INTRODUCTION
The thick accumulation of ancient unconsolidated sediments that characterize many northern Siberian river valleys often are exposed in lengthy sections (sometimes of several km) whose sediments record a complex depositional and post-depositional history (e.g., Sher, Reference Sher1971; Sher et al., Reference Sher, Kaplina, Giterman, Lozhkin, Arkhangelov, Kiselyov, Kouznetsov, Virina and Zazhigin1979; Kuzmina et al., Reference Kuzmina, Sher, Edwards, Haile, Yan, Kotov and Willerslev2011; Murton et al., Reference Murton, Goslar, Edwards, Bateman, Danilov, Savvinov and Gubin2015). The dynamic nature of northern landscapes, particularly in settings where fluvial and permafrost processes intersect, makes unraveling this environmental history a greater challenge than in more temperate regions. This challenge, in part, is why scientists working in this region have traditionally taken an interdisciplinary approach to Quaternary investigations, an approach that involves both physical and biological considerations of past landscapes. One such example is that of the Berelyokh mammoth “graveyard,” (also known as the Berelekh geological site; 70º29′N, 144º02′E, elevation=20 m asl), a mass deposit of 8431 Mammuthus primigenius Blum. remains found in the lower Indigirka drainage of Northeast Siberia (western Beringia; Vereshchagin, Reference Vereshchagin1977; Fig. 1). The origin and age of this unusual concentration of mammoth bones has been the focus of intermittent study over the last ~40 years (Pitulko, Reference Pitulko2011).

Figure 1 Map key to sites: (1) Berelyokh River mammoth site. (2) Smorodinovoye Lake (Anderson et al., Reference Anderson, Lozhkin and Brubaker2002). (3) Berelyokh exposure, Kolyma basin (Lozhkin and Postolenko, Reference Lozhkin and Postolenko1989). (4) Jack London and Sosedneye Lakes (Lozhkin et al., Reference Lozhkin, Anderson, Eisner, Rovako, Hopkins, Brubaker, Colinvaux and Miller1993). (5) Elgennya Lake (Anderson et al., Reference Anderson, Lozhkin, Belaya, Glushkova and Brubaker1997). (6) Alut Lake (Lozhkin and Anderson Reference Lozhkin and Anderson2011). (7) Elikchan 4 Lake (Lozhkin and Anderson, Reference Lozhkin and Anderson2011). (8) Tanon River exposure (Lozhkin and Glushkova, Reference Lozhkin and Glushkova1997). (9) Ledovyi Obryv (south exposure), Main River, Anadyr Basin (Lozhkin et al., Reference Lozhkin, Kotov and Ryabchun2000). (10) Gytgykaii and Patricia Lakes (Anderson and Lozhkin, Reference Anderson and Lozhkin2015). (11) Lake El’gygytgyn (Shilo et al., Reference Shilo, Lozhkin, Anderson, White, Stetsenko, Glushkova and Brigham-Grette2001; Lozhkin et al., Reference Lozhkin, Anderson, Matrosova and Minyuk2007).
Several recent publications (Nikolskiy et al. Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010, Reference Nikolskiy, Sulerzhitsky and Pitulko2011; Pitulko Reference Pitulko2011; Pitulko et al. Reference Pitulko, Basilyan and Pavlova2014) have revitalized interest in this well-known site. These newer investigations aimed to clarify the relationship of the paleontological material to an archeological site located ~200 m downstream from the main geological section (Vereshchagin and Mochanov, Reference Vereshchagin and Mochanov1972). These latter studies, however, also considered the timing and mechanisms responsible for the deposition of the bone horizon. In the original study of the Berelyokh site, Vereshchagin (Reference Vereshchagin1977) postulated fluvial, erosional (e.g., solufluction), and cultural processes as causative factors. He concluded, however, that “given the analysis of all the taphonomic factors the “cemetery” formed as the result of the deaths of mammoths in polynas and transportation of the carcasses to an oxbow of the ancient Berelyokh River where silt covered the bodies (Vereshchagin, Reference Vereshchagin1977, p. 49; translation by Lozhkin). The latest research in the Berelyokh valley on nearby sections (the original geological site no longer exists due to river erosion) showed that the ages of the archeological and faunal locales did not overlap and, thus, a cultural explanation could be eliminated (Nikolskiy et al., Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010; Pitulko et al., Reference Pitulko, Basilyan and Pavlova2014). These studies agreed with Vereshchagin in looking to the river as the mechanism responsible for the formation of the mammoth horizon. They postulated, however, that either mammoth bones, previously deposited on upstream sand bars, or the bodies themselves were swept into a low energy, secondary channel of the main river or into an oxbow lake. Nikolskiy et al. (Reference Nikolskiy, Sulerzhitsky and Pitulko2011) further suggested that, given a new set of radiocarbon dates, such exceptionally high spring-floods perhaps were linked to the Bølling warm interval.
While much research of the Berelyokh exposures has focused on the occurrence of frequent and long-term changes in the river system, less emphasis has been given to the permafrost processes that currently shape and have shaped the landscape of the this region. Thermokarst activity, related to the thawing and refreezing of unconsolidated sediments, has the potential to be as hazardous (e.g., thaw sinks and thaw slumps; Harris et al., Reference Harris, French, Heginbottom, Johnston, Ladanyi, Sego and van Everdingen1988; Lozhkin and Anderson, Reference Lozhkin and Anderson2016) to animal populations as are floods. Such processes should be considered when seeking explanations for the formation of the bone bed. As to the site’s age, the assignment of the original Berelyokh faunal site to the Bølling interval (~12,600–12,000 14C BP, ~15,000–14,000 cal yr BP; Nikolskiy et al., Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010) is inconsistent with some of the data from the original investigation. In this paper, we provide an alternative interpretation about the origin and age of the mammoth deposit based on previously published and unpublished research done by Lozhkin at the original Berelyokh geological site. Although we review prior work, we focus on the palynological data, because they provide: (1) key information for constraining the age of the mammoth “cemetery;” (2) additional information about the paleolandscapes of the Berelyokh valley; and (3) expand understanding of regional vegetation and climate histories, especially for Marine Oxygen Isotope Stages (MIS) 2 and MIS 3, from a poorly documented area of western Beringia. We dedicate this paper to the memory of O.V. Grinenko and B.S. Rusanov, who were key in encouraging, organizing, and participating in the first interdisciplinary research of the Berelyokh exposure.
BACKGROUND
Previous research
The Berelyokh bone horizon was first noted in the scientific literature by Grigoriev (Reference Grigoriev1957) following fieldwork in 1947 (see Pitulko, Reference Pitulko2011 for a detailed review of early research at the site). The first major scientific expedition, however, was not launched until 1970 under the auspices of the Geological and the Permafrost Institutes (Yakutia) and the North East Interdisciplinary Science Research Institute (NEISRI, Magadan) of the Yakutia and Far East Branches of the USSR Academy of Sciences, respectively. B.S. Rusanov (Geological Institute) was the overall chief of the expedition with Vereshchagin (Zoological Institute, Leningrad) leading the paleontological research. This early work was followed up by interdisciplinary expeditions in 1971, 1972, 1980, 1987, and 1988. Lozhkin, a member of the original research team, collected palynological, radiocarbon, lithological, permafrost, and geomorphological data to be used in paleoenvironmental interpretations (Lozhkin, A.V., unpublished data). No archeologist worked on the initial expeditions, but Mochanov was later invited to examine the cultural material (Vereshchagin and Mochanov Reference Vereshchagin and Mochanov1972; Mochanov, Reference Mochanov1977). Radiocarbon ages, completed at several labs as a means of dating control, suggested the site formed during the Late Pleistocene. As mentioned above, research on the chronology, stratigraphy, taphonomy, and archeological associations continued in the twenty-first century using material from newly exposed sections and faunal material placed in a storage pit near the 1970–1971 excavation (Nikolskiy at al., Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010; Pitulko, Reference Pitulko2011; Boeskorov, Reference Boeskorov2013; Pitulko at al., Reference Pitulko, Basilyan and Pavlova2014). This latter research confirmed a likely Late Pleistocene age for the site, as well as suggesting flooding as being responsible for the mammoths’ demise and/or the bone concentration.
