



Introduction
The temnospondyl Sclerocephalus haeuseri Goldfuss, Reference Goldfuss1847 was among the first Paleozoic tetrapod genera to be discovered (Goldfuss, Reference Goldfuss1847; von Meyer, Reference Meyer1858b), and because of its large size and abundance in many localities, it has gained much attention (Branco, Reference Branco1887; Ammon, Reference Ammon1889; Broili, Reference Broili1926; Boy, Reference Boy1988; Werneburg, Reference Werneburg1992; Meckert, Reference Meckert1993; Schoch, Reference Schoch2009; Schoch and Witzmann, Reference Schoch and Witzmann2009a). Species of Sclerocephalus were 1.5–2 m long aquatic predators known from lake deposits in southwestern Germany (Saar-Nahe Basin) and central Germany (Thuringian Forest Basin), confined to the lower Rotliegend (Autunian, Gzhelian–Asselian) of the late Paleozoic rock sequence (Boy et al., Reference Boy, Haneke, Kowalczyk, Lorenz, Schindler, Stollhofen, Thum, Lützner and Kowalczyk2012). Because of its best-known species, S. haeuseri, this taxon also has an exceptionally well-known ontogeny, documented by numerous larval and juvenile stages (Boy, Reference Boy1972; Lohmann and Sachs, Reference Lohmann and Sachs2001; Schoch, Reference Schoch2003; Werneburg, Reference Werneburg2017). Quite like Onchiodon in the eastern German basins, Actinodon frossardi Gaudry, Reference Gaudry1866 in France, and Eryops megacephalus Cope, Reference Cope1878 in North America, Sclerocephalus haeuseri was a heavily ossified taxon with a massive skull, pectoral girdle, and full complement of dermal scales. However, a puzzling feature of all hitherto known species of Sclerocephalus, consistent with other temnospondyls known from the Saar-Nahe Basin, is the very slow ossification of endochondral bones in the skeleton, as documented in the vertebrae, carpals, and tarsals, and especially the much-delayed ossification of the braincase and palatoquadrate (Boy, Reference Boy1988; Schoch and Witzmann, Reference Schoch and Witzmann2009a). So far, only small portions of the braincase (namely, the central sphenethmoid) have been known to become ossified in large adults of Sclerocephalus spp., whereas the remaining portions of the endocranium remained cartilaginous (Schoch and Witzmann, Reference Schoch and Witzmann2009a), consistent with the condition in close relatives such as Glanochthon latirostris (Jordan, Reference Jordan1849) and Archegosaurus decheni Goldfuss, Reference Goldfuss1847 (Witzmann, Reference Witzmann2006a, Reference Witzmannb).
A single newly identified specimen similar to Sclerocephalus haeuseri from a stratigraphically more ancient lake deposit in St. Wendel in the Saar region, southwestern Germany (Fig. 1) has a completely ossified endocranium, with fully established braincase, otic capsules, and epipterygoid regions. The specimen is recognized as representing a new species because of a suite of unique characters in the palate. Moreover, this find permits, for the first time, examination of the complete endocranial anatomy of a basal taxon in the stereospondylomorph clade, prompting an analysis of braincase evolution in temnospondyls.
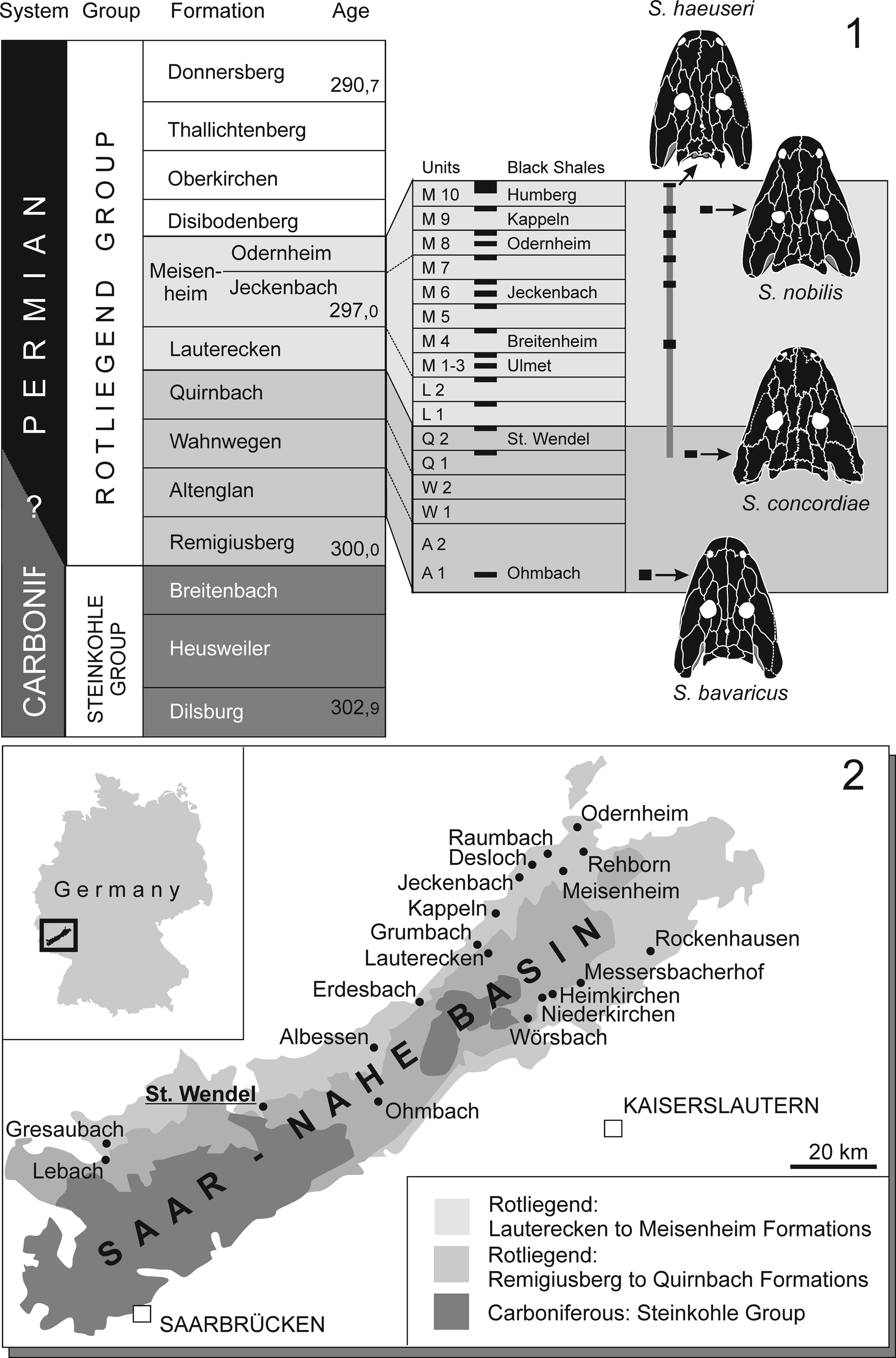
Figure 1. (1) Stratigraphic position of S. concordiae n. sp. and range of Sclerocephalus. (2) Geology of Saar-Nahe Basin with most important Rotliegend Lagerstätten mapped.
