Introduction
Farming systems based on no-till can improve soil health and functioning by increasing organic matter, enhancing microbial activity, and improving soil structure (Hobbs, Reference Hobbs2007; Triplett and Dick, Reference Triplett and Dick2008). To encourage adoption of no-till globally, the Food and Agriculture Organization (FAO) of the United Nations developed the concept, conservation agriculture (FAO, 2015). Conservation agriculture is based on three principles: (1) direct seeding of crops with minimum soil disturbance (no-till), (2) permanent soil cover by crop residues or cover crops and (3) diversity in crop rotation. The FAO views conservation agriculture as critical for achieving sustainability of global agriculture.
Organic producers are interested in no-till to improve soil health (Peigne et al., Reference Peigne, Ball, Roger, Strand and David2007; Maader and Bemer, Reference Maader and Bemer2012), but they are concerned about managing weeds without tillage (Sooby et al., Reference Sooby, Landeck and Lipson2007; Brainard et al., Reference Brainard, Haramoto, Williams and Mirsky2013). One approach that may reduce need for tillage to control weeds is to design rotations comprising crops with a diversity of life cycles (Anderson, Reference Anderson2005; Cavigelli et al., Reference Cavigelli, Teasdale and Conklin2008). For example, rotating warm-season crops such as corn (Zea mays L.) with cool-season crops such as winter wheat (Triticum aestivum L.) provides more opportunities for producers to prevent weeds from completing their life cycle and producing weed seeds. Arranging annual crops in 2-yr seasonal intervals, i.e., corn–soybean for a warm season sequence, is especially effective in suppressing annual weed dynamics. In conventional no-till systems, producers using a rotation with two cool-season crops followed by two warm-season crops eliminated the need for herbicides in three crops out of four (Anderson, Reference Anderson2005).
To further increase rotation complexity, perennial legumes such as alfalfa (Medicago sativa L.) or red clover (Trifolium pratense L.) can be added to the rotation. Population dynamics of weeds are disrupted in perennial legumes due to mowing for forage harvest and crop competition; it is difficult for annual weeds to establish and produce seeds (Ominski et al., Reference Ominski, Entz and Kenkel1999; Entz et al., Reference Entz, Baron, Carr, Meyer, Smith and McCaughey2002). Also, weed seeds remain on the soil surface where they are less likely to survive (Anderson, Reference Anderson2005). A perennial legume would help weed management while providing a 3-yr interval of no-till, and could be the starting point for a no-till, organic rotation.
We are seeking to develop a continuous no-till system for organic farming, and our research is guided by a complex rotation consisting of 3 yr of alfalfa followed by a 6-yr sequence of corn–soybean [Glycine max (L.) Merr.]–winter wheat–oat (Avena sativa L.)–soybean–corn (Anderson Reference Anderson2010). Alfalfa is grown for only 3 yr to avoid proliferation of weeds adapted to perennial legumes. Weed density in alfalfa usually start increasing in the fourth and fifth year of alfalfa (Anderson, Reference Anderson2015a) because alfalfa density declines across time (Undersander et al., Reference Undersander, Cosgrove, Cullen, Grau, Rice, Renz, Sheaffer, Shewmaker and Sulc2011).
We believe alfalfa will enhance success of no-till in organic farming, but it will be crucial to develop a uniform stand of alfalfa to effectively suppress weeds. In our proposed rotation, alfalfa is planted after corn. However, corn residue on the soil surface may interfere with seed placement. In an earlier study, density of red clover, which has similar seed size as alfalfa, was 15–40% lower following corn compared with spring wheat as preceding crops (Anderson, Reference Anderson2015b). Sowing alfalfa in the fall, following spring wheat harvest, is another option. Yet, weeds present at spring wheat harvest and wheat volunteers may interfere with legume establishment; organic producers till after spring wheat harvest to eliminate these weeds before sowing alfalfa.
Therefore, the objective of this study was to evaluate impact of preceding crop on alfalfa establishment, production and competition with weeds in a no-till, organic system. Weed community data were collected in the third forage year, preceding conversion to annual crops in the following year.
Materials and Methods
Study procedures
The study was established on a Barnes clay loam (Calcic Hapludoll) soil near Brookings, SD. The soil contained approximately 4% organic matter and had a soil pH of 6.9. Average yearly precipitation (30-yr record) is 584 mm. The cropping history of the site prior to the study was a corn–soybean–spring wheat rotation.