Modern vegetation
The Bereloykh River crosscuts the broad forest-tundra ecotone that spans much of far northern Northeast Siberia. Larix cajanderi is the only tree species in the lower Indigirka drainage (Karavaev, Reference Karavaev1958; Kremenetski et al., Reference Kremenetski, Sulerzhitsky and Hantemirov1998). The most common woody species within what is generally low-shrub tundra include Betula exilis, Duschekia fruticosa, and a variety of Salix species (e.g., S. polaris, S. glauca, S. alaxensis, S. nummularia, and S. pulchra). In contrast to areas farther south and east, where Pinus pumila is an important component of plant communities, this evergreen shrub is absent in the lower Indigirka drainage, reflecting both unsuitable soil conditions and a location near its climatic limits. The landscape near the Bereloykh site proper is open but does include widely dispersed forested areas with an understory of deciduous shrubs. Shrubs also border various watercourses (e.g., river and oxbow lakes) and can grow as dense thickets, which are scattered across the landscape.
The presence of active permafrost and fluvial processes in the Berelyokh valley has resulted in a mosaic of wet (e.g., Rubus chamaemorus) to dry-adapted (e.g., Asteraceae) plant communities. While such settings characterize valley microhabitats, they also are typical of permafrost disturbance and presence of frost polygon relief. For example, ice-wedge polygons and drained thermokarst basins support a variety of species dominated by Poaceae (e.g., Dupontia fisheri, Arctophila fulva, Alopecurus alpinum, Poa arctica) but with Cyperaceae (e.g., Eriophorum angustifolium), forbs (e.g., Saxifraga cernua, Potentilla hyparctica, Senecio congestus), and bryophytes (e.g., Sphagnum, Polytrichum spp., Marchania polymorpha; Billings and Peterson, Reference Billings and Peterson1980).
Permafrost
The area near the Berelyokh site has been and is currently being shaped by both fluvial and permafrost processes. The present-day river deeply meanders across the lowland, eroding cut banks, forming sand bars, and leaving abandoned channels that often develop into oxbow lakes. Many of these lakes eventually become thermokarst basins, which typically expand or drain as the lake waters melt the ice that is buried in the bordering banks. The local and regional presence of crosscutting ice wedges results in the development of a type of patterned ground called frost or ice-wedge polygon relief (Harris et al., Reference Harris, French, Heginbottom, Johnston, Ladanyi, Sego and van Everdingen1988). These polygons also can be a source of thermokarst activity, particularly when small ponds form in their centers (Billings and Peterson, Reference Billings and Peterson1980). While permafrost features typify higher fluvial terraces and interfluves, permafrost is lowered beneath rivers, lakes, and ponds because of the thermal properties of the water; thus, ice wedges and associated thermokarst activity do not characterize these settings.
The river valleys of northern Siberia, including the Berelyokh, usually include one or more terraces formed by the accumulation of channel and floodplain deposits. These terraces are rich in ice, including syngenetic ice wedges. The ice wedges are subject to melting when exposed (e.g., along a river’s cut bank) because of contact with comparatively warm air. As the ice melts, the sediments lose volume and form a mix of low-lying thermokarst ravines and higher “remnants” of the original sediments, termed baydzharaks (Komarov, Reference Komarov1930) or thermokarst mounds (Harris et al., Reference Harris, French, Heginbottom, Johnston, Ladanyi, Sego and van Everdingen1988). Such geomorphic forms were present at the Berelyokh geological site (Fig. 2). As melting continues, the baydzharaks diminish, the ravines join, and eventually can form a large “bowl” or thermocirque (also referred to as a retrogressive slide or retrogressive thaw slump; Harris et al., Reference Harris, French, Heginbottom, Johnston, Ladanyi, Sego and van Everdingen1988). Thermocirques also develop when ground ice melts, creating a structural weakness and subsequent downslope collapse of sediments. Thus, terraces along rivers in permafrost regions are not necessarily formed, at least not directly, by alluvial processes. Furthermore, these thermokarst processes result in a spatially and temporally complex sediment stratigraphy, leading to differences in the absolute depth of particular horizons and variability in the lateral extent and/or characteristics of some geomorphological features. The dynamic nature of these ancient northern landscapes provides potential difficulties in correlating exposures, even those separated by moderate distances. In the case of research along the Berelyokh River, the more recent investigations have relied heavily on radiocarbon results rather than lithological characteristics for correlating with the earlier work (Nikolskiy at al., Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010; Pitulko at al., Reference Pitulko, Basilyan and Pavlova2014).
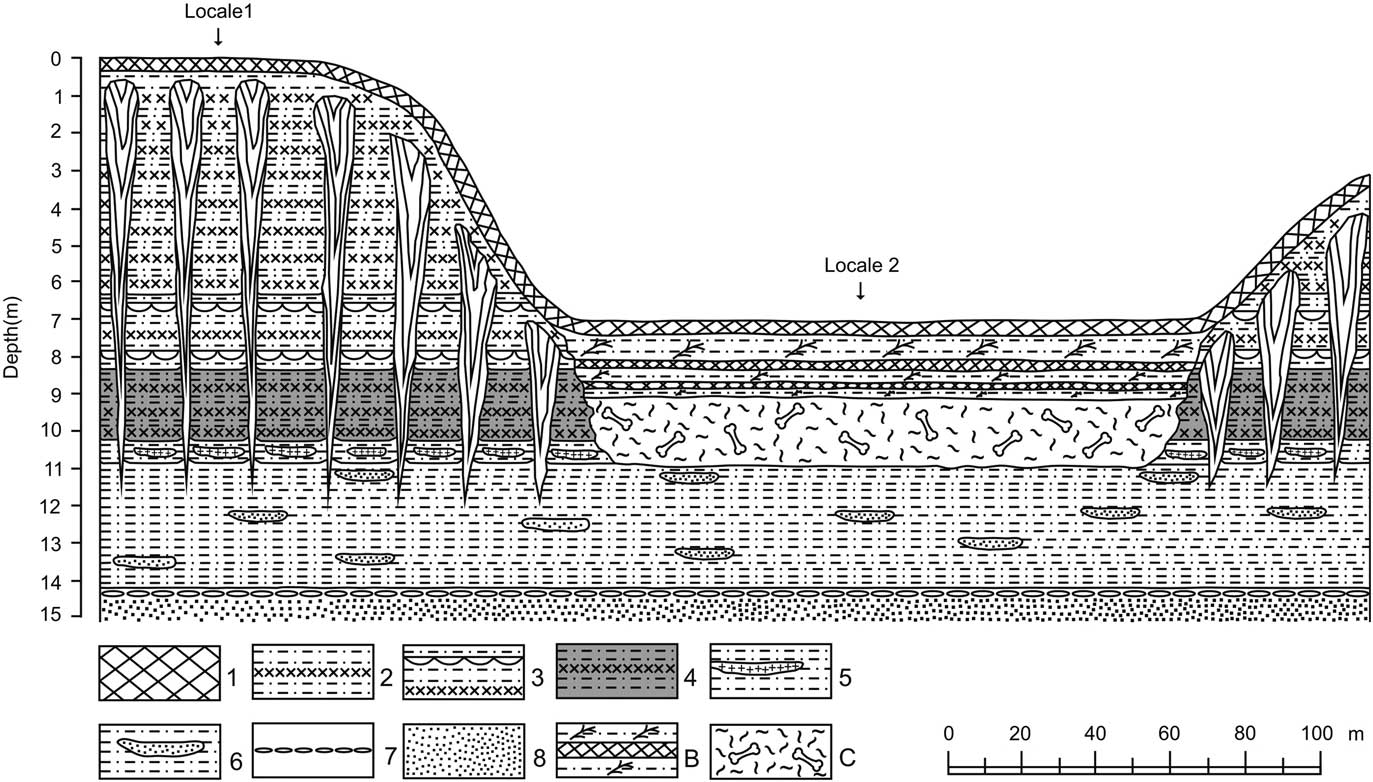
Figure 2 Schematic representation of the Berelyokh geological site, showing lithologic and permafrost features and locations of the bone horizon and sampling locales. Based on field notes of A.V. Lozhkin, 1970. Key to the lithologic units 1–8, B, and C are described in Table 1. Vertical exaggeration is 10×. Note that the area below the bone horizon (unit C) initially was covered by colluvium, which is not shown in the figure. Baydzharaks are located between the ice wedges.