Objectives
This study aims to: (1) name and describe the new species with particular emphasis on the endocranium, (2) elucidate its phylogenetic position, and (3) trace the character evolution in the braincase and palatoquadrate across the Temnospondyli.
Materials and methods
Following Boy (Reference Boy1987), the type locality of the new taxon was a clay pit (brickyard “Concordia”). In the late nineteenth and early twentieth centuries, dark mudstones were quarried there for the production of tiles. The mudstones intercalated with characteristic reddish-gray carbonate layers that produced specimens of the actinopterygian Paramblypterus sp., few small branchiosaurid skeletons (Apateon pedestris von Meyer, Reference Meyer1844), and rare but well-preserved specimens of Sclerocephalus spp., referred by Boy (Reference Boy1987, fig. 2) to S. haeuseri. The hitherto largest specimen from that locality was described in detail by Broili (Reference Broili1926) and later figured by Schoch and Milner (Reference Schoch and Milner2000, plate 1b). The horizon was reported by Boy (Reference Boy1987) and Boy et al. (Reference Boy, Haneke, Kowalczyk, Lorenz, Schindler, Stollhofen, Thum, Lützner and Kowalczyk2012) as being in the lower part (Q1) of the Quirnbach Formation (Autunian, lower Rotliegend). Stratigraphically, this horizon forms the second-oldest occurrence of the genus Sclerocephalus, with the oldest record (S. bavaricus [Branco, Reference Branco1887]) stemming from the somewhat older Altenglan Formation of Ohmbach (Boy, Reference Boy1987; Schoch and Witzmann, Reference Schoch and Witzmann2009a).
Materials examined
BSP AS I 575 (skull in palatal view, 12 cm skull length, with pectoral girdle and forelimbs), described and figured by Broili (Reference Broili1926) and Schoch and Milner (Reference Schoch and Milner2000, plate 1b), GPIT Am 699 (skull roof and partial palate in ventral view, 11.5 cm skull length). GZG.V 7998 (Fig. 2) comprises a nearly complete skull and anterior trunk and forms the largest specimen from the locality St. Wendel (Saar region, Germany). It is housed at the collection of the Geological Institute of Göttingen University, Germany, where it was identified by the author as an unprepared temnospondyl in a concretion. Preparation at the Staatliches Museum für Naturkunde Stuttgart gave a beautiful specimen that is unique among all known Sclerocephalus specimens in its extent of ossification throughout the preserved skeleton.

Figure 2. Type specimen of Sclerocephalus concordiae n. sp., GZG.V 7998. (1) Photograph of skull roof and cervical vertebrae in dorsal view. (2) Photograph of palate and dermal pectoral girdle in ventral view. (3) Line drawing of (1). (4) Line drawing of (2). bb = basibranchial; cl = clavicle; e = ectopterygoid; ep = epipterygoid; icl = interclavicle; f = frontal; ju = jugal; la = lacrimal; m = maxilla; n = nasal; nar = naris; o = palatal ossicle; or = orbit; p = parietal; pf = postfrontal; pl = palatine; pm = premaxilla; po = postorbital; pp = postparietal; prf = prefrontal; ps = parasphenoid; pt = pterygoid; q = quadrate; qj = quadratojugal; se = sphenethmoid; sm = septomaxilla; sq = squamosal; st = supratemporal; sta = stapes; ta = tabular; vo = vomer.
