INTRODUCTION
Marine mollusk shells are commonly found in prehistoric middens across Ireland, having likely been an important dietary component of many prehistoric communities (Murray Reference Murray, Murphy and Whitehouse2007). Limpets, periwinkles, and mussels all were exploited by prehistoric humans and their shells are widespread in archaeological sites across northern Europe (Gutiérrez-Zugasti et al. Reference Gutiérrez-Zugasti, Andersen, Araújo, Dupont, Milner and Monge-Soares2011). Shell middens have the potential to provide information on resource management and population movement, as well as past environments (Alvarez et al. Reference Alvarez, Godino, Balbo and Madella2011). Although mollusk shells provide carbon-rich material that can be measured for 14C content, accurate chronological control of prehistoric events requires knowledge of any in-built age in the carbon source used for shell construction. Equally, knowledge of any in-built age in the flesh of the limpets will affect the dating of bones, where limpets have been a dietary component (Milner Reference Milner, Conneller and Warren2006; Meiklejohn and Woodman Reference Meiklejohn and Woodman2012).
For shell building, mollusks obtain carbon from two sources, dissolved inorganic carbon (DIC) in sea water, and organic carbon from within the marine food chain. The proportion of carbon derived from metabolic sources may depend upon a number of factors, including habitat and the way that individual mollusk species feed (Tanaka et al. Reference Tanaka, Monaghan and Rye1986; Dettman et al. Reference Dettman, Reische and Lohmann1999). In some coastal areas, metabolic carbon originating from terrestrial plants and soils also could also be incorporated in mollusk shells. Feeding preference and habitat, on substrate containing old carbon such as limestone rock, can result in an apparently old age for both mollusk shell and flesh (Dye Reference Dye1994). Subsequent research has avoided areas with limestone geology on the assumption that old carbon is incorporated into the samples. From the point of view of archaeological sites, humans or animals feeding on mollusks also can incorporate this carbon into bone collagen. In this way, determining the existence of such an age offset and its size is an important aspect of using 14C for dating bones from coastal environments as well as shells.
It is necessary to consider the biology of the mollusks to understand how geological carbon could potentially be taken up into the bodies and shells. Patella vulgata, also known as the common limpet, is a microphagous herbivore gastropod found in the intertidal zones of rocky shores in northwestern Europe. The powerful radula (teeth-like structure in mollusks) of this species of P. vulgata is capable of gouging deeply into the substrate to remove microalgae as well as other larger objects (Hawkins et al. Reference Hawkins, Watson, Hill, Harding, Kyriakides, Hutchinson and Norton1989). The same mechanism, possibly combined with chemical dissolution using mucus, is used for creating the depression scar on which the P. vulgata makes its home (Lindberg and Dwyer Reference Lindberg and Dwyer1983). P. vulgata usually feeds during daytime or nocturnal high waters and travels an average of 0.4 m from its home scar, returning before exposure occurs at low tide. Even if relocation is necessary, they do not travel more than 2 m to a new home (Hartnoll and Wright Reference Hartnoll and Wright1977). This would suggest that an individual is usually grazing on one type of substrate during its lifetime. In contrast, mussels (Mylitus edulis) attach to the substrate and siphon water to filter out plankton and microscopic organisms. Importantly for this study, Ascough et al. (Reference Ascough, Cook, Dugmore, Scott and Freeman2005) also have shown that the feeding and habitat behavior of P. vulgata and M. edulis, from a coastline with no geological carbon influence, did not have a significant influence on 14C activity in their shells.