The importance of fluvial systems in affecting the deposition and post-depositional histories of faunal material is well described for arctic rivers (e.g., Harington Reference Harington2011; Sher, Reference Sher1971). Yet, permafrost also has a role. One of the most evident influences occurs in areas dominated by baydzharaks and ravines, also known as yedoma relief. With continued ice melt, faunal material originally preserved in the baydzharak will erode as the mounds decrease in size. Bones are selectively collected in the ravines. As further erosion occurs, either related to river or thermokarst, bones will move to lower elevations and ultimately form “bone trains,” as noted by various researchers at Berelyokh (e.g., fig. 5 in Nikolskiy et al., Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010). The formation of a thermocirque, whether through baydzharak erosion and/or hillside failure, is another means by which faunal material might be concentrated. Thaw sinks (Hopkins, Reference Hopkins1949; Kartashov, Reference Kartashov1966) are yet a third, but perhaps lesser-known, example of the potential role of permafrost in the deposition and preservation of faunal remains. As surface waters penetrate the lower sediments, often aided by root systems below shrub thickets, ice wedges melt and form a lenticular or funnel-shaped pit below the topsoil or duff. These pits fill with ground water that is rich in minerals and organic residues. The pits expand over time, as the relative warmth of the subsurface water continues the thawing of buried ice. The water-sediment mixture is sufficiently viscous to support the overlying topsoil, thereby hiding the subsurface traps. Escape from these traps can be difficult, as Lozhkin can attest from personal experience, because they are often >1 m deep and the watery muck acts like quicksand.
Study site
The 1970 geological site was found exposed in a cut-bank in the middle reaches of the meandering Berelyokh River (see Pitulko, Reference Pitulko2011 for photographs of the Berelyokh valley). Terraces, which occur along this stretch of the river, are formed by unconsolidated Quaternary sediments, which are perennially frozen and have a variable active layer that is generally <0.7 m thick. Researchers have labeled these terraces differently, in part because their characteristics, particularly heights with which the terraces typically are labeled, vary. We use the designations given in the original research: 8-m (thermocirque) and 15-m (high floodplain). High floodplain terraces are characteristic of northern Siberian rivers. The term refers to that portion of the floodplain that is only intermittently water-covered. Thus, these types of terraces typically include fluvial silts indicative of flood events, stable surfaces (e.g., paleosols) developed in times when water is absent, and because they are mostly free of standing water a variety of permafrost features (Table 1; Fig. 2). Pitulko (Reference Pitulko2011) mentioned a third 20-m terrace formed by yedoma. Lozhkin (unpublished data), however, noted that this surface was not present at the Berelyokh geological site, but yedoma does form a ~20- to 30-m-high landform in other parts of the valley.
Table 1 Sediment description Berelyokh site, 1970 expedition (Lozhkin, Reference Lozhkin1977)

Approximately 200 m separate the sections studied by Lozhkin (Reference Lozhkin1977) within the 15-m (hereafter referred to as locale 1) and 8-m terraces (locale 2; Fig. 2). A striking feature of the 8-m terrace, in addition to the bone horizon, is that its surface is bowl-shaped, which is a characteristic of thermocirques. The bone bed, which was exposed in frozen sandy loam located at a height of 8 m above the 1970 river level, measured ~180 m long and 1.5–2 m thick. Bones were randomly oriented and unevenly distributed throughout the horizon, with some areas having large concentrations and other parts with few faunal remains. At the time of the 1970–1971 investigations, bones were also scattered along the beach and in the river shallows. According to Vereshchagin (Reference Vereshchagin1977), 8431 bones of mammoths were collected and studied, belonging to at least 140 individuals (Table 2). This collection includes specimens that were not in situ, as Vereshagin assumed that the bone horizon reasonably was the source of the skeletal remains on the beach. In addition to the bones, a well-preserved rear mammoth leg was discovered. Small clumps of mammoth hair, occasionally attached to pieces of skin, also were found throughout the horizon. Seven other animal species were identified, but they comprise only 0.7% of the faunal material.
Table 2 Faunal remains from the Berelyokh bone horizon, 1970–1971 expedition (Vereshchagin, Reference Vereshchagin1977)

METHODS
A pressurized hose system was used during the 1970 field season to both thaw the frozen sediments and to clear over-burden from the site. Each of the thawed exposures was carefully cleaned to reveal in situ sediments prior to sampling for palynological and radiocarbon samples, which were collected from: (1) a baydzharak of the 15-m-terrace; (2) sediments above, below, and within the bone horizon of the 8-m-terrace; and (3) mammoth hair and teeth found within the silty, sandy loam of the bone horizon. Additionally, modern pollen samples, which represent the main plant communities that characterize the modern Berelyokh valley, were gathered from soil and silty sediments near the excavations (see Supplementary Material). These samples represent both floodplain and soils so that different depositional settings can be compared (i.e., the possibility of redeposited or long-distance transport of palynomorphs is greater in alluvium, potentially making such samples less representative of the local vegetation).
Palynological samples were prepared using heavy liquids as described in Faegri and Iversen (Reference Faegri and Iversen1975). Pollen samples were mounted in glycerin and a minimum of 300 identified pollen grains were counted for each level, although pollen sums more typically were 500–1000 grains. Identification was done at 400× and 600×, using a LOMO (Leningrad Optical Mechanical Factory) microscope. Identification was aided with published keys (Grichuk and Zaklinskaya, Reference Grichuk and Zaklinskaya1948; Gladkova et al., Reference Gladkova, Grichuk, Zaklinskaya, Sauer, Pokrovskaya, Radzevich, Samoilovich, Sedova and Stelmak1950; Sladkov, Reference Sladkov1967), and a lab reference collection of slides and photographs of palynomorphs obtained from herbarium and field collections of Northeast Siberian plants.
Radiocarbon assays of material collected during the 1970 field season were done at NEISRI (MAG) and Leningrad University (Institute of Archeology [LE] and Geographical Research Institute [LU]), using a benzol-based, liquid scintillation method. A lower mammoth leg, exposed in the middle of the day during the 1970 excavation, was immediately placed in a pit with ice and shaded by a tent. The next day, Lozhkin flew to Yakutsk, where the leg was stored overnight in the permafrost tunnel (today, the leg is on exhibition at the Alexeev Mammoth Museum in Yakutsk). Here Lozhkin removed several ~20×30-cm-sized pieces from the loose flap of skin on the upper part of the leg. He next flew to Magadan, where the mammoth skin was refrigerated, subsampled while still frozen, and a radiocarbon assay begun. The material was unrefrigerated for only ~6 hours; thus potential contamination from thawing and/or long storage is not a concern. Radiocarbon dates of plant material from locale 2 were dated about one month later, but samples were kept cool in the field and refrigerated in the lab. All age ranges noted in the text represent median calibrated dates.
RESULTS
Detailed description of faunal material and sediment characteristics have been described elsewhere (Vereschagin, Reference Vereshchagin1977; Pitulko, Reference Pitulko2011), but only a brief description of palynological results have been available previously (Lozhkin, Reference Lozhkin1977; Anderson and Lozhkin, Reference Anderson and Lozhkin2002). Therefore, we describe the latter results in more detail here, with fauna and lithology summarized in Tables 2 and 3. We also correct an error in the original radiocarbon reporting.
Table 3 Key ages related to the Berelyokh mammoth horizon.

a Calibration was done using CALIB 7.1 (Reimer et al., Reference Reimer, Bard, Bayliss, Beck, Blackwell, Bronch Ramsey and Buck2013).