The study involved a 4-yr interval. In the first year, spring wheat, corn and soybean were grown with conventional organic practices, where tillage was used to prepare a seedbed for all crops and control weeds during the early growing season in corn and soybean. Spring wheat (‘Briggs’) was planted at 130 kg ha−1, corn (DKC 47–10) at 61,000 seeds ha−1 and soybean (SD Genetics 2121) at 340,000 seeds ha−1. The seedbed was tilled before planting and weeds in corn and soybean were controlled with in-crop tillage. Spring wheat was planted in April and harvested in late July; corn and soybean were planted in May and harvested in October.
Alfalfa (Pioneer 54V54) was planted with a no-till drill following spring wheat harvest in the first year, and in the spring of the year following corn and soybean production (Table 1). Seeding rate was 14 kg ha−1 and seeding depth was 1 cm. Oat (‘Stallion’) at 40 kg ha−1 was planted as a companion crop with alfalfa in the corn and soybean treatments. Oat was planted first at 3 cm depth, followed by alfalfa planting on the same day. The drill was equipped with single disk openers. There was no tillage for seedbed preparation or weed control before planting alfalfa. Treatments were arranged in a randomized complete block design with six replications; plot size was 7×15 m2. The study was conducted twice, during 2011–2014 and 2012–2015.
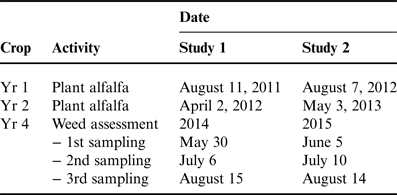
Table 1. Time of cultural activities for establishing alfalfa following three annual crops and for assessing weed infestations.
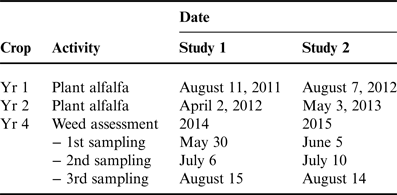
Forage yield of alfalfa was recorded in years 2, 3 and 4 (referred to as forage years 1, 2 and 3) when alfalfa reached the 1/10 bloom stage. Forage samples were hand harvested from an area 2×2 m2, with two subsamples randomly placed in each plot. Samples were oven-dried at 65°C until samples reached a constant weight. The remainder of the plot area was harvested with conventional equipment. In the first forage year, alfalfa was harvested twice when planted after spring wheat, but once when planted after soybean and corn. Alfalfa was harvested three times with all treatments in the second and third forage years in the late May, early July and mid-August. Alfalfa density was estimated by recording the number of 2.5 cm spaces in 1 m of row occupied by a plant stem, thus providing a non-destructive assessment of stand density. Density measurements were made at 8 random locations in each plot in early August of the first forage year, and early June in the second and third forage years.
In the third year, weed community variables also were measured in the 2×2 m2 forage quadrats at the three sampling dates (Table 1). Identification and density of specific weeds was recorded, and then weed community biomass was collected. Biomass data were expressed as dry weight; samples were oven-dried to a constant weight at 65°C.
Statistical analysis
Data were analyzed as a randomized complete block design. Data were initially examined for homogeneity of variance among years, and then subjected to analysis of variance to determine treatment effects and possible interactions among treatments and years. Main and interaction effects were considered significant at P ≤ 0.05; treatment means were separated with Fisher's Protected Least Significant Difference (LSD: 0.05).
Alfalfa forage yield, weed community density and weed biomass data were averaged across quadrats within a plot, and then analyzed with a repeated measures procedure. If interactions were found between the treatment and the date of sampling, data were analyzed within sampling date. The Fisher's Protected LSD was used for mean separation.
Results and Discussion
With all data, an interaction was not observed between treatments and studies; therefore, data were averaged across studies. Also, there was not an interaction between repeated sampling and treatments for weed community data, thus, data were averaged across sampling dates. An interaction was observed between alfalfa forage yield and year, and data were shown for individual years.
