
In recent years, the dissorophoid temnospondyls have assumed increasing importance in our attempts to understand amphibian evolution and the origins of the Lissamphibia. Most workers, including the author, now consider the dissorophoids to form the immediate lissamphibian stem, though this view is not yet universal. In the former view, the dissorophoids form a discrete adaptive radiation, first appearing in the Middle Pennsylvanian and last appearing in the Early Triassic, at which time the descendant Lissamphibia formed a second adaptive radiation which persists to the present day.
The component families Micromelerpetontidae, Amphibamidae, Branchiosauridae, Trematopidae and Dissorophidae are best known morphologically from well-preserved material from the Early Permian of North America and Europe. Consequently they are defined from somewhat derived forms. Their Pennsylvanian precursors, which might be expected to represent the basal conditions of the families, are not yet so well described, being mostly represented by small, flattened individuals. Four of the five families make their first appearances in the Asturian (late Middle Pennsylvanian) localities of Linton, Ohio; Mazon Creek, Illinois; and Nýřany in the Czech Republic. From Linton have been described the amphibamid Platyrhinops and the uncertainly placed Stegops; from Mazon Creek, the amphibamid Amphibamus; and from Nýřany, the micromelerpetontid Limnogyrinus, the amphibamid Playrhinops, the branchiosaurid Branchiosaurus, the trematopid Mordex and the uncertainly placed Nyranerpeton. The Dissorophidae make their first certain appearance slightly later in the Upper Pennsylvanian of Ohio and Oklahoma, USA (Vaughn Reference Vaughn1971; May et al. Reference May, Huttenlocker, Pardo, Benca and Small2011). An obvious conclusion is that the dissorophoids have a significant earlier history, not yet discovered, and must have been diversifying throughout the Pennsylvanian at least. The purpose of this study is to re-evaluate the earliest known trematopid material from Nýřany in the light of other recent descriptions of later trematopids.
The Nýřany Member of the Pilsen Basin represents an alluvial plain with many braided stream systems giving rise to extensive fluvial deposits (Opluštil et al. Reference Opluštil, Martínek and Tasáryova2005). Among these are many channel fills and it is likely to be that the Main Nýřany Coal (Gaskohle of Fritsch) was laid down in a small lake formed in an abandoned channel. It may have been an ox-bow lake but this is not likely to be provable now. The Main Nýřany Coal is a remarkable konservat-lagerstatte, still significantly understudied, despite being known since 1870. It includes a range of arthropods, fish and tetrapods, at least 800 specimens of the latter being present in collections across Europe. Some are represented by many tens of specimens and can form the basis of population and ontogenetic studies, but there are also many ‘accidental drop-ins' represented by one to a few individuals. Our understanding of the dissorophoid component of the fauna is complicated by the presence of many small specimens that appear to be juveniles and larvae, and it appears that the Nýřany water-body served as a breeding pool for a range of tetrapod taxa. This is particularly pertinent to the taxa being described here as it is probable, but not certain, that many of the small temnospondyls known from this assemblage may be larvae of the trematopids described here. This will be discussed in a later section of this work.
Institutional abbreviations. FMNH = Field Museum of Natural History, Chicago, USA; KUVP = Museum of Natural History, University of Kansas, Lawrence, Kansas, USA; MCZ = Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts, USA; NHMUK PV = Fossil Vertebrate Collection, Department of Earth Sciences, The Natural History Museum, London, UK; NMP = National Museum, Prague, Czech Republic; NMW = Naturhistorisches Museum, Wien (=Vienna), Austria; UT-BEG = Bureau of Economic Geology, University of Texas, Austin, Texas, USA.
1. Systematic history of the Nýřany trematopids
The amphibian Mordex calliprepes was described by Steen (Reference Steen1938, p. 260) on the basis of a single specimen without counterpart from the Asturian (=Westphalian D) ‘Plattelkohle' within the Main Nýřany Coal of Nýřany, now in the Czech Republic. Steen did not assign this form to any order or family but left it in a category of Amphibia incertae sedis, characterising it only by the presence of a small interfrontal ossification. Romer (Reference Romer1947, p. 167) suggested ‘with considerable doubt' that Mordex might be a predecessor to the Trematopsidae (hereafter referred to as the Trematopidae), a family of Early Permian temnospondyl amphibians. He noted that the configuration of the dermal bones of the skull roof was that of a primitive temnospondyl which had lost the intertemporal ossifications. The possible relationship to the Trematopidae was based on (1) the presence of large premaxillae each with a finger-like alary process overlapping the nasal, (2) the lacrimal broadly bordering the orbit margin, (3) the large tympanic embayment broadly bordered by the supratemporal and tabular on its dorso-medial edge. The alary process occurs in a similar state in many dissorophids although the premaxillae are less massive and, although its distribution is poorly known, it appears to occur in most post-edopoid temnospondyls (Milner Reference Milner, Taylor and Larwood1990). The lacrimal bordering the orbit occurs in colosteids, Dendrerpeton, trimerorhachoids and dissorophoids and is at least a primitive temnospondyl character. The large tympanic embayment occurs in dissorophids and amphibamids as well as trematopids. Of Romer's characters, only the enlarged premaxillae suggest that Mordex might be a trematopid. Gregory (Reference Gregory1950) suggested that Mordex was the European morphological counterpart of Platyrhinops (then referred to Amphibamus) from the contemporaneous locality of Linton, Ohio, and that both were primitive dissorophids. This view was elaborated further by Carroll (1964), who placed Mordex in synonymy with Amphibamus as A. calliprepes. Carroll argued that this species differed from the North American Amphibamus species only in some of the cranial proportions and showed no conclusive similarities to the Trematopidae. He noted that the interfrontal ossification reported by Steen might be an artefact of damage. In 1980, I followed this interpretation of the specimen, implying the reference to A. calliprepes of several undescribed amphibamid specimens from Nýřany (Milner Reference Milner and Panchen1980).
Subsequent re-examination of the holotype revealed that it does possess an elongate trematopid-like external naris and, while preparing a redescription of it as a trematopid, I was fortunate in locating, and being able to study, three undescribed larger specimens from Nýřany in the collections of the Naturhistorisches Museum in Vienna. These previously unrecognised specimens are also trematopid and initially I assumed that they might represent the adult of Mordex. After further work in Prague it also became clear the lectotype specimens of ‘Limnerpeton' laticeps Fritsch Reference Fritsch1881 and ‘Limnerpeton' macrolepis Fritsch Reference Fritsch1881, both from Nýřany, were trematopids rather than amphibamids. In our review of the genus Limnerpeton, Milner & Sequeira (Reference Milner and Sequeira2003) redescribed and refigured the types of ‘L'. laticeps and ‘L'. macrolepis fully and created the binomen Mordex laticeps for all the trematopid material from Nýřany. The type of laticeps is, unfortunately, a less informative juvenile specimen with fewer of the diagnostic characters of the taxon, but it does possess some, and the Law of Priority demanded that the combined taxon took the new combination Mordex laticeps (Fritsch) (Milner & Sequeira Reference Milner and Sequeira2003). I intended to describe the larger specimens as part of a wider study (Milner Reference Milner2007) but this was pre-empted by the publication by Werneburg (Reference Werneburg2012). However, the description of Fedexia striegeli (Berman et al. Reference Berman, Henrici, Brezinski and Kollar2010) and the publication of cladistic analyses of the Trematopidae (Berman et al. Reference Berman, Henrici, Brezinski and Kollar2010; Polley & Reisz Reference Polley and Reisz2011) led me to the conclusion that there are, in fact, two trematopid taxa occurring as accidentals in the Nýřany assemblage and that Mordex laticeps needed to be redivided into two distinct forms. This is the basis of this study. For the sake of completeness, the type of M. laticeps is briefly redescribed here, as are palatal elements of ‘L'. macrolepis, but the reader is referred to Milner & Sequeira (Reference Milner and Sequeira2003) for a full description of both specimens.
2. Systematic palaeontology
Amphibia Linnaeus Reference Linnaeus1758
Temnospondyli von Zittel Reference von Zittel1888
Dissorophoidea Bolt Reference Bolt1969
Olsoniformes Anderson et al. Reference Anderson, Henrici, Sumida, Martens and Berman2008
Family Trematopidae (as Trematopsidae)
Williston Reference Williston1910 recte Milner Reference Milner1985
Diagnosis. A family of dissorophoid-grade temnospondyls from the Upper Carboniferous and Lower Permian of Europe and North America, expanded here to comprise not only the genera Acheloma (including Trematops), Actiobates, Anconastes, Ecolsonia, Fedexia, Phonerpeton, Rotaryus and Tambachia (as per Polley & Reisz Reference Polley and Reisz2011) but also Mordex and Mattauschia. Unique characters diagnosing this expanded concept of the family are (1) greatly enlarged external naris replacing most the dermal exposure of the lacrimal which is a small compact bone bordering the orbit and extending a sliver of bone along the lateral margin of the naris, (2) unsculptured supratympanic flange of tympanic embayment with a slender squamosal component, a semilunar flange of the supratemporal and a tabular process which contacts the squamosal below the supratemporal (it can be argued that this is an olsoniform character with Dissorophidae having a more derived state of this character), (3) tympanic embayment with a ventral border sloping at less than 45° in large individuals so that at least anterior region of embayment tending to a horizontal slit. A non-unique derived character is (4) tympanic embayment generally closed posteriorly by tabular-quadrate contact in large (120mm+) skulls only. A feature of uncertain polarity of this family contra all other dissorophoid-grade temnospondyls is (5) anterior extension of the palatine ramus of the pterygoid contacting the vomer and excluding the palatine from the margin of the interpterygoid vacuity. This is the condition in basal temnospondyls but may be a reversal associated with the novel snout construction required by the enlarged nares.
Comment. This diagnosis and content corresponds in part to those in Polley & Reisz (Reference Polley and Reisz2011) and Schoch & Milner (Reference Schoch, Milner and Sues2014). Both include Ecolsonia and the latter includes a composite Nýřany Mordex (including Mattauschia). Some characters used by Polley and Reisz are not used here (caniniform teeth on premaxilla and maxilla, inflection of the prearticular along the medial ridge of the adductor fossa) either because they do not apply to the primitive Mordex and Mattauschia or they are not known in these taxa.
3. Mordex
Genus Mordex Steen Reference Steen1938
Type and only species. Mordex calliprepes Steen from the Asturian (=Westphalian D), Moscovian, Upper Carboniferous of the Czech Republic.
Diagnosis. As for M. calliprepes, the only species.
Mordex calliprepes Steen Reference Steen1938
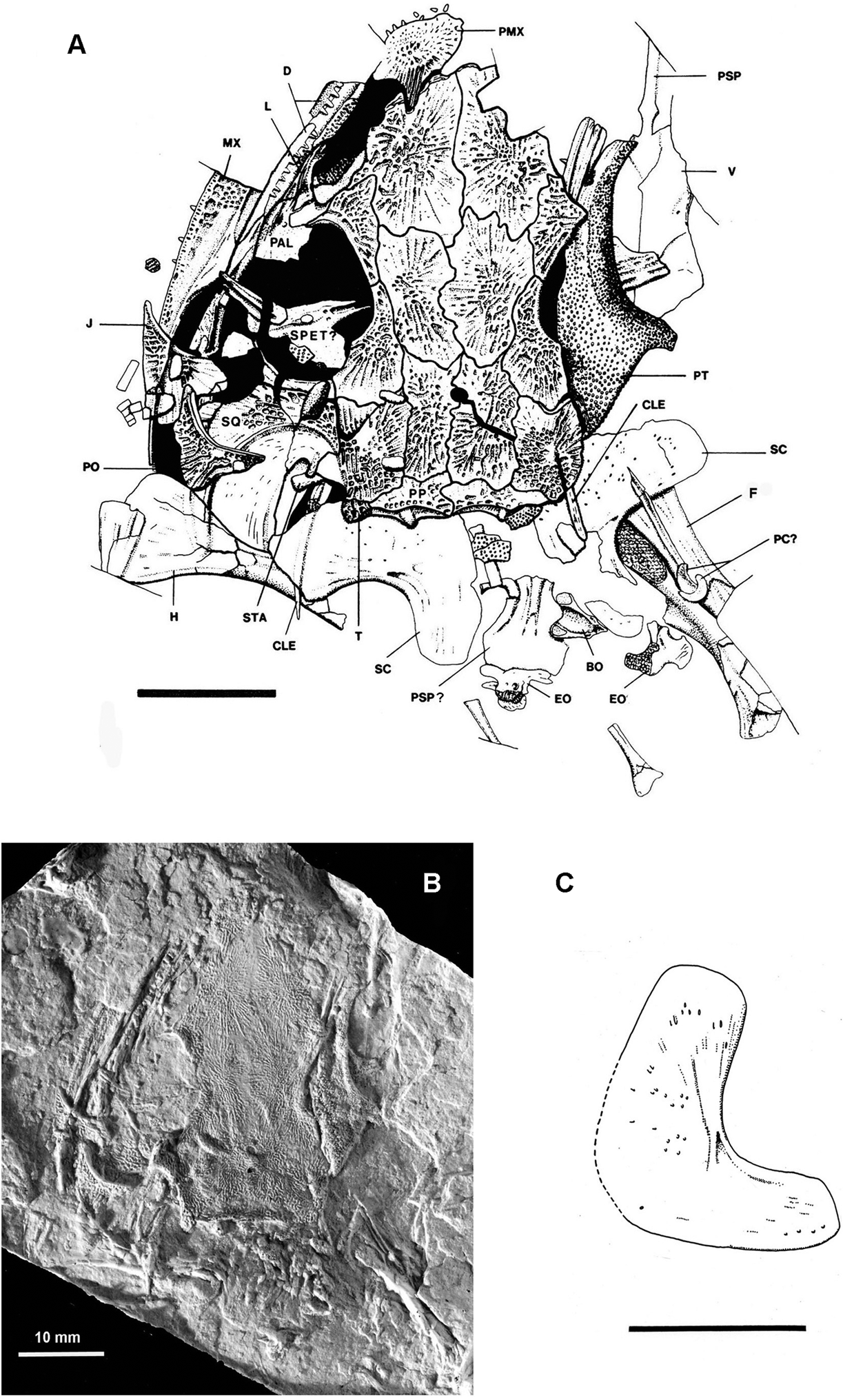
Figure 1 Mordex calliprepes (Steen), NHMUK PV R2817 (holotype and only specimen). (a) Interpretive drawing of entire specimen as preserved. (b) Photograph of silicone rubber peel of complete specimen. (c) Reconstruction of inner face of right scapulocoracoid of NHMUK PV R2817 in lateral aspect. Abbreviations: BO = basioccipital; CLE = cleithrum; D = dentary; EO = exocciptal; F = femur; H = humerus; J = jugal; L = lacrimal; MX = maxilla; PAL = palatine; PC? = probable pleurocentra; PMX = premaxilla; PO = postorbital; PP = postparietal; PSP = cultriform process of parasphenoid; PSP? = probable basal plate of parasphenoid; PT = pterygoid; SPET? = probable sphenethmoid; SC = scapulocoracoid; SQ = squamosal; STA = stapes; T = tabular; V = vomer. For identification of medial roofing bones see Figure 2. Scale bars = 10mm.
