


Introduction
Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) is one of the most important pests of cotton, pigeonpea, chickpea, maize, vegetables, fruits, and fruit trees worldwide (Sharma, Reference Sharma2001). The pest status of H. armigera is mainly the result of its physiological, behavioural, and ecological characteristics that enable it to survive in diverse habitats. Polyphagy, high mobility, high fecundity, and facultative diapause enable it to survive in adverse conditions (Fitt, Reference Fitt1989). Helicoverpa armigera is widely distributed in Asia, Africa, Australia and Mediterranean Europe (IIE, 1993) and causes an annual loss of over US$ 2 billion worldwide (Sharma, Reference Sharma2005). Having a status of high priority pest worldwide, it is routinely used in bioassays in the pesticide industry for monitoring the efficacy of pesticides, and as a tool for research on insect–host-plant interactions, insecticide resistance, transgenics, molecular markers, and biological, physiological and behavioural studies. To facilitate insect availability for use in a diverse array of research fields, it is desirable to manipulate various aspects of insect biology in order to have adequate numbers of insects at the appropriate stage of development for experimental purposes. Exposure of insects to temperatures below the typical rearing temperature of 27°C to slow down the rate of insect development can be exploited effectively for this purpose (Marrone et al., Reference Marrone, Ferri, Mosley and Mienke1985). Of the various developmental stages, eggs can be targeted for manipulating the insect biology in order to have adequate numbers of insects, when needed (Sharma et al., Reference Sharma, Taneja, Leuschner and Nwanze1992). Therefore, the present studies were undertaken to determine the effect of storage temperature on viability of H. armigera eggs at various temperatures.
Materials and methods
The experiments were conducted at the International Crops Research Institute for the Semi-Arid Tropics (ICRISAT), Patancheru, Andhra Pradesh, India. The H. armigera eggs were obtained from a laboratory culture maintained on chickpea-based artificial diet at 27±1°C, 65±5% RH, and 12 h photoperiod (Armes et al., Reference Armes, Bond and Cooters1992). The adults were released in a 30×30×30 cm cage, having nappy liners on the sides for oviposition, and fed on 10% sucrose solution in absorbent cotton. The nappy liners were replaced every morning. The eggs (0, 1, 2 and 3 days old) were exposed to −20, 0, 5, 10, 15, 20 and 27°C for 5, 10, 15 and 20 days. The temperatures 0 and −20°C were included in the studies to observe whether these lower temperatures could support H. armigera population build-up in, for example, cold Himalayan regions in Asia or whereever these low temperatures are experienced in regions where H. armigera is an important pest. There were 25 eggs in each replication, and replicated five times in a completely randomized design. The 0-, 1-, 2-, and 3-day-old eggs placed on a moist filter paper in a plastic cup (50 ml). For zero-day eggs, the nappy liners were changed at midnight (6 h before setting up the experiment). The eggs placed at different temperatures were examined daily for egg hatch. Observations were recorded at 12 h intervals on egg hatch and incubation periods at different temperatures.
Degree-day calculations and data analysis
The degree-days for different treatments were calculated with the following formula:
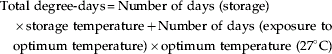
Data were subjected to analysis of variance (ANOVA) using GenStat version 8.0, to test the significance of differences among different treatments tested based on F-test at P=0.05. Since egg age, storage temperature and storage duration interact to influence egg hatch and incubation period, regression analysis was used to determine the combinations of these variables that would produce maximum percentage egg hatch. This model has a comparative advantage over the other regression models in the sense that it first computes the role of individual variable, and on the basis of that it gives best-fitted model in the forecasting. It uses several options, such as accumulated and pooled variables, forward selection and backward elimination, forward and backward stepwise regression, and/or all possible combinations to come out with best-fitted forecasting model. The Mallow Cp statistic regression model (Jackson et al., Reference Jackson, Lew and Woodson1995) was used to find the optimum storage time and temperature conditions to precise the forecasting of incubation period and percentage hatchability (Mallow's Cp statistic regression model=Dev/f+2×p−n where, Dev=deviance of the current model, f=dispersion parameter, p=number of fitted parameters of the current model, and n=number of units).
