



Introduction
Foraminifers are ubiquitous elements in marine ecosystems, found from the estuaries to the deep sea, and they represent one of the best-studied groups of microfossils due to their abundance in the fossil record and importance for biostratigraphy and palaeoenvironmental analyses. Many Phanerozoic foraminiferal faunas are recognized worldwide, and much is known about their distribution in the modern oceans. For the Antarctic region, however, the knowledge on living assemblages is still fragmentary (see Majewski Reference Majewski2010, Majda et al. Reference Majda, Majewski, Mamos., Grabowski, Godoi and Pawłowski2018), and only a few pre-Quaternary assemblages are known.
The notorious patchiness of the foraminiferal fossil record of West Antarctica prevents a comprehensive view of its Cenozoic faunas, but a few assemblages are known from the following deposits: Cretaceous–Palaeocene López de Bertodano Formation and Palaeocene Sobral Formation on James Ross Island, Seymour Island and Vega Island (Huber Reference Huber1986, Reference Huber1988); Eocene La Meseta Formation on Seymour Island (Gaździcki & Majewski Reference Gaździcki and Majewski2012; see also Bitner Reference Bitner1996 for brachiopod-incrusting foraminifers and Badaró Reference Badaró2019 for rare agglutinated specimens); Oligocene Polonez Cove Formation on King George Island (Gaździcki Reference Gaździcki1989, Majewski & Gaździcki Reference Majewski and Gaździcki2014); Early Miocene Cape Melville Formation, also on King George Island (Birkenmajer & Łuczkowska Reference Birkenmajer and Łuczkowska1987), Middle Miocene deposits off Weddell Sea (Majewski et al. Reference Majewski, Olempska, Kaim and Anderson2012) and Miocene Hobbs Glacier Formation (HGF) on James Ross Island (Jonkers et al. Reference Jonkers, Lirio, Del Valle and Kelley2002, Concheyro et al. Reference Concheyro, Salani, Adamonis and Lirio2007); Pliocene Cockburn Island Formation on the homonymous island (Hennig Reference Henning1910, Holland Reference Holland1910, Gaździcka & Gaździcki Reference Gaździcka and Gaździcki1994, Gaździcki & Webb Reference Gaździcki and Webb1996); and Pleistocene deposits on Vega Island (Caramés & Concheyro Reference Caramés and Concheyro2013, Concheyro et al. Reference Concheyro, Caramés, Amenábar and Lescano2014).
As for East Antarctica, the record is also scarce. Although a series of ocean drilling projects provided a better understanding of the Cenozoic foraminifers from Ross Sea (see Majda et al. Reference Majda, Majewski, Mamos., Grabowski, Godoi and Pawłowski2018), the knowledge on assemblages from shallower settings - presumably recorded in outcrops on land - is still very fragmentary (see Majewski et al. Reference Majewski, Tatur, Witkowski and Gaździcki2017, Reference Majewski, Bart and McGlannan2018).
In order to fill a gap in and improve the knowledge of the Antarctic fossil foraminifers, here we report a new foraminiferal assemblage for the Miocene HGF and the first possible indigenous foraminifers for the Plio-Pleistocene Weddell Sea Formation (WSF) on Seymour Island, West Antarctica, as well as some associated microfossils. We also discuss their palaeoenvironmental and palaeoecological significance and investigate their taxonomic similarities with other assemblages from both West and East Antarctica.
Materials and methods
Stratigraphic sections were defined and sampled by Antonio C. Rocha Campos (Universidade de São Paulo) and his team during field activities on Seymour Island (Fig. 1) in the summers of 2011 and 2012, during the 29th and 30th Brazilian Antarctic operations. We have adopted the same profile nomenclature and notation used by Rocha Campos' team during their field campaigns and recorded in their field notes.