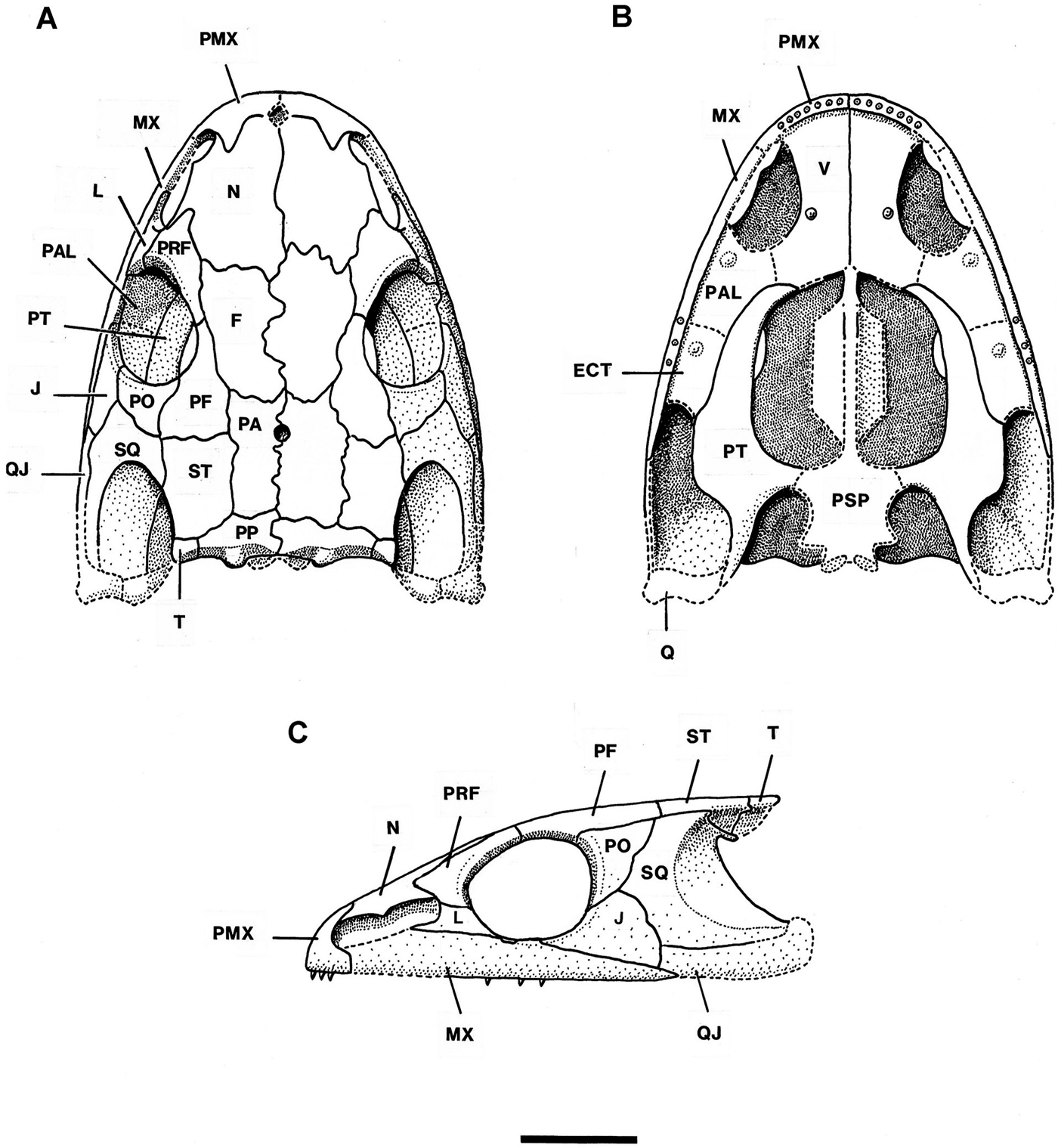
Figure 2 Mordex calliprepes (Steen). Reconstruction of skull of NHMUK PV R2817 in (a) dorsal, (b) ventral and (c) lateral aspects×2. Abbreviations: ECT = ectopterygoid; F = frontal; J = jugal; L = lacrimal; MX = maxilla; N = nasal; PA = parietal; PAL = palatine; PF = postfrontal; PMX = premaxilla; PO = postorbital; PP = postparietal; PRF = prefrontal; PSP = parasphenoid; PT = pterygoid; Q = quadrate; QJ = quadratojugal; SQ = squamosal; ST = supratemporal; T = tabular; V = vomer. Scale bar = 10mm.
Synonymy.
1938 Mordex calliprepes Steen, p. 260, text-fig. 42, pl. V fig. 1.
1947 Mordex calliprepes Steen; Romer, p. 167, fig. 29.
1950 Mordex calliprepes Steen; Gregory, p. 855.
1964 Amphibamus calliprepes (Steen); Carroll, p. 241.
1980 Amphibamus calliprepes (Steen); Milner, p. 453.
1982 Amphibamus calliprepes (Steen); Milner, p. 645.
1986 Mordex calliprepes (Steen); Milner, p. 672, fig. 2.
1986 Amphibamus laticeps (Fritsch); Milner, p. 672, non Fritsch Reference Fritsch1881.
1994 Amphibamus laticeps (Fritsch); Werneburg, pp. 464–465, non Fritsch Reference Fritsch1881.
1994 ‘Limnerpeton' laticeps (Fritsch); Clack & Milner, p. 188, non Fritsch Reference Fritsch1881.
2003 Mordex laticeps (Fritsch); Milner & Sequeira, p. 129, non Fritsch Reference Fritsch1881.
2012 Mordex laticeps (Fritsch); Werneburg; pp. 22–31 abb.18b, non abb. 18a, 19. Partim!
2014 Mordex laticeps (Fritsch); Schoch & Milner, pp. 64–65, non fig. 32. Partim!
Holotype and only specimen. NHMUK PV R2817, a subadult represented by a partly disarticulated skull together with elements of the anterior postcranial skeleton. Only one counterpart is known (Fig. 1a, b) and this has been acid etched, probably by Margaret Steen in the mid-1930s. Other specimens were implied to belong to this species (as Amphibamus calliprepes) by Milner (Reference Milner and Panchen1980). These all belong to Platyrhinops or Mattauschia.
Diagnosis. A primitive trematopid with the following characters.
Derived: External naris elongate and divided into two lobes by lateral flange of nasal; developing dermal ornament suggestive of relatively fine even pitting.
Primitive: Prefrontal–postfrontal common suture excluding frontal from orbit margin; very small marginal teeth with no evidence of pseudocanine enlargement.
Polarity uncertain: Relatively massive premaxilla in a small skull; frontals widen anteriorly; postorbital and squamosal forming anteroposteriorly narrow strut dividing orbit from tympanic embayment; humerus lacking supinator process.
Locality. Nýřany, 13km southwest of Plzeň, the Czech Republic.
Horizon. Main Nýřany Coal, Nýřany Member, Kladno Formation of the Plzeň Basin. Dated as late Asturian (308–307Ma) within the Moscovian, Upper Carboniferous (Opluštil et al. Reference Opluštil, Schmitz, Cleal and Martínek2016).
3.1. Description
3.1.1. Skull roof (Figs 1a, b, 2a, c)
The skull roof possesses all the dermal bones to be expected in a dissorophoid temnospondyl and none shows any trace of lateral-line sulci. The snout is slightly elongate compared with that of Platyrhinops and the external naris is extremely large extending back almost to the orbit margin (Fig. 1a). The orbits are relatively large and also widely separated by the broad frontals and by the prefrontal–postfrontal connections, much more so than in NMP M470 – the small Mattauschia laticeps. The orbits were probably laterally directed as they border the maxillae, there being no lacrimal–jugal connection distal to the orbit. The pattern of crushing suggests that a vertical left cheek of a deep-skulled animal has been crushed sideways and outwards. The squamosal embayment is very large and forms a deep semicircular emargination in the squamosal. A small pineal foramen is situated in the anterior half of the common parietal suture. To the left of the skull are several featureless rectangular platelets of bone which may be displaced sclerotic ossicles.
All dermal bones are either entirely covered with shallow pitting or are pitted centrally with peripheral striate–radiate ornament in regions of relatively rapid growth. The ornamentation is much lighter than in Platyrhinops lyelli skulls of similar size and this by itself suggests that this specimen may represent an immature individual which was still growing when it died. If the central pits are representative of the final state of the ornament, then there is a resemblance to derived trematopids such as Acheloma rather than other primitive trematopids which have much coarser pits on the skull roof. There are no thickened ridges of bone. The left premaxilla is preserved and can be seen to be an unusually robust element possessing a prominent posterodorsal alary process which interdigitates with the anterior edge of the nasal. Such processes occur in most post-edopoid temnospondyls but are highly developed in the dissorophoids. The shape of the premaxilla suggests the presence of an internarial foramen, although there is no certainty that the premaxilla is complete mesially, particularly as its posteroventral ramus, which contacts the maxilla, is missing. Such a massive premaxilla is found in many but not all trematopids. It is typical in large skulls but it is unusual in such a small skull. About eight short, blunt, conical teeth are present in the premaxilla. Only the posterior half of the left maxilla is present. It is of uniform depth from the level of the leading edge of the orbit to the level of the middle of the orbit, then tapering gradually to a point. There is an anterodorsal groove in which the lacrimal sits and another groove on the sloping posterodorsal face into which the jugal must have fitted. The dermal sculpture on the maxilla is shallower than that on other dermal bones. The posterior maxillary teeth are small simple cones, but the anterior teeth are not visible in this specimen.
The nasals and frontals are large, distinctly asymmetrical and still growing proportionately in relation to the rest of the skull roof, their medial, anterior and posterior margins bearing striate ornament. Even without the discovery of a larger specimen, this suggested that the type represents a partly grown individual, the adult form of which might have had a differently shaped snout. The nasals interdigitate both with the premaxillae and the prefrontals. The frontals are wider anteriorly than posteriorly and do not extend beyond the level of the anterior orbit margin. Steen (Reference Steen1938) diagnosed Mordex by its possession of a tiny interfrontal between the nasals and frontals. Carroll (1964, p. 242) suggested that this ‘interfrontal' was probably an artefact produced by the flaking off of a piece of matrix. Study of the specimen confirms that this is so and that an exfoliated flake of coal from the posteromedial corner of the impression of the left nasal has left a surface resembling a small unornamented bone. The prefrontal and postfrontal are primitively large compared with those of most Permian dissorophoids, and meet along the dorsomedial edge of the orbit, thus excluding the frontal from the orbit margin. The postfrontal enlarges posteriorly as a substantial rectangular bone, an unusual condition in dissorophoids.
The lacrimal is perhaps the most interesting component of the skull roof. The left lacrimal is visible as a poorly sculptured U-shaped bone extending from partway along the distal side of the long external naris to narrowly border the orbit margin and to suture with the prefrontal and the palatine. It bears a single, apparently unbranched lacrimal duct. There is a smooth bone surface underlying the posterior region of the external naris and this may also be part of the lacrimal. The reduced lacrimal is of the shape characteristically associated with an elongate external naris extending almost from naris to orbit. Because the anterior end of the maxilla is not preserved in NHMUK PV R2817, the complete shape of the external naris cannot be determined but it appears to have been an elongate ovoid, slightly narrowed in the middle by a convexity in the anterodistal border of the nasal. This type of external naris otherwise occurs only in the temnospondyl family Trematopidae.
Bordering the outer orbit margin, immediately behind the lacrimal, is an unsculptured dorsal exposure of the palatine. Some smooth bone surface representing the dorsal face of the palatal region of the palatine can be seen within the left orbit, and the dorsal exposure extends smoothly from it to form part of the orbit rim. Behind it, the maxilla forms the orbit rim for a few millimetres after which the anterior region of the jugal forms the orbit margin. The jugal is of characteristic dissorophoid shape, narrowing to an anterior point at about the level of the mid-orbit and, as noted above, not contacting the lacrimal or the palatine. Posteroventrally, the jugal is excluded from the skull margin by the maxilla, and on its dorsomedial corner it bears a shallow slot, onto which fits the distal corner of the postorbital. The postorbital is of typical dissorophoid shape, being shallowly triangular with a concave anterior edge.