Biomass of alfalfa and weeds in the third forage year
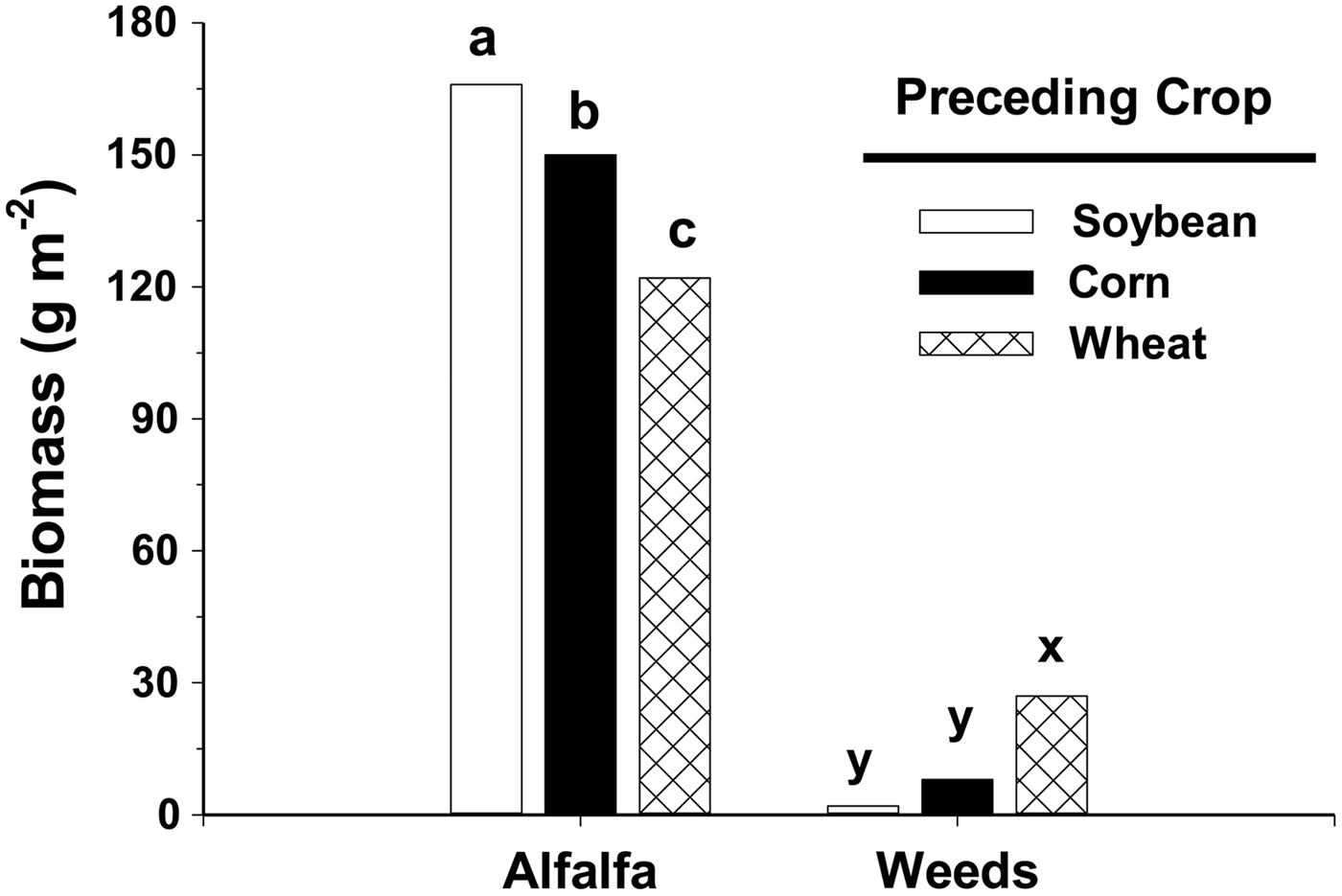
Weed biomass in alfalfa following spring wheat was 27 g m−2, more than five times higher than biomass in alfalfa following corn or soybean (Fig. 1). Only 2 g m−2 of weeds were present when alfalfa followed soybean. Weed biomass in alfalfa following spring wheat comprised 18% of the plant community, but only 1% of the plant community when alfalfa followed soybean. Weed biomass in alfalfa did not differ between corn and soybean as preceding crops.