Fossil pollen spectra
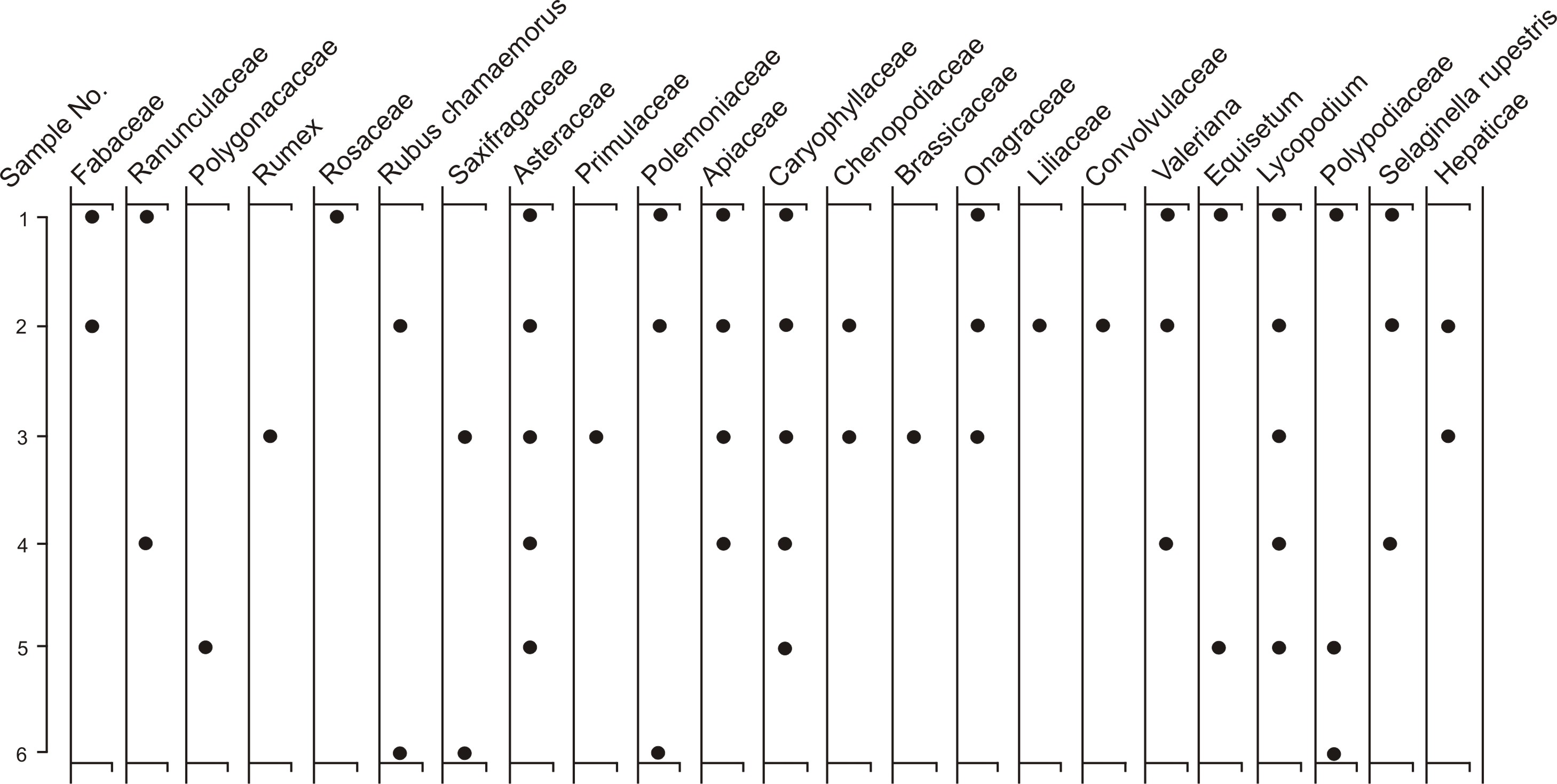
Three pollen zones were defined at locales 1 (Fig. 3) and 2 (Fig. 4). These zones show similar characteristics allowing the two sections to be correlated by their pollen stratigraphy. Pollen spectra from the mammoth remains (Fig. 5) do not display a similar zonation but show greatest similarity to zone BR2.


Figure 4 Pollen percentages from the mammoth bone bed (locale 2) horizon from the 1970 Berelyokh excavation with (a) major taxa and (b) minor taxa. Calibrated ages are listed in parentheses below 14C ages. Percentages are calculated as described for Fig. 2. See Table 3 for lithology. See Table 1 for sediment description of lithological units.

Figure 5 Pollen from mammoth remains within the bone bed from the 1970 excavations from the Berelyokh geological site with (a) major taxa and (b) minor taxa. Percentages are calculated as described for Fig. 2, and samples are presented in stratigraphic order. Key to samples: (1 and 2) mammoth skull; (3) silty loam from hair and skin preserved on front part of mammoth skull; (4) plant remains and hair attached to mammoth skull; (5) silty loam within a tooth from a lower mammoth jaw of mammoth; (6) silty loam and hair from a lower jaw of mammoth; (7) silty loam with hair found below a mammoth tusk; and (8) silty loam from an area with abundant mammoth bones.
Zone BR1 is most fully represented at locale 1 (1450–675 cm). Only the upper part of this zone is found at locale 2 (525–495 cm), because the lower portion of the exposure was covered by slope overburden. Zone BR1 at locale 1 is characterized by marked fluctuations in subsum values, particularly in the earliest part of the zone. Poaceae is the dominant pollen taxon with variable but significant percentages of Betula and Alnus. Cyperaceae and Artemisia, taxa typical of glacial assemblages, are generally <10%. Larix pollen is present in five samples in locale 1 and in one of the two samples at locale 2. At locale 1, the pollen composition of secondary herbaceous and shrub taxa is at its greatest for this record (20 types). The numbers of minor taxa are fewer at locale 2 (eight types).
Zone BR2 (locale 1, 675–520 cm; locale 2, 495–222 cm) is defined in more detail in locale 2 but both sites show similar patterns. Poaceae also dominates this zone, with a decrease in Betula and Alnus pollen. Artemisia pollen is a secondary herb component in the locale 2 record but a minor contributor in the locale 1 record. Cyperaceae percentages are <10%, and Larix occurs in a single sample in lower zone BR2 at locale 2. Caryophyllaceae dominates the 14 minor taxa at locale 2 but is absent at locale 1 (eight minor taxa).
Pollen zone BR3 (locale 1, 520–0 cm; locale 2, 222–0 cm) is marked by decreases in Poaceae pollen and increases in arboreal pollen and Sphagnum spores. Larix pollen continues to occur rarely, and minor taxa are somewhat decreased (locales 1 and 2 with 8 and 11 taxa, respectively). Of all zones, BR3 most closely approximates the modern spectra.
Like palynological spectra from locale 2 (bone bed 210–370 cm, or mid-to-upper zone BR2), the results from the faunal material are dominated by herb taxa, particularly Poaceae with lesser but significant pollen percentages of Artemisia, Cyperaceae, and Caryophyllaceae pollen. The decrease in Betula and Alnus pollen in samples 1–5 and, to a lesser extent, the generally higher Poaceae percentages also parallel trends in zone BR2 locale 2, as does the variety of minor herb types (12 types).
Site chronology and the age of the Berelyokh mammoth leg
At the time of Lozhkin’s original research, radiocarbon dates were available only from locale 2. The biostratigraphy, however, allowed for correlation with the pollen assemblages defined for locale 1 (Lozhkin, Reference Lozhkin1977). Pitulko (Reference Pitulko2011), Pitulko et al. (Reference Pitulko, Basilyan and Pavlova2014) and Nikolskiy et al. (Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010) provide comprehensive lists of radiocarbon dates for the various Berelyokh sections sampled over the past 40+ yr. Unfortunately, the age of the mammoth leg was incorrectly reported as 13,700 ± 400 BP in these papers, as well as earlier in Lozhkin (Reference Lozhkin1977) and Lozhkin and Parii (Reference Lozhkin and Parii1985). The correct date is 13,700 ± 80 BP (~16,800 cal yr; Table 3). The age from the skin seems to be given little consideration in the more recent discussions of the age of the Berelyokh bone bed. Possibly the sample is downplayed because of the supposedly large standard error or fear of possible contamination prior to the radiocarbon assay. Such contamination, however, is highly unlikely (see Methods).
DISCUSSION
We first describe the vegetation history of the Berelyokh site as it has implications for both the origin and age of the bone bed. The latter topics are discussed in the subsequent sections. We then consider the possible role of the Bølling warm interval in site formation.