The skull table is made up of parietals, supratemporals, postparietals and tabulars, all of which are uniformly sculptured. It is relatively short and wide and the tabular–tabular width is greater than the tabular–orbit length. This is a feature of most dissorophoids apart from the Micromelerpetontidae. The parietals are rectangular bones of uniform width, with the pineal foramen in the anterior half of the common suture. The supratemporals are rectangular bones but narrow slightly posteriorly where they form the dorsal margins of the tympanic embayments. The ventrolateral face of the supratemporal is visible as a crushed triangular lappet overlapping the stapes. The many fine cracks on this lappet produced by crushing have been left out of Figure 1a for clarity. The lappet appears to have been unsculptured. The postparietals and tabulars are anteroposteriorly narrow, the postparietals being relatively wide bones which occupy most of the occipital edge of the skull table. The occipital face of each bone is broadly concave but with a central convexity and there is also a convexity posterior to the common suture of the postparietals. The lateral convexities and the thickenings on the occipital face behind them undoubtedly represent buttressing associated with the sutural connections to the underlying exoccipitals. The tiny tabular is slightly expanded laterally and has such a short sutural connection to its neighbours that it may detach readily post-mortem as the right tabular appears to have done. This resembles the situation in Platyrhinops, in which some specimens have no preserved tabulars attached to the skull, although the few that do, demonstrate that the bone was present in life (Milner Reference Milner1982a). The large squamosal forms most of the anterior border of the tympanic embayment and its posterior border is entirely concavely curved. Its medial face is lightly striated and, at its posterodorsal end, it curves sharply round to form a short process appressed to the anteroventral face of the supratemporal component of the supratympanic flange. This small squamosal component to the supratympanic flange is the condition which Bolt (Reference Bolt1974b) described as characteristic for trematopids, the equivalent structure in dissorophids being a large semilunar flange.
3.1.2. Palate and braincase (Figs 1a, 2b)
The only palatal element noted by Steen was the right pterygoid, the quadrate ramus of which is partly obscured by the skull roof. She failed to recognise the vomers and anterior cultriform process, which appear as a detached unit to the right of the pterygoid (Fig. 1a). The vomers and part of the left palatine are visible in dorsal aspect only and hence their dentition is almost unknown. The ectopterygoids are not visible. Those elements present suggest a typical dissorophoid palate (Fig. 2b) except that the pterygoid apparently extends forward to contact the vomer thus excluding the palatine from the margin of the interpterygoid vacuity.
The vomers are anteroposteriorly elongate as in trematopids and some dissorophids and one shows the shape of the medial border of an ovoid internal naris. There is nothing to suggest the presence of a vomerine pit at the anterior end of the vomers. The median suture is comparatively straight except just anterior to the cultriform process insertion where one vomer puts a small lappet of bone over the other. The dorsal face of the left vomer bears a shallow furrow extending from its anterior edge to a point near the posteromedial corner of the internal naris. Such a groove characterises many dissorophoids and is associated with the upward reflexion of the medial region of each vomer as may be seen in the cross-sections of the snouts depicted by Carroll (1964, fig. 6b – Tersomius) and Bolt (Reference Bolt1974a, fig. 3 – Acheloma). In Bolt's sections, the portion of the vomer distal to the furrow is labelled as ‘unnamed septum'. Dilkes (Reference Dilkes1990) referred to comparable structures in Phonerpeton as the medial vomerine septum and used it as a character to define the Trematopidae. It has subsequently been reported in Tambachia and Anconastes (Sumida et al. Reference Sumida, Berman and Martens1998) but the appropriate region is not visible in Actiobates or Ecolsonia. The condition in Mordex is significantly less derived and more like the generalised dissorophoid condition of Tersomius than the sharp ridge seen in Phonerpeton. Essentially it lacks the median vomerine septum.
The parasphenoid possesses a slender cultriform process, expanded anteriorly where it contacts the vomers. The expansion is rhomboidal and appears to suture with the vomers. Just posterior to the anterior terminal expansion of the cultriform process is a second expansion which probably marks the anterior attachment of the sphenethmoid to the parasphenoid. A structure in the left orbit may represent a poorly preserved sphenethmoid. There is an elongate structure of poorly defined outline but with distinct bilateral symmetry running from left to right across the left orbit. It appears to bear a straight ridge on which is a small projection, followed by a pair of pits and then a groove.
Part of the left palatine is preserved in dorsal aspect where it extends into the orbit margin between the lacrimal and the maxilla. All that can be said of its palatal component is that it was relatively broad and, to judge by the position of a thickening on the dorsal side, probably bore a palatal fang.
The pterygoid is of typical temnospondyl form comprising three regions. (1) An elongate palatine ramus extending anteriorly and narrowing gradually. The attempted reconstruction of the palate of NHMUK PV R2817 (Fig. 2b) suggests that the short straight anterior edge represents a short common suture with the vomer as in Acheloma (Bolt Reference Bolt1974a, fig. 1b) or Ecolsonia (Berman et al. Reference Berman, Reisz and Eberth1985, fig. 4). The medial edge of the palatine ramus is shallowly concave unlike those of dissorophids, which are extremely concave, indicating that in Mordex the interpterygoid vacuities were not as large as in most dissorophoids and were straighter sided. (2) A central flattened basisphenoid region which bears medially the base of the basisphenoid articulation and, on its posterior edge, a small transverse pterygoid flange which extends into the subtemporal fossa. The pterygoid bulges ventrally in this region, presumably to accommodate adductor musculature. It is not possible to ascertain whether the basisphenoid articulation was a sutured joint or an open mobile joint as in Ecolsonia. (3) The quadrate ramus, also relatively long, which extends around the posteromedial edge of the adductor fossa. Only the base of this ramus is visible, the rest having presumably been crushed under the skull roof. This ramus was probably originally orientated in a vertical plane. Much of the ventral surface of the pterygoid is coated with a dense shagreen of denticles which are not uniform in size or distribution. The distal side of the palatine ramus bears very few denticles while the medial edge bears densely packed small denticles which occur around the edge of the interpterygoid vacuity almost to the basipterygoid articulation. The rest of the ventral surface is covered by distinctly larger denticles, also closely packed. These extend over the pterygoid flange and the preserved portion of the quadrate ramus. As observed by Carroll (1964), the ventral surface of the pterygoid flange cannot have been used for muscle insertion. Carroll (1964) and Clack & Milner (Reference Clack and Milner2010, p. 288) describe how in Platyrhinops lyelli, the interpterygoid vacuities are covered by rhomboidal platelets of bone which bear denticles and form a complete denticulate surface across the palate (e.g., Clack & Milner Reference Clack and Milner2010, figs 2, 6, 7). In Mordex, several isolated denticle-bearing platelets are scattered around the specimen and these are presumably remnants of a similar structure across the interpterygoid vacuities. Such denticulate palatal platelets are widespread in temnospondyls wherever permitted by the circumstances of preservation and their structure and distribution have been discussed recently by Gee et al. (Reference Gee, Haridy and Reisz2017).
Four isolated bones scattered between and behind the scapulocoracoids of the specimen may represent displaced elements of the palate and occiput. One is a plate of bone apparently with symmetrical anterolateral concavities and some superficial grooves, which also suggest bilateral symmetry. The general shape and symmetry suggest that it could be the basal plate of the parasphenoid and the grooves may be comparable to the deep furrows across the ventrolateral surface of the parasphenoid in the primitive trematopid Tambachia (Sumida et al. Reference Sumida, Berman and Martens1998, text-fig. 7). Next to it is another small symmetrical triangular ossification which may be the basioccipital seen in dorsal aspect. Most of its surface is taken up by a broad channel, presumably the floor of the foramen magnum. The medial corner of the triangle has two small expansions behind it, while the other two corners are expanded as cartilaginous facets for the exoccipitals. Behind the presumed parasphenoid base and basioccipital are two more small but complex ossifications, which may be the exoccipitals.
The reconstruction of the palate (Fig. 2b) is inevitably tentative and is based on the few known palatal elements and their mirror images superimposed on the skull outline as deduced from the dermal roofing bones. The ectopterygoid and quadrate are entirely hypothetical, and the sphenethmoid and the basal plate of the parasphenoid are effectively so, as the presumed structures in the specimen have little information content. The quadrate ramus of the pterygoid appears narrower than in the specimen as it is assumed to be orientated in a vertical plane.
3.1.3. Mandible (Fig. 1a)
Only a few pieces of the left mandible are recognisable. Twelve short conical teeth are visible on a strip of dentary accompanied by a patch of coronoid denticles. The coronoid sutures are not visible. Behind the dentary is the anterior region of a broader structure which appears to be the left surangular. Behind the detached left postorbital and under the flattened distal end of the left humerus is a raised outline which appears to be a continuation of the curvature of the dentary and surangular and is probably the posterior end of the left mandible.
3.1.4. Stapes (Fig. 1a)
The left stapes is present, although partly obscured by the supratemporal. The preserved middle shaft is slightly under 4mm long and uniform in width. The distal end may be broken and it is possible that the shaft was relatively long and slender as in other dissorophoids. No stapedial foramen is visible.
3.1.5. Hyobranchial element (Fig. 1a)
A blade-like bone bearing a ridge along its length is visible in the left orbit to the left of the presumed sphenethmoid. Clack & Milner (Reference Clack and Milner2010, figs 5c, 6c) found similar elements in the palatal region of Platyrhinops and concluded that they were ceratohyals retained in terrestrial adults and modified for tongue manipulation purposes as in living salamandrids. Mordex appears to possess a similar element, indicating that it too underwent a metamorphosis converting its branchial skeleton into a lingual support skeleton characterised by blade-like ceratohyals. The presence of such structures in a trematopid as well as an amphibamid suggests that this form of lingual support skeleton may have been a general character of metamorphosing dissorophoids of the subgroup Xerodromes. In the zatracheid Acanthostomatops there appears to have been a distinctly different rearrangement of the branchial skeleton at metamorphosis to produce a differently structured lingual support system (Witzmann & Schoch Reference Witzmann and Schoch2006; Witzmann Reference Witzmann2013), suggesting that such adaptations to a bimodal life history occurred more than once.
3.1.6. Postcranial skeleton (Fig. 1a, c)
The only elements of the axial skeleton which can be identified are two pleurocentra resting on the right humerus, and two or three anterior ribs scattered behind the skull. The centra are both small crescent-shaped ossifications, broader on the convex face. Both the single intercentrum and the smaller paired pleurocentra of typical temnospondyl vertebrae are crescent-shaped wedges (the ‘rhachitomous condition'), but the small size of the wedges in NHMUK PV R2817 makes it almost certain that they are pleurocentra. There are two short ribs at the back of the specimen and a possible third rib under some denticle-bearing plates of bone immediately behind the skull table. All three structures are exceptionally short and straight, as are the anterior ribs of many early tetrapods, and one has a slightly bicipital head with a large tuberculum and a small capitulum.
The only preserved components of the pectoral girdle are the scapulocoracoids and parts of the cleithra. The pectoral elements were vaguely outlined by Steen (Reference Sequeira and Milner1938), but illumination of a cast of the specimen at a low angle of incidence reveals clearly the internal faces of both scapulocoracoids. The scapular blade is a thin flat plate of bone, most of the surface of which is smooth. On the inner face, a few striations radiate from the region of the supraglenoid foramen, which is very prominent (Fig. 1a, c). Above the foramen is a broad ridge, the supraglenoid buttress, which supports the back of the vertical scapular blade and which forks to either side of the foramen. The coracoid extends backwards well behind the glenoid region, forming an elongate plate of bone of almost equal length to the height of the scapula (Fig. 1c). This shape is similar to that in trematopids (Olson Reference Olson1941, figs 11b, 11c) and in Ecolsonia (Berman et al. Reference Berman, Reisz and Eberth1985, fig. 11a) but contrasts with that of dissorophids in which the coracoid is short and the scapular blade is tall (Dissorophus Demar Reference Demar1968, text-fig. 13). The form of the glenoid fossa is unknown in Mordex as the outer faces of the scapulocoracoids are not exposed. Largely crushed under the left scapular blade, but projecting from each side of it, is a slender spike-like structure, slightly expanded at one end, which is presumed to be the stem of the left cleithrum. On the right side of the specimen is one spike of bone extended backwards from under the right supratemporal to overlay the scapulocoracoid, while another slender shaft of bone also projects back from above the scapula. These two bones may represent the shafts of the cleithrum and the clavicle, but it is not possible to identify either of them with certainty.
Three incomplete limb bones are visible. On the left is the distal end of a humerus which is flattened but appears to be broader-ended than that of Platyrhinops and does not appear to be perforated by any foramina or bear a supinator process. An associated fragment may pertain to the left radius or ulna. The limb bones may have still been poorly ossified at the ends, which are exceptionally crushed. On the right is a larger limb element that appears to be a femur bearing an adductor crest. No dermal scales are visible on this specimen.
4. Mattauschia
Genus Mattauschia
Type and only species. Mattauschia laticeps (Fritsch Reference Fritsch1881) from the Asturian (=Westphalian D), Moscovian, Upper Carboniferous of the Czech Republic.
Diagnosis. As for Mattauschia laticeps, the only species.
Etymology. After H.M. Mattausch, a resident of Nýřany (then Nürschan), who made a large collection of Nýřany amphibian material and donated it to the Museum für Naturkunde, in Vienna in 1898 (Steindachner Reference Steindachner1899, p. 32), including the large skull described here. His collection was second in size only to that of Anton Fritsch, but he is little remembered now.
Mattauschia laticeps (Fritsch Reference Fritsch1881) Milner & Sequeira Reference Milner and Sequeira2003

Figure 3 Mattauschia laticeps (Fritsch), lectotype NMP M470 (=Fritsch Orig. 94). Skull and pectoral region preserved in ventral aspect. Abbreviations: CL = clavicle = H = humerus; ICL = interclavicle; PAL = palatine; PMX = premaxilla; SC = scapulocoracoid; ST = supratemporal. Scale bar = 10mm.