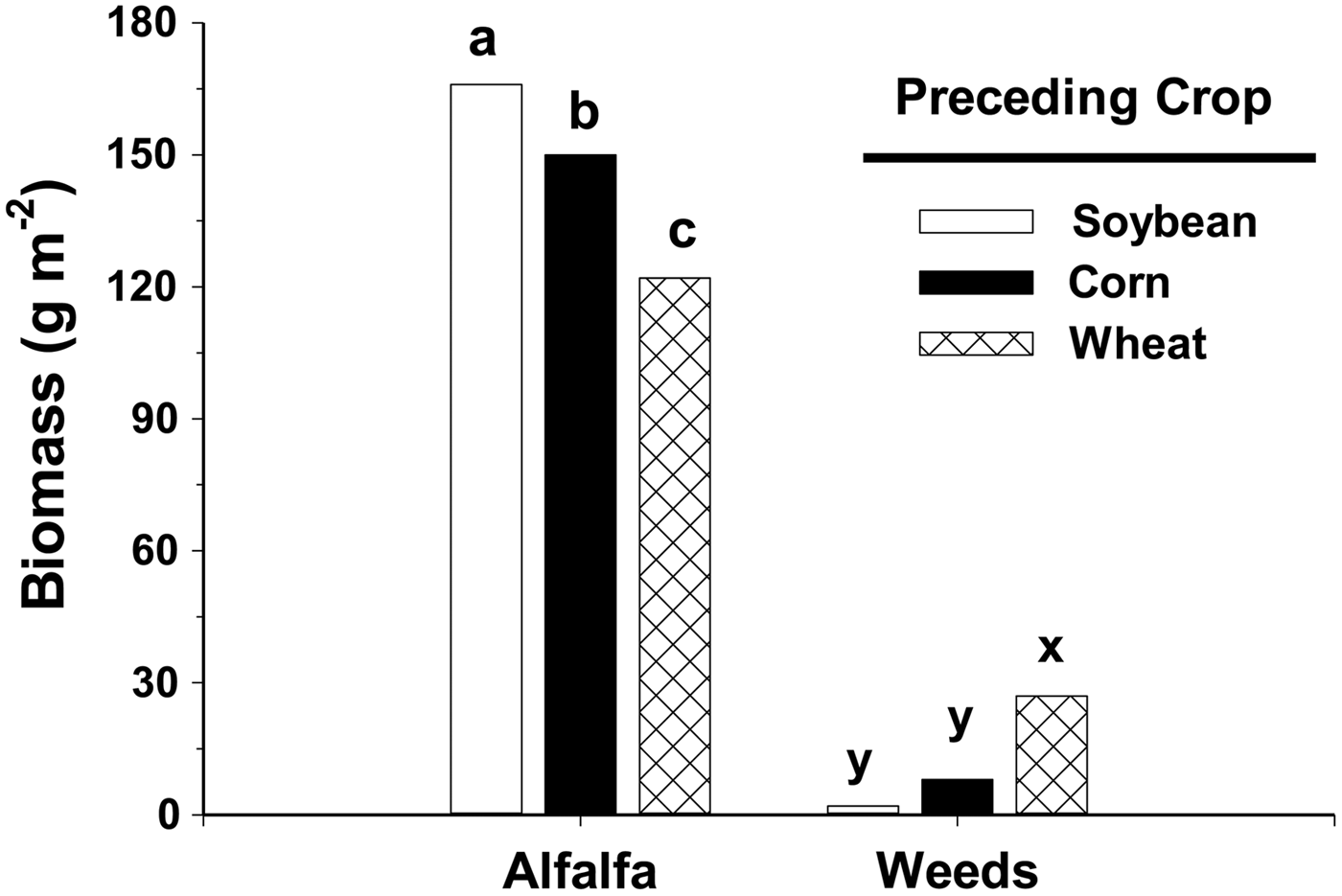
Figure 1. Biomass (dry weight) of alfalfa and weeds as affected by the preceding crops, soybean, corn or spring wheat. Data collected in the third forage year and averaged across three sampling dates and two studies. Bars within either alfalfa or weeds with the same letter are not significantly different as determined by Fisher's LSD (0.05).
