Sesame has been cultivated for centuries for its seed, which is used to produce high-quality cooking oil or as a complementary ingredient in confectionary goods (Nayar and Mehra Reference Nayar and Mehra1970). Historically, sesame production has predominantly occurred in Asia using dehiscent varieties that easily shatter when disturbed, and thus require delicate hand-harvesting to minimize seed loss. However, in the United States during the 1950s, nondehiscent varieties were developed to facilitate mechanical harvest (Langham et al. Reference Langham, Wiemers, Janick and Whipkey2002). Since this improvement, sesame production has expanded in the United States. Nevertheless, published literature on sesame production in mechanized systems is limited, making it difficult to develop science-based recommendations for producers.
The adoption of conservation tillage in sesame systems has been utilized to improve soil moisture retention and lower input costs, resulting in higher net returns (Keeling and Segarra et al. Reference Keeling, Segarra and Abernathy1989). However, with the elimination of primary tillage early-season weed control is reliant entirely on herbicides. Currently, S-metolachlor is the only herbicide registered for broadleaf weed control in sesame. Because S-metolachlor cannot control emerged seedlings, planting into a stale seedbed is essential. Therefore, an effective preplant burndown application is necessary to provide weed control prior to S-metolachlor application.
Preplant burndown herbicide applications usually consist of nonselective broad-spectrum herbicides such as paraquat and glyphosate, which adequately control most winter annual weed species (Shaw Reference Shaw1996). Glyphosate and paraquat are commonly applied for preplant burndown in conservation tillage systems because they effectively control a wide range of species, are relatively inexpensive, and can be applied close to planting due to their lack of soil activity (Brown and Whitwell Reference Brown and Whitwell1985; White and Worsham Reference White and Worsham1990). Unfortunately, wild radish (Raphanus raphanistrum L.) and cutleaf evening-primrose (Oenothera laciniata Hill) are not sufficiently controlled by glyphosate or paraquat (Culpepper et al. Reference Culpepper, Carlson and York2005). Though these weeds are winter annuals, failure to control them prior to planting results in early-season competition that commonly extends well into the summer (Webster and MacDonald Reference Webster and MacDonald2001; Webster and Nichols Reference Webster and Nichols2012). Aside from these two species, horseweed [Conyza canadensis (L.) Cronq.] and volunteer peanut (Arachis hypogaea L.) often escape control with paraquat and glyphosate as well (Bruce and Kells Reference Bruce and Kells1990; Johnson et al. Reference Johnson, Baldwin and Mullinix2000; Webster Reference Webster2013).
Glyphosate and paraquat provide 42% to 60% and 24% to 56% control of cutleaf evening-primrose, respectively (Culpepper et al. Reference Culpepper, Carlson and York2005; Reynolds et al. Reference Reynolds, Crawford and Jordan2000). Likewise, glyphosate and paraquat typically provide approximately 80% control of wild radish (Culpepper et al. Reference Culpepper, Carlson and York2005; Schroeder Reference Schroeder1989). Additionally, glyphosate and paraquat commonly only provide approximately 60% and 25% control of horseweed, respectively (Eubank et al. Reference Eubank, Poston, Nandula, Koger, Shaw and Reynolds2008; Keeling and Henniger et al. Reference Keeling, Henniger and Abernathy1989; Wilson and Worsham Reference Wilson and Worsham1988). Therefore, 2,4-D and dicamba are common tank-mix options to broaden the spectrum of glyphosate or paraquat mixtures. The addition of 2,4-D (0.56 kg ha−1) or dicamba (0.56 kg ha−1) to glyphosate or paraquat has been shown to increase cutleaf evening-primrose and wild radish control to >90% (Culpepper et al. Reference Culpepper, Carlson and York2005; Reynolds et al. Reference Reynolds, Crawford and Jordan2000; Schroeder Reference Schroeder1989). Horseweed control is also improved to >93% when 2,4-D or dicamba is added to glyphosate or paraquat (Eubank et al. Reference Eubank, Poston, Nandula, Koger, Shaw and Reynolds2008; Keeling and Henniger et al. Reference Keeling, Henniger and Abernathy1989; Wilson and Worsham Reference Wilson and Worsham1988).