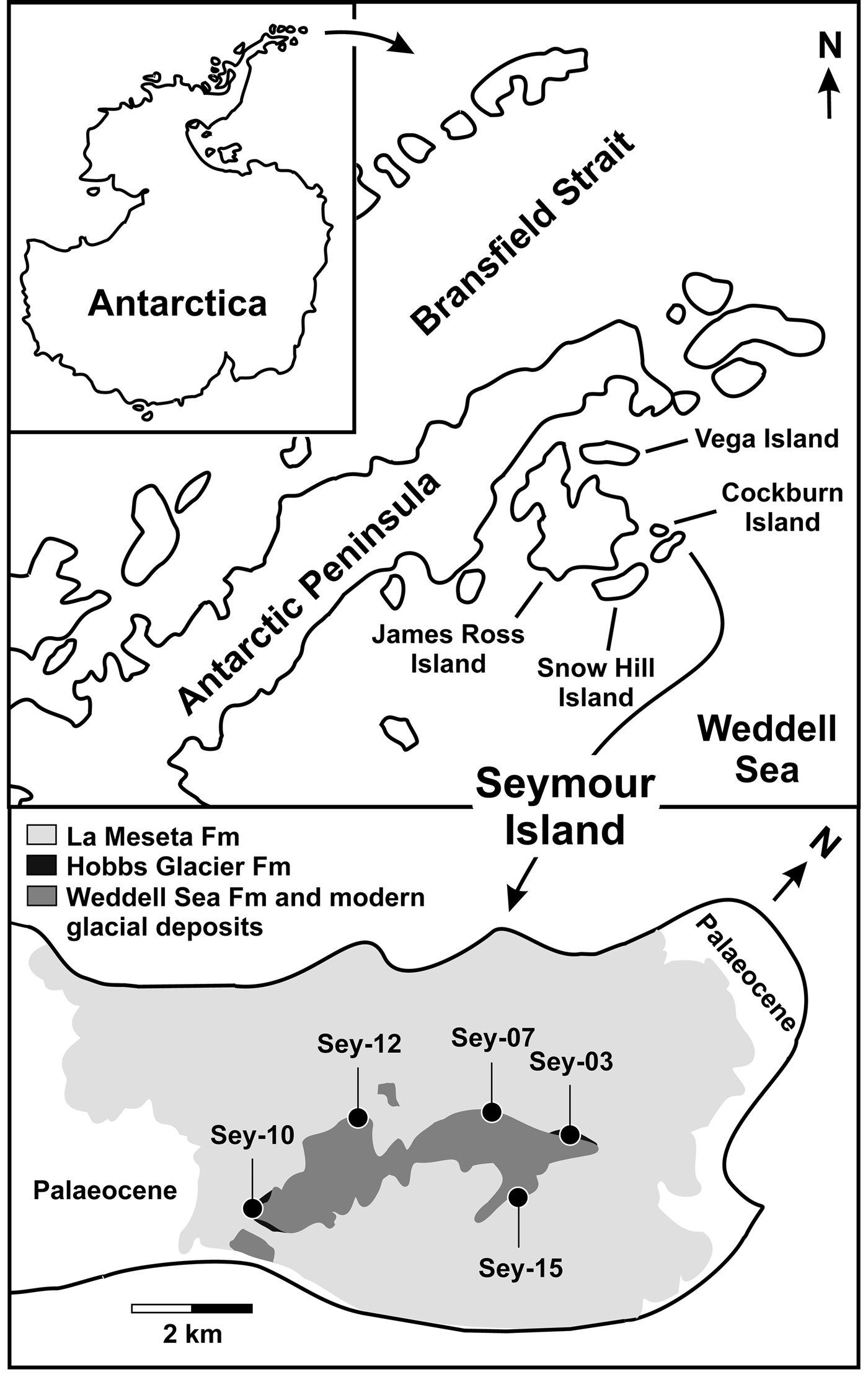
Fig. 1. Geologic map of the north-east portion of Seymour Island, showing the locations of the analysed outcrops. Fm = formation. (Modified from Montes et al. Reference Montes, Nozal, Santillana, Marenssi and Olivero2013, with supplementary information from Marenssi et al. Reference Marenssi, Casadío and Santillana2010 and Rocha Campos et al. Reference Rocha Campos, Kuchenbecker, Duleba, Santos and Canile2017.)
The foraminifers herein reported come from the Sey-03 section (64°14′05.2″S, 56°37′12.7″W), but other important and possibly indigenous microfossils occur in some levels of the Sey-07 (64°14′20.1″S, 56°38′16.5″W), Sey-10 (64°15′26.8″S, 56°39′22.2″W), Sey-12 (64°14′43.4″S, 56°39′27.3″W) and Sey-15 (64°14′32.6″S, 56°37′10.2″W) sections (Fig. 2).