The coast of Ireland lies between the Atlantic Ocean to the west and the Irish Sea to the east. Outcrops of limestones are common and, in the north, are overlain by extensive basalts (GSNI 2018). Sellafield, on the Cumbrian coast in the northeast of the Irish Sea, is a nuclear reprocessing and decommissioning complex that processes all forms of radioactive waste. Normally we would expect the 14C of the Irish Sea to be similar to that of the “global” ocean based on measurements from pre-nuclear testing shells (Harkness 1983; Blake 2005; Butler et al. 2009). However, radioactive waste including 14C is discharged to the aquatic environment as low-level waste, primarily as DIC. This 14C mixes with the existing 14C in the seawater and is dispersed across the Irish Sea with the currents (Muir et al. Reference Muir, Tierney, Cook, MacKinnon, Howe, Heymans, Hughes and Xu2016) and is incorporated into the shells of mollusks. The levels of concentration of discharged 14C are monitored and published in an annual report as activity in Bequerels (Bq). In 2007, a total of 4.7 TBq of 14C was discharged into the Irish Sea (Nuclear Decommissioning Authority 2007) and in 2017, a total of 3.6 TBq of 14C was discharged (Nuclear Decommissioning Authority 2017).
In this paper we measure the 14C of shells and flesh of intertidal limpets (P. vulgata) from both volcanic and limestone rocks at six sites from around the Irish coast to determine variation in mollusk 14C content. We are particularly interested in documenting any species-specific, shell-flesh, and granite/basalt-limestone 14C offsets. It was expected that the discharge of 14C into the Irish Sea from the Sellafield nuclear reprocessing site located on the opposite shore would accentuate the difference between ingested limestone and dissolved inorganic carbon of the seawater in specimens from the west coast of Northern Ireland and help identify the need for reservoir corrections.
METHODS
Sample Collection
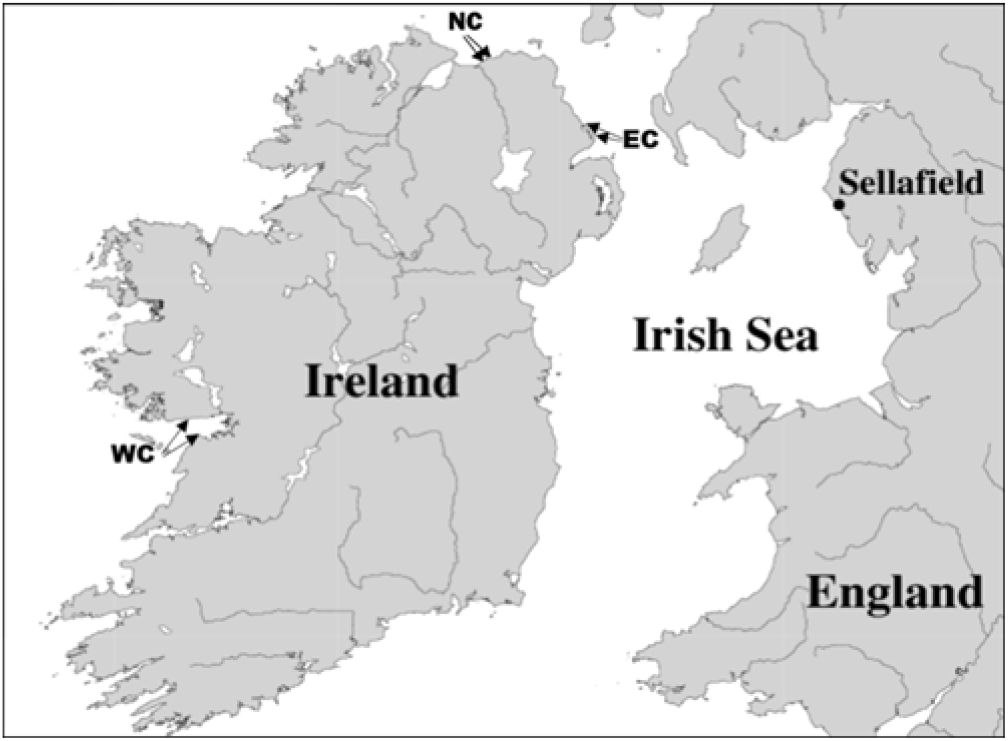
Five individual limpets (Patella vulgata) were collected living on intertidal limestone and basalt rocks sites on the east coast of Northern Ireland (EC) at Portmuck and The Gobbins, Islandmagee, in October 2007. Five limpets were also collected living on limestone and granite rocks on the west coast of Ireland (WC) at Black Head and near Loughaunbeg, Galway Bay. Analyses were completed but the results were not published at that time. Further samples were collected in July and September 2018 from the north coast (NC) at Whiterocks, Portrush, and Rinagree Point, Portstewart. The locations of the sample sites are shown in Figure 1 and geological substrate is included in the results tables. A full list of samples collected can be found in Tables 3 and 4 in the appendix. For comparison, samples of mussels (M. edulis) were also collected from two of the sites, as M. edulis is a filter-feeding mollusk and should not be affected by substrate type.