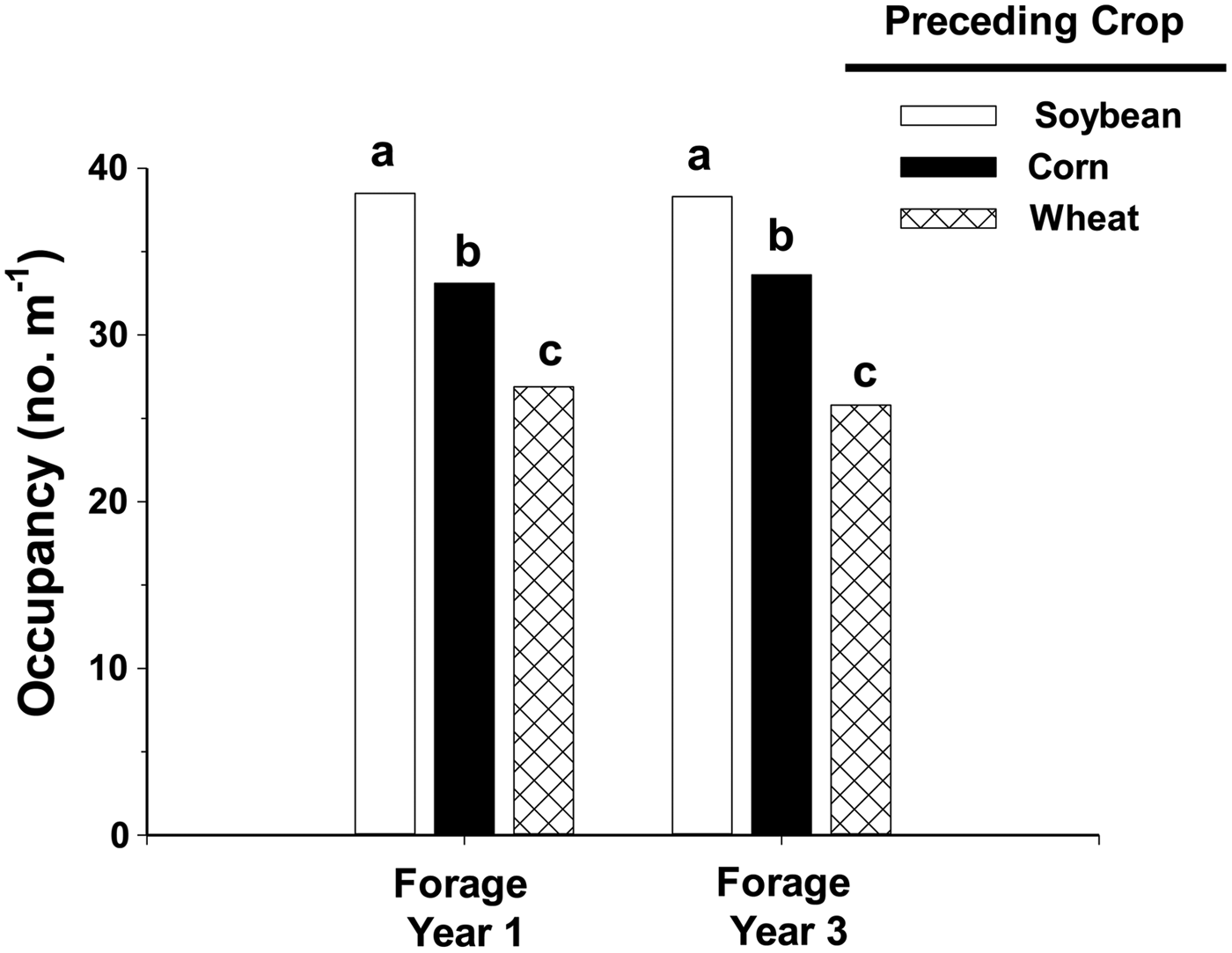
Weeds produced more biomass in alfalfa following spring wheat because of a lower stand density. Alfalfa following spring wheat occupied only 26 spaces out of a possible 40 in the third forage year, in contrast with alfalfa occupying more than 38 spaces when following soybean (Fig. 2). Difference in alfalfa density among treatments was observed in both forage years 1 and 3, which indicates that initial stand establishment varied with preceding crop. Alfalfa establishment following spring wheat harvest was reduced due to competition from spring wheat volunteers and fall growth of annual weeds. Few volunteers of soybean and corn established in alfalfa and seedlings of warm-season weeds that infested soybean and corn the previous year did not emerge until after alfalfa seedlings were established. Similar reduction of alfalfa density due to weeds has been shown elsewhere; Wilson and Burgener (Reference Wilson and Burgener2009) and Bradley et al. (Reference Bradley, Kallenbach and Roberts2010) found that weed interference can reduce alfalfa density 20–30%.