Though neither 2,4-D nor dicamba are considered to have substantial PRE activity, they do possess enough soil activity to injure sensitive crops such as cotton (Gossypium hirsutum L), soybean [Glycine max (L.) Merr.], and rapeseed (Brassica napus L.) if planting is not sufficiently delayed (Moyer et al. Reference Moyer, Bergen and Schaalje1992; York et al. Reference York, Culpepper and Stewart2004). The need to delay planting following 2,4-D and dicamba applications has been well documented for cotton and soybean (Bruce and Kells Reference Bruce and Kells1990; Everitt and Keeling Reference Everitt and Keeling2007; Keeling and Henniger et al. Reference Keeling, Henniger and Abernathy1989; Krausz et al. Reference Krausz, Kapusta and Mathews1993; Thompson et al. Reference Thompson, Steckel, Ellis and Mueller2007; Wilson and Worsham Reference Wilson and Worsham1988; York et al. Reference York, Culpepper and Stewart2004). However, there are no data in the literature documenting sesame sensitivity to these herbicides or the nature of the sesame’s response to them. If sesame proves to be sensitive to soil residual activity of 2,4-D or dicamba, it is unknown whether the result will simply be stand reduction, or if plants will emerge but will be affected by stunting and altered reproductive physiology throughout the entire season. If stand reduction alone is the result, then it would be desirable to know the sesame population required before yield reduction occurs and replanting should be considered. However, research on this question is lacking. Therefore, the objectives of these studies were 1) to identify the impact of preplant application timing of 2,4-D and dicamba on sesame emergence, plant height, and yield, and 2) to determine the level of stand reduction that can be experienced without significant associated yield loss.
Materials and Methods
Two separate experiments (a preplant herbicide study and a population density study) were conducted at one location in 2015 and two locations in 2016 (29°24′10.37′′N, 82°10′16.60′′W and 29°24′24.59′′N, 82°8′43.89′′W) at the University of Florida’s Plant Science Research and Education Unit in Citra, FL. All experiments were conducted under conventional tillage, and hand-weeding was used to maintain weed-free conditions throughout the duration of the experiments. In both experiments, plots included four 7.62-m long rows with 76-cm inter-row spacing. Fertilizer, fungicides, and irrigation were used as needed, in accordance with local production practices. In 2015, sesame was planted in one location on April 30, and in 2016, sesame was planted in two locations on May 5, thus providing three location-years for analysis. In 2015, the soil type was Arredondo fine sand (loamy, siliceous, semiactive, hyperthermic Grossarenic Paleudult) with <1% organic matter. In 2016, soil types were Arredondo fine sand and Candler sand (hyperthermic, uncoated Quartzipsamment) with <1% organic matter. Rainfall totals for 30 d before planting (DBP) were 6.6 and 6.3 cm in 2015 and 2016, respectively (Florida Automated Weather Network 2016).
Sesame Response to Preplant Herbicides
The herbicide 2,4-D (Weedar 64, Nufarm Inc, Alsip, IL) was applied at 0.53 and 1.06 kg ha−1, and dicamba (Clarity, BASF Corporation, Research Triangle Park, NC) was applied at 0.56 and 1.12 kg ha−1. A CO2-pressurized backpack sprayer calibrated to deliver 187 L ha−1 with 11003 flat-fan nozzles was used for all applications. Applications of 2,4-D and dicamba occurred at five timings: 30, 15, 10, 5, and 0 DBP. Tillage was conducted to ensure that no vegetation was present at the time of treatment applications to maximize herbicide contact with the soil. Immediately prior to the 0 DBP application timing, ‘S38’ sesame (Sesaco Corporation, Austin, TX), a branching variety, was planted at a rate of 26 seeds m−2 (i.e., 20 seeds m−1). Counts of emerged sesame seedlings were conducted on the same 1 m of row in each plot at 1, 2, and 3 wk after planting (WAP) to measure the effects of 2,4-D and dicamba on sesame seedling emergence and survival. Heights of ten random plants per plot were measured at 3 and 6 WAP.
When 90% of sesame plants had mature seed capsules at the top of the plant, glyphosate was applied to terminate plant growth and initiate dry down (Langham et al. Reference Langham, Riney, Smith and Wiemers2008). When seeds reached 6% moisture, the middle two rows of each plot were machine-harvested, and seed weight was recorded. This experiment was conducted as a randomized complete block design with four replications using a factorial arrangement of three factors: herbicide (2,4-D and dicamba), herbicide rate (low and high), and application timing (30, 15, 10, 5, and 0 DBP).
