


Introduction
The recently developed allele-specific DNA markers for defining the allele variation in major loci controlling growth habit (vernalization, photoperiod response and plant stature) (Korzun et al., Reference Korzun, Roder, Ganal, Worland and Law1998; Ellis et al., Reference Ellis, Spielmeyer, Gale, Rebetzke and Richards2002; Yan et al., Reference Yan, Helguera, Kato, Fukuyama, Sherman and Dubcovsky2004; Fu et al., Reference Fu, Szücs, Yan, Helguera, Skinner, von Zitzewitz, Hayes and Dubcovsky2005; Beales et al., Reference Beales, Turner, Griffiths, Snape and Laurie2007) have allowed more efficient characterization of wheat germplasm. These markers have also contributed to the deeper understanding of the effect of allele variation on the agronomically important traits as well as to more effective management of selection activities during the breeding process. Our previous works on the identification, distribution and effect of the semi-dwarfing Rht alleles on some agronomic traits in Bulgarian bread wheat showed that the allele 192 bp at the microsatellite locus Xgwm 261, which is linked to Rht8 gene (2DS), prevailed in the cultivars developed during the last 50 years (Ganeva et al., Reference Ganeva, Korzun, Landjeva, Tsenov and Atanassova2005; Zheleva et al., Reference Zheleva, Todorovska, Jacquemin, Atanassov, Christov, Panayotov and Tsenov2006).
This study was motivated by the lack of more complete information concerning the allele variation at the major loci for growth habit and their effect on maturity and other productive agronomic traits in Bulgarian bread wheat varieties. The aim was to define and to assess the allele variation and distribution at loci for growth habit genes (Vrn, Ppd and Rht) in Bulgarian germplasm collections from the Institute of Plant Genetic Resources (IPGR), Sadovo and Dobroudzha Agricultural Institute (DAI), G. Toshevo, Bulgaria. The obtained results could also prove useful for planning and implementation of molecular markers in practical wheat-breeding programmes.
Material and methods
Plant material
A total of 52 old and modern Bulgarian wheat genotypes from the genebank collections of the IPGR, Sadovo and DAI, Bulgaria were used in this study. The modern cultivars were developed in the period 1960–2008.
Microsatellite analysis
Microsatellite analysis of the Xgwm261 locus was performed as described by Röder et al. (Reference Röder, Korzun, Wendehake, Plaschke, Tixier, Leroy and Ganal1998), using Cy5′-labelled forward and unlabelled reverse primers. Fragment analysis was carried out on AFL Express II sequencer (Amersham Biosciences). The size of the fragments was determined with the program Allele locator, version 1.03.
Ppd-D1
The alleles of the pseudo-response regulator Ppd-D1 (2D), which determine the sensitivity or insensitivity to day length, were detected using primers according to Beales et al. (Reference Beales, Turner, Griffiths, Snape and Laurie2007). Following Beales et al. (Reference Beales, Turner, Griffiths, Snape and Laurie2007), the photoperiod-insensitive allele was labelled as Ppd-D1a. The alternative allele, inferring photoperiod sensitivity, was designated Ppd-D1b by analogy.
Vrn-A1
The dominant spring alleles (Vrn-1Aa and Vrn-1Ab) were identified utilizing genome-specific primers (VRN1AF and VRN1-INT1R) described by Yan et al. (Reference Yan, Helguera, Kato, Fukuyama, Sherman and Dubcovsky2004), by variation in the promoter region of the Vrn-A1 locus. The primer combination Intr1/A/F2/Intr1/A/R3 was used to differentiate the dominant spring allele VrnA1c from vrn-A1 (the recessive winter allele).
Vrn-D1
Vrn-D1 intron-1 alleles were detected using Intr1/D/F, Intr1/D//R3 and Intr1/D/R4 primers (Fu et al., Reference Fu, Szücs, Yan, Helguera, Skinner, von Zitzewitz, Hayes and Dubcovsky2005). The primer pair Intr1/D/F and Intr1/D//R3 was used for the amplification of the dominant spring allele Vrn-D1, while the primer pair Intr1/D/F and Intr1/D/R4 was used for the amplification of the recessive winter allele vrn-D1.
Rht-1
The primer pairs BF/WR1 and BF/MR1 (Ellis et al., Reference Ellis, Spielmeyer, Gale, Rebetzke and Richards2002) were used for the amplification of the alleles Rht-B1a (wild type) and Rht-B1b (dwarf), respectively.
Results and discussion
Distribution of the alleles of the major growth habit genes (Ppd-D1, Vrn-1, Rht-1 and Rht8) within Bulgarian bread wheat genotypes (Triticum aestivum L.)
The assessment of Vrn-1(5A/5B/5D), Ppd-D1(2DS), Rht-1(4B) and Rht8(2D) allele variation in 52 old and modern Bulgarian wheat cultivars and landraces showed that the recessive (vrn) alleles of Vrn-1 genes responsible for winter growth habit prevailed in both the old and the modern Bulgarian genotypes (72.7 and 93.3%, respectively).