Figure 1 Location map of sample collection sites.
Radiocarbon Analyses
Mollusk individuals were heated in Milli-Q® water until the flesh easily separated from the shell. The digestive organs were identified under a low-powered binocular microscope, then cut away from the remaining flesh and discarded, ensuring that the entire digestive tract was removed. The flesh was treated with 4% HCl for 1 hr at 80ºC, rinsed five times with Milli-Q® water, dried overnight at 70ºC, and ground with mortar and pestle until all material passed through a 250-μm sieve to ensure homogeneity. A subsample (~100 mg) of each shell was etched in 1% HCl (2 mL per 100 mg) overnight to remove surface carbonates, rinsed five times with Milli-Q® water, dried overnight at 70ºC, and coarsely ground with a mortar and pestle.
Subsamples of mollusk flesh for 14C analysis were individually loaded into quartz sample tubes with 30 mg of pre-combusted CuO and a strip of Ag foil then sealed under vacuum. Sealed tubes were combusted to CO2 at 900°C for at least 4 hr. Approximately 12 mg of each pretreated shell was weighed into 4 mL Exetainer® vials (LabCo, Inc.), evacuated, injected with 1.5 mL of concentrated (85%) phosphoric acid to hydrolyze the shell to CO2, and heated at 80ºC for 20 min or until all bubbling ceased.
Samples were converted to graphite on an iron catalyst using the hydrogen reduction method (Vogel et al. Reference Vogel, Southon and Nelson1987) and the 14C/12C ratio and 13C/12C were measured by accelerator mass spectrometry at the 14CHRONO Centre, Queen’s University Belfast. The radiocarbon age and one standard deviation were calculated using the Libby half-life of 5568 years following the conventions of Stuiver and Polach (Reference Stuiver and Polach1977). The ages were corrected for isotope fractionation using the AMS measured δ13C (not given) which accounts for both natural and machine fractionation. 14C data are reported as F14C (Reimer et al. Reference Reimer, Brown and Reimer2004).
A statistical analysis of the groups of measurements, using one-way analysis of variance (ANOVA), was carried out between the sets of limpets at each site as well as testing the P. vulgata and M. edulis samples as three groups at the east and west coast sites. ANOVA is a test of differences with the null hypothesis being that two sets of data are the same and can be used to test more than two groups of samples together. In this study we are aiming to accept the null hypotheses, so a P value of above 0.05 is needed to be confident that there are no statistical differences between data sets.