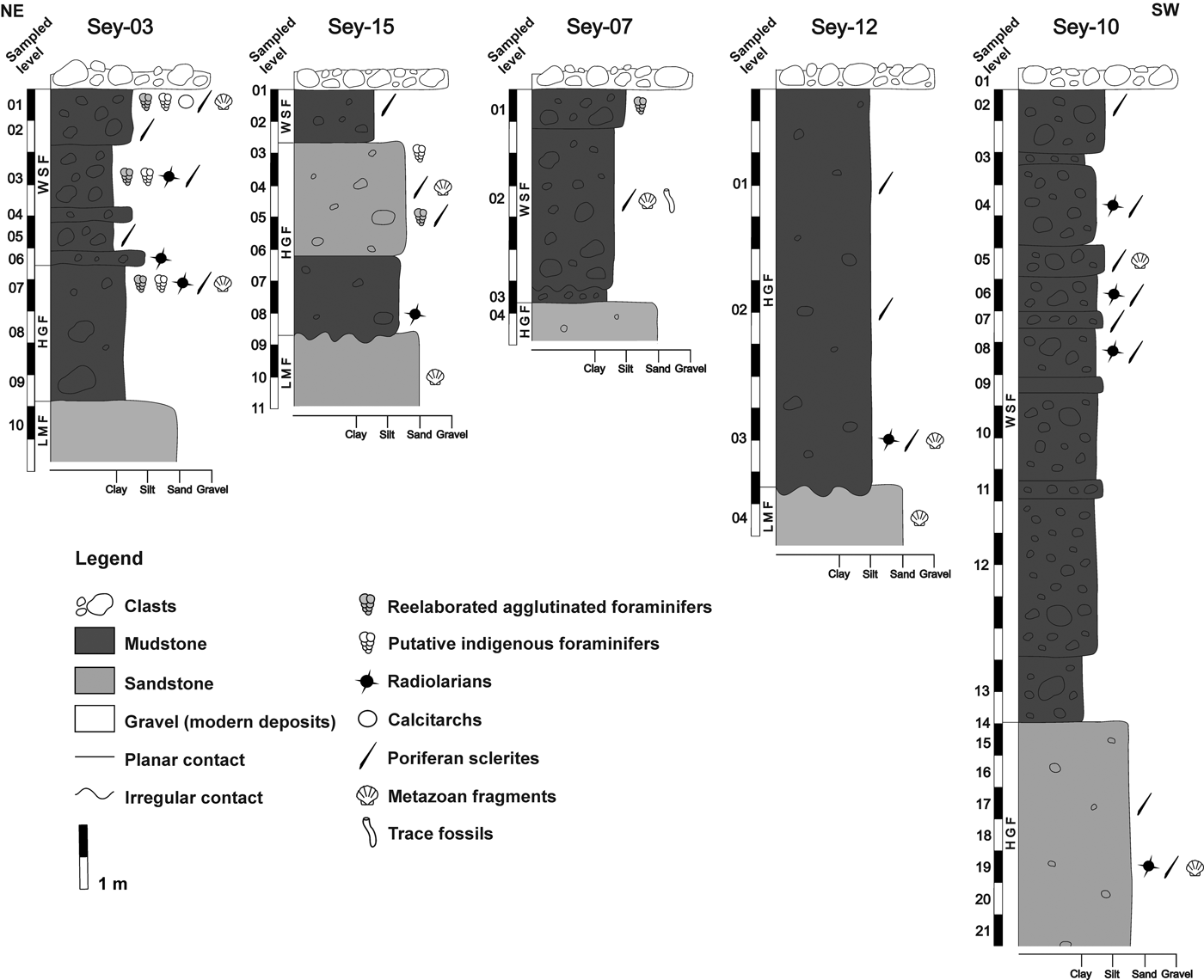
Fig. 2. Stratigraphic sections of the Hobbs Glacier Formation and Weddell Sea Formation on Seymour Island, showing lithologic and palaeontologic data. Sections aligned in a NE–SW transect. LMF = La Meseta Formation; HGF = Hobbs Glacier Formation; WSF = Weddell Sea Formation.
