Research Institute, a Presidential Fellowship from The Ohio State University, and a Springer Fellowship from the NMNH (Smithsonian Institution). DFW was supported by a James R. Welch Scholarship from the Association of Applied Paleontological Sciences and a Peter Buck Fellowship from the NMNH (Smithsonian Institution).
Supplementary material
To view supplementary material for this article, please visit the Dryad Digital Repository: http://doi.org/10.5061/dryad.dd0b1