Vegetation history
The BR1 pollen assemblage indicates the establishment of Larix-forest tundra early in the zone (locale 1; Fig. 3). This vegetation possibly was replaced by deciduous shrub tundra. The underrepresentation of Larix pollen does not rule out the continued presence of scattered stands of trees or even open forest-tundra as is found in the area today. High values of nonarboreal pollen, however, suggest a predominantly open landscape and likely a restriction of woody taxa to protected sites. These latter taxa included Betula, Alnus, and Salix shrubs, and possibly occurred as thickets in sheltered valley sites.
Like today, Pinus pumila was absent during zone BR1, even though the evergreen shrub shares similar temperature requirements with Larix, possibly indicating the lack of deep snow-cover required to protect the evergreen shrub from winter desiccation (Kozhevnikov, Reference Kozhevnikov1981). Although the absence of Pinus pumila may be primarily due to winter-dry conditions, it may equally be explained by the presence of cold, wet soils. The latter explanation would be consistent with the section sedimentology, which indicates the presence of permafrost conditions, documented by massive and segregated ice structures and syngenetic ice wedges (Table 1; Fig. 2). Minor taxa reflect a vegetational mosaic of wet (e.g., Rubus chamaemorus) to dry-adapted (e.g., Asteraceae) communities. Such settings characterize valley microhabitats and areas of permafrost disturbance and/or presence of frost-polygon relief. Although identification of palynomorphs is often not possible to species, the families and genera noted in zone BR1, as well as the other pollen zones at both sites, are consistent with thermokarst settings and characterize modern surface samples collected near the site (see Background Modern vegetation and Supplementary Material).
Greater percentages of Bryales spores in the upper part of zone BR1 accompanied by lesser values of Artemisia pollen suggest moister conditions during the latter part of the zone. The increase in Bryales spores also generally corresponds with more consistently high arboreal pollen, the latter representing taxa that would benefit from increased effective moisture. The spore increase also does not correlate with shifts in sediment type, indicating the change is not dependent on the depositional environment. The continued absence of Pinus pumila suggests that the shift in assemblage may reflect a change in summer conditions.
Variability in the BR1 pollen spectra is highlighted by the subsums with alternating intervals of increased woody and non-woody taxa. Similar pre-Holocene fluctuations in vegetation have been documented in other section and lake data from western Beringia (Anderson and Lozhkin, Reference Anderson and Lozhkin2001; Lozhkin and Anderson Reference Lozhkin and Anderson2011). These assemblages have been assigned to the Karginski interstade in Siberia and, more generally, to MIS 3, dating to 29,000–57,000 yr BP (Lisiecki and Raymo, Reference Lisiecki and Raymo2005). The assignment of zone BR1 to MIS 3 is consistent with radiocarbon dates of >42,000 14C BP (>45,600 cal yr BP) found below bone-bearing beds in various localities and latest Pleistocene ages for the overlying bone bed (Vereshchagin, Reference Vereshchagin1977; Lozhkin, Reference Lozhkin1977; Mochanov, Reference Mochanov1977; Pitulko et al., Reference Pitulko, Basilyan and Pavlova2014).
Zone BR2, more completely defined at locale 2 (Fig. 4), is characterized by conditions that were markedly colder and drier than present and with less temporal variation in vegetation types as compared to zone BR1. The decline in arboreal pollen taxa, particularly Betula and Alnus, suggests the predominance of arctic graminoid-forb communities. The vegetation was a mosaic of various tundra types (e.g., intermittent cover of herbs and Selaginella rupestris in elevated areas; Cyperaceae-moss and graminoid communities in wet to moderately-wet sites associated with poorly drained soils, permafrost activity, and snow beds; and Artemisia-xeric forb-graminoid communities in well-drained locales along river banks, terraces, and talus slopes). As in zone BR1, these minor taxa reflect plant communities adapted to disturbance related to permafrost and/or fluvial processes.
Protected locations along the Berelyokh valley would have provided suitable habitats for the persistence of Salix and Betula shrubs under the harsh, glacial conditions. It is unlikely, however, that Duschekia, which requires warmer, moister climates, was present. Thus, Alnus pollen probably represents long-distance transport (Lozhkin, Reference Lozhkin1977). Woody taxa likely continued to be most common along watercourses, where they likely formed thickets. Away from the river, they likely occurred as scattered subshrubs. The low percentages of Ericales pollen and Sphagnum spores suggest that moderately moist heath communities were limited in extent.
The dominance of Poaceae pollen is a feature of both lacustrine (e.g., Lake El’gygytgyn, Jack London Lake, and Gytgykaii Lake; Fig. 1) and non-lacustrine glacial assemblages (e.g., Berelyokh-Kolyma, Tanon River, and Main River-Ledovyi Obryv) of western Beringia during the Sartan/MIS 2 glaciation (27,400–12,400 14C yr BP; 31,990–14,600 cal yr BP; Anderson and Lozhkin, Reference Anderson and Lozhkin2001, Reference Anderson and Lozhkin2002). The secondary importance of Artemisia and Cyperaceae to Poaceae pollen in the Berelyokh sections also corresponds with other glacial spectra from western Beringia. An increase in Caryophyllaceae pollen seen in locale 2 is reminiscent of Lake El’gygytgyn stadial assemblages (Lozhkin et al., Reference Lozhkin, Anderson, Matrosova and Minyuk2007). Palynological data from the bone horizon (locale 2, 210–370 cm; Fig. 4) and the mammoth remains themselves (Fig. 5) are consistent with an interpretation of herb-dominated tundra dating to MIS 2. As in the sections, minor taxa associated with the faunal remains reflect a variety of microhabitats that are typical of glacial conditions. The 13,700±80 14C yr BP (~16,800 cal yr BP) date of the mammoth leg falls within MIS 2, suggesting the bone horizon was formed during the latest Pleistocene. Additionally, radiocarbon dates and palynological spectra in zone BR3 indicate that zone BR2 predates late-glacial climatic amelioration, which is marked in Beringia by a dramatic increase in Betula pollen (>30%), referred to as the birch zone (Hopkins, Reference Hopkins1982).
The decline in Poaceae and increase in arboreal pollen (particularly Betula and Alnus) and Sphagnum spores in zone BR3 mark the shift from glacial to interglacial conditions and the replacement of herb or herb-subshrub tundra by Betula-Alnus shrub tundra and open Larix forest in the Berelyokh basin. The presence of Larix pollen early in the zone (locale 2) is consistent with interpretations of a widespread deciduous forest/forest-tundra across Beringia during the late glaciation-early Holocene (Edwards et al., Reference Edwards, Brubaker, Anderson and Lozhkin2005). In upper zone BR3, increases of Alnus pollen, which are independent of significant sediment changes at the terrace site, suggest the growing importance of the shrub in the regional vegetation, where it likely was found in the understory of the Larix forest and in thickets along the banks of rivers and lakes. Higher percentages of Ericales pollen and Sphagnum spores indicate the local development of heath communities and generally mark greater effective moisture. The reduction in herb pollen (e.g., particularly Poaceae) reflects the spread of woody species as climate ameliorated. Minor taxa, however, continue to represent a variety of xeric to mesic settings. The continued presence of active permafrost processes, often associated with cool, wet soils, would limit both local and regional establishment of Pinus pumila.
Origin of the Berelyokh “cemetery”
The cause of death of large numbers of mammoths and the origin of such a dense bone concentration have been intriguing questions from the first stages of research at the Berelyokh site. While famine, epidemic, and drought, typical causes of death in modern elephant populations (Haynes, Reference Haynes1991), could bring about the demise of many individuals, these factors would not likely result in massive numbers of dead mammoths in a single location or provide a process that would concentrate the bones in an apparently short period of time. Given the site’s location and aspects of the lithology, Vereshchagin (Reference Vereshchagin1977) as well as the more recent investigations (Nikolskiy et al., Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010; Pitulko et al., Reference Pitulko, Basilyan and Pavlova2014) suggested that the river was the most likely mechanism ultimately responsible for the formation of the mammoth horizon. Details of site formation, however, differed between the earlier and later studies. In contrast to Vereshchagin (Reference Vereshchagin1977), who proposed the river simply washed the remains into an oxbow lake, Pitulko et al. (Reference Pitulko, Basilyan and Pavlova2014) proposed a more complex geomorphological history. It begins with the deposition of faunal remains on sand bars during MIS 3 or earlier. The river channel subsequently deepened, forming a high floodplain on the south bank of the river. Eventually, an oxbow formed at the “cemetery” site, when the main channel migrated to another part of the valley. Younger mammoth bones were carried by high water and accumulated in the oxbow, thereby forming the bone bed between 12,500–12,180 14C yr BP (~14,800–14,200 cal yr BP). A small portion of the fauna represents redeposition from interstadial deposits, thus giving a mix of ages reported for the site (Table 3). Floodplain deposits subsequently covered the oxbow sediments, with some of the bones later being eroded and deposited in younger oxbow lakes. Although changes in river channel and oxbow formation are characteristic of meandering rivers, such as the Berelyokh, Nikolskiy et al. (Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010), and Pitulko et al. (Reference Pitulko, Basilyan and Pavlova2014) suggest that exceptionally high flood event(s) would be needed to carry and concentrate such a large number of bones.