Population Density Study
In a separate study, sesame was planted and thinned to various densities to determine the relationship between plant population and yield. The purpose of this study was to investigate the yield reductions that would occur if herbicide injury resulted in stand loss. S38 sesame was planted at a rate of 26.3 seeds m−2 (i.e., 20 seeds m−1). At 21 d after planting (DAP), plots were thinned to five plant densities: 26.3, 19.7, 13.2, 6.6, and 1.3 plants m−2. At seed maturity, plots were harvested and seed weight was recorded using the same methods described for the preplant herbicide study. The experiment was arranged in a randomized complete block design with four replications and was conducted once in 2015 and twice in 2016.
Statistical Analysis
Statistical analysis was conducted using the agricolae, stats, and investr packages in R (version 0.98.1091, RStudio Inc, Boston, MA). Data in both studies were subjected to ANOVA to test for main effects and interactions. Fisher’s protected LSD was utilized to separate means (α=0.05).
Additionally, regression analyses were conducted for both experiments. For the sesame response to preplant herbicides experiment, emergence 3 WAP and yield data were regressed over application timing to identify more precisely the application timing that was associated with a 10% yield reduction. Data were grouped into treatment categories of herbicide and rate combinations and then fit to asymptotic nonlinear regression curves. The asymptotic model used was
 $$Y\,{\equals}\,Y_{{{\rm asym}}} \left[ {1-{\rm exp}\left( {-aI/Y_{{{\rm asym}}} } \right)} \right],$$
$$Y\,{\equals}\,Y_{{{\rm asym}}} \left[ {1-{\rm exp}\left( {-aI/Y_{{{\rm asym}}} } \right)} \right],$$
where Y was the response variable [stand or yield; expressed as percent of the nontreated control (NTC)], Y asym was the asymptotic Y value, I was the explanatory variable [application timing (DBP) in the preplant herbicide experiment and stand (plants m−2) in the density experiment], and a was the initial slope of the curve at low I values. The fitted regression curves were used to estimate the DBP values associated with a 10% reduction in the response variable compared to the NTC (GR10). Reduction of 10% was chosen based on the overall variance of the data. Fisher’s protected LSD values in the first statistical analysis ranged from 9 to 10, reflecting the overall variance of the data. The GR10 values were then compared using their respective 95% confidence intervals.
Furthermore, yield data from the preplant herbicide experiment were asymptotically regressed over 3 WAP stand count data. The same procedure was followed for the density experiment yield data. Logarithmic rate constants were obtained from each treatment regression curve and compared using 95% confidence intervals. Additionally, YR10 values (10% reduction in yield expressed as plants m−2) were obtained from stand count regression curves and compared using 95% confidence intervals. Like GR10 values, YR10 values were based on 10% reductions to account for the data’s overall variance.
Results and Discussion
Sesame Response to Preplant Herbicides
No interactions were detected between year-location and main effects (P>0.22). Therefore, data were pooled across years and locations. However, interactions were detected between herbicide, rate, and application timing for all collected data.
Sesame emergence was not affected when 2,4-D was applied at 0.53 kg ha−1 30 to 10 DBP (Table 1). However, when 2,4-D at 0.53 kg ha−1 was applied 5 and 0 DBP, sesame emergence was reduced 17% and 58%, respectively, when evaluated 3 WAP. 2,4-D at 1.06 kg ha−1 and dicamba at 0.56 kg ha−1 did not reduce emergence when applied 30 to 15 DBP; however, reductions in emergence were observed when either treatment was applied 10 to 0 DBP. Sesame emergence evaluated 1 and 2 WAP was not reduced from dicamba at 1.12 kg ha−1 until applications were made 10 DBP. However, by the 3 WAP evaluation, dicamba at 1.12 kg ha−1 reduced emergence when applied 15 DBP.
Table 1 Effect of 2,4-D and dicamba application timing on sesame emergence, plant height, and yield expressed as percentage of the nontreated control.Footnote a

a Abbreviations: DBP, days before planting; WAP, weeks after planting.
b Asterisk signifies that the mean is different from the nontreated control according to Fisher’s LSD (α=0.05).