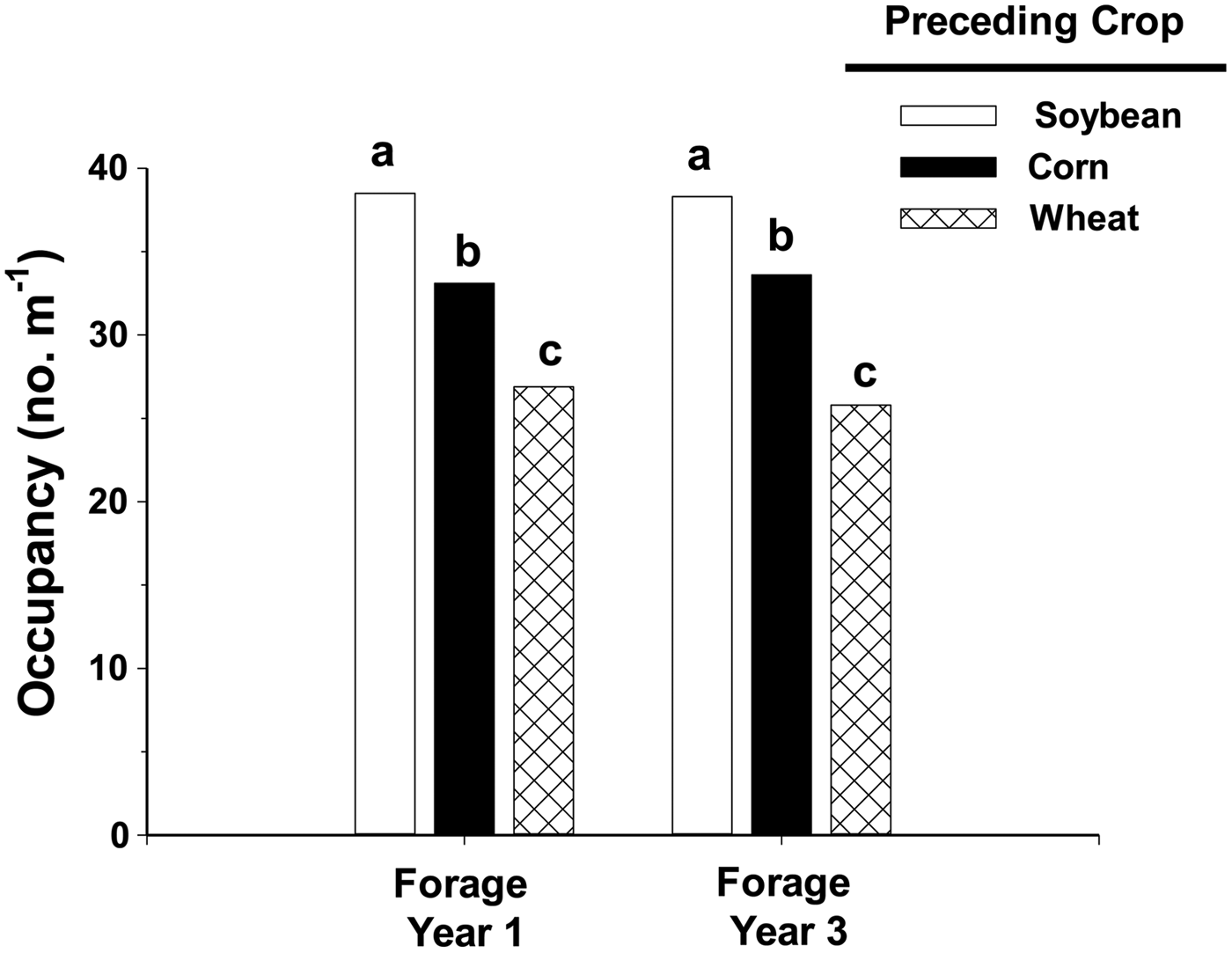
Figure 2. Stand estimates (occupancy) of alfalfa in the first and third forage years when established after spring wheat, corn or soybean. Maximum possible value is 40 spaces m−1 of row. Bars within a forage year with the same letter are not significantly different as determined by Fisher's LSD (0.05).
Alfalfa density, as assessed by occupancy, was less following corn than following soybean (Fig. 2). We attribute this to corn residues on the soil surface interfering with proper seed placement during alfalfa planting, as previous research showed that stand density of red clover was reduced by corn stalks in a no-till system (Anderson, Reference Anderson2015b).
Alfalfa also produced the highest forage quantity following soybean in the third forage year (Fig. 1). Alfalfa yielded 166 g m−2 following soybean but only 122 g m−2 following spring wheat, a reduction of 27%. Alfalfa following corn also yielded 10% less than alfalfa following soybean. These yield differences reflect differences in stand establishment, as both highest yield (Fig. 1) and plant density, as noted by occupancy values (Fig. 2), occurred when alfalfa followed soybean. The lower alfalfa density also led to more weed interference, which decreased forage yield. Weed interference can reduce alfalfa yield 15–30% (Wilson and Burgener, Reference Wilson and Burgener2009; Dillehay and Curran, Reference Dillehay and Curran2010).