Figure 4 Mattauschia laticeps (Fritsch), NMW 1898-X-47. (a) Photograph of silicone rubber peel of complete specimen. (b) Interpretive drawing of entire specimen, as preserved. Abbreviations: ART = articular; COR = coronoid; D = dentary; F = frontal; J = jugal; L = lacrimal; MX = maxilla; N = nasal; PA = parietal; PAL = palatine; PMX = premaxilla; PRA = prearticular; Q? = quadrate; QJ = quadratojugal; SA = surangular; SPL 1 = first splenial; SPL 2 = second splenial; SQ = squamosal; ST = supratemporal; T = tabular; V = vomer. For identification of circumorbital bones, see Figure 6. Scale bars = 10mm.

Figure 5 Mattauschia laticeps (Fritsch), NMW 1898-X-47. (a) Left nasal. (b) Left antorbital region showing lacrimal with posterior duct openings and dorsal exposure of palatine. (c) Anterior end of left mandible showing left symphyseal thickening and symphyseal tooth. NMW 1983-32-81. (d) Isolated premaxilla from a large skull, sculpture omitted where eroded. NMW 1983-32-79. (e) Isolated left squamosal – acid etched mould. (f) Interpretive drawing of left squamosal in lateral aspect from silicone cast. Abbreviations: LD = lacrimal duct openings; MX = maxilla; PAL = palatine; PRF = prefrontal; V = vomer. Scale bars = 10mm.

Figure 6 Mattauschia laticeps (Fritsch). Reconstruction in (a) dorsal aspect and (b) left lateral aspect, based on NMW 1898-X-47. Abbreviations: F = frontal; J = jugal; L = lacrimal; MX = maxilla; N = nasal; PA = parietal; PAL = palatine; PF = postfrontal; PMX = premaxilla; PO = postorbital; PP = postparietal; PRF = prefrontal; PT = pterygoid; Q = position of quadrate; QJ = quadratojugal; SQ = squamosal; ST = supratemporal; T = tabular. Scale bar = 10mm.

Figure 7 Mattauschia laticeps (Fritsch), NMW 1983-32-79. Reconstruction of left mandible in dorsal aspect. Natural size. Abbreviations: ART = articular; COR = coronoid; D = dentary; PRA = prearticular; SA = surangular; SPL 1 = first splenial; SPL 2 = second splenial. Scale bar = 10mm.
Synonymy (not including possible larvae).
1879 Microdon laticeps Fritsch p. 28 nomen nudum.
1881 Limnerpeton laticeps Fritsch pp. 148–51, plate 31 fig. 1, text-fig. 92, non plate 36, fig. 1, text-figs 89–91.
1881 Limnerpeton macrolepis Fritsch pp. 151–52, plate 32, figs 1–5.
1938 Limnerpeton laticeps Fritsch; Steen p. 261 Partim!
1938 Limnerpeton macrolepis Fritsch; Steen pp. 263–64.
1947 Potomochoston salamandroides (Fritsch); Romer p. 146, non Fritsch, non Steen.
1986 Mordex calliprepes (Steen); Milner, p. 672, Partim! Non fig. 2.
1986 Amphibamus laticeps (Fritsch); Milner p. 672.
1994 Amphibamus laticeps (Fritsch); Werneburg pp. 464–65.
1994 ‘Limnerpeton' laticeps (Fritsch); Clack & Milner p. 188.
2003 Mordex laticeps (Fritsch); Milner & Sequeira pp. 126–32 figs 1–5. Partim!
2007 Mordex laticeps (Fritsch); Milner p. 118A.
2012 Mordex laticeps (Fritsch); Werneburg; pp. 22–31 abb. 18a, 19, non abb. 18b. Partim!
2014 Mordex laticeps (Fritsch); Schoch & Milner, pp. 64–65 fig. 32a, Partim!
Lectotype (after Milner Reference Milner and Roček1986). NMP M470/471 (=Fritsch Orig. 94; St. 123), a small skull in ventral aspect and a partial postcranial skeleton in counterpart. The specimen comprises three blocks of coal and a galvanotype of one block. NMP M470 comprises the anterior half of the specimen with the bone preserved as figured by Fritsch (Reference Fritsch1881, plate 31) and in this work (Fig. 3). A second fitting slab bears the posterior half of the specimen in which the bone has been etched away to leave a mould in the coal, and there is a galvanotype made from the acid-etched posterior slab. NMP M471 is the counterpart block corresponding to the posterior end of the anterior block and the entire posterior block. It does not include any cranial material. It should be noted that Fritsch's Plate 31 incorporates substantial rearrangement of the postcranial elements to fit his plate format. In the original specimen, the skeleton topology is still represented across the slab despite some disarticulation. The lectotype skull is figured and briefly reviewed here but the reader is referred to the full description of this specimen by Milner & Sequeira (Reference Milner and Sequeira2003).
Diagnosis (based mainly on NMW 1898-X-47). Because this taxon is argued to be the basal trematopid with no synapomorphies beyond the family character states, the diagnosis entirely comprises primitive and unpolarised character states.
A primitive trematopid with the following characters.
Primitive: External nares relatively broad ovoids and nasals correspondingly slender; nasal without lateral lappet tending to separate external naris into anterior and posterior regions; prefrontal–postfrontal common suture excluding frontal from orbit margin; dentary with space for about 75 small teeth on each jaw ramus; prearticular not extending to back of adductor fossa and not expanded posteriorly; maxillary/anterior mandibular jaw margin convexly curved when viewed from above or below; dermal sculpture of coarse pitting.
Polarity uncertain: Postorbital and squamosal forming anteroposteriorly narrow strut dividing orbit from tympanic embayment; sculptured supratemporal contribution to supratympanic flange; mandibular teeth slightly inwardly curved.
Referred material. NMP M472 (the lectotype of ‘Limnerpeton' macrolepis Fritsch), a small disarticulated and scattered specimen (redescribed fully by Milner & Sequeira Reference Milner and Sequeira2003, pp. 129–32 figs 4, 5). NMW 1898-X-47, a large incomplete skull and left mandible in dorsal aspect. Only one counterpart is known (Figs 4, 5a–c, 6, 7) and this was acid etched and cast by Angela C. Milner during 1984 (figured by Werneburg Reference Werneburg2012 fig. 19). NMW 1983-32-79, an isolated left squamosal from a large skull, in external aspect. Only one counterpart is known (Fig. 5e, f) and this was acid etched and cast by Angela C. Milner in 1983. NMW 1983-32-81, an isolated premaxilla from a large skull (Fig. 5d).
Werneburg (Reference Werneburg2012) has attributed many small dissorophoid larvae (‘branchiosaurs') from Nýřany to the binomen Mordex laticeps. This attribution will be discussed in a separate section later in this paper.
Locality. Nýřany, 13km southwest of Plzeň, the Czech Republic.
Horizon. Main Nýřany Coal, Nýřany Member, Kladno Formation of the Plzeň Basin. Dated as late Asturian (308–307 Ma) within the Moscovian, Upper Carboniferous (Opluštil et al. Reference Opluštil, Schmitz, Cleal and Martínek2016).
Description
NMP M470 has a skull length of about 33mm whereas NMW 1898-X-47 has an approximate midline length of 120mm. The squamosal NMW 1983-32-79 is almost identical in size to that of the larger skull. NMW 1898-X-47 is relatively large for a Late Carboniferous/Early Permian dissorophoid and may reasonably be presumed to represent the adult size. The other specimens are a quarter to a third of this size in linear dimensions and although clearly terrestrial post-metamorphic individuals with well-ossified scapulocoracoids and pelves are taken to be juveniles or subadults with some characters incompletely expressed.
The description of the cranial material is based on NMP M470, NMP M472 and NMW 1898-X-47 (with reference to NMW 1983-32-79 for the squamosal and NMW 1983-32-81 for the premaxilla). The description of the postcranial elements is, of necessity, based entirely on NMP M470. Because of the differences in size and structure between the major specimens, they are described separately to avoid ambiguities about the nature of characters in different sizes of individual. The smaller specimens are described first, followed by NMW 1898-X-47, incorporating a description of the isolated premaxilla and squamosal.
4.1. NMP M470 and M472
4.1.1. General features
The skeleton of the lectotype NMP M470 is disarticulated but the various elements have been only slightly displaced. The skull is represented by the underside of the greater part of the skull roof, with one detached bone in the orbit (Fig. 3). Unfortunately, damage to the narial region means that the shape of the external naris cannot be determined. The palate is largely missing but both mandibles are crushed together superimposed on the left cheek region. The orbits are relatively closely spaced, much more so than the slightly larger specimen of Mordex. Immediately behind the skull are several elements of the pectoral girdle together with a humerus and numerous neural arches and ribs. The counterpart posterior slabs bear more neural arches and trunk ribs, some forelimb elements, the right half of the pelvis and many elements of the hind limbs. Dorsal and gastral dermal scales are scattered throughout the slabs and are still in associated sheets around the posterior trunk region, as described by Milner & Sequeira Reference Milner and Sequeira2003. NMP M472 is a completely disarticulated and scattered specimen. It was described fully by Milner & Sequeira (Reference Milner and Sequeira2003). The description of the palate is summarised here as it includes information not found in any other specimen.
4.1.2. Skull (Fig. 3)
The skull of NMP M470 was about 33mm in midline length. The bones are described as they appear in ventral aspect. An impression of some of the sculpture on the dorsal surface of the right supratemporal is preserved and shows honeycomb-type pitting, much more robust than in the larger skull of M. calliprepes. There are squarish nasals, elongate frontals showing a slight expansion at the anterior end, and narrow prefrontals which extend back to meet the postfrontals, excluding the frontals from the orbit margin. The postfrontals have narrow anterior extensions and a relatively expanded ovoid posterior region. The parietals are large rhomboidal elements, narrower anteriorly next to the interorbital region. The pineal foramen is relatively large and situated just anterior to the middle of the interparietal suture. There are no intertemporals, and the supratemporals are large square elements, extending posteriorly behind the level of the parietals. The postparietals are wide rectangular bones, about half the area of the parietals and narrowing laterally where they extend behind the supratemporals. Each postparietal bears an occipital flange bordering the foramen magnum. The left tabular is present though slightly detached. It is a small element separated widely from the parietal. It bears a tiny posteriorly directed process on its posterolateral corner. The lateral edge overlapping the tympanic embayment region has a convex expansion. The left postorbital is a triangular bone, wider than long and apparently only slightly wedged between the supratemporal and squamosal. Part of a large left squamosal is visible. It appears to have a concave posterior edge forming the anterior margin of a large tympanic embayment but this region is damaged. In the left orbit is a bone which appears to be the right premaxilla with an attached nasal fragment. It has a curved row of tooth bases indicative of its identity. It is evidence of a robust and distinctive premaxilla such as is also present in Mordex (Fig. 1).
Little of the palate is clearly visible in NMP M470 but there are several fragments of densely denticulate bone wedged between the two mandibles. These are probably pieces of the palatine and pterygoid, or possibly ectopterygoid. They demonstrate that the lateral palatal bones bear a dense covering of denticles, also seen in Mordex. Though comprising damaged and disarticulated material, NMP M472 is a useful source of information about the palate, as vomers, a pterygoid and the parasphenoid are present. The vomers possess smooth concave edges which appear to be the anterior margins of the interpterygoid vacuities. No vomerine fangs are visible but a dense covering of denticles is present. The left pterygoid is relatively complete. The ventral face of the central region and the palatine ramus are covered in a dense sheet of denticles. The palatine ramus widens from an anterior point suggesting that the palatine narrowed posteriorly and that the ectopterygoid may have been narrow. The quadrate ramus is broad but may have been crushed from a vertical to a horizontal plane. The basipterygoid process is broad but the surface for articulation with the basisphenoid is obscured. The parasphenoid is largely visible in ventral aspect. The broadly rectangular basal plate bears a sharply defined triangle of denticles at its junction with the cultriform process. Carotid foramina with elongate entry grooves are present on either side of the denticle patch. The cultriform process is relatively slender. Anteriorly it is damaged but appears to have had a single terminal expansion.
4.1.3. Mandible (Fig. 3)
In NMP M470, parts of the middle and posterior regions of both mandibles are present. The tooth-bearing strip of bone overlying the outer edge of the left orbit appears to be the right dentary together with part of a denticle-bearing coronoid. The preserved fragment of dentary has space for about 36 teeth. The tooth bases and positions suggest that the dentary teeth were small, equal in size and closely spaced. Medial to the tooth row is a distinct shelf, apparently the medial surface of the dentary on which the coronoids would have been situated. At the anterior end is a row of denticles, distinctly smaller than the main tooth row, and this is presumably a row of coronoid denticles. The left mandible is represented by the entire posterior region which is severely crushed. The angular is visible in ventral aspect and can be recognised by its dermal ornament in the form of radiating striations. Behind it is a mould of the articular, visible as its bilobed condylar surface.
4.1.4. Axial skeleton
The axial skeleton of NMP M470 is disarticulated but not much scattered. One articulated series of four trunk vertebrae are 13.8mm in total length. The distance from the back of the skull to the ilium is 74mm, which suggests a presacral column of about 22 vertebrae. The neural arches are present as separately ossified halves with very low neural spines and distinct anterior and posterior zygapophyses. The middle trunk series bear prominent rib articulations. In the same middle trunk region, at least two isolated intercentra are present. They have been dorsoventrally flattened and appear as small ovals of bone, pointed at both ends and with a medial suture, implying that the intercentrum has recently ossified from two separate halves and that the animal was a post-metamorphic juvenile when it died. The trunk ribs are relatively small and straight or slightly curved. The anterior trunk ribs, represented by a clump of ribs just behind the skull, are larger with expanded heads. One or two pairs situated next to the pectoral girdle have slightly expanded distal shafts. No uncinate processes are visible. The posterior trunk ribs are shorter, thinner and unossified distally. Close to the pelvis are four caudal haemal arches, each comprising a pair of rod-like bones fused to give an articulating surface proximally and terminating together as a fused point distally.