Although reductions in emergence were commonly detected when herbicides were applied 5 DBP, the effect increased dramatically (often more than 3-fold) when the application was made at 0 DBP. For example, 2,4-D at 0.53 kg ha−1 reduced sesame emergence 12% to 19% when applied 5 DBP, but when applied 0 DBP, reductions of 57% to 59% were observed. Additionally, the response to dicamba was more severe than the response to 2,4-D applied at nearly equivalent rates. Differential sensitivity of plants to 2,4-D and dicamba is common and usually varies by plant family. For example, legumes are generally more sensitive to dicamba than they are to 2,4-D (Johnson et al. Reference Johnson, Fisher, Jordan, Edmisten, Stewart and York2012; Lyon and Wilson Reference Lyon and Wilson1986; Prostko et al. Reference Prostko, Grey, Johnson, Jordan, Grichar, Besler, Brewer and Eastin2003; Thompson et al. Reference Thompson, Steckel, Ellis and Mueller2007; Wax et al. Reference Wax, Knuth and Slife1969), while some plant families such as Malvaceae are generally the opposite (Everitt and Keeling Reference Everitt and Keeling2009; Johnson et al. Reference Johnson, Fisher, Jordan, Edmisten, Stewart and York2012; Marple et al. Reference Marple, Al-Khatib and Peterson2008). However, there is no published literature on sesame’s sensitivity to 2,4-D or dicamba.
Sesame plant height was not reduced at either evaluation interval when 2,4-D at 0.53 kg ha−1 was applied 30 to 5 DBP (Table 1). However, when applied 0 DBP, 2,4-D at 0.53 kg ha−1 caused 29% and 36% reductions in plant height at the 3 and 6 WAP evaluation intervals, respectively. Sesame plant height was not reduced when either 2,4-D at 1.06 kg ha−1 or dicamba at 0.56 kg ha−1 was applied 30 to 10 DBP and 30 to 5 DBP with either evaluation timing, respectively. At the 6 WAP evaluation, 2,4-D at 1.06 kg ha−1 and dicamba at 0.56 kg ha−1 applied 0 DBP resulted in 41% and 43% reduction in plant height, respectively. Dicamba at 1.12 kg ha−1 did not reduce sesame plant height at either evaluation interval when applied 30 to 15 DBP. However, plant height reductions ranging from 14% to 68% were observed at both evaluation intervals when dicamba at 1.12 kg ha−1 was applied 10 to 0 DBP.
Sesame grain yield was less responsive to herbicide application timing than were emergence and height. Both rates of 2,4-D and dicamba at 0.56 kg ha−1 did not cause yield reductions when applied 30 to 5 DBP (Table 1). However, when applied 0 DBP, 2,4-D at 0.53 and 1.06 kg ha−1 and dicamba at 0.56 kg ha−1 reduced yield by 18%, 29%, and 26%, respectively. Dicamba at 1.12 kg ha−1 did not reduce yield when applied 30 to 10 DBP. Nevertheless, when applied 5 and 0 DBP, dicamba at 1.12 kg ha−1 reduced yield 22% and 48%, respectively. Interestingly, application timings that reduced emergence or plant height early in the season did not necessarily cause significant yield reductions for some treatments. For example, 2,4-D at 1.06 kg ha−1 and dicamba at 0.56 kg ha−1 caused reductions in emergence and plant height when applied 5 DBP, but did not affect yield. Although plant number and overall stature can be reduced by as much as 30% to 40% from applying 2,4-D or dicamba close to planting, the remaining plants tend to flower and yield normally, compensating for the stand loss. This speaks to the yield compensatory ability of sesame to changes in population.
While the difference between the 0 and 5 or 5 and 10 DBP application timings is noteworthy, a more precise estimation of sesame’s response to applications between the tested timings would be beneficial. Burndown applications are most effective when applied close to planting. Shortening the interval between burndown and planting is especially critical in sesame as there are no herbicide options for POST control of broadleaf weeds. Additionally, in order to prevent sesame injury, S-metolachlor is best applied 3 to 6 d after planting (Sperry et al. Reference Sperry, Ferrell, Leon, Rowland and Mulvaney2016). Due to the delayed application of S-metolachlor, weeds may emerge in the time period between burndown and PRE applications, especially Palmer amaranth (Amaranthus palmeri S. Wats.), which emerges almost continuously during the summer months (Jha and Norsworthy Reference Jha and Norsworthy2009). Consequently, the ability to apply 2,4-D or dicamba 1 to 2 d closer to planting could potentially provide higher levels of early-season weed control.