Weed species density in alfalfa as affected by preceding crop
The weed community consisted primarily of downy brome, dandelion, wild buckwheat (Polygonum convolvulus L.), a mix of green [Setaria viridis (L.) Beauv.] and yellow foxtail [Setaria pumila (Poir.) Roem. & Schult.] (referred to as a foxtail complex), and common lambsquarters (Chenopodium album L.). Density of downy brome and dandelion varied among treatments; highest density of both species occurred when alfalfa followed spring wheat (Table 2). For example, 10.7 downy brome plants m−2 infested alfalfa following spring wheat, whereas downy brome was not observed in alfalfa following soybean. Also, dandelion density was 13-fold higher in alfalfa following spring wheat than following soybean, 5.3 versus 0.4 plants m−2, respectively. Gaps in the canopy due to interference by weeds and volunteer wheat during alfalfa establishment allowed downy brome and dandelion to establish and increase in population in alfalfa following spring wheat. Downy brome and dandelion were observed in alfalfa following corn also, but at a lower density than following spring wheat (Table 2). These species, downy brome and dandelion, are prominent weeds in alfalfa throughout the Northern Great Plains (Ominski et al., Reference Ominski, Entz and Kenkel1999; Moyer and Acharya, Reference Moyer and Acharya2006).
Table 2. Weed density in alfalfa as affected by preceding crop. Data averaged across three samplings in the third forage year and across two studies. Total density is the sum of all weed species observed. Means within a column followed by identical letters are not significantly different based on Fisher's LSD (0.05).

Density of the other species, wild buckwheat, foxtail complex and common lambsquarters, did not vary among treatments. Weed community density was 24.4 plants m−2 when alfalfa followed spring wheat or 4-fold higher than 6.1 plants m−2 observed in alfalfa following soybean (Table 2). Even though densities of individual weed species did not vary between alfalfa following corn or soybean, weed community density was 14.1 plants m−2 when alfalfa followed corn, 2-fold higher than weed density in alfalfa following soybean. Alfalfa density was 13% lower when following corn compared with soybean (based on occupancy values, Fig. 2); consequently, alfalfa following corn was less competitive with weeds.
Alfalfa forage yields
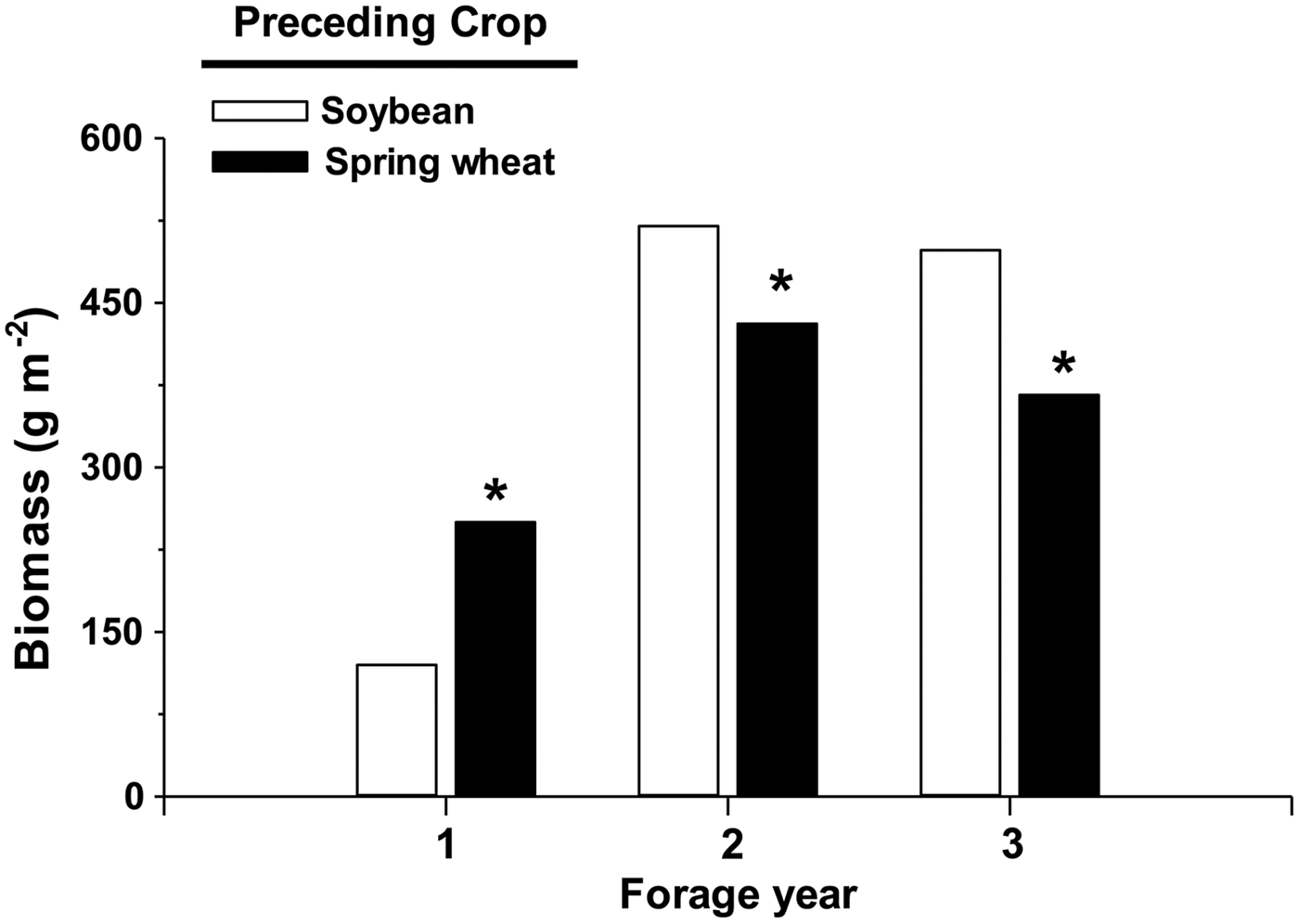
We were concerned that differences in number of harvests in the first forage yield (two harvest times for alfalfa following spring wheat compared with one harvest for alfalfa following soybean or corn), may lead to less forage when alfalfa was established after soybean or corn. But, alfalfa forage yield summed across 3 yrs was 8% higher following soybean than spring wheat (1140 versus 1050 g m−2, respectively; P = 0.047). Higher alfalfa yields in forage years 2 and 3 when alfalfa followed soybean compensated for higher yield of alfalfa following spring wheat in forage year 1 (Fig. 3). Alfalfa also yielded 8% less following corn than following soybean across 3 yr (data not shown), which is attributed to lower stand density of alfalfa following corn (Fig. 2).