Results
The percentage egg hatch and incubation period were significantly influenced by egg age (P<0.001, F=28.8, df=3 for egg hatch; and P<0.001, F=1378.6, df=3 for incubation period), storage temperature (P<0.001, F=815.1, df=5 for egg hatch; and P<0.001, F=489.6, df=5 for incubation period), storage duration (P<0.001, F=206.9, df=4 for egg hatch; and P<0.001, F=827.4, df=4 for incubation period), and their interaction (P<0.001, F=2.6, df=34 for egg hatch; and P<0.001, F=16.5, df=34 for incubation period). Egg hatching of 0-, 1-, 2-, and 3-day-old eggs at 27±1°C varied from 69.6 to 98.0% (table 1). Egg hatch ranged from 8.0 to 96.8% across storage durations at 5 to 20°C (table 1). None of the 0-, 1-, 2- or 3-day-old eggs hatched when stored at −20 and 0°C. The 0- and 1-day-old eggs stored for 5 days at 5°C showed 34.4 to 60.8% egg hatching, while it was 8.0 and 8.8% in 2- and 3-day-old eggs stored for 10 days at this temperature (table 1). Two and 3-day-old eggs stored for 5 days at 15°C hatched in 1.0 to 1.2 days with 73.6 to 90.0% hatchability, when exposed to ambient temperature in the laboratory (27±1°C).
Table 1. Percentage egg hatch and incubation period of Helicoverpa armigera eggs stored at different temperatures for different durations.

* Controls kept at 27±1°C. –, Eggs hatched earlier than the storage duration in the incubator.
The regression model with the optimum Mallow Cp statistic for any of the identified linear and quadratic terms did not improve the precision of forecasting in egg hatch beyond 67.0%.
Model I=Egg hatch (%)=32.92+2.082 (temperature) (R2=66.7%).
Model II=Egg hatch (%)=30.57+1.560 (egg age)+1.996 (temperature)+0.058 (age×temperature) (R2=66.6%).
Model III=Egg hatch (%)=41.86−0.733 (storage duration)+1.807 (temperature)+0.019 (storage duration×temperature) (R2=67.0%).
For eggs of different age groups (0, 1, 2 and 3 days old) stored at different temperatures, the egg incubation period ranged from 5.4 to 20.9 days (table 1). The incubation period of different aged eggs stored at 5°C for 5 days did not differ from the eggs stored at 27±1°C. The 2- and 3-day-old eggs stored for 10 days at 5°C hatched 1.2 and 0.9 days later than the eggs stored at 27±1°C, respectively. The regression model for incubation period explained 86.9% variation (Incubation period (days)=2.775−1.054 (egg age in days)+0.296 (storage duration in days)−0.005 (temperature)−0.011 (age×storage)+0.012 (age×temperature)−0.039 (storage×temperature)+0.007 (age×storage×temperature) (R2=86.9%)). However, in the regression model with the optimum Mallow Cp statistic, the identified linear and quadratic terms were comparable to the full model in forecasting the incubation period (Incubation period (days)=2.612−0.701 (egg age)+0.258 (storage duration)−0.033 (duration×temperature) (R2=84.2%).
At constant storage temperatures, egg hatchability increased with an increase in temperature from 10 to 20°C (fig. 1a), while incubation period decreased across the egg age (table 2). Although the 0-day-old eggs could be stored for 20 days at 10°C, their hatchability was reduced to 36.0% (table 1). The 3-day-old eggs could not be stored at 10°C even for 5 days (tables 1, 2). The regression model with the optimum Mallow Cp statistic, the linear and quadratic terms precised the forecasting to 66.7% for egg hatch (Egg hatch (%)=22.82−6.355 (egg age)−0.771 (temperature)+0.216 (egg age×temperature) (R2=66.7%)), and 63.2% for incubation period (Incubation period=27.8+14.07 (egg age)+2.062 (temperature)−0.355 (egg age×temperature) (R2=63.2%)).
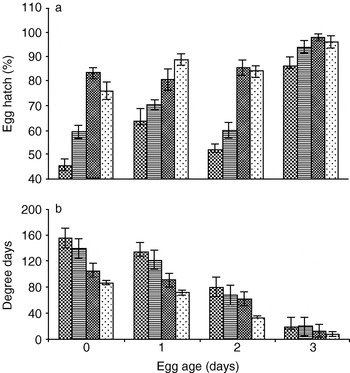
Fig. 1. Percentage egg hatch (a) and degree-days (b) required for Helicoverpa armigera eggs to hatch at different temperatures (  , 10°C; , 15°C; , 20°C; , control (27±1°C)).
, 10°C; , 15°C; , 20°C; , control (27±1°C)).
Table 2. Incubation period of Helicoverpa armigera eggs at different temperatures.
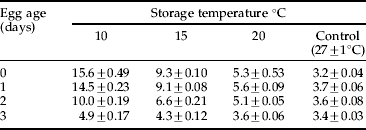
The day degrees required for egg hatching decreased with an increase in temperature from 10 to 27°C, and egg age from 0 to 3 days (fig. 1b). The day degree requirements were highest for 0-day-old eggs at 10°C (156.1 day degrees), being lowest (8.0 day degrees) at 27°C (fig. 1b). At 10°C constant temperature, the 0- and 1-day-old eggs hatched in 15.6 and 14.5 days (table 2) with egg hatchability of 45.6 and 64.0%, respectively (fig. 1a). Although the incubation period was higher, the hatchability was lower for 0- and 1-day-old eggs stored at constant 10°C (table 2), these can be stored for 10 days at 10°C, with hatchability of >75.0% (table 1).