Repositories and institutional abbreviations
BSP, Bayerische Staatssammlung für Geologie und Paläontologie, Munich, Germany; GPIT, Geologisch-Paläontologisches Institut, Tübingen, Germany; GZG, Geologisches Zentrum Universität Göttingen, Germany.
Systematic paleontology
Temnospondyli Zittel, Reference Zittel1888
Rhachitomi Watson, Reference Watson1919, sensu Schoch, Reference Schoch2013
Eryopiformes Schoch, Reference Schoch2013
Stereospondylomorpha Yates and Warren, Reference Yates and Warren2000
Sclerocephalidae Jaekel, Reference Jaekel1909
Genus Sclerocephalus Goldfuss, Reference Goldfuss1847
Type species
Sclerocephalus haeuseri Goldfuss, Reference Goldfuss1847
Sclerocephalus concordiae new species
Figures 2–4
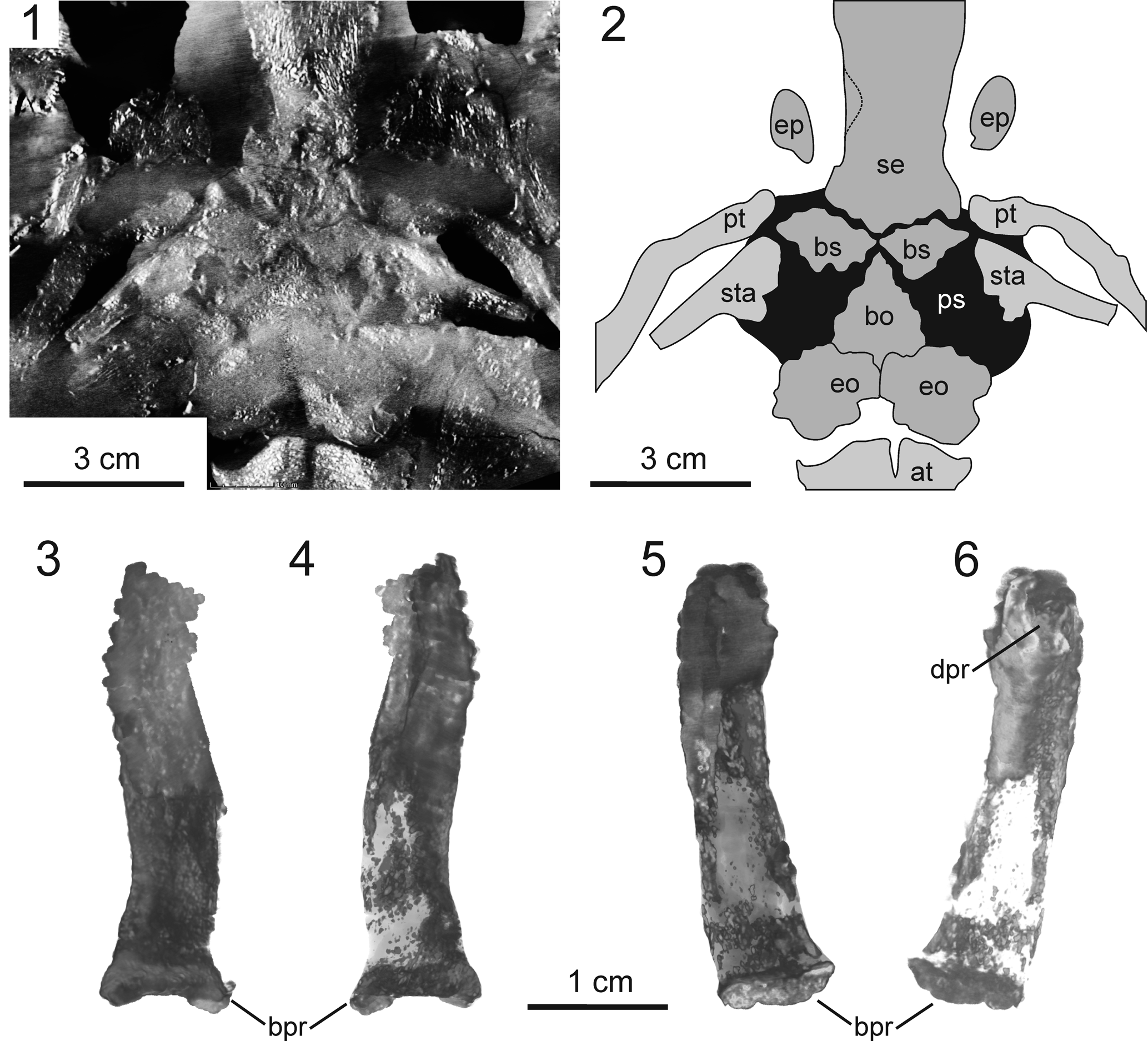
Figure 3. Morphology of endocranium in Sclerocephalus concordiae n. sp., GZG.V 7998. (1) CT image of braincase, occiput, and atlas in dorsal view, horizontally sectioned at mid-level of auditory capsules. (2) Interpretative drawing of (1). (3–6) Epipterygoid: (3) right element in dorsal view; (4) right element in ventral view; (5) left element in ventral view; (6) left element in dorsal view. at = atlas; bo = basioccipital; bpr = basal process (epipterygoid); bs = basisphenoid; dpr = dorsal process; eo = exoccipital; ep = epipterygoid; ps = parasphenoid; pt = pterygoid; se = sphenethmoid; sta = stapes.