Figure 3. Biomass (dry weight) of alfalfa following spring wheat or soybean, summed across samplings within a year. Asterisk above a bar within a forage year signifies that means within a forage year differed significantly at the 0.05 level of probability.
Summary
Alfalfa established after soybean is more suppressive of weeds than alfalfa following spring wheat or corn in a no-till organic system. In the third forage year, weed biomass in alfalfa following spring wheat was 18% of the plant community, but only 1% when alfalfa followed soybean (Fig. 1). Alfalfa yield was also higher following soybean than spring wheat.
Weed biomass increased because alfalfa density following spring wheat was only 71% of alfalfa following soybean (Fig. 2). Two species, downy brome and dandelion, were especially prominent in alfalfa following spring wheat because of a sparse stand, but these species were rarely observed when alfalfa followed soybean. Based on this study, we suggest organic producers establish perennial legumes after soybean to improve competitiveness with weeds.
Our research program is approaching success with a continuous no-till system. We have found that red clover (medium type) can be converted to annual crops with a mowing strategy, thus eliminating the need for tillage (Anderson, Reference Anderson2015c). Therefore, a perennial legume can be included in a no-till organic rotation to help manage weeds. Also, we have shown that weeds can be controlled following small grain harvest with cover crops or underseeded annual clovers that winter kill, thus eliminating the need for tillage during a 2-yr cool-season interval (Anderson, Reference Anderson2015a, Reference Anderson2015d). Thus, weeds can be controlled successfully without tillage in at least 6 yr of our proposed 9-yr rotation (Anderson, Reference Anderson2010).
Organic producers are seeking to restore and protect soil health (Sooby et al., Reference Sooby, Landeck and Lipson2007); a no-till system can contribute to that goal (Triplett and Dick, Reference Triplett and Dick2008). Incorporating a perennial legume into a complex rotation such as described above (Anderson, Reference Anderson2010, Reference Anderson2015a) will enhance the probability of successful no-till in organic farming. Furthermore, perennial legumes in the rotation can increase corn yield (Stanger and Lauer, Reference Stanger and Lauer2008), promote soil aggregation (Triplett and Dick, Reference Triplett and Dick2008) and improve nutrient cycling (Karlen et al., Reference Karlen, Hurley, Andrews, Cambardella, Meek, Duffy and Mallarino2006), which will lead to more sustainable cropping systems (Hobbs, Reference Hobbs2007; Krischenmann, Reference Kirschenmann2007).