It is important to note that soils at both experimental locations favored maximum herbicide mobility and availability. Prior research has concluded that the soil mobility of 2,4-D and dicamba increase as soil organic matter content decreases (Grover Reference Grover1977). The soils used for these experiments both contained <1% organic matter, ensuring that there was a high likelihood for herbicide injury. Future work should focus on the interactions of preplant-applied 2,4-D and dicamba with sesame planted in soils with varying texture and organic matter content, to define the relationship between application timing and soil type.
Regression Analysis
The analysis shown in Table 1 allowed us to observe when significant reductions in sesame emergence, plant height, and yield occurred relative to discrete application timings. However, to more accurately predict when these herbicides can be safely applied, a regression analysis was conducted. There were no differences between GR10 values for emergence 1 and 2 WAP (data not shown); however, differences in GR10 values were present for emergence 3 WAP. At the 3 WAP emergence evaluation, 2,4-D at 0.53 kg ha−1 had the lowest GR10 value of 6.4 DBP (Table 2). These data indicate that 2,4-D at 0.53 kg ha−1, applied at 6.4 DBP, resulted in only a 10% reduction in total emergence at 3 WAP. In contrast, dicamba at 0.56 and 1.12 kg ha−1 resulted in GR10 values of 12.3 and 15.7 DBP, respectively. Also 2,4-D at 1.06 kg ha−1 had a lower GR10 value (10.7) than did dicamba at 1.12 kg ha−1, suggesting that sesame was more sensitive to preplant-applied dicamba than it was to 2,4-D. Similarly, cotton and soybean have been shown to be more tolerant to preplant applications of 2,4-D than those of dicamba. Everitt and Keeling (Reference Everitt and Keeling2007) reported no reduction in cotton stand when 2,4-D at 1.12 kg ha−1 was applied 7 DBP, although dicamba applied at 0.14 and 0.28 kg ha−1 caused 29% stand reduction when applied 14 DBP. Likewise, Keeling et al. (1989a) reported that 2,4-D did not result in any injury when applied 15 to 18 DBP, although dicamba caused >15% injury when applied at the same timing. In soybean, Thompson et al. (Reference Thompson, Steckel, Ellis and Mueller2007) reported that dicamba caused up to 60% higher injury to soybean than did 2,4-D when both were applied 0 DBP. Therefore, like other sensitive crops, sesame planting must be delayed longer after preplant applications of dicamba compared with 2,4-D.
Table 2 GR10 Footnote a values from nonlinear regressionb of 2,4-D and dicamba application timing on sesame emergence, determined 3 weeks after planting (WAP), and yield, expressed in days before planting (DBP). These data represent the number of DBP that 2,4-D and dicamba should be applied resulting in 10% reduction in emergence and final yield.

a GR10, days before planting that result in 10% reduction in emergence or yield compared to the nontreated control.
b The asymptotic model used the following equation: Y=Y asym[1 – exp(–aI/Y asym)], where Y is the response (stand or yield), Y asym is the asymptotic Y value, I is the explanatory variable [application timing (DBP)], and a is the initial slope of the curve at low I values.
c 95% confidence intervals are given in parentheses.
Sesame yield was most affected by dicamba at 1.12 kg ha−1, which had a GR10 value of 10.3 DBP, higher than those of all other treatments (Table 2). The GR10 values for both rates of 2,4-D and dicamba at 0.56 kg ha−1 ranged from 2 to 3.7 DBP, with no difference between treatments. Similarly, Krausz et al. (Reference Krausz, Kapusta and Mathews1993) reported no difference in soybean yield compared to NTC when 2,4-D (0.56 kg ha−1) was applied 0 DBP. Also, Everitt and Keeling (Reference Everitt and Keeling2007) reported 2,4-D and dicamba applied to cotton 7 DBP at rates up to 1.12 and 0.28 kg ha−1, respectively, did not cause any reductions in yield. A possible explanation for sesame’s higher tolerance to 2,4-D than dicamba may be the differing soil degradation rates of the herbicides. Altom and Stritzke (Reference Altom and Stritzke1973) reported that the half-life of 2,4-D on three different soils ranged from 4 to 5 d, while the half-life of dicamba ranged from 17 to 32 d.