Our recognition of the HGF and WSF is based on their known distributions on Seymour Island and their characteristic lithofacies (Gaździcki et al. Reference Gaździcki, Tatur, Hara and Del Valle2004, Marenssi et al. Reference Marenssi, Casadío and Santillana2010, Montes et al. Reference Montes, Nozal, Santillana, Marenssi and Olivero2013, Rocha Campos et al. Reference Rocha Campos, Kuchenbecker, Duleba, Santos and Canile2017). On Seymour Island, the HGF can be recognized by its two main lithofacies: the lower muddy, matrix-supported diamictite and the upper pebbly sandstone (Marenssi et al. Reference Marenssi, Casadío and Santillana2010), while the WSF also consists of a muddy, matrix-supported diamictite, but it shows a higher density of clasts (Gaździcki et al. Reference Gaździcki, Tatur, Hara and Del Valle2004). The typical muddy diamictite of the HGF is found only in Sey-03 section, being replaced by a silty diamictite in Sey-12 and Sey-15; and as the silty stratum is overlaid by the typical pebbly sandstone in the Sey-15 section, this confirms it represents a minor variation of the same lithofacies.
We analysed c. 500 g of rock for each sampled level. Most of the incipiently lithified matrix was easily disaggregated using only water. The sediment then passed through sieves of 1 cm, 0.5 mm and 0.063 mm. Non-disaggregated, light to moderately consolidated matrix pieces retained in the sieve of 1 cm were reprocessed using the kerosene disintegration method (for details, see Thomas & Murney Reference Thomas and Murney1985).
Sediments retained in the sieves of 1 cm and 0.5 mm were entirely screened, but due to the large amount of disaggregated material from the sieve of 0.063 mm, we used trichloroethylene (a high-density liquid) to concentrate (by floating) the relatively small and unfilled specimens of this portion. Approximately 10 g of the processed waste was screened in order to find any non-floated microfossils, but only relatively large, linear poriferan sclerites were found. The sediment generated through the kerosene method was screened separately and, again, only linear sclerites were found.
The foraminifers and associated microfossils are deposited in the micropalaeontological collection of the Laboratório de Paleontologia Sistemática of the Instituto de Geociências, Universidade de São Paulo (catalogue GP/5E, microslides 4307–4365).
Results
Microfossils of the Hobbs Glacier Formation
The most abundant and diverse assemblage of the HGF occurs in the Sey-03 section (Table I). Although usually scarce, microfossils include foraminifers, radiolarians, poriferans and other remains (Fig. 3).

Fig. 3. Foraminifers and associated microfossils from the Hobbs Glacier Formation (HGF) and Weddell Sea Formation (WSF) on Seymour Island. 1. Psammosphaera fusca. WSF. 2. Favulina hexagona. WSF. 3. Oolina globosa. HGF. 4. Globocassidulina cf. subglobosa. WSF. 5. Bolivina sp. HGF. 6. Globigerinita uvula. WSF. 7. Rosalina cf. globularis. HGF. 8. Indeterminate agglutinated foraminifer. HGF, WSF. 9. Larcopyle sp. HGF, WSF. 10. Cenosphaera sp. HGF. 11. Calcitarch. WSF. 12. Demospongian sclerite. HGF, WSF. 13. Hexactinellid sclerite. HGF. 14. Detail of 13. 15. Geodiid sclerite. WSF. 16. Calcareous tube. WSF. 17. Detail of 16, showing large rhombohedral crystals on inner tube surface.
Table I. Microfossils of the Sey-03 section.

HGF = Hobbs Glacier Formation; WSF = Weddell Sea Formation.
The foraminiferal assemblage of the HGF occurs in the diamictite's muddy matrix of the seventh level of the Sey-03 section and it is composed of the benthic Oolina globosa and Rosalina cf. globularis associated with the radiolarian Larcopyle sp. Microfossils from other outcrops represent non-foraminiferal or single foraminiferal specimens: 16 specimens of the radiolarian Cenosphaera sp. were recovered from the 19th level of the Sey-10 section; 7 triaxonic sclerites, representing hexactinellid poriferans, where found in the first level of the Sey-12 section; and a fragmented specimen of the benthic foraminifer Bolivina sp. was found in sample from the third level of the Sey-15 section. Linear sclerites of demospongians occur in some strata, as well as fragmented serpulids, bivalves, gastropods, and echinoderms re-elaborated (i.e. exhumed and displaced after a preceding final burial) from the Eocene La Meseta Formation and large, poorly preserved (i.e. completely filled and partially covered by micrite) agglutinated foraminifers re-elaborated from an unknown deposit.