Figure 4. Cranial reconstruction of Sclerocephalus concordiae n. sp. (1) S. haeuseri, skull in dorsal view. (2) S. haeuseri, skull in ventral view. (3) S. nobilis, skull in dorsal view. (4–8) S. concordiae: (4) skull roof; (5) palate; (6) skull in lateral view; (7) same as (6) with cheek removed to expose endocranium; (8) endocranium in dorsal view. Braincase elements in black, palatoquadrate in dark gray. (1–3) from Schoch and Witzmann (Reference Schoch and Witzmann2009b). ap = anterior palatal depression; bo = basioccipital; bs = basisphenoid; ch = choana; e = ectopterygoid; eo = exoccipital; ep = epipterygoid; f = frontal; ipv = interpterygoid vacuity; ju = jugal; la = lacrimal; m = maxilla; n = nasal; ot = otic; p = parietal; pf = postfrontal; pl = palatine; pm = premaxilla; po = postorbital; pp = postparietal; prf = prefrontal; ps = parasphenoid; pt = pterygoid; q = quadrate; qj = quadratojugal; se = sphenethmoid; sq = squamosal; st = supratemporal; sta = stapes; stf = subtemporal fenestra; ta = tabular; vo = vomer.
Holotype
Nearly complete skull (18.5 cm skull length), anterior axial skeleton, pectoral girdle, left humerus, and proximal head of ulna (GZG.V 7998, Fig. 2).
Diagnosis
Large-sized species with: (1) completely ossified endocranium in adults (central portion of sphenethmoid already ossified at 12 cm skull length); (2) basipterygoid articulation fully formed; (3) epipterygoid forming an anterodorsally elongate rod; (4) cultriform process and interpterygoid vacuities short, the latter measuring only 40% the length of the skull; (5) palatine and ectopterygoid teeth enlarged; (6) elongated vomer reaching near posterior end of palatine; (7) palatal branch of pterygoid short and wide; and (8) stapes suturally attached to the parasphenoid.
Occurrence
Quirnbach Formation (Q1, lower part), locally developed reddish-brown limestone horizon in mudstone sequence (Boy, Reference Boy1987), Gzhelian–Asselian boundary, (upper Carboniferous–lower Permian interval); Concordia pit, Sankt Wendel, Saarland, southwest Germany (Boy et al., Reference Boy, Haneke, Kowalczyk, Lorenz, Schindler, Stollhofen, Thum, Lützner and Kowalczyk2012).
Description
The skull of Sclerocephalus concordiae shares the straight rather than rounded, tapering margins with those of large specimens of S. haeuseri (Fig. 3), especially from the Jeckenbach lake horizon (M6), lower Meisenheim Formation (Boy, Reference Boy1988; Schoch and Witzmann, Reference Schoch and Witzmann2009a). It also shares all autapomorphies of the genus, notably: (1) the slender supratemporal, (2) small and narrow lacrimal, (3) preorbital region less than twice the length of the postorbital skull table, (4) palatine and ectopterygoid with continuous row of teeth, and (5) interclavicle wide and posteriorly triangular (Schoch and Witzmann, Reference Schoch and Witzmann2009a). An additional derived feature shared by Sclerocephalus spp. and Glanochthon spp. (Schoch and Witzmann, Reference Schoch and Witzmann2009b), the posteromedial process of the vomer, is well developed in S. concordiae.
Whereas the proportions of most elements in the skull roof fall within the individual range of morphologies known from Jeckenbach S. haeuseri (except for the supratemporal being shorter and wider), the palate and endocranium are quite distinct from those in other species of Sclerocephalus. The major difference and unique feature of the holotype of S. concordiae is the complete ossification of endocranial elements, which include the braincase, epipterygoid, and quadrate as well as the basipterygoid and occipital regions. In these features, S. concordiae resembles large specimens of Edops craigi (Romer and Witter, Reference Romer and Witter1942), Eryops megacephalus (Sawin, Reference Sawin1941), Mastodonsaurus giganteus Jaeger, Reference Jaeger1824 (Schoch, Reference Schoch2002a), and Gerrothorax pulcherrimus Fraas, Reference Fraas1913 (Witzmann et al., Reference Witzmann, Schoch, Hilger and Kardjilov2012). Further differences between S. concordiae and S. haeuseri include the size and proportion of the interpterygoid vacuity, the sutural attachment of the stapes to the parasphenoid, and the fully ossified basipterygoid articulation.