Discussion
The responses of insects to environmental extremes have an important bearing on their life cycle, survival, and perennation to the next season. Cooling and freezing, and rates of temperature change influence physiological responses of insects, including H. armigera (Sinclair et al., Reference Sinclair, Vernon, Klok and Chown2003). Prediction of temperature-related cotton bollworm, Heliothis zea (Boddie) (Lepidoptera: Noctuidae) population (degree days) model suggested that growth and development can vary as much as 2 to 3 weeks from the normal time, depending on whether the temperature is above or below the average (Wilson & Barnett, Reference Wilson and Barnett1983). Physiological studies on the eggs of several dipteran insect species have demonstrated that egg development is temperature dependent, and the eggs absorb water during the early phases (0 to 3 days) and increase their volume up to 42.4% (Timm, Reference Timm1988). Insects adjust their life cycles to the seasonally variable environmental conditions so that growth, development, and reproduction coincide with favourable conditions (Wipking, Reference Wipking2000).
Thuja bark beetle Phloeosinus perlatus Chapuis (Coleoptera: Scolytidae) eggs stored at variable temperatures between 13 to 31°C (Matsuda et al., Reference Matsuda, Nakamura and Itao1997), and predatory water mite, Piona exigua Viets eggs at constant temperatures between 10 and 28°C (Butler & Burns, Reference Butler and Burns1989) increased the development time, and decreased the incubation period. However, in the case of the pentatomid predatory bugs, Podisus nigrispinus (Dallas) and Supputius cincticeps (Stål) egg incubation periods increased and egg viability decreased with an increase in temperature, 29°C being the optimum for egg development (Didonet et al., Reference Didonet, Zanuncio, Sediyama and Picanco1996). For eggs of the medfly Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) exposed to a lower threshold temperature (9°C) over 7 days and then transferred to 26°C, the survival rate was close to 50%, but all the eggs died between 17 to 18 days (Putruele et al., Reference Putruele, Garrido and Del Pino1997). In case of codling moth Cydia pomonella (Linnaeus) (Lepidoptera: Tortricidae), the eggs develop below 10°C with the base temperature of 1 or 2°C lower than 10°C (Saethre & Hofsvang, Reference Saethre and Hofsvang2002), but if the eggs are stored at 0°C for 14 days, the insect dies before the adult stage. Storage at 0°C for 21 days causes embryonic mortality (Yokoyama & Miller, Reference Yokoyama and Miller1989). Studies on southern corn rootworm Diabrotica undecimpunctata howardi Barber (Coleoptera: Chrysomelidae) indicated that egg age, storage temperature and storage duration have a significant influence on incubation period and percentage egg hatching. One-day-old eggs incubated at 15°C for 3 weeks was optimum for 50% egg hatching (Jackson et al., Reference Jackson, Lew and Woodson1995). There was a positive and linear relationship between temperature and developmental rates of the apple sawfly Hoplocampa testudinea (Klug) (Hymenoptera: Tenthredinidae) and sugarcane rootstalk borer weevil Diaprepes abbreviatus (Linnaeus) (Coleoptera: Curculionidae) eggs, with a thermal threshold of 6.9 and 12.0°C, respectively (Lapointe, Reference Lapointe2001; Graf et al., Reference Graf, Hopli and Hohn2002). The information on the influence of temperature on egg development and viability is a viable input for forecasting for egg hatch and timing of control measures against the neonate H. armigera larvae.
Results of these studies suggested that temperature is an important regulatory factor in the population dynamics of H. armigera. In the present study, day degrees required for hatching of H. armigera eggs decreased with an increase in temperature, and were highest for 0-day-old eggs at 10°C, and lowest at 27°C. Although the incubation period was longer, the hatchability was lower for 0- and 1-day-old eggs at constant 10°C. It is safer to store the 0- and 1-day-old eggs of H. armigera for 10 days at 10°C, they will hatch within 1.6 to 2.0 days after restoration at 27°C with a hatchability of >75.0%. This information can be used for forecasting egg hatching under different environmental conditions, and for egg storage at an appropriate temperature for planning and execution of experiments involving H. armigera. These studies too have implications on the estimation of H. armigera population build-up at different fluctuating and constant temperature regimes under field conditions and as a tool to judge infestation levels of migratory moths from warmer climates to regions with lower winter temperature. Apart from the application of this information on storage of H. armigera eggs at lower temperatures to delay the development and emergence of neonate larvae for use in studies on insect–host-plant interactions, bioassay of insecticides and transgenic plants, insecticide resistance, molecular markers, and biological, physiological, and behavioural studies; this information could also be used for predicting population dynamics of this pest to design appropriate control strategies.