RESULTS
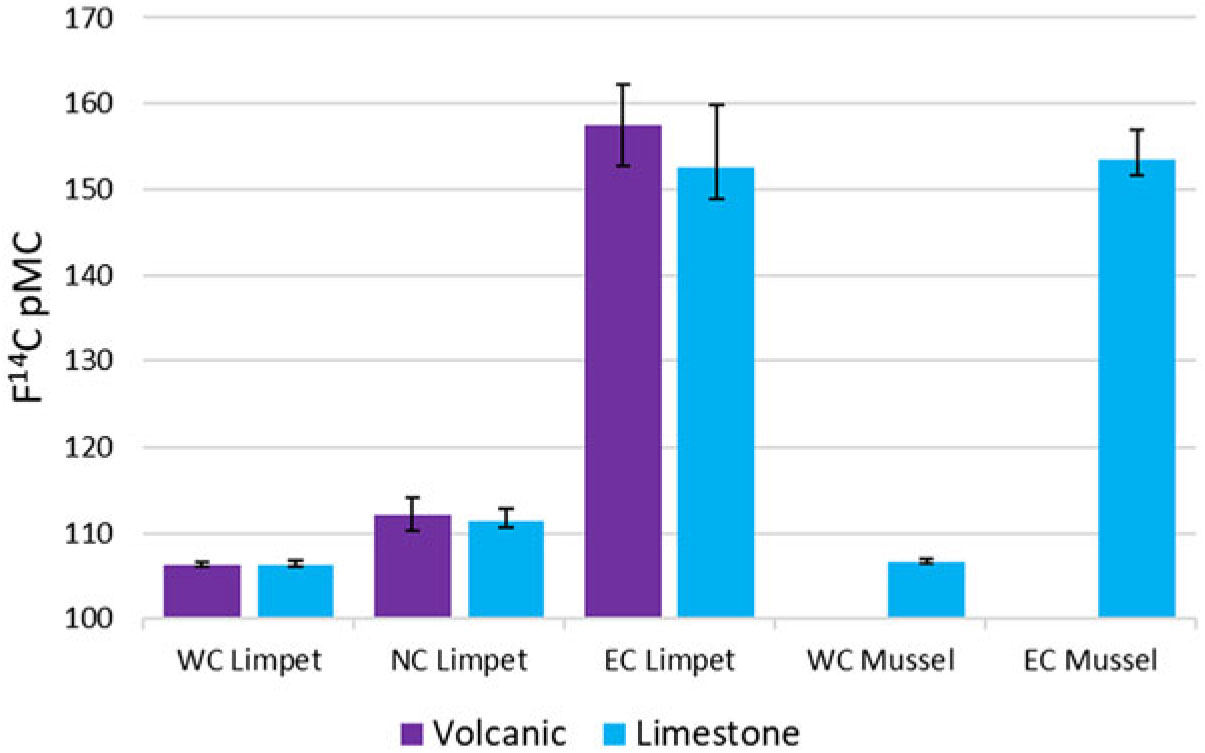
A summary of the F14C results are presented in Tables 1 and 2 and Figure 2. A full list of the results is presented in Tables 3 and 4 in the appendix. A significant difference between east coast compared to north and west coast results is apparent, with the east coast results showing an enrichment of 14C. A smaller enrichment of 14C is also apparent in the north coast results.
Table 1 14C-AMS measurements of P. vulgata from the coast of Ireland presenting the weighted means and standard deviations of the groups of measurements. The P values represent the analysis of differences between P. vulgata on limestone and basalt/granite substrates.

Table 2 14C-AMS measurements of M. edulis from the coast of Ireland presenting the weighted means and standard deviations of the groups of measurements. The P values represent the analysis of differences between P.vulgata and M. edulis on limestone and basalt/granite substrates.


Figure 2 14C activity of shell and algae samples as per cent modern carbon (pMC) with an associated error derived from the standard error of each group to express the natural variability in the shells.
The results from the P. vulgata shell samples indicate that there is no statistical difference between the volcanic and limestone locations. The results from the west coast shell samples indicate no statistical difference (ANOVA F 1,8 = 0.1.69E-06, P = 0.999). The results from the east and north coast shell samples also indicate no statistical difference between, but with lower confidence (ANOVA F 1,8 = 0.444, P = 0.52 and ANOVA F 1,8 = 0.748, P = 0.412). The results from the flesh samples indicate no statistical difference between volcanic and limestone locations on the west coast (ANOVA F 1,8 = 0.466, P = 0.514) but a significant difference on both east and north coasts (ANOVA F 1,8 = 15.40, P = 0.004 and ANOVA F 1,8 = 49.09, P = 0.0001). P values and standard deviations are included in Table 1. All the flesh samples are depleted in 14C compared to the shell samples, although the shell and flesh from the west coast are not statistically different (ANOVA F 1,8 = 3.88, P = 0.084).