4.1.5. Appendicular skeleton (Fig. 3)
The pectoral girdle of NMP M470 is represented by the partly hidden interclavicle, both clavicles, and both scapulocoracoids, also each partly hidden. No cleithra could be recognised although it is possible that the cleithral shafts might be present, unrecognised, among the anterior trunk ribs. The interclavicle is a small rhomboid, about as long as wide and fully ossified. It is visible in internal aspect and the nature of its dermal ornament is unknown. The clavicles are long structures with narrow blades and the articulated dermal pectoral girdle would have been a narrow strut-like structure rather than a large plate-like structure. The scapulocoracoids are fully ossified, an unusual feature in such a small temnospondyl. Each scapulocoracoid is a flat element with a broad crescent-shape or kidney shape (Fig. 3). There is a thickened strut extending down the posterior concave edge and a foramen on the edge of the posteroventral extension of the coracoid. The only anterior limb elements which can be identified are the humeri and a radius. The left humerus, close to the skull, is a slender element comparable in proportions to those of branchiosaurids and dissorophids. Neither end is greatly expanded and no supinator process is visible. In this, it resembles that of Mordex rather than those of more derived trematopids. The right humerus, partway down the trunk is similar and is accompanied by a slender radius.
The lateral face of the right side of the pelvic girdle is visible (Milner & Sequeira Reference Milner and Sequeira2003, fig. 3). The ilium, ischium and pubis are sutured together, an unusual feature in such a small temnospondyl. The ilium bears a large acetabulum and a relatively stout, posterodorsally directed, dorsal shaft. The dorsal shaft is simple with only a slight suggestion of dorsal expansion. The pubis is represented by a poorly defined patch of bone, as if it were just in the process of ossification. The ischium is fully ossified and is a large slightly curved plate of bone extending well behind the ilium. It appears to have extended forwards under the pubis which is wedged between it and ilium so that much of the area immediately below the acetabulum is occupied by the pubis. The visible femur is relatively large but poorly preserved. Both sets of tibiae and fibulae are present. The tibia is the larger element, relatively straight with a highly expanded proximal end and a less expanded distal end. The smaller fibula is slightly bowed and is expanded principally at the distal end. Numerous pedal phalanges are scattered around the posterior part of the specimen.
4.1.6. Scales
Large numbers of dispersed and clumped scales are associated with NMP M470. There appear to have been both dorsal scales and ventral gastralia present. The dorsal scales are very small circular structures while the gastralia are larger ovoid scales with a thickening along one edge.
4.2. Large specimens
4.2.1. Skull roof (Figs 4, 5a, b, 5d–f, 6)
The large skull, NMW 1898-X-47, probably belonged to an adult as the coarse reticulate pitting shows no evidence of ‘stretching' except on the anterolateral edge of the squamosal and the sutures are poorly expressed. Their position may be deduced from the ornament patterns on the bones and the furrows between ornamented areas but the actual sutures are visible only in some regions and have had to be depicted in oversimplified form in Figure 4. There is no evidence of lateral-line sulci.
Very little can be seen of the marginal tooth-bearing series of bones. Only the right premaxillary is present as a very crushed, poorly preserved structure. An alary process appears to be present as in the type skull. An isolated premaxilla NMW 1983-32-81 (Fig. 5d) may belong to this taxon. It is about 22mm along the jaw margin with a very long alary process and heavily pitted dermal ornament. The ornament identifies it as temnospondyl and the only temnospondyls from Nýřany which were large enough to possess such a premaxilla are Capetus, Mattauschia and possibly Mordex (the premaxilla of Cochleosaurus is very different in shape). The premaxilla of Capetus has finer pitting along the jaw margin and a shorter alary process (Sequeira & Milner Reference Sequeira and Milner1993, text-fig. 4) and this specimen can be deduced to be dissorophoid by elimination. A reason for positive attribution to Mattauschia is the smooth curve on the right side as preserved corresponding to the margin of the large oval external naris. A long alary process partly bordering the naris can be seen in Fedexia (Berman et al. Reference Berman, Henrici, Brezinski and Kollar2010, figs 4b, 5b). The middle section of the left maxilla is present in NMW 1898-X-47 and bears heavy ornament. The anterior region is missing and the posterior region is obscured by the left jugal. The middle region does not form part of the orbit margin, being excluded by the palatine. No premaxillary or maxillary teeth are preserved.
The external naris is a very large ovoid fenestra of a shape and size otherwise found only in Fedexia (Berman et al. Reference Berman, Henrici, Brezinski and Kollar2010, figs 4–7) and in indeterminate juvenile trematopids (e.g., MCZ 2531) from the Texas Permian (Dilkes Reference Dilkes1993, text-fig. 4). Mordex, Ecolsonia and more derived trematopids share an elongate but relatively narrow external naris with a slight waisting produced by a convexity on the distal margin of the nasal. In this specimen however, the external naris is much wider with a smoothly convex medial margin formed by a smoothly concave distal edge to the nasal. The nasal (Fig. 5a) is unusually slender for a temnospondyl, broadening slightly posteriorly. It does not resemble that of any other dissorophoid and appears to be a unique feature of Mattauschia. Even the nasals of Fedexia are broader relative to their length. The prefrontal is a typical triangular bone bordering the orbit and with a deep rim along its orbital edge. It is relatively large and primitively contacts the postfrontal posteriorly. Because the external naris is unusually large, it is bordered by the prefrontal that has a shallowly concave anterodistal face as a result. The lacrimal (Fig. 5b) is even more extremely modified in relation to the external naris and resembles that in Fedexia, Ecolsonia and some trematopids. Most of the enlargement of the naris has been at the expense of the anterior region of the lacrimal which is a compact rhomboidal bone wedged between naris, orbit and maxilla. It extends a slender sliver of bone anteriorly, dorsal to the maxillary, along the distal margin of the naris. The main sculptured dermal surface is triangular but posteriorly there is a wedge of smooth bone within the orbit margin between palatine and prefrontal. This wedge bears the two posterior openings of the lacrimal duct. Behind the lacrimal, forming the distal border of the orbit is a dorsal exposure of the palatine (Fig. 5b). It bears some dermal sculpture and forms the orbit border between the lacrimal and the jugal, thus conforming to the typical large dissorophoid condition (Bolt Reference Bolt1974b). The broad smooth medial face of the dorsal extension of the palatine forms a major part of the orbit wall. The left jugal is incompletely preserved but is of typical dissorophoid type, namely a triangular bone with a centre of ossification at the level of the back of the orbit. Because of damage to the specimen, it cannot be observed whether or not the jugal forms part of the skull margin.
The frontals are relatively short rectangular ossifications, about twice as long as wide, and widening only slightly in the region of the prefrontal–postfrontal contact which excludes them from the orbit margin. The parietals are incomplete posteriorly but appear to be of conventional temnospondyl configuration. The large pineal foramen is very slightly wider than long. The postorbital is a shallow, roughly triangular bone forming the posterior margin of the orbit. Posteriorly, it is wedged shallowly between the squamosal and the supratemporal. Its distal end forms a slightly elongate extension ahead of the squamosal. The postfrontal has a relatively small contact with the orbit margin and is expanded posteriorly. Medially it forms a shallow wedge, narrowing the contact between the frontal and the parietal. The supratemporal is an anteroposteriorly elongate rectangular bone which broadly borders the dorsal margin of the large tympanic embayment. This tympanic border bears an expansion of sculptured bone partway along the margin. This was depicted as a laterally directed expansion by Milner (Reference Milner and Roček1986, fig. 2), but appears to be the ventrally directed supratemporal contribution to the supratympanic flange crushed outwards. If so, it demonstrates that this structure bore dermal sculpture in Mordex as in Ecolsonia. The supratemporal component to the supratympanic flange is not visible and is presumably crushed under the skull table. The squamosal is of a type frequently found in dissorophoids in that the sculptured dermal region is roughly crescent shaped, bordering the anterior edge of the tympanic embayment, while there is a large smooth surface inside the tympanic embayment. The sculptured region is unusually anteroposteriorly narrow, particularly at the medial end, which is widely separated from the tabular. Distally, the squamosal extends only about halfway back along the ventral side of the tympanic embayment. The isolated left squamosal NMW 1983-32-79 (Fig. 5e, f) merits separate description at this point, as it is less crushed and abraded than that in NMW 1898-X-47 and its precise sutural margins can be seen. There is slight crushing anterior to the stapedial notch and the resulting cracks and raised wedge of bone have been depicted in Figure 5f. The squamosal is slightly larger than that in the large skull, the sculptured region having a maximum width of 32mm as against 29mm in NMW 1898-X-47. The sculptured region is crescent shaped, the semicircular concave edge forming the anterior half of the tympanic border. Within the concavity of the tympanic border is a lightly striated thick shelf of bone bordering an acute V-shaped stapedial notch. The dorsomedial region of this shelf forms the anterior region of a supratympanic flange, but does not show the expansion of this flange that one would expect in a true dissorophid (Bolt Reference Bolt1974b). Extending around the anterior and lateral edges of the bone is a lightly striated irregular shelf which formed an undercutting sutural connection with the supratemporal, postorbital, jugal and quadratojugal. It has the same relationship to the sculptured region as that in Micromelerpeton (Boy Reference Boy1972, abb. 4g).
The quadratojugal of NMW 1898-X-47 is a relatively massive bone. The anterior region is lost and so its anterior extent and possible contact with the maxilla are unknown. The dermal sculpture is exceptionally rugose and it is probable that an expanded sculptured flange of bone hung down the cheek below the level of the jaw margin. This is the condition in Ecolsonia (Berman et al. Reference Berman, Reisz and Eberth1985, figs 7c, 8c) and the quadratojugal of NMW 1898-X-47 bears a distinct resemblance to that of Ecolsonia seen in dorsal aspect (Fig. 4 and Berman et al. Reference Berman, Reisz and Eberth1985, fig. 8a). Medial to the dermal exposure of the quadratojugal is the smooth ventral face of the embayment, made up largely of the smooth squamosal flange. In some temnospondyls, the quadratojugal contributes to the posterior region of this smooth flange but no suture can be seen which might suggest a quadratojugal–squamosal boundary in this region. Posteriorly, the sculptured region of the quadratojugal seems to extend a slender process of dermal bone above the quadrate although sutures are not clear in this region. It is possible but not certain that this is part of a crushed quadrate–tabular contact enclosing the back of the tympanic embayment. The tabular is only partly preserved on the edge of the specimen. It is a quite substantial bone with at least a slight posterolateral extension. Again, this may be part of a process of bone enclosing the back of the embayment. The quadrate is not clearly visible but there are two lumps of unsculptured bone, side by side at the posterior margin of the embayment and these probably represent the bilobed condyle of the quadrate, corresponding to the bilobed cartilage-lined pockets on the articular. There is no evidence of a blade-like anterior quadrate process in the tympanic embayment as found in some dissorophids (e.g., Cacops, see Fig. 10e).
4.2.2. Palate (Fig. 4)
Because NMW 1898-X-47 is preserved in dorsal aspect, only a few parts of the palate are visible in the narial, orbital and otic regions (Fig. 4). Through the large left external naris, can be seen part of the left vomer bordering the internal naris. The vomer appears to have an upturned rim bordering the internal naris and small denticles can be seen on the edge of this rim. The choanal margin appears be bordered by denticles, a situation which also occurs in Platyrhinops lyelli from Linton (Clack & Milner Reference Clack and Milner2010). The choana is also relatively large but narrower than the external naris. The anteromedial and posteromedial faces of the internal naris are almost perpendicular to one another and are separated by a very sharp curvature through 90°.
A fragment of the dorsal face of the palatine and much of the palatine ramus of the pterygoid can be seen in the left orbit. They demonstrate that the interpterygoid vacuities are large with very curved lateral margins. The pterygoid extends well forwards of the palatine and probably contacts the vomer, excluding the palatine from the interpterygoid vacuity margin.
4.2.3. Mandible (Figs 4, 5c, 7)
The mandible of NMW 1898-X-47 is 135mm long in a straight line from the mandibular symphysis to the back of the articular. It is visible in dorsal aspect and is crushed dorsoventrally, particularly at the posterior end. Traces of most of the ten constituent bones presumed to be present are visible, although the coronoid denticles obscure the margins of the individual coronoids. The sutural relationships of the mandibular elements are, where visible, almost identical to those in Tersomius (Carroll 1964, fig. 8).
The dentary is about 110mm long and extends back to just behind the level of the anterior end of the adductor fossa. It bears a single row of marginal teeth, there being space for up to 75 teeth. About 40 teeth are present in approximate alternation, although occasional sequential pairs of teeth are visible, the whole being presumably the product of a ‘zahnreihe system'. The marginal teeth are small, no more than 3mm in height and are simple cones, slightly bucco-lingually flattened at the unicuspid tip, and slightly incurved lingually. Each tooth bears about 12 grooves around the base, the external manifestation of simple labyrinthodont infolding. This large number of small marginal teeth is a distinct character separating Mattauschia from Ecolsonia, which has about 33 larger teeth in each mandibular ramus. Both genera share the inward curvature of the marginal dentition. At the anterior end of the dentary is a thickening at the mandibular symphysis (Fig. 5c). This thickening does not bear any certain symphyseal fangs but there is a detached tooth lying on top of the thickening that also bears two pits that may represent fang pits. If so, the symphyseal fangs are no larger than the marginal teeth, a situation similar to that in Tersomius (Carroll 1964, fig. 8). Lingual to the marginal teeth is a short shelf extending up to the coronoid region. This region is largely covered by a shagreen of minute denticles commencing just lateral to the symphyseal thickening and ending about 5mm anterior of the adductor fossa. The denticle-bearing strip is about 12 denticles wide. Sutural borders of the anterior and middle coronoids are completely obscured by the denticles but the posterior margin of the third coronoid extends behind the denticles and forms the anterobuccal margin of the adductor fossa, contacting the dentary and surangular.