A fluvial interpretation, however, leaves several questions. The Indigirka lowland is a geomorphic feature that not only characterizes the modern geography but also persisted through the Late Pleistocene. The lowland, however, was more extensive during MIS 2, being ~1100 km to the north of the modern coastline (-50 m sea level) at ~15,000 14C yr BP (~18,000 cal yr BP) and ~1000 km to the north of the modern mouth of the Indigirka River (-37 m sea level) at ~12,000 14C yr BP (~14,000 cal yr BP; Lozhkin, Reference Lozhkin2002). Given the presence of such a broad and low-lying floodplain, it is difficult to image a spring flood (1) whose strength was not dissipated by spreading across the lowland; and (2) which would have retained sufficient strength to either drown the mammoths or carry away their remains. Similarly, the regional topography (i.e., no constrictions of the river channel by high, stable banks that could increase river flow) makes a flash flood, which could sweep away an unsuspecting herd, unlikely. The role of an oxbow channel in this history is also a bit puzzling in that, by definition, an oxbow is separated from the main channel, thereby requiring a truly exceptional flood to carry the bones or carcasses into an abandoned part of the river rather than just carrying the remains farther along the channel. If these floods were carrying bones previously deposited upstream, a more mixed assemblage might be expected, rather than one that is 99.3% mammoth. Additionally, it seems unlikely that the mammoth leg and the abundant clumps of hair and skin fragments found throughout the bone horizon would be preserved if not immediately buried in an anoxic environment, which would be difficult along a sand bar or potentially within an oxbow.
Although fluvial processes certainly have been an important element in the formation of late Quaternary landforms of the Berelyokh valley, the role of natural causes in the origin of the bone bed and/or cause of death may relate to non-fluvial landscape processes, as suggested for other mammoth sites in Eurasia. For example, sedimentological studies in the Don, Dnieper, and Desna regions of eastern Europe indicated that large accumulations of mammoth remains, associated with steep-sided topography within loess-covered valleys, were due to massive mud-slides triggered by water-saturated sediments (Lavrushin et al., Reference Lavrushin, Bessudov, Spiridonova, Kuralenko, Nedumov and Kholmovoy2015). Although not a massive burial, Lozhkin and Anderson (Reference Lozhkin and Anderson2016) commented on the dangers of Late Pleistocene permafrost landscapes for fauna in western Beringia, as in the entrapment and subsequent drowning of the baby mammoth Dima in a buried thermokarst pit in the upper Kolyma region.
We propose an alternative to the fluvial-based interpretation for the formation of the Berelyokh bone bed, suggesting, instead, that thermokarst activity was central to the simultaneous destruction of a large group of animals and the formation of the mammoth horizon. As described above (Background, Permafrost), the formation of thaw sinks, when surface waters gradually melt underlying ice wedges, is a characteristic landform in high-floodplain terraces within the Yana-Indigirka-Kolyma lowlands. The sinks expand over time, often aided by the penetration of the lower sediments by root systems (in the case of the Berelyokh site, these are likely Salix and/or Betula). The mix of organics, silt, and water is sufficiently viscous to support a surficial layer of topsoil or duff, thereby hiding the subsurface traps.
We suggest that such a process occurred in the ancient Berelyokh valley. In this scenario, a series of smaller traps or perhaps a larger pit formed below the MIS 2 surface of the 15-m terrace. This high floodplain terrace, with its open, gentle terrain, would be a likely route for travel and/or an area for grazing within the river valley for the mammoths. As they were crossing or grazing within the area, the topsoil gave way, plummeting the animals into the water and muck-filled depression(s). The sediments within these thermokarst sinks are analogous to water-rich fluvial silts, which Guthrie (Reference Guthrie1990) has described as entrapping large animals, such as moose, in modern Alaskan floodplains. Such sediments would provide an equally difficult setting for even strong animals to escape. In the case of the Bereloykh site, we suggest that the mammoths’ struggles as they attempted to free themselves from the trap(s) destabilized the slope, causing the hillside to collapse and burying the animals. Conversely, the presence of the sinks themselves perhaps resulted in a weaken hillside that underwent a massive slope failure and catastrophic collapse under the weight of the mammoths. Such failures are typical of retrogressive thaw slumps or thermocirques (Washburn, Reference Washburn1979; see Murton et al., Reference Murton, Edwards, Lozhkin, Anderson, Bakulina, Bondarenko, Cherepanova, Danilov, Boeskorov, Goslar, Stanislav, Gubin, Korzun, Lupachev, Savvinov, Tikhonov, Tsygankova and Zanina2017 for an extreme examples of such a feature). Unfortunately, thermokarst traps themselves, compared to thermokarst lakes, for example, do not leave a characteristic sedimentary “signature.” The presence of active permafrost processes along this portion of the Berelyokh valley, however, is clear from lithological (e.g., the presence of syngenetic ice wedges or thermokarst lake deposits found in the more recent studies near the Berelyokh site) and palynological data (persistence of tundra vegetation, taxa associated with frost polygons). The bowl-like surface configuration of the 8-m terrace also suggests a permafrost origin, related to the formation of a thaw slump. Chronological evidence that this was a single event is considered below.
Age of the Berelyokh “cemetery”
The history of radiocarbon dating of the Berelyokh site might be considered as complex as the paleoenvironmental record itself, including: (1) materials collected and dated in 1970–1971; (2) materials collected in 1970–1971, placed in an underground storage pit at the geological site or archived at the Zoological Institute Leningrad, and then dated more than 40 years later; and (3) materials collected during field work in the twenty-first century, which provided dates for several sections located ~1 km distant from the original geological site (see Pitulko, Reference Pitulko2011, and Pitulko et al., Reference Pitulko, Basilyan and Pavlova2014 for complete listing of Berelyokh dates). The results of these multiple analyses indicate the faunal material falls into two age groups: the latest MIS 2 and MIS 3. The latter group includes two mammoth samples and three from other species (Table 2), all of which were dated after long storage. Although the results might be questioned because of the delayed analyses, deposits of MIS 3 age occur in this portion of the Berelyokh valley, as indicated by the palynological data from the original geological site and the non-faunal radiocarbon dates from the more recently studied sections. We agree with Pitulko (Reference Pitulko2011) that it is most likely that the interstadial ages are from specimens that were redeposited through fluvial erosion and/or during the formation of the thermocirque (this paper).
Five radiocarbon dates from the mammoth leg, mammoth tusks, and unidentified plants provided the original age model for the site. The plant macrofossils were from a paleosol layer above the bone deposit, and indicated that the “cemetery” pre-dated ~11,900 14C yr BP (~13,800 cal yr BP). These ages are similar to the ~11,830 14C yr BP (~13,700 cal yr BP) date obtained from Salix roots and branches found in the same stratigraphic layer but not in association with the area of the greatest bone accumulation (Lozhkin, Reference Lozhkin1977). Dates from the leg and tusks suggest the mammoth remains were deposited during late MIS 2, probably some time between ~13,700–12,900 14C yr BP (~16.800–15,600 cal yr BP). These ages are in agreement with the palynological data from the Berelyokh site, which indicate the mammoth remains accumulated under cool, dry glacial conditions (Fig. 4 and 5). The Berelyokh chronology and biostratigraphy also parallel Beringian-wide trends for the transition from full-glacial herb-dominated tundra to late-glacial shrub tundra (Hopkins, Reference Hopkins1982).