Skull roof
The dermal ornament consists of tall and thickened polygonal ridges, most similar in size and distribution to those of the neotype of S. haeuseri from the M 9 horizon of Pfarrwald at Heimkirchen (Boy, Reference Boy1988; Schoch and Witzmann, Reference Schoch and Witzmann2009a, figs. 3C, 5C; Schoch et al., Reference Schoch, Ebert and Robert2019). The dermal bones are unusually thick compared with those of similar-sized or even larger specimens from other localities. There are no traces of lateral line sulci; these are generally poorly developed in the genus Sclerocephalus and were absent in most specimens of S. haeuseri (Boy, Reference Boy1988), as well as in large adults of S. nobilis (Krätschmer and Resch, Reference Krätschmer and Resch2005) (Schoch and Witzmann, Reference Schoch and Witzmann2009a). A pervasive feature of GZG.V 7998 is the serration of sutures, most obvious in the medial series and cheek. The midline suture is markedly stepped where the nasals and frontals meet, a typical feature of Sclerocephalus.
The premaxilla is relatively short, and the region anterior to the naris is not expanded like in S. bavaricus or other sterospondylomorphs. The lacrimal has pointed ends and is smaller and slenderer than in most specimens of S. haeuseri. The orbits are similar in size to those of S. haeuseri and S. bavaricus, substantially larger than in S. jogischneideri Werneburg, Reference Werneburg1992. The interorbital width is greater than in most specimens of S. haeuseri (0.21) but still within the upper range of that taxon, consistent with larger specimens specifically from the Jeckenbach (M6) and Kappeln (M9) horizons (Schoch and Witzmann, Reference Schoch and Witzmann2009a, fig. 2). Additional features present only in large adult S. haeuseri are the very wide jugal, the laterally extended orbits, the lateral excursion of the maxilla to accommodate a larger tooth, and the large, well-ossified septomaxilla.
The supratemporal is somewhat shorter and wider than in S. haeuseri and S. nobilis but much longer than in S. bavaricus and S. jogischneideri. The tabular is massive and has a ventrally expanded descending lamella most similar to that of rhinesuchids (Marsicano et al., Reference Marsicano, Latimer, Rubidge and Smith2017). Postparietal and tabular form a much shallower occipital margin than in S. haeuseri, where it is deeply concave (Schoch and Witzmann, Reference Schoch and Witzmann2009a). Likewise, the squamosal embayment is more consistent with that of S. bavaricus and stratigraphically older (L1–M8) S. haeuseri than the classical samples from Heimkirchen, which were referred to the chronosubspecies S. haeuseri haeuseri (Boy, Reference Boy1988).
Palate
In the palate, the differences between GZG.V 7998 and adult specimens of S. haeuseri and S. nobilis are more profound. Most conspicuous is the smaller size of the interpterygoid vacuities, which in S. concordiae are of equal length as vomer and premaxilla combined. A decrease in the size of the interpterygoid vacuities has been reported as an ontogenetic trend in various temnospondyls, notably eryopids (Boy, Reference Boy1990), and in this feature S. concordiae is more consistent with Onchiodon than with other species of Sclerocephalus. The choana is largely obscured by the lower jaw, but in GPIT Am 669 it is exposed and somewhat more broadly oval than in S. haeuseri.
The palatine and ectopterygoid bear a continuous series of large teeth, most of which have transversely broadened bases much like in stereospondyls (Schoch and Milner, Reference Schoch and Milner2000). Their exact numbers are difficult to determine because sutures are not preserved and some regions are obscured by the mandible. The most complete series of ectopterygoid and posterior palatine teeth is on the right side and consists of a continuous row of 10 teeth (Fig. 2.4). Like in other specimens, the palate was covered by dentigerous plates, some of which are preserved in a disarticulated state in the anterior part of the palate (Figs. 2, 4).
Almost the entire palate is covered by small denticles, including the vomer as well as the quadrate ramus and transverse process of the pterygoid. Only the posterior half of the basal plate (parasphenoid) and the thin cultriform process remain smooth. Medial to the choana, denticles are emplaced on a marked ridge; these are not larger than denticles in other vomerine regions. The anterior region of the basal plate bears a semilunar dentigerous area, with the apex pointing anteriorly. The plate is more massive than in smaller specimens of S. concordiae and S. haeuseri, and the basipterygoid processes are larger and laterally expand well beyond the lateral margins of the plate. They bear anterolateral facets for the articulation with the pterygoid, which is fully ossified and resembles those of Eryops megacephalus and Actinodon frossardi (Gaudry, Reference Gaudry1866; Sawin, Reference Sawin1941). The basipterygoid region is triangular in outline and of “swollen” appearance, clearly offset from the medial region by a deep groove. Posterolaterally, the basal plate forms a raised margin for muscular attachments, consistent with many stereospondylomorphs (Yates and Warren, Reference Yates and Warren2000). The stapes is firmly attached to the ventrolateral margin of the plate.