Specimens of O. globosa are well preserved (i.e. complete tests, with no signs of abrasion or dissolution and unaltered colour and transparency), while those of R. cf. globularis show a moderate to good state of preservation (i.e. complete tests, with only small and surficial signs of abrasion or dissolution). All radiolarians are coated with clay minerals and are partially filled by sediment.
Although present in strata from both formations, Larcopyle sp. are relatively abundant only in the HGF, especially in the 19th level of the Sey-10 section (in which it co-occurs with Cenosphaera sp.) and in the Sey-12 section, from where 21 and 15 specimens were recovered, respectively.
Microfossils of the Weddell Sea Formation
As in the case of the HGF, microfossils occur in different localities of the WSF, but are more abundant and diverse in strata of the Sey-03 section (Fig. 3, Table I).
The two main foraminiferal assemblages occur in the upper portion of the Sey-03 section. The lower assemblage is found in the diamictite's muddy matrix of the third level and it is composed of the benthic G. cf. subglobosa and Larcopyle sp. specimens. The upper assemblage occurs in the diamictite's silty matrix of the first level and it is composed of the benthic Favulina hexagona and Psammosphaera fusca and the planktonic Globigerinita uvula. We have also found rare spherical calcistarchs (probably closed dinoflagellate cysts, but without an operculum suture to confirm this identification) and a demospongian sclerite of the sterraster type (exclusive to the Geodiidae family and with the typical spherical morphology of those from the Geodia genus) in a sample from the first level of the Sey-03 section.
Specimens of F. hexagona, P. fusca and G. uvula are well preserved (i.e. complete specimens, with no signs of abrasion or dissolution and unaltered colour and transparency), while Bolivina sp. specimens are poorly preserved (i.e. fragmented and with opaque, yellowish colour). Specimens of G. cf. subglobosa are considered only to be in a good state of preservation because, although complete and unfilled, their coating of clay minerals prevents observation of other taphonomic features. Larcopyle sp. specimens present the same taphonomic features of those from the HGF. The hexactnellid sclerites vary by having one or two broken rays (Fig. 3.13–14), which could be considered good preservation for such delicate structures. On the other hand, the geodiid sclerite is complete and has all of its countless rays immaculately preserved (Fig. 3.15).
Trace fossils with an unusual mode of preservation occur in the second level of the Sey-07 section. This assemblage is composed of 48 hollow and cylindrical to conical objects of carbonate composition (Fig. 3.16), sometimes closed at one end, and are herein interpreted as moulds of the tunnel walls produced by worm-like animals. Under the electron microscope, the moulds have a smooth external surface and a rhombohedral crystalline internal texture (Fig. 3.17).
Discussion
The taphonomy of the assemblages and their palaeoenvironmental significance
In general, an autochthonous or parautochthonous nature of the small benthic foraminifers (i.e. F. hexagona, G. cf. subglobosa, O. globosa and P. fusca) would be deduced due to their good to excellent state of preservation and lack of sedimentary filling; and although not as well preserved as most benthic forms, the lack of sedimentary filling in the planktonic specimens would suggest that they are contemporaneous to the former. In the same sense, the well-preserved, complex sclerites of geodiids and hexactinellids - common poriferans in the modern Antarctic seas (Uriz Reference Uriz, Hooper and van Soest2002, McClintock et al. Reference McClintock, Amsler, Baker and van Soest2005) - would also be considered contemporaneous with the deposition of the Miocene and Plio-Pleistocene diamictites. All of these well-preserved specimens contrast with the usually poorly preserved (i.e. filled and covered by lithified micrite) agglutinated foraminifers from some strata of both the HGF and WSF, which certainly were re-elaborated from older deposits.
However, studies on the Quaternary glacial–marine deposits of East Antarctica have shown that the recurring advance and retreat of ice sheets was responsible for exhuming specimens from deposits of earlier glacial cycles and for mixing them with indigenous specimens, especially in the grounding-zone wedges (see Bart et al. Reference Bart, Coquereau, Warny and Majewski2016, Majewski et al. Reference Majewski, Bart and McGlannan2018, Prothro et al. Reference Prothro, Simkins, Majewski and Anderson2018). In these mixed assemblages, it can be difficult or even impossible to differentiate between indigenous and re-elaborated elements from a single assemblage, since a good state of preservation (i.e. non-filled and morphologically pristine tests) is not necessarily a sign of indigenous nature (Bart et al. Reference Bart, Coquereau, Warny and Majewski2016, Prothro et al. Reference Prothro, Simkins, Majewski and Anderson2018) - possibly because re-elaborated tests could have been exhumed and then rapidly buried in new sediments, or even been reincorporated still inside sedimentary clasts, which would ensure some protection against physical damage.