The dominant alleles Vrn-A1a and Vrn-A1b, which determine the spring growth habit, were found only in two cultivars, the modern one – ‘Gladiator 113’ and the old one – ‘Karnobatska ranozreika’. A combination of two dominant alleles Vrn-A1b and Vrn-B1 was identified only in cv. ‘Karnobatska ranozreika’. Alleles Vrn-A1a and Vrn-A1b separately or in combination with other Vrn-1 alleles have been observed in high proportion in spring wheat genotypes from Argentina, Canada, USA and China (Yan et al., Reference Yan, Helguera, Kato, Fukuyama, Sherman and Dubcovsky2004; Fu et al., Reference Fu, Szücs, Yan, Helguera, Skinner, von Zitzewitz, Hayes and Dubcovsky2005; Santra et al., Reference Santra, Santra, Allian, Campbell and Kidwell2009). Similar to the results in some other studies (Fu et al., Reference Fu, Szücs, Yan, Helguera, Skinner, von Zitzewitz, Hayes and Dubcovsky2005; Iqbal et al., Reference Iqbal, Navabi, Yang, Salmon and Spaner2007), the allele Vrn-A1c was not found in Bulgarian wheat germplasm. On the other hand, the spring allele Vrn-B1 was found only in the old Bulgarian cultivars and landraces, while Vrn-D1 allele was detected in a few old and in one modern cultivar. The photoperiod sensitive allele (Ppd-D1b) prevailed in the old cultivars (90.9%). A significant increase in the frequency of the photoperiod-insensitive allele (Ppd-D1a) was observed in the modern Bulgarian cultivars (96.7%) (Fig. 1). The same tendency has also been observed for the modern Chinese wheat (Yang et al., Reference Yang, Zhang, Xia, Laurie, Yang and He2009). In our study, the allele Rht-1a (tall type) was found in both old cultivars and landraces as well as in the modern ones. The semi-dwarfing allele (Rht-1b) was detected in several modern cultivars. This may be related to the greater use of CIMMYT germplasm in Bulgarian breeding in the previous years (Table 1). At the Xgwm261 (2D) locus located close to the Rht8 gene, the alleles of 164, 212 and 216 bp length were found only in the old cultivars and landraces.
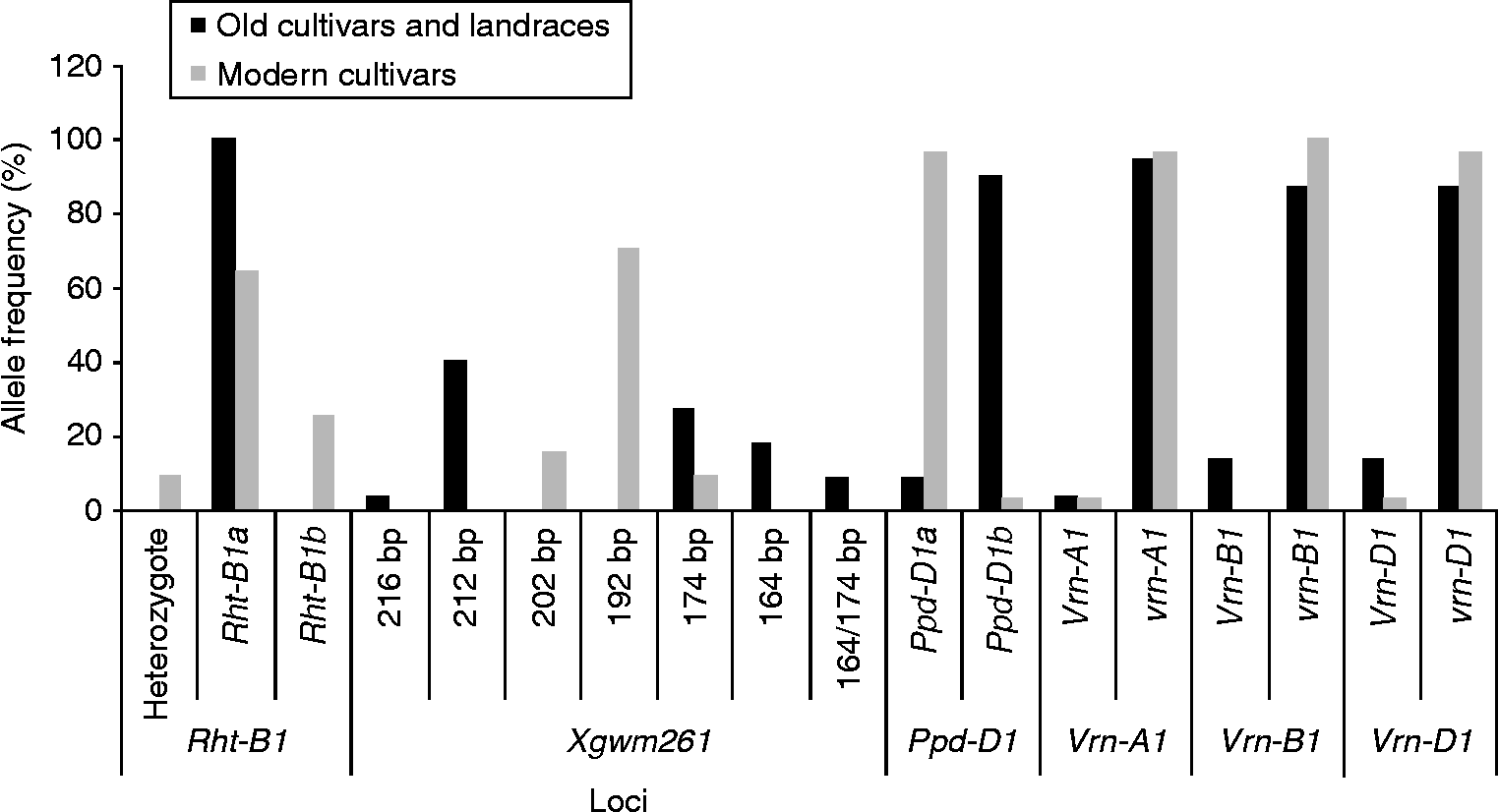
Fig. 1 Distribution of alleles for major growth habit genes among the old and modern Bulgarian wheat genotypes.
Table 1 Rht-1, Xgwm261(Rht8), Ppd-D1 and Vrn-1 allele composition in the modern Bulgarian bread wheat cultivars (T. aestivum L.) and their heading datesa

a The data were recorded during 2005–2009.
Allele combinations of the major growth habit genes (Ppd-D1, Vrn-1, Rht8 and Rht-1) in the modern Bulgarian bread wheat and their influence on some agronomic traits