On the ventromedial side of the mandible, slivers of bone representing the medial edges of the splenial and postsplenial are visible. They appear to be of orthodox temnospondyl configuration. Behind the postsplenial is a more extensively exposed prearticular. Anteriorly, this bone narrows to a point wedged between the postsplenial and the coronoid series. Posteriorly, it broadens out to form the posterolingual face of the mandible and the medial border of the adductor fossa. The buccal border of the adductor fossa is formed by the surangular which is wedged between the dentary and the coronoid anteriorly and extends back to border the articular laterally. The lingual border of the adductor fossa is formed by the prearticular extending back to border the articular medially, as in Tersomius (Carroll 1964, fig. 8). There is no medial inflection of the prearticular as in the Permian trematopids described by Dilkes (Reference Dilkes1990). At the posterior end of the mandible, the articular is visible as a relatively large articular surface of ‘unfinished' bone. As in other dissorophoids, the articular surface is anteroposteriorly deep and consists of a two-lobed structure, there being a smaller buccal lobe and a larger lingual lobe separated by a ‘waist'.
5. Systematic discussion
5.1. The family Trematopidae
The family Trematopidae was proposed (as the Trematopsidae) by Williston (Reference Williston1910) for Trematops milleri and, tentatively, Acheloma cumminsi from the Lower Permian of Texas. Until the 1990s, the prevailing view of the family was based on the major review of the Texas and Oklahoma material published by Olson (Reference Olson1941), who described the further species Trematops willistoni, Acheloma pricei and Acheloma whitei.
Subsequently, further taxa were added to the Trematopidae, including Trematopsis seltini Olson Reference Olson1956, Ecolsonia cutlerensis Vaughn Reference Vaughn1969, Trematops stonei Olson Reference Olson1970, Actiobates peabodyi Eaton Reference Eaton1973, Anconastes vesperus Berman et al. Reference Berman, Reisz and Eberth1987, Tambachia trogallas Sumida et al. Reference Sumida, Berman and Martens1998, Mordex laticeps Milner & Sequeira Reference Milner and Sequeira2003, Fedexia striegeli Berman et al. Reference Berman, Henrici, Brezinski and Kollar2010, Rotaryus gothae Berman et al. Reference Berman, Henrici, Martens, Sumida and Anderson2011 and Acheloma dunni Polley & Reisz Reference Polley and Reisz2011. Of these forms, Trematopsis seltini has subsequently been shown to be a poor specimen of the dissorophid Cacops (Milner Reference Milner1985). Subsequently, Dilkes (Reference Dilkes1990) has revised the small trematopid material from the earlier Texas red-bed assemblages and placed it all in the new genus Phonerpeton. In the following discussion, the genus Acheloma is taken to comprise the species cumminsi (including milleri, thomasi and willistoni) and dunni, and Phonerpeton is based on the species pricei and whitei.
This content of the family is largely uncontroversial and is as used by Polley & Reisz (Reference Polley and Reisz2011) and Schoch & Milner (Reference Schoch, Milner and Sues2014). However, there have been doubts raised about the status of Actiobates and Ecolsonia within the family and it is useful to discuss these before going on to consider the relationships of Mordex and Mattauschia.
5.2. The status of Actiobates and Ecolsonia
Actiobates peabodyi Eaton is known from a single specimen (KUVP 17941) from the Upper Pennsylvanian Stanton Limestone from near Garnett in Kansas (Fig. 8). It comprises the anterior half of a slightly disarticulated skeleton exposed in ventral aspect but with the left side of the dorsal surface of the skull prepared out. Eaton (Reference Eaton1973) described it as having both dissorophid and trematopid characteristics and used it, together with Ecolsonia (then known only from a single poor specimen) and Longiscitula (reassigned to Dissorophidae by Milner Reference Milner2003a), as a basis for reducing the family Trematopidae in synonymy with the family Dissorophidae. However, Berman et al. (Reference Berman, Reisz and Fracasso1981) and Berman et al. (Reference Berman, Reisz and Eberth1985) noted it as a probable or certain trematopid, and Milner (Reference Milner1985) identified it as a primitive trematopid having a mixture of trematopid features and generalised dissorophoid features. Berman et al. (Reference Berman, Reisz and Eberth1987) argued that it was the most primitive trematopid, and that the dissorophid-like proportions of the skull and some of the individual elements were attributable either to the primitive position of this genus within the family, or to the subadult status of the type specimen. Subsequently, most authors have followed this conclusion although the type and only specimen requires redescription.

Figure 8 Actiobates peabodyi Eaton, holotype KUVP 17941. Stanton Limestone, Upper Pennsylvanian; Garnett, Kansas. Photograph of specimen in ventral aspect with inset showing proximal end of stapes with perforation (STA). Scale bars = 10mm.
One alternative opinion can be dealt with here. Boy (Reference Boy1981) proposed a set of relationships for the Dissorophoidea in which Actiobates is the sister taxon of a clade comprising the Trematopidae, Dissorophidae (excluding Amphibamus) and Doleserpetontidae. In Boy's scheme, Actiobates shares with them the frontal entering the orbit margin, but unlike them it primitively retained a stapes perforated by a foramen for the stapedial artery. The original specimen has never been illustrated until now so I include a photograph of the palate with an inset of the perforated stapes (Fig. 8). The two stapes of Permian trematopids known at that time both lacked a visible stapedial foramen, as did the stapes of the Permian dissorophid genera Broiliellus and Dissorophus and the amphibamid Tersomius. The several synapomorphies of Actiobates with the Trematopidae outnumber this retained primitive feature and if all the described morphologies are genuine, the loss of the stapedial foramen might be regarded as a convergent feature of dissorophids, amphibamids and trematopids. However at the time that Boy produced this argument, the preservation of the known Permian specimens was not good and it was not clear that the stapedial foramen was really absent in these forms. Bolt & Lombard (Reference Bolt and Lombard1985, p. 86) noted that the foramen is never absent in well-preserved temnospondyl material. Foraminate stapes have subsequently been reported in the trematopids Anconastes (Berman et al. Reference Berman, Reisz and Eberth1987, p. 260), Fedexia (Berman et al. Reference Berman, Henrici, Brezinski and Kollar2010, fig. 8b), Acheloma dunni (Polley & Reisz Reference Polley and Reisz2011, fig. 8) and Rotaryus (Berman et al. Reference Berman, Henrici, Martens, Sumida and Anderson2011, fig. 2b) and the amphibamid Tersomius (Maddin et al. Reference Maddin, Fröbisch, Evans and Milner2013, fig. 2c). Thus the presence of a stapedial foramen is entirely compatible with a position within the Trematopidae. In conclusion, Boy's evidence for placing Actiobates outside the dissorophid–trematopid group is insufficient. Likewise, Actiobates is not a trematopid–dissorophid intermediate, but simply a small primitive trematopid with some generalised dissorophoid features.
Berman et al. (Reference Berman, Reisz and Eberth1985, pp. 29–30) argued that Ecolsonia was a primitive offshoot of the Dissorophidae and that the enlarged external naris was a convergent similarity to that in the Trematopidae. In part, they formulated their argument by a three-taxon cladistic comparison using Ecolsonia, Trematops (now Acheloma and hereafter referred to as such) and Dissorophus, with the primitive temnospondyl Dendrerpeton as the outgroup. One problem with this approach is that comparison was made using some large, highly derived, terminal members of families, several of the character states of which were not necessarily those of the primitive members of the families they represented. Berman et al. (Reference Berman, Reisz and Eberth1985) discussed 15 characters of which five were used to associate the three genera as dissorophoids, three supported an Ecolsonia–Acheloma relationship and seven supported an Ecolsonia–Dissorophus relationship. The association of the three genera as dissorophoids is not controversial and the first five characters do not need to be discussed.
The three character states stated to be shared by Ecolsonia and Acheloma were as follows.
Character 6. ‘Ring intercentra' seems not to be valid as later study of Acheloma has failed to confirm the presence of ring intercentra in that genus (Dilkes & Reisz Reference Dilkes and Reisz1987, pp. 8–10).
Character 7. ‘Elongate, partially divided external naris' is valid and is used here as part of the diagnosis of an expanded Trematopidae.
Character 8. ‘Absence of typical dissorophid dermal armour' is the primitive temnospondyl condition and cannot be used as a derived character. The presence of dorsal armour is here taken to be a derived character defining the Dissorophidae. Its absence defines nothing.
The seven characters used to support an Ecolsonia–Dissorophus relationship were as follows.
Character 9. ‘Box-like configuration of the skull roof, with abrupt right-angle union between the cheek and skull table'. I suggest that this character is a manifestation of the presence of the large tympanic embayments on either side of the skull and is related to the requirement for such large tympanic openings to be suitably braced. The small slit-like embayments and associated skull shape of advanced trematopids are unique to that group and not the primitive temnospondyl condition. There is no reason why the trematopid condition should not be derived from the dissorophid skull shape, which would then have to be considered as the stem-dissorophoid skull shape. Large trematopids do possess a thickened curved postorbital and forms such as Fedexia and Phonerpeton have open embayments and steep-sided cheeks and are more dissorophid-like than Acheloma cumminsi from the Arroyo Formation. I will argue below that Mattauschia, Ecolsonia and Acheloma show a morphotypic series of progressively more specialised embayments and surrounding cheeks (Fig. 10).
Character 10. ‘Greatly variable development of skull-roof sculpturing' is too variable a character within the Dissorophidae for there to be any certainty that it is the primitive dissorophid condition. Although a feature of the large Dissorophus and Ecolsonia, other dissorophids show more uniform pitting (e.g., Broiliellus texensis FMNH UC684, Broiliellus olsoni UT-BEG 31189.8) and the variable sculpture patterns may be size linked and restricted to large derived forms. A variant of this character was also used by Daly (Reference Daly1994, p. 50) to include Ecolsonia in the Dissorophidae.
Character 11. ‘Short preorbital length of the skull' also occurs in some trematopids apart from the Arroyo material. Increase in snout length was later used by Berman et al. (Reference Berman, Reisz and Eberth1987, pp. 266–67) as a defining character for Acheloma (=Trematops) within the Trematopidae. They pointed out that the primitive trematopid Actiobates has dissorophid-like snout proportions, which would appear to negate the use of a short snout as a derived character relative to the trematopid condition.
Character 12. ‘Greatly expanded interpterygoid vacuities' is also a basic dissorophoid character that is found in the smaller and more primitive trematopids. The small interpterygoid vacuities of Acheloma (=Trematops) are not a primitive retention but have arisen within the Trematopidae as most trematopids including the more primitive forms, have large ‘dissorophid-like' interpterygoid vacuities (Berman et al. Reference Berman, Reisz and Eberth1987, p. 267).
Character 13. ‘Tabular horn bends sharply posteroventrad at about a right angle to the skull table to contact a vertically orientated posterodorsal process of quadrate' is a derived character of some late large dissorophids such as Dissorophus and Cacops but is not found in many other dissorophids which primitively retain an open embayment. Within the dissorophines (sensu Demar Reference Demar1966, Schoch & Milner Reference Schoch, Milner and Sues2014), the earlier and more primitive Broiliellus has simple armour and an open embayment whereas Dissorophus has more complex armour and a closed embayment. Within the eucacopines (also sensu Schoch & Milner Reference Schoch, Milner and Sues2014) the early Scapanops and Conjunctio have simple armour and an open embayment while the later Cacops has complex armour and a closed embayment. It appears that embayment closure by tabular and quadrate outgrowth has arisen independently twice within the armoured Dissorophidae and its presence in Ecolsonia must be a convergent feature.
Character 14. ‘Semilunar curvature of squamosal that narrows the tympanic embayment from above'. This character defines a wider group than the armoured Dissorophidae, as it not only occurs in Ecolsonia but also in the trematopid Phonerpeton (Dilkes Reference Dilkes1990) and the amphibamid Eoscopus (Daly Reference Daly1994) although it is not so well developed in the three later forms. Daly (Reference Daly1994) has argued it to be a general dissorophoid character subsequently lost within some subgroups.
Character 15. ‘Posterodorsal process of quadrate possesses an anterior expansion' is shared by Ecolsonia, Cacops and Dissorophus but as it is an outgrowth of an ossified posterodorsal quadrate process closing the tympanic embayment, it follows that it can only occur in dissorophoids with closed embayments. Therefore, as with Character 13, it must have arisen independently as the closed embayments arose independently.
In conclusion, of the characters listed by Berman et al. (Reference Berman, Reisz and Eberth1985), only character 7 is accepted as unambiguously valid supporting a trematopid position for Ecolsonia. Of the other characters, 6 is based on an earlier misinterpretation of trematopid vertebrae; 8 is a primitive temnospondyl character; 9 is probably a primitive dissorophoid character with the trematopid condition as the more derived condition; 10 is derived but probably a size-linked homoplasy which does not occur in all dissorophids; 11 and 12 are based on ‘primitive' features of Acheloma (=Trematops) which can be shown to be derived within the Trematopidae, with the implication that the Ecolsonia–Dissorophus condition is actually the primitive state; 13 and 15 are derived but homoplastic because they are associated with tympanic embayment closure which must have occurred independently within the dissorophines, aspidosaurines and Ecolsonia; 14 is probably a primitive dissorophoid character. Characters 10–15 illustrate the problems of cladistic analysis based on selected genera as representatives of families when those genera are highly derived members of those families. It appears that, contra Berman et al. (Reference Berman, Reisz and Eberth1985), Ecolsonia cannot be placed within a Dissorophidae defined on derived characters.
5.3. Ontogeny and relationships within the Trematopidae
The disparity of characteristics of the small and large individuals of the Nýřany trematopids must ultimately lead to a further consideration of the permutation of size and character states in other trematopid taxa. It appears that some features of the skull construction of large trematopids developed gradually after metamorphosis and that the small post-metamorphics were significantly different from the large ones. This needs to be discussed before consideration of the relationships of the Nýřany genera.
5.3.1. Closure of the tympanic embayment
Comparison of the midline skull lengths and otic embayment closure gives the following result. Note that Mordex, Fedexia, Actiobates, Anconastes, Tambachia and Rotaryus are all single-specimen genera.