In order to try to differentiate such elements, one can consider their state of preservation and their abundance in a given assemblage, with abundant and well-preserved specimens (i.e. unfilled and chemically and physically unaltered tests) being considered the most probable candidates for indigenous components and rare and poorly preserved specimens (i.e. filled and altered tests) considered as probably re-elaborated (Bart et al. Reference Bart, Coquereau, Warny and Majewski2016, Prothro et al. Reference Prothro, Simkins, Majewski and Anderson2018), and also compare stable isotopes data from each particular species (Majewski et al. Reference Majewski, Bart and McGlannan2018). However, these criteria are not applicable to depauperated assemblages such as those herein reported, and the question of their contemporaneity to the host deposits or their re-elaboration, only partially or as a whole, must remain open.
Nevertheless, alternative criteria suggest the possibility that at least some specimens can be in fact indigenous. Oolina globosa occurs associated with an indigenous assemblage in the HGF on James Ross Island (Jonkers et al. Reference Jonkers, Lirio, Del Valle and Kelley2002), indicating such species as an original element for this geological unit. As for the WSF, specimens as fragile as our unfilled P. fusca and the calcareous tubes would hardly resist being reworked, unless they were still inserted into a more or less consolidated matrix that would only be disaggregated during sample processing. But this seems improbable, since these microfossils were found in the readily friable matrix and not in the most consolidated fragments that needed to be disaggregated through the kerosene method, which are the most probable candidates for reworked diamictite fragments from earlier glacial cycles. So, it is probable that these specimens represent not only indigenous, but also autochthonous or parautochthonous remains, in view of both their inherent fragility and excellent preservation and their occurrence in the more friable parts of matrices. In contrast, excluding the large agglutinated foraminifers (whose taphonomic features indicate them as the oldest re-elaborated elements of the assemblages), Bolivina sp. and R. cf. globularis specimens are the more probable candidates to have been re-elaborated from previous glacial strata, in view of their fragmentation and altered colour (Bolivina sp.) and their surficial damage and sedimentary filling (R. cf. globularis).
If one assumes the autochtonous–parautochtonous hypothesis for the small and fragile benthic foraminifers, the paucity of the assemblages can be explained by taphonomic factors. In the first case, agglutinated forms, such as P. fusca, may have been easily obliterated by the degradation of the organic cement of the test or by the compaction of the deposits, while calcareous tests may have been dissolved by acid interstitial waters (Berkeley et al. Reference Berkeley, Perry, Smithers, Horton and Taylor2007). In fact, the absence or scarcity of foraminifers in other West Antarctic units (e.g. Gaździcki & Majeswki Reference Gaździcki and Majewski2012, Caramés & Concheyro Reference Caramés and Concheyro2013, Badaró Reference Badaró2019) may be explained by similar causes. Moreover, the original communities may have been dominated by allogromids - as in some modern Antarctic environments (see Gooday Reference Gooday2002) - which have an extremely low potential for fossilization, if any.
While the glacial–marine palaeoenvironment of the HGF has been well established by other works (e.g. Pirrie et al. Reference Pirrie, Crame, Riding, Butcher and Taylor1997, Concheyro et al. Reference Concheyro, Salani, Adamonis and Lirio2007, Marenssi et al. Reference Marenssi, Casadío and Santillana2010), the depositional setting of the WSF is still debatable. The WSF, formerly also interpreted as glacial–marine (e.g. Malagnino et al. Reference Malagnino, Olivero, Rinaldi and Spikermann1981, Zinsmeister & DeVries Reference Zinsmeister and DeVries1982), was recently reinterpreted as fully glacial by Gaździcki et al. (Reference Gaździcki, Tatur, Hara and Del Valle2004), who hypothesized a moraine depositional setting. However, our putative indigenous - and possibly autochthonous or parautochthonous - specimens may substantiate a different interpretation.