Neurocranium
The neurocranium forms a continuous ossification from the basisphenoid region through to the anterior end of the braincase level with the posterior rim of the choanae (Fig. 3). The basisphenoid region is visible only in the microcomputed tomography (micro-CT) images, where a symmetrical, anteriorly concave ossification similar to that of Eryops megacephalus is present (Fig. 3; Sawin, Reference Sawin1941). Ossifications of the ear capsule are difficult to trace in the pyrite-rich posterior portion of the endocranium, but bone is certainly present in the floor region (Fig. 3.1).
In GZG.V 7998, the central part, which formed the tallest portion, is broken into two halves so that the lateral walls face nearly ventrally. They are largely smooth but bear a raised area at about midlevel that separates anterior and posterior regions. The anterior region houses two depressions, a deeper dorsal one with a foramen and a shallower anteroventral one. In life, the lateral walls were probably aligned at an angle between 30° and 40° from the vertical plane, by analogy with Eryops (Sawin, Reference Sawin1941). The anterior sphenethmoid part (ethmoid portion) is substantially lower because the preorbital region was much flatter than the interorbital portion of the skull. In ventral view, the ethmoid portion has a stepped lateral margin, with the posterior third of the portion being substantially wider than the anterior two-thirds. Level with this lateral step, the cultriform process broadens, flooring almost the entire width of the anteriormost ethmoid portion. The posterior part of the ethmoid portion sutures with the frontal by means of paired dorsolateral lateral wings. The anteriormost part of the ethmoid portion bears paired ventral descending lamellae that frame the lateral margins of the cultriform process.
Palatoquadrate
The most remarkable element in the skull is the epipterygoid, which is unusually large and well ossified even compared with many Triassic stereospondyls (Schoch and Milner, Reference Schoch and Milner2000; Witzmann et al., Reference Witzmann, Schoch, Hilger and Kardjilov2012). On both sides, it is detached from its base at the anterior margin of the basipterygoid process (pterygoid) and aligned anteriorly, in parallel with but dorsal to the lateral wall of the sphenethmoid, reaching well into the interorbital region. This orientation is probably a result of crushing, consistent with the laterally rotated portions of the braincase (Fig. 2). On the left side, the margin of the facet for the articulation with the pterygoid is exposed (Fig. 3). As revealed by micro-CT data, the bone has a massive, fully ossified ventral portion but forms a slender blade dorsally. The whole element has a gently S-shaped long axis. In life, its long axis was probably aligned anterodorsally. Close to the dorsal end, the posterodorsal surface bears a small buttress (posterodorsal process). This process probably extended close to the skull roof in the region where supratemporal, postfrontal, and parietal meet. In contrast to Eryops megacephalus and Edops craigi, the epipterygoid lacks an expanded, ossified footplate and does not continue posteriorly along the pterygoid.
Stapes
The ear ossicle of immature and medium-sized specimens of S. haeuseri was described by Boy (Reference Boy1988) and Schoch (Reference Schoch2002c). Although consistent in the two-headed proximal region and the elongated shaft, it differs from that of more advanced stereospondylomorphs in having a substantial quadrate process. This is also found in Archegosaurus decheni (Witzmann, Reference Witzmann2006a), but not in Eryops megacephalus (Sawin, Reference Sawin1941; personal observation). The stapes of S. concordiae was consistent with the primitive temnospondyl condition in retaining a large quadrate process (Schoch, Reference Schoch2019). However, in contrast to all other known specimens of Sclerocephalus spp., both stapes of the holotype of S. concordiae are firmly attached to the parasphenoid, which is well exposed on both sides, and the tight suturing is also revealed by micro-CT data. An articulation of the stapes with the parasphenoid is reported for various stereospondyls (Schoch and Milner, Reference Schoch and Milner2000; Arbez et al., Reference Arbez, Dahoumane and Steyer2017). Sutured or fused stapes (to the parasphenoid) have been reported in Lyrocephalus euri Wiman, Reference Wiman1913 (Säve-Söderbergh, Reference Säve-Söderbergh1936), and arguably Eryops megacephalus (Sawin, Reference Sawin1941) and Tersomius texensis Case, Reference Case1910 (Carroll, Reference Carroll1964). In Mastodonsaurus giganteus a tight suturing occurs in large adults (Schoch, Reference Schoch2002c), whereas in Stanocephalosaurus amenasensis Dahoumane et al., Reference Dahoumane, Nedjari, Ait-Quari, Taquet, Vacant and Steyer2016 it occurs only on one side (Arbez et al., Reference Arbez, Dahoumane and Steyer2017).
Hyobranchium
The basibranchial is partially preserved, lying on the ventral surface of the basal plate (Fig. 2). The slender central and posterior portions are exposed, whereas the anterior end is obscured by the cultriform process. Micro-CT data reveal that the anterior end is broadened, as known from other species of Sclerocephalus (Boy, Reference Boy1988; Schoch and Witzmann, Reference Schoch and Witzmann2009a). Other hyobranchial elements, and branchial denticles, are absent.
Postcranial axial skeleton
The first nine vertebrae are preserved, with the associated ribs slightly dislocated but well preserved. The intercentra are wedge shaped and massive, with well-established parapophyses. As described by Boy (Reference Boy1988), the atlas has a low and slender neural arch, the axis has a tall and anteriorly inclined one, and the fourth vertebra is markedly lower than vertebrae 3 and 5. The ribs are bicapitate, with the atlantal rib being the shortest and most slender, whereas the following ribs have much broadened heads and uncinate processes.