The chronostratigraphic importance of the palynological data from the mammoth horizon might be questioned, if the samples originated within a slump deposit where mixing of different-aged sediments is likely. The herb-dominated assemblage from the bone bed, however, is consistent with spectra from undisturbed MIS 2-aged sediments (unit D, zone BR2) and contrasts strongly to the shrub-dominated spectra of the late glaciation/early Holocene (zone BR3). Samples taken from the hair and teeth of the mammoth also are dominated by nonarboreal pollen, and add further evidence that the deposition of the mammoth remains occurred during glacial times.
Nikolskiy et al. (Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010) postulated an age of 12,400–12,200 14C yr BP (~14,600–14,300 cal yr) for the bone bed, based on radiocarbon assays of nine mammoth mandibles (~12,700–11,900 14C yr BP; ~15,100–13,800 cal yr BP) from Vereshchagin’s storage pit and analysis of other sample types from the more recent investigations. Pitulko et al. (Reference Pitulko, Basilyan and Pavlova2014) proposed a generally similar age for the mammoth horizon (12,600–12,180 14C yr BP; ~15,000–14,200 cal yr BP), relying primarily on dates of non-mammoth fauna (one sample from a mammoth) and of plant remains collected from the more recently analyzed sections. Thus, the results of both studies suggested that the “cemetery” formed more recently than proposed in the original age model. All chronologies, however, are consistent with an event that predates the ~11,900 14C yr BP (~13,800 cal yr BP) paleosol.
In discussing their revised chronology, Nikolskiy et al. (Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010, p. 21) questioned the precision of the dates done in the 1970s, commenting that ages obtained from different laboratories “may introduce a significant distortion if used together as a single data set.” They also cited conclusions drawn by Sher and Plakht (Reference Sher and Plakht1988), who questioned the accuracy of dates that assigned “warm” Late Pleistocene paleobotanical horizons from Siberia to MIS 3. Sher and Plakht (Reference Sher and Plakht1988) postulated that the long-term storage of organic material in unrefrigerated laboratories caused contamination that resulted in finite rather than infinite dates and that these horizons were more correctly of MIS 5 age. Subsequent research on continuous lake sediments (Lozhkin and Anderson, Reference Lozhkin and Anderson2011) showed that: (1) intervals of climatic amelioration did occur during the Late Pleistocene interstade; (2) ages from MIS 3 sections are likely correct; and (3) concerns about contamination and cross-lab comparisons were overstated. Certainly, contamination was not an issue in the 1970 excavation, as the mammoth leg was processed almost immediately (see Methods) and other organic material was not stored for any length of time.
Nikolskiy et al. (Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010) also remarked that collagen dates from permafrost regions are usually reliable, citing Vasilchuk et al. (Reference Vasilchuk, Punning and Vasilchuk1997), but collagen dating is not necessarily straightforward. For example, Higham et al. (Reference Higham, Jacobi and Ramsey2006) noted that collagen dates, especially from older samples, are highly dependent on analytical technique and post-depositional influences on collagen preservation. Vasilchuk et al. (Reference Vasilchuk, Punning and Vasilchuk1997) also commented on the greater dating reliability of frozen carcass remains as compared to bones, because of greater taphonomic issues related to skeletal material deposited in dynamic landscapes. Although the Berelyokh samples are clearly from a permafrost landscape, the bones would not necessarily have been frozen quickly if initially deposited on sand bars or on banks of oxbow lakes/abandoned channels. Furthermore, sediments underlying lakes and rivers have significantly deeper permafrost levels, which could further delay any freezing of the bones, as compared to other terrestrial settings. The bones taken from the 1970–1971 storage pit of course, had been removed from their original depositional environment and, even though this was only for several decades, some skewing of ages may have occurred. Additionally, skeletal material was collected from the beach adjacent to the site and stored with in situ material excavated from the exposure. The inability to separate samples from different collection areas adds another possibly confounding factor.
The age ranges of the above chronologies encompass a period when conditions were shifting from the cold, dry full-glaciation (herb tundra) to the more moderate late glaciation (Betula shrub tundra). Unfortunately, only the original investigation at Berelyokh included palynological analysis. The available pollen spectra are consistent with a late full-glacial age, and given the pollen stratigraphy from sites in northern western Beringia, indicate the bone bed most likely formed prior to 12,400 14C yr BP (~14,600 cal yr BP; Anderson and Lozhkin, Reference Anderson and Lozhkin2002). Regional biomization also supports this conclusion indicating that western Beringia was populated by dry tundra and dwarf shrub tundra at 15,000 cal yr BP (~12,600 14C yr BP) with areas of low to high shrub tundra established by 14,000 cal yr BP (~12,000 14C yr BP; Binney et al., Reference Binney, Macias-Fauria, Edwards, Anderson, Lozhkin, Kaplan and Nield2017). Thus, both local and regional paleobotanical data suggest that the mammoth “cemetery” formed during the earlier interval defined by the radiometric age-ranges. While the more recent studies have added significantly to the number of dates from the Berelyokh exposures, we consider the radiocarbon date on the mammoth leg to be more reliable than the bone dates because: (1) the leg has fewer problems related to radiocarbon method; (2) post-mortem alteration is less likely; and (3) its age is more consistent with the palynological data.
The wide spread in the radiocarbon ages, nonetheless, has implications. Specifically, an age model that includes all radiocarbon results indicates that the mammoth “cemetery” accumulated over hundreds or perhaps even thousands of years. Such an age range, of course, would argue against a catastrophic event related to either permafrost processes or several closely timed floods. The suite of radiocarbon dates, however, clearly is not without issues related to analytical methods and the post-depositional history of the samples. Again, we suggest that the mammoth leg is the least problematical of all these samples.
The occurrence of other species in the Berelyokh collection (Table 2) might also indicate that the faunal material represents more than a single catastrophic event. As noted above, the presence of some redeposited specimens is strongly suggested by the MIS 3 radiocarbon dates (e.g., Panthera spelaea, Coelodonta tichorhinus) and may relate to collections made along the beach. In contrast, the frozen Gulo gulo mummy likely is contemporaneous with the mammoths, as the body was found laying on its back, crushed under a mammoth skull. It also is probable that the Gulo gulo skeletal remains represent animals that scavenged the site. The other species we suggest most likely represent re-deposition of older (although not necessarily from MIS 3) bones. Even with the occurrence of these species, the preponderance of mammoth bones (99.3%) in the Berelyokh assemblage is consistent with the proposed thermorakarst scenario.
The role of climate in the formation of the Berelyokh “cemetery”
A final consideration is the role of climate in the genesis of the Berelyokh “cemetery.” Nikolskiy et al. (Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010, Reference Nikolskiy, Sulerzhitsky and Pitulko2011) suggested that the large-scale floods experienced in the Berelyokh valley are related to warmer and snowier conditions during the Bølling interval (12,600–12,000 14C yr BP; 14,600–14,000 cal yr BP). This interpretation disagrees with Lozhkin (Reference Lozhkin1977), who argued that the bones accumulated under relatively cool, dry conditions.
Paleobotanical data from continuous lake records in western Beringia indicate that warming, as reflected in the shift from full-glacial herb tundra to late-glacial shrub Betula tundra, occurred regionally as early as ~12,500 14C yr BP (~14,800 cal yr BP) and as late as -12,000 14C yr BP (14,000 cal yr BP) with most ages falling at ~12,400 14C yr BP (~14,600 cal yr BP; Anderson and Lozhkin, Reference Anderson and Lozhkin2002). The Bølling oscillation is essentially synchronous with the broader herb-birch zone transition, and the widespread establishment of Betula shrub tundra points to a rise in summer temperature and precipitation. Little independent evidence, however, exists to indicate that increased precipitation, particularly snow depth (the greatest source for spring floods), was sufficient to cause catastrophic flooding. Analog-based climate reconstructions from Lake El’gygytgyn (Fig. 1) show only a moderate increase in mean annual precipitation (Melles et al., Reference Melles, Brigham-Grette, Minyuk, Nowaczyk, Wennrich, DeConto and Anderson2012) during early MIS 1 (i.e., early birch zone), with the greatest increase occurring in summer (Lozhkin et al., Reference Lozhkin, Anderson, Matrosova and Minyuk2007). These reconstructions are based on pollen percentages, which suggest shrub Betula was present on the Chukchi Plateau as early as ~12,800 14C yr BP (~15,300 cal yr BP). Pollen accumulation rates, however, indicate the plant was not abundant until ~9900 14C yr BP (~11,300 cal yr BP). Shrub Betula likely was more common in the lowlands and along river valleys, such as the Berelyokh, as compared to the upland El’gygytgyn site. Nonetheless, the difference in timing of Betula expansion, if the Lake El’gygytgyn age model is correct, suggests that regional climates probably remained relatively dry until the early Holocene. Additionally, climate models indicate that conditions were cooler and drier than present across Beringia before ~12,000 14C yr BP (~14,000 cal yr BP; Bartlein et al., Reference Bartlein, Anderson, Edwards and McDowell1991).