Adding the results for the M. edulis samples also indicates that there is no statistical difference between the two species. Statistical analysis of the three groups of west coast limestone shell samples (ANOVA F 2,12 = 0.876, P = 0.441) and three groups of east coast limestone shell samples (ANOVA F 2,12 = 0.355, P = 0.708) indicates no statistical difference between species. The three groups of flesh samples also indicate no statistical difference on the west coast (ANOVA F2,11 = 0.278, P = 0.762) or on the east coast (ANOVA F 2,12 = 1.51, P = 0.259). P values and standard deviations are included in Table 2.
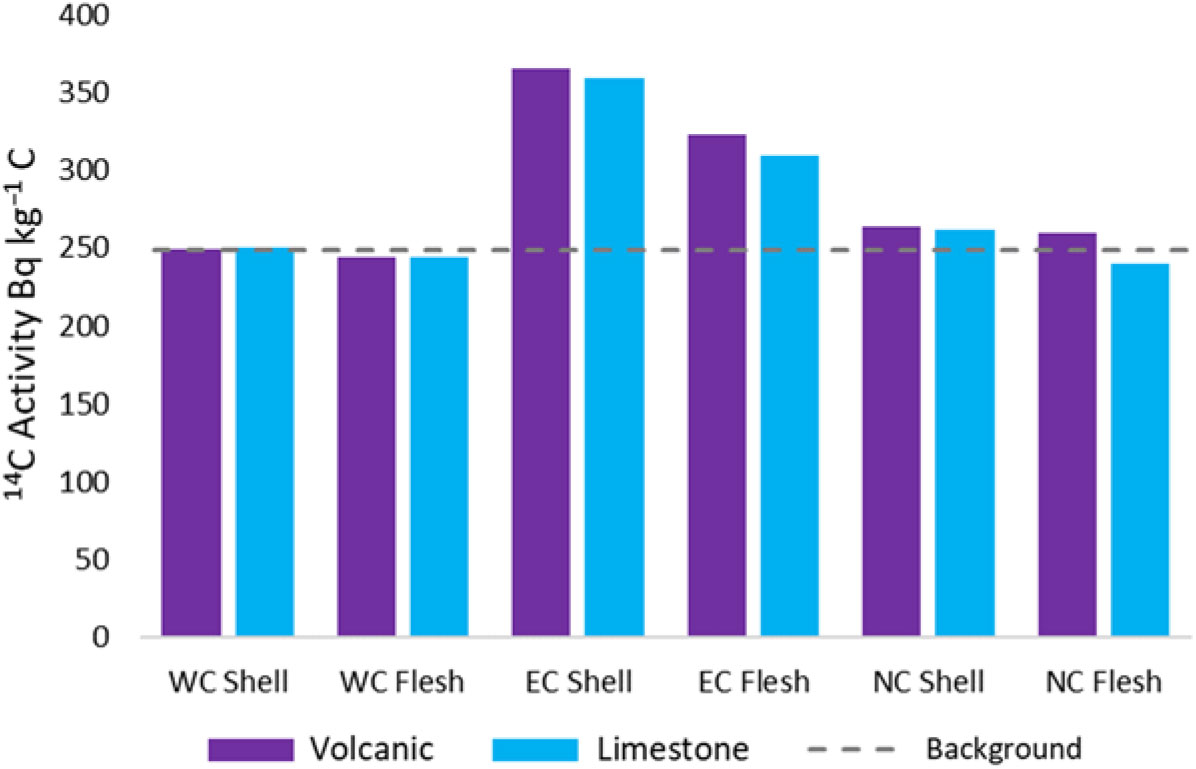
As a comparison to data collected from Irish Sea shells by Muir et al. (Reference Muir, Tierney, Cook, MacKinnon, Howe, Heymans, Hughes and Xu2016), the F14C results were converted to 14C activity in Bq kg−1 (Mook and van der Plicht Reference Mook and van der Plicht1999) and are presented in Figure 3. An existing “background” value of 249±1 Bq kg–1 C has been defined using shells collected on the west coast of Ireland (Muir et al. Reference Muir, Tierney, Cook, MacKinnon, Howe, Heymans, Hughes and Xu2016), representing natural production of DIC and atmospheric fallout. The west coast P. vulgata samples match this background value well. The east coast samples are also consistent with the enrichment of marine biota (flesh) samples due to Sellafield shown in the Muir et al. study, where values ranged between 275±1 Bq kg–1 C to 287±1 Bq kg–1 C. Actual levels of 14C enrichment vary with location and the level of discharge from Sellafield.