The Pennsylvanian genera Mattauschia and Mordex are represented by small short-snouted juveniles (33–39mm skull) with an open squamosal embayment, and a large specimen (120mm skull) with an enclosed embayment. Fedexia is represented by a single incomplete 103mm skull with an open embayment to judge by the condition of the tabulars, Ecolsonia is represented by complete or partial large skulls in the 135–150mm range with closed embayments and highly comparable with the large Mattauschia. Actiobates has a 70mm skull with an open embayment and Anconastes has a 100mm skull with an embayment of uncertain status. The Permian taxa comprise Tambachia with a 65mm skull with open embayment, Rotaryus with a 48mm skull and open embayment, Phonerpeton consisting of skulls in the 60–90mm range with open embayments and Acheloma with 150–180mm skulls with closed embayments. In summary, skulls from 30–110mm midline length have open embayments, whereas skulls from 120–180mm in length have the embayments closed by a tabular–quadratojugal strut. Is this an unqualified taxonomic feature or an ontogenetic phenomenon in all trematopids?
In this work, skulls assigned to Mattauschia, include a small skull with an open embayment and a large one with a closed embayment. A later potential instance of this is Acheloma in the Arroyo of Texas where the large skulls are accompanied by smaller 50–70mm skulls (‘Trematops willistoni' of Olson; e.g., FMNH UC 1584, MCZ 2475) with very narrow notches combined with open embayments. The former was not discussed by Dilkes (Reference Dilkes1990) and the latter was simply attributed to Phonerpeton sp. and they have been neglected in all recent literature. They have the massive horizontal cheek region of Acheloma but the otic embayment is not closed at the back. They are consistent with being half-grown post-metamorphics of Acheloma cumminsi. A third possible example of the problem is Phonerpeton pricei, described by Dilkes (Reference Dilkes1990) as a small trematopid with open otic embayments, from the Archer City to Petrolia Formations of Texas. However in the collections at the Field Museum, Chicago, is FMNH UC 481 – a 150-mm skull from the Archer City Formation which demonstrates that either a second large form lived alongside Phonerpeton, or that at least some of the Phonerpetons are juveniles of an earlier Acheloma species where the flattening of the otic embayment was less highly developed. This specimen is not mentioned by Dilkes (Reference Dilkes1990).
The possibility must be reconsidered that all trematopid genera grew to 150mm or more in skull length but that tympanic embayment closure only took place at 110–120mm midline skull size. If this is so, the types of Mordex, Actiobates, Anconastes, Tambachia and Phonerpeton may all be immature and not ontogenetically equivalent to the larger specimens of Mattauschia, Fedexia, Ecolsonia and Acheloma.
5.3.2. Nostril shape
Nostril shape must almost certainly change with ontogeny, at least in the more derived trematopids. Dilkes (Reference Dilkes1993) figured a small snout (MCZ 2531) from the Lower Permian of Texas in which the external naris is a simple oval like those of Mattauschia and Fedexia. Dilkes interpreted this as a juvenile with an estimated skull length of 53mm. This is the most logical interpretation, although the possibility exists that this is a late-surviving relic of the Mattauschia/Fedexia snout morphology. Phonerpeton skulls of 60–70mm figured by Bolt (Reference Bolt1974a fig. 1) and Dilkes (Reference Dilkes1990 figs 3, 4) show a fully elongate bilobed naris implying either a very rapid morphological change during growth or the existence of a mixture of juvenile and dwarf species in this size range. This might explain the confusing mixture of snout shapes and nostril shapes combined within Phonerpeton. It may explain the variable position of Phonerpeton in relation to Acheloma in the cladograms of Polley & Reisz (Reference Polley and Reisz2011) and Berman et al. (Reference Berman, Henrici, Martens, Sumida and Anderson2011).
5.3.3. Snout shape and length
Finally, there is the issue of whether the smaller skulls can be determined as growing into Acheloma-shaped adults or Ecolsonia-shaped adults. An example of this problem may be the status of Ecolsonia and Anconastes both from the Cutler Formation of north-central New Mexico. They were described as an aberrant dissorophid and a trematopid respectively but in some recent analyses they are apparently different grades of trematopid. However, as noted by Sumida et al. (Reference Sumida, Berman and Martens1998), Anconastes, because incomplete, possesses only two of the six characters that they used to define the Trematopidae and both of these (external naris and pseudocaniniform teeth) are present in Ecolsonia. As Anconastes and Ecolsonia also share a peculiar mosaic covering of dorsal osteoderms, not reported in any other trematopids, it is possible that Anconastes is no more than a half-grown Ecolsonia. They have never been directly compared. The German trematopid Tambachia shares three characters with Anconastes (Sumida et al. Reference Sumida, Berman and Martens1998) namely absence of internarial fenestra (also in Ecolsonia), suborbital process of lacrimal reduced also (in Ecolsonia) and no palatine dorsal exposure but maxilla entering orbit margin (Ecolsonia does have a palatine exposure excluding the maxilla from the orbit margin but this could be size linked). It is perfectly possible that a large Tambachia would be more Ecolsonia-like than Acheloma-like.
5.3.4. Summary
Clearly a simple cladistic analysis of a series of genera represented partly by juveniles and partly by adults could give flawed results if this is not taken into account. Further investigation is beyond the scope of this paper and no taxonomic changes are proposed here but this problem merits a further focussed study. If such late ontogenetic changes do take place within trematopid species, any character analysis that does not take account of this is likely to be highly compromised. In the following discussion, the relationships of Mattauschia are discussed solely in relation to equally large trematopids (midline skull length 120mm or more) which are unambiguously adult, whereas the relationships of Mordex are simply discussed with extreme caution.
5.4. Relationships of Mordex and Mattauschia
Mordex and Mattauschia both belong to the Dissorophoidea because they possess the following derived characters: (1) a reduced jugal extending forwards to a point at the level of the middle of the orbit, thus permitting the maxillary and palatine to enter the orbit margin in large skulls, (2) a large tympanic embayment extending from quadrate to tabular resulting in extreme concavity of the posterior edge of the squamosal, (3) a prominent transverse flange on the posterodistal edge of the pterygoid. A feature shared by Mattauschia with comparably large dissorophids and trematopids is the posterior closure of the tympanic region by extensions of the tabular and the quadratojugal.
5.4.1. Mattauschia
The relationships of Mattauschia will be considered first. It is compared only with genera with comparably large skulls, namely Fedexia, Ecolsonia and Acheloma, in order to avoid the complication of ontogenetic factors (Fig. 9a). The character states involved are sufficiently few that a ‘hand-worked' presentation seems sufficient to the author.

Figure 9 (a) A hypothesis of relationships of four trematopid genera. Character states defining nodes are: Node 1 (Trematopidae) small v-shaped lacrimal and enlarged external naris, flattened tympanic embayment and large quadratojugal; Node 2 reduced prefrontal and postfrontal permit frontal to enter orbit margin, reduction to 30–40 large marginal teeth with caniniforms on premaxilla and maxilla; Node 3 external naris divided into anterior and posterior lobes by nasal flange, tympanic embayment further flattened; Taxon 4 (Acheloma) elongate straight-sided snout, internarial fenestra, even fine dermal ornament. (b) Cladogram with Mordex inserted, its small size reflecting its juvenile status. Mordex shares naris shape with Node 3 and fine ornament with Acheloma, both contra Fedexia. However, it lacks the Node 2 characters that Fedexia shares with Ecolsonia and Acheloma.
Mattauschia is a trematopid because it possesses (1) the enlarged external naris reaching almost to orbit with the dermal portion of lacrimal small and triangular, and the prefrontal entering the naris margin, (2) an unornamented supratympanic flange of the tympanic embayment with a slender squamosal component and a suture indicative of a significant supratemporal component (in the isolated squamosal), the remaining characters of the flange being obscured, (3) a tympanic embayment with a ventral border sloping at less than 45° in large individuals so that at least the anterior region of embayment tends to a horizontal slit, this is combined with an enlarged deep quadratojugal (this is not seen in Fedexia which has a deep embayment) (Fig. 10).

Figure 10 Tympanic embayments of trematopids and Cacops in lateral aspect. Quadratojugal shaded. (a) Mattauschia (based on Fig. 10). (b) Ecolsonia (based on Berman et al. Reference Berman, Reisz and Eberth1985, fig. 8c). (c) Anconastes (based on Berman et al. Reference Berman, Reisz and Eberth1987, fig. 8), degree of closure of tympanic embayment is not known; (d) Acheloma (based on Dilkes & Reisz Reference Dilkes and Reisz1987, fig. 3); (e) Cacops (based on Williston Reference Williston1910, pl. 8, sutures after Fröbisch & Reisz Reference Fröbisch and Reisz2012 fig. 2).
Mattauschia is more primitive than any later trematopid because (1) it retains a prefrontal–postfrontal contact, (2) it has a marginal dentition of 75 small teeth per ramus as opposed to 30–40 large teeth per ramus, (3) it lacks the inflexion of the prearticular on the mandible (situation unknown in Fedexia). Given the visible pattern of dentition, it probably lacks the pseudocaniniform teeth on the premaxilla and the anterior maxilla, but this is not demonstrable.
Mattauschia and Fedexia are more primitive than later trematopids because (1) the external nares are large ovals not divided into anterior and posterior lobes by a lateral flange of the nasal. The nasals are long rectangular straight sided elements, (2) the tympanic embayment of is less flattened and trematopid-like than those of Ecolsonia and the Permian trematopids.
Mattauschia, Fedexia and Ecolsonia are more primitive than later trematopids such as Acheloma because (1) they lack the elongate straight-sided snout of later forms, (2) they lack the internarial fenestra (3) they lack the very fine dermal sculpture, (4) they lack the derived otic notch reduced to a slit.
Two features of uncertain significance are as follows. One is the very narrow distance between the orbit and the squamosal embayment in Mattauschia which separates it from all later trematopids but might be an autapomorphy. The other is the humerus of the small type specimen which lacks a supinator process, whereas more derived trematopids possess one. This might be a significant phylogenetic feature but could also be based on ontogenetic change with the supinator ossifying only in larger individuals.
5.4.2. Mordex
Mordex is a trematopid because it possesses (1) the enlarged external naris reaching almost to orbit with the dermal portion of lacrimal small and triangular extending a finger of bone along the lateral margin of the naris, together with the prefrontal entering the naris margin, (2) an unsculptured supratympanic flange of the tympanic embayment with a slender squamosal component, a semilunar flange of the supratemporal and a tabular process which contacts the squamosal below the supratemporal, (3) the palatine ramus of the pterygoid extending forwards to reach the vomer and exclude the palatine from the margin of the interpterygoid vacuity. A character of uncertain significance found in some but not all trematopids is the relatively massive premaxilla.
Mordex is more primitive than any later trematopid because, like Mattauschia, (1) it retains a prefrontal–postfrontal contact, (2) it has a marginal dentition of small teeth per ramus as opposed to fewer large teeth per ramus. The status of the inflexion of the prearticular on the mandible is unknown. Given the visible pattern of dentition, it probably lacks the pseudocaniniform teeth on the premaxilla and the anterior maxilla, but this is not demonstrable.
At this point, there is a character conflict between Mordex and Fedexia (Fig. 9b). Mordex is more primitive for the reasons listed above, but more derived than Mattauschia in that it possesses a nasal with a flange dividing the external naris into two lobes, and it also possesses very fine dermal sculpture like most later trematopids. Mordex also shows the very narrow distance between the orbit and the squamosal embayment like Mattauschia and a humerus which lacks a supinator process. Given that the reduction in size of the circumorbital series resulting in loss of the prefrontal–postfrontal contact occurs repeatedly in the Dissorophoidea (within Branchiosauridae, Micromelerpetontidae and Amphibamidae), it is suggested that this is not a character of high uniqueness and that Mordex might be closer to higher trematopids than is Fedexia. The ambiguity of this situation is reflected in Figure 9b.
5.4.3. General conclusions
It seems clear that these two genera are the most primitive trematopids, they have only a few of the character states that genera such as Phonerpeton and Acheloma share, but character acquisition is sequential so this is to be expected in early more primitive forms. My conclusion is that Mattauschia is certainly the most basal trematopid yet described, while Mordex might be the most primitive relative of the main trematopid clade from Ecolsonia crownwards based on the possession of the characteristic external naris and fine dermal sculpture.
6. Chronology and evolution of the Dissorophoidea and Trematopidae
6.1. Dissorophoidea
Representatives of most dissorophoid families appear simultaneously in the fossil record in the Asturian/late Middle Pennsylvanian of Nýřany, Mazon Creek and Linton. The contemporaries of Mordex and Mattauschia (trematopids) include Platyrhinops and Amphibamus (derived amphibamids), Limnogyrinus (micromelerpetontid) and Branchiosaurus (branchiosaurid) and these are sufficiently distinct to suggest that the dichotomies producing these four families had occurred significantly earlier and hence that recognisable trematopids and other dissorophoids will ultimately be found in Langsettian–Bolsovian (=‘Westphalian A-C') beds and possibly even earlier. There are very few records of pre-Asturian temnospondyls and these are demonstrably more primitive forms with one possible exception. Milner (Reference Milner1982b) described a poorly preserved small skull from the Langsettian of Parrsboro, Nova Scotia as a small Dendrerpeton. This specimen was collected from a lycopsid trunk and is likely to have been a relatively terrestrial form. The 1982 interpretation was a conservative one mainly intended to counter the view that the specimen was a Eugyrinus-like form (Romer Reference Romer1966) and hence a dvinosaurian. I interpreted the specimen as having an intertemporal and hence a dendrerpetontid but the preservation is sufficiently poor that the possibility exists that it is an early dissorophoid. Two features that support this are the incipient pterygoid flange and the deep striate–radiate dermal sculpture very reminiscent of that of Stegops divaricata – a dissorophoid of uncertain family relationships found at Linton. Because the specimen is poorly preserved, its relationships are likely to remain uncertain but it does suggest the possibility that further lycopsid trunk finds might include early dissorophoids.