In order to admit the deposition in moraines for the WSF, it would be necessary to interpret all of our specimens as re-elaborated. Nonetheless, although our assemblages could be interpreted as the results of glacial re-elaboration, to harmonize with the glacial depositional setting proposed by Gaździcki et al. (Reference Gaździcki, Tatur, Hara and Del Valle2004), both the inherent fragility and state of preservation of some of our microfossils may indicate that at least some microfossils are indigenous. The case is especially significant due to the occurrence of P. fusca, a species with a very fragile test; and it is even more critical considering our specific P. fusca specimens, which lack any sedimentary filling that would represent a strong diagenetic reinforcement and therefore would not resist significant reworking. In addition, had they been re-elaborated, the most probable source would be the underlying HGF, but there is no correlation between the foraminifers of both units, and they show no taxonomic nor taphonomic similarities with the large, micrite-filled agglutinated foraminifers that certainly are the oldest re-elaborated specimens, nor even with the Bolivina sp. and R. cf. globularis specimens, the most probable candidates to represent subsequent re-elaboration. In the same sense, the presence of the small, tubular trace fossils strongly suggests the marine influence for at least one stratum of the WSF, since even though these fossils consist of moulds of the former tunnel walls, they are still very delicate and would not resist any significant reworking. In fact, they seem to have barely survived the sample treatment only due to the presence of relatively large carbonate crystals, which act as a structural reinforcement of diagenetic origin.
Finally, the well-preserved Bryozoan colony reported by Gaździcki et al. (Reference Gaździcki, Tatur, Hara and Del Valle2004), if reinterpreted in light of the alternative autochnous–parautochnous hypothesis, should represent not a reworked element, but the colonization of pebbles that reached the shores of the Weddell Sea sometime between the latest Pliocene and the early Pleistocene, when the sea level was still relatively high, and would also be considered as evidence for a marine influence on the deposition of the WSF.
Taxonomic similarities with other Antarctic assemblages
The sparse and poorly diverse foraminiferal assemblages of the HGF and WSF could not be correlated with other fossil or modern assemblages of the Antarctic Peninsula, including the assemblages previously reported for the HGF (Jonkers et al. Reference Jonkers, Lirio, Del Valle and Kelley2002, Concheyro et al. Reference Concheyro, Salani, Adamonis and Lirio2007). However, some taxa were also found in Cenozoic deposits of both West and East Antarctica.
As for the HGF, single specimens of O. globosa were reported for the Recent deposits of King George Island (Majewski Reference Majewski2005), the Pleistocene deposits of Vega Island (Caramés & Concheyro Reference Caramés and Concheyro2013) and the Pliocene Cockburn Island Formation in the homonymous island (Gaździcki & Webb Reference Gaździcki and Webb1996), as well as the already mentioned occurrence in the HGF on James Ross Island (Jonkers et al. Reference Jonkers, Lirio, Del Valle and Kelley2002). In turn, R. cf. globularis occurs in the Recent deposits of King George Island (Majewski Reference Majewski2005), and in East Antarctica, it occurs in deposits as old as the Oligocene (Leckie & Webb Reference Leckie and Webb1986, Webb & Strong Reference Webb and Strong1998, Strong & Webb Reference Strong and Webb2000), in the Miocene Fisher Bench Formation (Majewski et al. Reference Majewski, Tatur, Witkowski and Gaździcki2017), in Pliocene deposits of the Ross Sea (Webb & Strong Reference Gaździcki, Tatur, Hara and Del Valle2000) and in the Sørsdal Formation of Vestfold Hills (Quilty Reference Quilty2010). Thus, our specimens represent the oldest occurrence known so far for West Antarctica.