Pectoral girdle
The extensive clavicles and interclavicle are largely preserved, but the posterior halves are present only as an impression in the matrix. These dermal bones are proportionally much thicker than in smaller specimens. Whereas the interclavicle is of similar size to those in juveniles of S. concordiae and S. haeuseri, the clavicle is proportionally larger than in smaller stages, with the dermal pectoral girdle as wide as the posterior portion of the skull. The latter feature is also found in the large neotype of S. haeuseri from the Heimkirchen locality (level M9, see Schoch and Witzmann, Reference Schoch and Witzmann2009a). The most characteristic feature of the pectoral girdle in S. concordiae is the ornament, consisting of high and much-thickened ridges, a notable difference from the tall but thin ridges of S. haeuseri (neotype) and the rather pustular ridges of S. nobilis. The cleithrum forms a massive, gently curved element almost exactly the length of the humerus, with a slightly broadened dorsal portion, which differs from that of Eryops megacephalus in the absence of a dorsoposterior process that framed the scapula (Miner, Reference Miner1925).
Humerus
The humerus is slightly more elongate than that of S. nobilis but similar in its heavy appearance. The shaft is longer and slenderer than in other species of Sclerocephalus, and the radial condyle is fully ossified, forming a prominent portion. A substantial supinator process is present and somewhat more clearly offset from the shaft than in S. nobilis. On the proximal head, the deltopectoral crest is well developed.
Etymology
The species name concordiae refers to the type locality, the Concordia clay pit at Sankt Wendel.
Remarks
The high degree of ossification in the holotype is probably an adult feature, but in S. haeuseri, adults of equal or even larger size are known from several horizons (M6, M9, M10) and lack a fully ossified neurocranium as well as a fully ossified epipterygoid. In addition, in the well-known ossification sequence of S. haeuseri, the central portion of the sphenethmoid formed long before the tarsals, carpals, and pubis started to ossify (Schoch and Witzmann, Reference Schoch and Witzmann2009a, fig. 2A), and thus the condition in S. concordiae cannot be considered simply an extension of the ontogenetic trajectory of S. haeuseri.
Results
With Sclerocephalus being one of the key taxa among eryopiform temnospondyls, the present findings add to our understanding of temnospondyl endocrania. To place S. concordiae in a phylogeny of eryopiform temnospondyls, a cladistic analysis was performed, employing the data matrix of Schoch and Witzmann (Reference Schoch and Witzmann2009b).
Data matrix
The original data matrix contained 54 characters (Schoch and Witzmann (Reference Schoch and Witzmann2009b) to which the following three characters were added: (55) interpterygoid vacuities longer than vomer and premaxilla (0), or equal to or shorter (1); (56) neurocranium cartilaginous or only partially ossified (0), or fully ossified with sphenoid and ethmoid portions (1); (57) squamosal embayment framed by parallel squamosal and tabular margins (0), or forming medially rounded extension, constricting the posterior skull table (1). Four taxa were added, which all belong to the genus Sclerocephalus: S. concordiae, S. bavaricus (Boy, Reference Boy1988), S. jogischneideri (Werneburg, Reference Werneburg1992), and S. nobilis (Schoch and Witzmann, Reference Schoch and Witzmann2009a).
Analysis
The analysis of 57 characters and 22 taxa found a single most parsimonious tree requiring 102 steps (CI = 0.59, RI = 0.82). The analysis was conducted in the ACCTRAN mode, under the Traditional Search option. In a variant analysis, the probably juvenile specimen of Sclerocephalus stambergi (Klembara and Steyer, Reference Klembara and Steyer2012) was added to the matrix, which resulted in three most parsimonious trees and a lower resolution (see the following).
Results
In the obtained topology (Fig. 5), the same general branching pattern was retained, except for Melosaurus uralensis von Meyer, Reference Meyer1857 and Intasuchus silvicola Konzhukova, Reference Konzhukova1956, which form sister taxa here but were found to be a grade toward Stereospondyli by Schoch and Witzmann (Reference Schoch and Witzmann2009b). The genus Sclerocephalus forms a monophyletic group, with S. bavaricus, S. jogischneideri, and S. concordiae being successive sister taxa to the terminal sister taxa S. haeuseri and S. nobilis. Notably, Sclerocephalus nests below Cheliderpeton vranyi Fritsch, Reference Fritsch1877, Glanochthon angusta Schoch and Witzmann, Reference Schoch and Witzmann2009b, and G. latirostris (monophyletic) and the “archegosauroids” Melosaurus uralensis, Intasuchus silvicola, Archegosaurus decheni, Platyoposaurus stuckenbergi Trautschold, Reference Trautschold1884, and Australerpeton cosgriffi Barberena, Reference Barberena1998.

Figure 5. Topology obtained from cladistic analysis. Sclerocephalus concordiae n. sp. nests within the Sclerocephalidae clade, which forms the sister taxon to all other stereospondylomorphs.