Even if the Berelyokh “cemetery” was formed during the late-glacial transition, a permafrost, as opposed to a fluvial, origin still seems more likely. For example, Edwards et al. (Reference Edwards, Grosse, Jones and McDowell2016) noted that thermokarst activity rapidly increased in the Yukon Flats of central Alaska following late-glacial climatic amelioration. If the Berelyokh deaths occurred during post-glacial warming, the ice-rich silts would have been susceptible to increased thermokarst activity, and the terraces perhaps presented a greater danger to the large animals. As thermokarst activity (e.g., thawing of ice wedges) also occurs under cold, dry conditions (Shilo et al., Reference Shilo, Lozhkin, Titov and Shumilov1983), the creation of the bone bed does not require a Bølling-type warming.
Although we believe that the formation of the Berelyokh “cemetery” occurred under glacial conditions, the dating of the faunal remains represents a broad range of ages that encompasses the herb zone and herb-birch zone transition. If falling in the younger interval, we propose that the bone bed was deposited during the early part of the glacial-interglacial transition based on the available palynological data. Because the late-glacial climate history of western Beringia does not include the fluctuating climates described from northwestern Europe (except perhaps for a weak Younger Dryas signal; Kokorowski et al., Reference Kokorowski, Anderson, Mock and Lozhkin2008), we suggest the use of Bølling is inappropriate for this region, and the interval may be better referred to simply as the late-glacial warming.
Regardless of the climatic conditions, the Berelyokh “cemetery” provides a cautionary tale about the dangers of dynamic northern landscapes. Edwards et al. (Reference Edwards, Grosse, Jones and McDowell2016) warn against naively projecting the nature of changes to thermokarst landscapes under global warming scenarios based on paleo-studies. Any future alterations of these regions, however, will not only potentially shift important geochemical processes (e.g., release of CO2 and CH4), but could provide an increasingly uncertain land surface, especially for large arctic and subarctic mammals.
CONCLUSIONS
The Bereloykh mammoth “cemetery,” containing >8400 mammoth bones, has been the focus of multiple investigations into the age and origin of this remarkable stratigraphic horizon. Most recently, Nikolskiy et al. (Reference Nikolskiy, Basilyan, Sulerzhitsky and Pitulko2010, Reference Nikolskiy, Sulerzhitsky and Pitulko2011) and Pitulko et al. (Reference Pitulko, Basilyan and Pavlova2014) postulated that the bone horizon was created by one or more flood events that occurred during the late-glacial Bølling warming, when a marked increase in snow accumulation resulted in catastrophic spring flooding. However, the abundance of permafrost features found throughout the 8-m and 15-m terraces at the Berelyokh site and the persistence and dominance of arctic tundra from MIS 3 to the present are indicative of a terrain shaped both by fluvial and permafrost processes that can provide an alternative explanation for site formation.
Thermokarst relief is a terrain often associated with ice wedges, frost polygons, and thermokarst lakes. Thaw sinks or thermokarst traps, however, originate with the subsurface melting of ice wedges and the development of lenticular pits and are both lesser-known and poorly studied. As the buried ice melts, a viscous mix of sediment and ground water that is sufficiently dense to support an upper layer of soil fills the holes, thereby forming a subterranean trap. We suggest that a mammoth herd traveling or grazing across a high floodplain (now the 15-m terrace), where the concealed traps were formed, broke through the surface and became entrapped in the underlying muck. Their struggles resulted in the local collapse of the terrace slope (forming a thermocirque on the present-day 8-m terrace) and burial of the mammoths. Alternatively, the sinks caused a weakening of the hillside, which gave way catastrophically under the weight of the mammoths and subsequently buried the animals.
While fluvial influences cannot be totally dismissed, several factors suggest a thermokarst explanation may be more parsimonious. Specifically:
(1) The presence of a well-preserved mammoth leg, the abundance of clumps of hair scattered throughout the bone horizon, and numerous pieces of skin still attached to the bones indicate a depositional environment favorable to the preservation of organic material. Such preservation would not occur in scenarios where bones were deposited along sand bars and/or redeposited following flooding.
(2) A permafrost mechanism would account for the surficial impression of a thermocirque on the 8-m terrace and for a decrease in the level of relief of ≥7 m between the two terraces. No fluvial mechanism has been proposed to specifically account for either of these features at the original geological site. Additionally, the deposits above the bone horizon suggest that this portion of the Berelyokh terrace was removed from the fluvial system, as indicated by the accumulation of sediments into the bowl-like depression through intermittent solifluction and the establishment of stabilized, organic-rich surfaces.
(3) The random orientation of the bones and lack of layered sediments seem to be as or more suggestive of collapse and slumping of terrace sediments than does deposition of the bones in an active river or oxbow channel.
(4) The entrapment and collapse argument might more reasonably account for the location of the bone horizon in what was already a terrace well-elevated above the river level, even in pre-Holocene times.
(5) The broad expanse and gentle gradient of the Indigirka lowland during the last glacial maximum/late glaciation decreases the likelihood that a flood of sufficient strength to carry away remains of live mammoths would have occurred. Additionally, the geomorphology suggests even waters of an exceptional flood would quickly spread across the lowland, dissipating its strength and making deposition of 8400 bones in oxbows or a high floodplain terrace unlikely.
(6) A Bølling oscillation has not been documented in any area of western Beringia, making this climatic event an improbable reason for site formation. In contrast, the thermokarst scenario is not dependent on climate variation, as these permafrost processes occur during glaciations and interglaciations.
Despite a suite of radiocarbon dates from the original 1970–1971 investigation and from subsequent research of sections ~1 km distant, the age of this stratum is still not definitive. We consider the radiocarbon date on the mammoth leg found within the bone bed itself to be more reliable than the bone dates from the geological site, because: (1) the leg has fewer problems related to radiocarbon method as compared to bone collagen dating; (2) post-mortem alteration of frozen skin is less likely than in skeletal specimens, most of which were in storage for >30 years prior to dating; (3) the depositional/post-depositional history of the leg is less questionable as compared to the bones, which are from both in situ and beach localities; (4) radiocarbon dates on several bones fall within MIS 3, suggesting that at least some of the collection was redeposited, whereas the preservational quality of the leg indicates it has remained in place; and (5) the age from the leg is more consistent with the palynological data, which indicate deposition under cool, glacial conditions. The herb-dominated assemblages that are associated with the bone horizon, the mammoth remains themselves, and the underlying, undisturbed sediments are characteristic of the last glacial maximum in western Beringia, making a late-glacial age, an interval marked by the widespread establishment of Betula shrub tundra, problematic.
The Berelyokh investigation represents a classic example illustrating the need for a broad range of interdisciplinary studies to unravel complex environmental histories, such as occur in settings where permafrost and fluvial processes interact. It also speaks to the hazards of thermokarst landscapes for animal populations. Although active permafrost disturbance can occur under a wide range of climates, such hazards to these northern populations are likely to increase as the Arctic warms.
ACKNOWLEDGMENTS
This research was supported by the Far Eastern Branch of the Russian Academy of Sciences (project 15-I-2-067) and the Russian Foundation for Basic Research (project 15-05-06420). We thank Y.A. Korzun for assistance in preparing the figures and Mary Edwards for helpful discussions. We also appreciate the useful comments of two unnamed reviewers.
SUPPLEMENTARY MATERIAL
To view supplementary material for this article, please visit https://doi.org/10.1017/qua.2018.3