Figure 3 14C activity in P. vulgata as Becquerel per kilogram (Bq kg−1 C).
DISCUSSION
It is evident that the west coast samples are the only ones that do not show 14C enrichment and provide the most stable results. The east coast locations are in the northern half of the Irish Sea, where 14C discharges from Sellafield have a large effect. The north coast locations were assumed to be out of range of the Sellafield discharge, but the slight enrichment indicates that there is some influence, possibly from currents being funnelled between Rathlin Island and the north coast (Pingree and Griffiths Reference Pingree and Griffiths1980; Hill et al. Reference Hill, Brown and Fernand1997). The high standard deviations for the east and north coast samples could be explained by the varied discharge of 14C from Sellafield and, therefore, varied concentrations of 14C in the seawater DIC. The live samples that were collected were not aged, as the growth rings were not easily distinguishable, so uptake of Sellafield derived 14C may not be uniform across all the samples.
Statistically, the results indicate that there is no difference in the 14C activity of the shells of Patella vulgata living on limestone and those living on other non-carbonate substrates. This, along with the comparison to the filter feeder results from the Mytilus edulis, suggests that this species does not metabolise carbon from the limestone at these locations. The uptake of Sellafield 14C in the east and north coast samples highlights the influence of seawater DIC in both shell and flesh. Although Tanaka et al. (Reference Tanaka, Monaghan and Rye1986) and Dettman et al. (Reference Dettman, Reische and Lohmann1999) concluded that metabolic carbon contributed up to 40% of the carbon in shell precipitation, Gillikin et al. (Reference Gillikin, Lorrain, Bouillon, Willenz and Dehairs2006) revised this figure to approximately 10% depending on the species. The results of this study suggest that, for P. vulgata on the Irish coastline, this figure could be lower. The depletion in 14C activity of the limpet flesh is unexplained but may be due to input of older terrestrial organic carbon, possibly from fossil fuel-based compounds especially at the east coast site. On the west coast, the flesh is not significantly depleted compared to the shell. Further study of mollusks from this coast, in areas where there is no anthropogenic input, would provide more data to investigate any differences between shell and flesh.
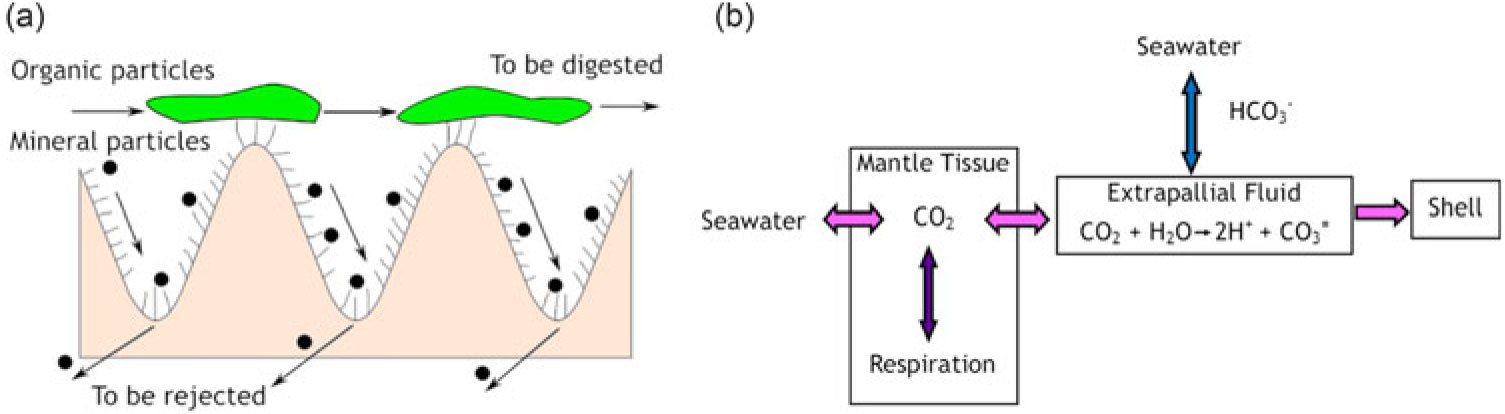
The biology and anatomy of the digestive system of P. vulgata may be key to understanding the results of this study. Inorganic and organic particles are sorted in the limpet digestive system. Inorganic particles are removed by ciliary action and only organic particles are directed to the digestive gland where most of the digestion occurs (Figure 4a). Absorption only occurs in the digestive gland and not in the rest of the digestive system as it would in some other mollusk species. The lowest pH value for the limpet digestive system is 5.4 (Yonge Reference Yonge1925). The solubility of limestone is very low at this pH value, but