As for the WSF taxa, comparisons can also be made with the assemblages reported by Caramés and Concheyro (Reference Caramés and Concheyro2013). These authors reported specimens of Globocassidulina sp. that are superficially similar to our clay-covered G. cf. globosa specimens. In addition, G. cf. subglobosa occur in the La Meseta Formation (Gaździcki & Majewski Reference Gaździcki and Majewski2012), in the Oligocene Polonez Cove Formation (Majewski & Gaździcki Reference Majewski and Gaździcki2014), in the Early Miocene Cape Melville Formation (Birkenmajer & Łuczkowska Reference Birkenmajer and Łuczkowska1987) and in the Cockburn Island Formation (Gaździcki & Webb Reference Gaździcki and Webb1996). As for P. fusca, this species occurs in the Cretaceous–Palaeocene López de Bertodano Formation (Huber Reference Huber1988) and in the Cape Melville Formation (Birkenmajer & Łuczkowska Reference Birkenmajer and Łuczkowska1987), as well as in the Recent slope and abyssal plain of the Weddell Sea (Cornelius & Gooday Reference Cornelius and Gooday2004) and in inlets of King George Island, where it is found at depths of 20–250 m (Majewski Reference Majewski2005, Reference Majewski2010). Favulina hexagona was also reported (as Oolina hexagona) for the Cape Melville Formation (Birkenmajer & Łuczkowska Reference Birkenmajer and Łuczkowska1987), but there is no other record of this species for the Cenozoic deposits of the Antarctic Peninsula, although other Favulina sp. are known, such as Favulina epibathra (reported as Conolagena epibathra) from the Pleistocene deposits of Vega Island (Caramés & Concheyro Reference Caramés and Concheyro2013) and a Favulina sp. (reported as Lagena sp. 1) with a reticulated pattern composed of both pentagons and hexagons (instead of the strictly hexagonal pattern of F. hexagona) from the HGF (Jonkers et al. Reference Jonkers, Lirio, Del Valle and Kelley2002). As for the planktonic G. uvula, the species does not occur in other Antarctic deposits, but it was reported (as Tenuitellinata uvula) for the Neogene and Pleistocene deposits of the sub-Antarctic portion of the Kerguelen Plateau (Li et al. Reference Li, Radford and Banner1992). None of our G. uvula nor the Kerguelen Plateau specimens presents bula, and specimens from both localities have similar sizes and small tubers on the umbilical sides of the tests, suggesting they were part of a same population.
Regarding non-foraminiferal microfossils, our smooth calcitarchs do not resemble the crystalline calcisphaerid dinolagellates from the Cretaceous–Palaeogene deposits of the Weddell Sea (cf. Fütterer Reference Fütterer1990); instead, they are similar to the smooth thoracosphaerids from the Quaternary of the Ross Sea (Villa & Wise Reference Villa and Wise1998). As for the spumellarian radiolarians, they are common in the Neogene of the Weddell Sea, especially the prunoid morphotypes (i.e. with an external ellipsoidal shell and an internal spongy structure), such as some Larcopyle sp. (Lazarus Reference Lazarus1990, Lazarus et al. Reference Lazarus, Faust and Popova-Goll2005). Despite their overall good preservation and absence of sedimentary filling, our Larcopyle sp. specimens are common only in some levels of the HGF and are especially rare in the WSF, which suggests that the specimens found in the latter were exhumed from the Miocene strata. This interpretation is indeed consistent with the biostratigraphy of the Weddell Sea spumellarians, since the majority of Larcopyle spp. and subspp. (specially some Larcopyle polyacantha subspecies that resemble our specimens) have a chronostratigraphic distribution restricted from the Miocene to the Early Pliocene (Lazarus et al. Reference Lazarus, Faust and Popova-Goll2005).
Conclusion
The question of the indigenous or re-elaborated origin of the foraminiferal assemblages remains open, as their low abundance and diversity, associated with the complex taphonomical processes that prevailed in glacial–marine palaeoenvironments, do not allow further inferences. Yet, the fragility and state of preservation of P. fusca specimens and of the moulds of trace fossil walls strongly suggest their indigeneity for the WSF, while the presence of well-preserved specimens of O. globosa, previously known from an indigenous assemblage of the HGF on James Ross Island, also suggests its indigeneity to the HGF on Seymour Island.
In either case, the assemblages herein reported fill a gap in the knowledge on the Antarctic fossil foraminifers, since just two foraminiferal assemblages of the HGF were previously known and, along with the Destruction Bay Formation of King George Island, the WSF was the only known Cenozoic unit of West Antarctica for which foraminifers were unknown.
It is somewhat remarkable that, except for P. fusca, the other benthic species do not occur in modern Antarctic communities, and that, with the exception of G. cf. globosa, they are rare in the few other geological units in which they occur. However, if there is any palaeobiological significance in these aspects, it is hidden by the complex taphonomy of the assemblages.
Acknowledgements
We thank Antonio C. Rocha Campos and Wania Duleba for making available to us the samples from Seymour Island. We also thank both anonymous reviewers for their thorough commentaries on an earlier draft of this paper. This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (grant no. 5570362009-7) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (grant no. 1267397).
Author contributions
Victor C.S. Badaró prepared and analysed the samples, identified the microfossils, interpreted the data and wrote and revised the manuscript. Setembrino Petri identified the microfossils, interpreted the data and revised the manuscript.