Yield GR10 values were lower than emergence GR10 values, which shows sesame’s ability to compensate in yield for reduced stand (Table 2), often through increased branching. For example, the emergence GR10 for dicamba at 0.56 kg ha−1 was 12.3 DBP, but the yield GR10 was 3 DBP. Also, it is important to note that the confidence intervals for yield GR10 values were smaller than those for emergence. Lower variance in the yield data suggests that plants either recovered from herbicide injury or that the plants that survived to maturity may have escaped prolonged herbicide effects. Soybean injury from preplant applications of 2,4-D has been shown to decrease over time, suggesting that some crops can fully recover from early-season herbicide injury (Krausz et al. Reference Krausz, Kapusta and Mathews1993). The data in Table 2 suggest that some level of sesame stand loss from applications of 2,4-D and dicamba can occur without affecting yield. Therefore, to further understand the relationship between sesame stand and grain yield, an analysis was conducted using stand count and yield data from the preplant herbicide experiment as well as the density experiment to investigate the compensatory ability of sesame in the presence and absence of herbicides.
Stand-Yield Regression with Density Experiment
No interactions were detected between location-years and main effects; therefore, data were pooled across locations and years for both the density study and preplant herbicide study (P=0.24 and P=0.22, respectively).
Sesame yield was not affected when plant population was 26.3, 19.7, or 13.2 plants m−2 (Table 3). However, sesame populations of 6.6 and 1.3 plants m−2 reduced yield by 15% and 53%, respectively. Sesame yield did not decrease until population was reduced to 6.6 plants m−2, indicating that the population needed to produce yields similar to the NTC was between 6.6 and 13.2 plants m−2. The difference between 6.6 and 13.2 plants m−2 reflects 25% and 50% of the NTC population density, which are large differences in plant stand. Regression analysis was conducted to identify the population at which yield was significantly reduced compared to the NTC. The analysis allowed greater resolution relative to how much density can be lost before yield is significantly affected and replanting should be considered.
Table 3 Effect of sesame plant density on seed yield, expressed as percentage of a nontreated control.

a Means followed by the same letter are not different according to Fisher’s protected LSD at P=0.05.
Regression analysis of the population study data indicated the YR10 (10% yield reduction from NTC) of sesame to be 8.6 plants m−2 (Table 4). This value indicates that an approximately 68% population reduction only reduces yield 10%. In many other crops, a 68% population reduction would be considered complete crop failure, especially for determinate crops. Cotton, also an indeterminate crop, has been shown to produce similar yields to the NTC with population reductions of up to 83% (Bednarz et al. Reference Bednarz, Shurley, Anthony and Nichols2005). Soybean has a similar compensation capacity and typically only suffers a 2% to 12% reduction in yield after stand is decreased by 25% to 67% (Carpenter and Board Reference Carpenter and Board1997; Conley et al. Reference Conley, Abendroth, Elmore, Christmas and Zarnstorff2008; Hicks et al. Reference Hicks, Lueschen and Ford1990).
Table 4 Parameters of nonlinear regressionFootnote a of the effect of sesame population on yield for sesame treated with 2,4-D and dicamba applied preplant, compared to nontreated control (NTC).
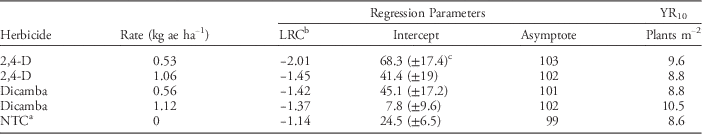
a The asymptotic model used the following equation: Y=Y asym[1 – exp(–aI/Y asym)], where Y is the response (yield), Y asym is the asymptotic Y value, I is the explanatory variable (plants m−2), and a is the initial slope of the curve at low I values.
b Abbreviations: LRC, logarithmic rate constant (relative slope); YR10, 10% yield reduction from NTC.
c 95% confidence intervals are given in parentheses. Values within columns not followed by parentheses are not statistically different.
Unlike US sesame varieties that are nondehiscent, shattering varieties have been shown to exhibit more determinate behavior and are not able to compensate as well for population reductions (Noorka et al. Reference Noorka, Hafiz and El-bramawy2011; Roy et al. Reference Roy, Mamun and Jahan2009). A review of the literature revealed that optimum yields occurred when plant populations ranged from 16.7 to 51 plants m−2 in shattering sesame varieties (Caliskan et al. Reference Caliskan, Arslan, Arioglu and Isler2004; El Naim et al. Reference El Naim, El day and Ahmed2010). The current study, using US varieties, found sesame populations of 8.6 plants m−2 to yield 90% to 100% compared to the NTC (Table 4). Delgado and Yermanos (Reference Delgado and Yermanos1975) found that sesame yield reductions of >10% occur at a density value between 17.5 and 8.8 plants m−2 using US varieties, although regression analysis was not utilized to find a more exact value.