The contribution of the allelic composition at the Vrn, Ppd and Rht loci to the variation of the heading date and other agronomic traits was studied in the modern Bulgarian bread wheat cultivars developed after 1960. In these cultivars, the dominating allele is 192 bp at locus Xgwm261 (2D), which is considered to be a diagnostic for the presence of Rht8 gene.
Here, it was confirmed that some of the commercial Bulgarian cultivars carried the Rht8 gene alone or in combination with the Rht-1 gene. Similar results have been reported by Ganeva et al. (Reference Ganeva, Korzun, Landjeva, Tsenov and Atanassova2005), using gibberellin test and isogenic lines of cv. ‘Mercia’. The combination of different Rht genes in Bulgarian cultivars could be considered as successful, as these cultivars have found extensive use in the breeding practice. In the last few years, the lands sown with the cultivars ‘Enola’ (Rht-B1b+Rht8) and ‘Aglika’ (Rht-B1a+202 bp allele at the Xgwm 261 locus) comprise 19 and 10%, respectively, of the total wheat-sown areas in 2009, followed by ‘Sadovo1’ (8.5%), ‘Pobeda’ (6.1%) and ‘Milena’ (3.7%) (Table 1). The photoperiod insensitivity of the modern Bulgarian wheat cultivars is due to the presence of the Ppd-D1a allele, which has been introduced from the main donors of Rht8 (192 bp allele at the Xgwm261 locus) in Bulgarian wheat such as the Russian cultivars ‘Bezostaya 1’, ‘Skorospelka’ and ‘Kavkaz’.
All modern Bulgarian wheat cultivars excluding ‘Laska’ (vrn-A1/vrn-B1/Vrn-D1) and ‘Gladiator 113’ (Vrn-A1a/vrn-B1/vrn-D1) were of the winter type (vrn-A1/vrn-B1/vrn-D1). Ten allele combinations for growth habit and plant stature were detected in the old cultivars and landraces. In contrast, the major combination of alleles of the photoperiod sensitivity (Ppd-D1) and the Vrn-1(5A/5B/5D) genes in the modern Bulgarian wheat gene pool Ppd-D1a//vrn-A1/vrn–B1/vrn-D1 amounted to 93.1%. The results showed that most of the early-heading cultivars possessed the allelic combination Rht-B1b//192 or 202 bp allele at the Xgwm261 locus//Ppd-D1a//vrn-A1/vrn–B1/vrn-D1 (Table 1).
Additional studies on the allele variation in other genes affecting the photoperiod sensitivity in wheat (Ppd-B1 and Ppd-A1), the vernalization response (Vrn-B3 and Vrn-D4) and the earliness per se gene(s) would generalize to a great extent our knowledge of their effect on maturity and other agronomic traits.
Assessment of the effects of these genes on the heading date and grain yield in diverse Bulgarian production environments is also a task of current priority.