The large Mattauschia specimen also demonstrates that large dissorophoids with closed tympanic embayments were already present in the Asturian, and hence that such closed embayments may characterise large dissorophoids throughout their stratigraphical range, and are not just characteristic of the later Lower Permian forms.
6.2. Trematopidae
In evolutionary terms, the internal relationships of the Trematopidae proposed here suggest that trematopids originated as a distinct lineage prior to the Asturian and may have originated as relatively large dissorophoid temnospondyls. The characteristic naris and tendency to a slit-like tympanic embayment in large individuals were acquired early in their evolution, but much of their subsequent evolution appears to relate to a shift in diet from smaller, possibly arthropod, prey to larger, possibly vertebrate, prey. Mordex and Mattauschia retained a generalised dissorophoid skull shape and had many small marginal teeth and could perhaps be envisaged as terrestrial predators on smaller prey such as arthropods which required gripping and crushing. By the Late Pennsylvanian/basal Permian, forms such as Fedexia, Ecolsonia, Actiobates and Anconastes had larger piercing teeth including highly differentiated premaxillary and maxillary teeth consistent with an important holding and piercing function. This suggests a more robust prey that required impaling and may represent a shift to larger vertebrate prey. In the more derived Permian trematopids, the snout is elongated and in the terminal form Acheloma, the snout tip became massive and the anterior palatal bones were built up medially, reducing the interpterygoid vacuities. This suggests a greater capacity to resist forces generated by prey held in the front of the mouth.
The sequence of progressively more derived trematopid tympanic embayments (Fig. 10a–d) shows a trend from a large laterally situated embayment with some incipient elongation to a highly derived slit-like embayment in an elevated position on the cheek. Dilkes & Reisz (Reference Dilkes and Reisz1987, p. 11) suggested that the derived embayment must have borne a long shallow tympanum which was only capable of receiving low-frequency sound. If so, this suggests that the perception of high-frequency air-borne sound had become unimportant to the derived Trematopidae. This appears to conflict with terrestrial adaptations such as the absence of lateral-line sulci, a body covering of osteoderms and well-ossified carpals and tarsals, all found in later forms. A possible life-style might be that of a terrestrial lurking predator on other smaller vertebrates, a niche filled by toads such as Bufo and Ceratophrys today. The general terrestrial adaptations, the dentition adapted for holding and subduing vertebrates, and the ears capable of perceiving ground-borne low-frequency vibrations such as vertebrate footfalls, are most consistent with such a life-style. The evolution of trematopids would thus be the modification of a terrestrial arthropodivore into a terrestrial predator on small vertebrates.
7. The Nýřany dissorophoid larva problem
The 800 or more tetrapod specimens in the Nýřany assemblage include at least 150 tiny larval or paedomorphic dissorophoids (skull length 12mm or less). Fritsch (Reference Fritsch1879) described a selection of these as Branchiosaurus salamandroides. Boy (Reference Boy1972) demonstrated that Permian ‘branchiosaur' assemblages included a distinct form Micromelerpeton, and was able to argue that related forms (as the family Micromelerpetontidae) were found in many ‘branchiosaur' assemblages. In a short note, Milner (Reference Milner and Roček1986) identified the Nýřany micromelerpetontid as Limnogyrinus elegans. Werneburg (Reference Werneburg1994, Reference Werneburg2012) provided full descriptions of this taxon, which is readily separated from other small dissorophoids. Milner & Sequeira (Reference Milner and Sequeira2003) also described some specimens as part of their evaluation of the problematic genus Limnerpeton.
The remaining material was redescribed as Branchiosaurus salamandroides by Werneburg (Reference Werneburg1987), but Milner (Reference Milner2003b, p. 112, 2007) and Milner & Sequeira (Reference Milner and Sequeira2003, p. 140) suggested that this material included larval amphibamids and trematopids as well as Branchiosaurus sensu stricto. The extensive work to elaborate on this suggestion progressed slowly and was overtaken by a taxonomic analysis by Werneburg (Reference Werneburg2012). Werneburg segregated the non-Limnogyrinus material into four taxa, Branchiosaurus salamandroides, larvae of the amphibamid Platyrhinops fritschi, larvae of the trematopid Mordex laticeps and a new small form Nyranerpeton amilneri. While agreeing with the general result of Werneburg's work, I find many of his individual assignments highly problematic, and feel that further analysis will be required particularly as there are now two trematopids to be considered.
Before discussing identification of larvae, a number of points need to be made. Firstly, it must be emphasised that this is difficult material with many crushed specimens possessing skulls in the 5–10mm range, and that a significant proportion of these larvae must remain as Dissorophoidea incertae sedis. Many show one or two potentially useful character states but are indeterminate for most others. As a consequence, they can be partially categorised but not fully. Secondly, there is the problem of identifying taxonomically useful characters that might persist unaltered from larva to adult. The shape of the skull table, the short gap between the orbit and otic embayment, the length of the presacral column and the degree of elongation of the humerus and the manual phalanges appear to be characters that define animals at all sizes. A third point is that the material can be quite readily separated into micromelerpetontid larvae (Limnogyrinus (Fig. 11a) and probably Nyranerpeton) in contrast to larvae attributable to Xerodromes (Schoch & Milner Reference Schoch, Milner and Sues2014), which are the more derived dissorophoids including Amphibamidae, Trematopidae, Dissorophidae and Branchiosauridae. The problem is discriminating among the Xerodromes larvae and small adults. Although we have no transported material identified as armoured Dissorophidae, such taxa may well have been present and represented by larvae, if not by adults. The larger amphibamids and trematopids are sufficiently rare that the absence of recognised dissorophids cannot be taken as profoundly significant. Also, we cannot assume that Branchiosaurus is represented only by small paedomorphic individuals. It has been demonstrated that Apateon gracilis from Niederhässlich metamorphoses at about 18mm skull length into a terrestrial adult, unlike most other species (Werneburg Reference Werneburg1991; Schoch & Fröbisch Reference Schoch and Fröbisch2006). Branchiosaurus salamandroides is the most primitive branchiosaurid (Schoch & Milner Reference Schoch and Milner2008) but no specimen has been described with skulls larger than 13mm, so it is possible that metamorphosed Branchiosaurus lurk among the larger specimens. Thus we have to consider hypothetical larval dissorophids and adult branchiosaurids as well as small branchiosaurids, an amphibamid and two types of trematopid.

Figure 11 Small and larval dissorophoids found in the Nýřany assemblage. (a) the micromelerpetontid Limnogyrinus elegans. (b) The branchiosaurid Branchiosaurus salamandroides. (c) The olsoniform larval morphotype, probably trematopid. (a–c) Are reconstructions in dorsal aspect based on multiple specimens. (d) The amphibamid Platyrhinops fritschi, an interpretive drawing of the only specimen – NMP M340, which is incomplete and insufficient for a full reconstruction. Scale bar = 1mm.
Branchiosaurus salamandroides (Fig. 11b) can be most readily separated out by postcranial characters, namely a reduced presacral column of 21–22 vertebrae combined with elongate humerus and femur and manual phalanges that are shorter than the humerus. The humerus+femur: skull midline ratio is 88–96%. Trunk ribs 3–10 are substantially more robust than posterior trunk ribs. No specimen shows evidence of a palatal dentition and there is no retroarticular process on the mandible. In Werneburg's (Reference Werneburg2012) work and my unpublished study, about a third of the potentially determinate small Xerodromes larvae correspond to these features, including, fortunately, the lectotype specimen (NMP M332).
The amphibamid Platyrhinops fritschi is represented by a single incomplete determinate larva (NMP M340) in my opinion (Fig. 11d). The presacral column is incomplete but the humerus is elongate and the longest manual phalanges are longer than the humerus. The remarkably large manus is a characteristic of Platyrhinops and is present here in a larva with a 7mm skull. The specimen shows a short retroarticular process, a cultriform process stouter than that of Branchiosaurus, and the rib differentiation is less extreme. As recorded by Werneburg (Reference Werneburg2012), it has ossified hyobranchial elements and single-spike branchial denticles like Branchiosaurus, demonstrating that it is a larva and not just a small juvenile of Platyrhinops. Werneburg assigned ten other specimens to P. fritschi (apart from the adult holotype) but in my unpublished study, they were all determined as Mordex laticeps before it became clear that two trematopids were present.
The remaining material which Werneburg described as Mordex laticeps and most of his Platyrhinops fritschi, can best be categorised as Olsoniformes incertae sedis (Fig. 11c). They are characterised by a presacral column of 23–24 vertebrae, shorter more robust humerus and femur, with manual phalanges no longer than the humerus. The humerus + femur midline skull length is 70–83%. Where the palate is visible, palatal denticles are present on vomers, palatines and anterior pterygoids. Where preservation permits, it can be seen that the orbit and the squamosal embayment are separated by a very narrow distance as in the large Mordex and Mattauschia specimens. Almost two-thirds of the Nýřany Xerodromes larvae correspond to this general morphology but I cannot identify any character that would enable me to identify individuals as Mordex larvae, Mattauschia larvae or dissorophid larvae. Pending further study, I suggest that these be treated as indeterminate olsoniform larvae. They demonstrate that at least one species of large olsoniform dissorophoid was breeding in the Nýřany water body with individuals present in larger numbers than Branchiosaurus or Limnogyrinus.
8. Palaeobiogeography and palaeoenvironments of the Trematopidae
The presence of Mordex and Mattauschia in the Asturian (=late Middle Pennsylvanian) of Europe, together with Actiobates, Fedexia and Anconastes in the Upper Pennsylvanian of North America, demonstrates that the primitive Trematopidae were widespread throughout southern Euramerica from the late Carboniferous, despite being known from only a few occurrences. The advanced Trematopidae are well represented in the Lower Permian (up to Artinskian) of North America (Texas, New Mexico, Oklahoma, and Ohio) and Europe (Germany) and so occupied at least the same geographic area during the Early Permian. No trematopid has been recorded from post-Artinskian horizons. The presence of primitive and advanced trematopids across the same range indicates some degree of dispersal, rather than developing endemism. In the cladogram of Berman et al. (Reference Berman, Henrici, Martens, Sumida and Anderson2011), the European Early Permian Rotaryus is the sister taxon of the North American Early Permian Phonerpeton + Acheloma, indicating that these have a common origin more derived than the European Mattauschia or the North American Fedexia.
Mordex and Mattauschia are unusual in being the only trematopids recognised in a coal-lake assemblage. Their recognition is a reminder that, although much of the Nýřany assemblage is autochthonous and derived from a pool/channel ecosystem, the richness of the fauna is due to the presence of erratics from other neighbouring ecosystems (Milner Reference Milner and Panchen1980). The three larger specimens of Mattauschia may be presumed to represent occasional drowned individuals of a species from a neighbouring terrestrial habitat. If one accepts that many of the small dissorophoids at Nýřany are trematopid larvae, this implies that the trematopids were using the water body as a breeding pool. This might be the explanation for the occasional large specimen in the Nýřany assemblage, namely that they died during breeding activities, rather than being the result of random accidents.
Other early trematopids also appear to have been unusually terrestrial. Actiobates from the Late Pennsylvanian Garnett Formation of Kansas is the only amphibian associated with an assemblage of seven amniotes (synapsids and a diapsid) and found in stream channel infills in tidal mudflats. The exact horizon of Fedexia within the Casselman Formation of Pennsylvania is uncertain but probably a freshwater limestone derived from a drying ephemeral lake in a period of pronounced seasonality (Berman et al. Reference Berman, Henrici, Brezinski and Kollar2010). Ecolsonia was argued to be terrestrial by Berman et al. (Reference Berman, Reisz and Eberth1985) using morphological criteria. The horizons in the Cutler Formation which produced the specimens were a channel fill (VanderHoof quarry – holotype) and a sandy shale in a crevasse-splay deposit (Morfin bonebed – referred material) and appear to have been formed in an environment of alternating seasonal wetness and aridity. This may have been a more typical environment for early trematopids. It seems likely then that Mordex and Mattauschia were terrestrial forms living in dryer areas of the Pilsen Basin and that their presence in the Nýřany assemblage was indeed accidental. It is also a reminder that however terrestrial these amphibians had become, they were still tied to water for reproduction and may have needed to migrate and aggregate to breed as many of their descendants still do.
9. Acknowledgements
This paper is dedicated to Jenny Clack, a colleague and dear friend for over 40 years. Whereas most of us are content to have steadily advanced the study of early tetrapods, Jenny has taken one of the most fundamental transformations in vertebrate evolution and revolutionised our understanding of it. I am also particularly grateful to her for finding the time to rescue some of my unfinished work from oblivion, a true friend indeed.
For permission to study material in the collections in their care, I am indebted to Angela C. Milner (Natural History Museum, London), Heinz A. Kollman (Naturhistorisches Museum, Wien), Vojtek Turek, Milada Maňourová and Boris Ekrt (National Museum, Prague), John R. Bolt (Field Museum of Natural History), Larry Martin, John Chorn and Hans-Peter Schultze (University of Kansas), and Dave Berman and Amy Henrici (Carnegie Museum). My thanks also go to Dave Berman for freely communicating results of his research prior to publication, and to John Bolt, Larry Martin, John Chorn, Dave Berman and Amy Henrici for their hospitality during my visits to North America. Particular thanks go to my wife Angela for undertaking the acid etching, conservation and casting of the Nýřany specimens, to Sandy Sequeira for assistance with photography and preparation of figures, to Jenny Clack for constructive criticism of an earlier draft of the manuscript and to Florian Witzmann and Rainer Schoch for refereeing and improving the final submission. Study trips were supported by the University of London Central Research Fund (Vienna 1983), the Natural Environment Research Council (grant GR3/7491 – Prague 1993), the Royal Society of London (North America 1995) and the Leverhulme Trust (award F/07112/B – Vienna and Prague 2001).