Stand count data (3 WAP) from the preplant herbicide study were also regressed to investigate the effect of stand on yield in sesame that had been treated with 2,4-D and dicamba compared to the population study data (expressed as NTC in Table 4). No differences in YR10 values were detected between treatments and the NTC. The YR10 values ranged from 8.6 to 10.5 plants m−2, which is equivalent to a 60% to 67.5% reduction in stand, while only reducing yield 10% or less. Despite high levels of stand reduction that can occur and consequently be alarming to growers early in the crop’s development, the decision to replant should only be considered if stand falls below 8.6 to 10.5 plants m−2.
Despite early-season reductions in sesame emergence and plant height from preplant-applied 2,4-D and dicamba, plants that survived to maturity appeared to grow and yield normally. Therefore, it was assumed that reductions in yield were simply due to stand reduction and no lasting physiological effects from herbicide exposure affected yield. In order to test this assumption, regression parameters were compared between herbicide-treated plants (preplant herbicide experiment) and nontreated plants (density experiment). Parameters included logarithmic rate constants (relative slope), y-intercepts, and asymptotes (Table 4). Regression curves for all treatments revealed no differences between logarithmic rate constants values or asymptotes (Table 4). This suggests that the reductions in yield observed in the preplant herbicide experiment were due simply to reductions in sesame stand. Consequently, it seems that stunting caused by herbicide exposure did not affect reproductive physiology and yield was not affected. This also confirms that plants that survive to maturity either recovered from herbicide injury or escaped exposure all together.
Intercepts did however differ between treatments. The 0.53 kg ha−1 rate of 2,4-D regression produced an intercept of 68.3, which was higher than those of all other treatments and the NTC (Table 4). Conversely, the dicamba at 1.12 kg ha−1 regression produced an intercept of 7.8, which was lower than those of all other treatments and the NTC. These differences in intercepts mirror the trends that were present in earlier data: 2,4-D at 0.53 kg ha−1 was the least injurious and dicamba at 1.12 kg ha−1 was the most injurious. Theoretically, the y-intercept (x=0 plants m−2) would have a yield of 0; however, these intercepts are predicted values based on the fitted regression model. Therefore, the intercepts reflected the measured stand 3 WAP in the preplant herbicide experiment. For example, 2,4-D at 0.53 kg ha−1 had an intercept much higher than did other treatments, because sesame stand was rarely reduced to the lower end of the graph (0 to 6.6 plants m−2) and most data points were concentrated towards the high end of the curve which increased the y-intercept. Conversely, dicamba at 1.12 kg ha−1 had an intercept much lower than did other treatments, because sesame stand was severely reduced and most of the data was concentrated on the lower end of the graph. Regardless, the differences observed in intercepts only reflect sesame emergence 3 WAP and not any physiological differences in the ability of sesame to compensate.
In summary, higher tolerance of sesame was observed to preplant applications of 2,4-D than to preplant applications of dicamba. The findings of the current studies support applications of 2,4-D and dicamba closer to sesame planting than the current labels suggest for cotton or soybean. This allows a wider window for applications and planting to avoid unfavorable environmental conditions. For example, to avoid injury to cotton the product labels suggest waiting 30 d after 0.53 kg ha−1 2,4-D applications and 21 d plus the accumulation of 2.54 cm of rainfall or irrigation for dicamba at 0.56 kg ha−1 (Anonymous 2004, 2010). For soybean, the product label recommends a 15 d plant back period after 1.06 kg ha−1 of 2,4-D, and 14 d plus 2.5 cm of rainfall or irrigation for dicamba at 1.12 kg ha−1 (Anonymous 2004, 2010). Reductions in sesame emergence and plant height did occur when these herbicides were applied less than a week before planting sesame, especially for higher rates of dicamba. The response of sesame to higher rates of 2,4-D and dicamba must be taken into account, as higher rates are sometimes required to control larger weeds. Also, it is important to note that yield reductions from herbicide injury are most likely less than the yield reductions that would occur as result of weed presence. Wilson and Worsham (Reference Wilson and Worsham1988) reported that after integrating 2,4-D at various rates into burndown applications with paraquat and glyphosate applied 0 DBP, soybean yield was higher than it was when the crop was faced with weed competition. Currently, there are no POST herbicide options available in sesame; therefore, some level of herbicide injury may have to be tolerated.