some dissolution may be possible. Carbonic anhydrase, an enzyme that catalyses the reversible CO3 to CO2 reaction, is present in limpet tissues but is not known to be secreted to the stomach (Rao Reference Rao1975; Lindberg and Dwyer Reference Lindberg and Dwyer1983). The uptake of CO2 for shell formation is mainly by diffusion or fluid exchange between seawater and the extrapallial fluid (the fluid in the space between the body and shell where mineral precipitation takes place). CO2 uptake for tissue formation is mainly from metabolic respiration but also by diffusion from seawater (McConnaughey and Gillikin Reference McConnaughey and Gillikin2008). A model for the uptake of CO2 is presented in Figure 4b. This model explains how the seawater DIC has the greatest influence on mollusk shell as well as a strong influence on the flesh and this is supported by the results of this study. The east coast results, reflecting the Sellafield enrichment, show that the CO2 in the mantle tissue is not only derived from respiration, but also from diffusion from seawater. The greatest influence of Sellafield 14C is in the shell, as the carbon used for the shell building is mainly derived from seawater.

Figure 4 (a) sorting area in the P. vulgata digestive system (b) model of likely transport routes for inorganic carbon, based on figure from McConnaughey and Gillikin (Reference McConnaughey and Gillikin2008).
CONCLUSION
This study aimed to determine if off-sets from seawater 14C were required when dating the shells of Patella vulgata that have fed on carbonate limestone substrates. Previous studies have suggested that foraging gastropods would metabolise old carbon, which would then be incorporated into their shells. The 14C of shells and flesh from P. vulgata, which fed on either substrate, were measured. The results indicate that the majority of shell carbon comes from ambient seawater DIC. Any effects on the 14C of P. vulgata from foraging on limestone are insignificant with respect to isotopic analysis. Shells from the east coast of Ireland are enriched by Sellafield derived carbon, causing higher variances within sample groups. However, these samples also indicate that there is no statistical difference between P. vulgata that fed on carbonate or volcanic substrates. The reason for this may be because of the primitive digestive system of P. vulgata. This study is relatively small, in respect to sample size, and a larger study would be needed to confirm these results. The study also indicated that the flesh is depleted in 14C compared to the shell. A larger study would provide more data so that this finding could be investigated further.
The shells of this species that are often found in archaeological sites should be suitable for radiocarbon dating, even if they are thought to be from carbonate geology, although further analysis of archaeological samples would be required to confirm these findings.
APPENDIX
Full Results of AMS Measurements
Table 3 14C-AMS measurements of P. vulgata from the coast of Ireland.

Table 4 14C-AMS measurements of M. edulis from the coast of Ireland. * Insufficient sample recovered following pretreatment.