The variant analysis with Sclerocephalus stambergi found the same general topology, albeit with Sclerocephalus bavaricus, S. stambergi, and Cheliderpeton vranyi forming an unresolved polytomy with all other species of Sclerocephalus, whose topology remained unaltered.
Discussion
The monophyly of Sclerocephalus is well supported, based particularly on synapomorphies in the preorbital region (lacrimal morphology, snout tip), as outlined by Schoch and Witzmann (Reference Schoch and Witzmann2009a). However, two homoplasies are apparent: S. bavaricus shares with Glanochthon spp. and “archegosauroids” the anterior extension of the premaxilla, and S. concordiae is similar to eryopids in the small size of the interpterygoid vacuities in large adults. The topology of branching within the genus Sclerocephalus is largely consistent with the stratigraphical occurrence of taxa: S. bavaricus forms the basalmost and stratigraphically oldest taxon, followed by somewhat younger S. concordiae and finally S. haeuseri and S. nobilis. S. jogischneideri, from the Upper Oberhof Formation of the Thuringian Forest Basin, which correlates with the highest Glan Group or lowest Nahe Group (Lützner et al., Reference Lützner, Kowalczyk, Schneider, Lützner and Kowalczyk2012), probably represents the stratigraphically youngest occurrence of Sclerocephalus. The genus Glanochthon was found here to nest with Cheliderpeton vranyi, Archegosaurus decheni, and other long-snouted stereospondylomorphs but according to new evidence may have originated from within the Sclerocephalidae (Schoch, Reference Schoch2021).
Discussion
The present findings provide some interesting insights into the evolution and diversification of temnospondyl endocrania. Although braincase elements and the epipterygoid have been reported from numerous taxa, endocrania are rarely known completely because even large temnospondyls often had poorly ossified braincases. This was already a topic for Case (Reference Case1933), who coined the term “progressive chondrification” in Triassic stereospondyls: compared with the heavily ossified endocrania of Edops craigi and Eryops megacephalus, large stereospondyls such as Cyclotosaurus robustus von Meyer and Plieninger, Reference Meyer and Plieninger1844, Parotosuchus nasutus von Meyer, Reference Meyer1858a, Metoposaurus diagnosticus von Meyer, Reference Meyer1842, and Batrachosuchus watsoni Haughton, Reference Haughton1925 had surprisingly little bone in the braincase and palatoquadrate (Watson, Reference Watson1919, Reference Watson1956, Reference Watson1962).
This view was challenged by more-detailed data on Benthosuchus spp. (Ochev, Reference Ochev1958, Reference Ochev1972; Getmanov, Reference Getmanov1989), Mastodonsaurus giganteus (Schoch, Reference Schoch2002b, Reference Schochc), and Gerrothorax pulcherrimus (Witzmann et al., Reference Witzmann, Schoch, Hilger and Kardjilov2012), all of which had much more fully ossified endocrania, adding to similar observations for Eryops megacephalus (Sawin, Reference Sawin1941) and Edops craigi (Romer and Witter, Reference Romer and Witter1942). Recently, the detailed studies of Robinson et al. (Reference Robinson, Ahlberg and Koentges2005) on Dendrerpeton acadianum Owen, Reference Owen1853 and of Arbez et al. (Reference Arbez, Dahoumane and Steyer2017) on the capitosaur Stanocephalosaurus amenasensis highlighted the importance of micro-CT data for the understanding of temnospondyl endocrania.
Neurocranium
The temnospondyl braincase falls into three distinct regions that usually remained well separated but occasionally are fused by co-ossification in heavily ossified taxa. These are the sphenethmoid region, the otic capsules, and the basisphenoid–basioccipital region, which forms the floor of the brain. In S. concordiae, all three regions are well ossified and were confluent, comparable to the condition in Eryops megacephalus and Edops craigi. Furthermore, as in Benthosuchus korobkovi Ivakhnenko, Reference Ivakhnenko1972 (Getmanov, Reference Getmanov1989), the roof of the otic capsules and their medial connection (tectum synoticum) is fully ossified in S. concordiae. Even in the giant taxon Mastodonsaurus giganteus, this condition was reached only in the largest specimens (Schoch, Reference Schoch2002b). These data suggest that co-ossification of the braincase was a late adult feature of hyperossified taxa, confirming that the holotype of S. concordiae was fully mature or even of old age. Like Eryops megacephalus, S. concordiae retained fully ossified basisphenoid and basioccipital portions, which were successively reduced in trematosauroids and capitosauroids (Case, Reference Case1933; Schoch, Reference Schoch2002a) but retained in plagiosaurids (Witzmann et al., Reference Witzmann, Schoch, Hilger and Kardjilov2012). Apart from these general observations, clear-cut evolutionary changes in the temnospondyl braincase are not readily apparent.
Epipterygoid
The palatoquadrate has rather divergent morphologies in temnospondyls for two reasons. First, it varies in the extent of ossification, ranging from fully cartilaginous in pedomorphic taxa to almost entirely ossified in large or heavily ossified forms. In most cases, bone is confined to the ascending process and the footplate, and this region is generally referred to as epipterygoid. Second, the ascending process and the articulation of the epipterygoid with the braincase underwent some evolutionary modification that gave way to divergent morphologies of the element (Fig. 6).
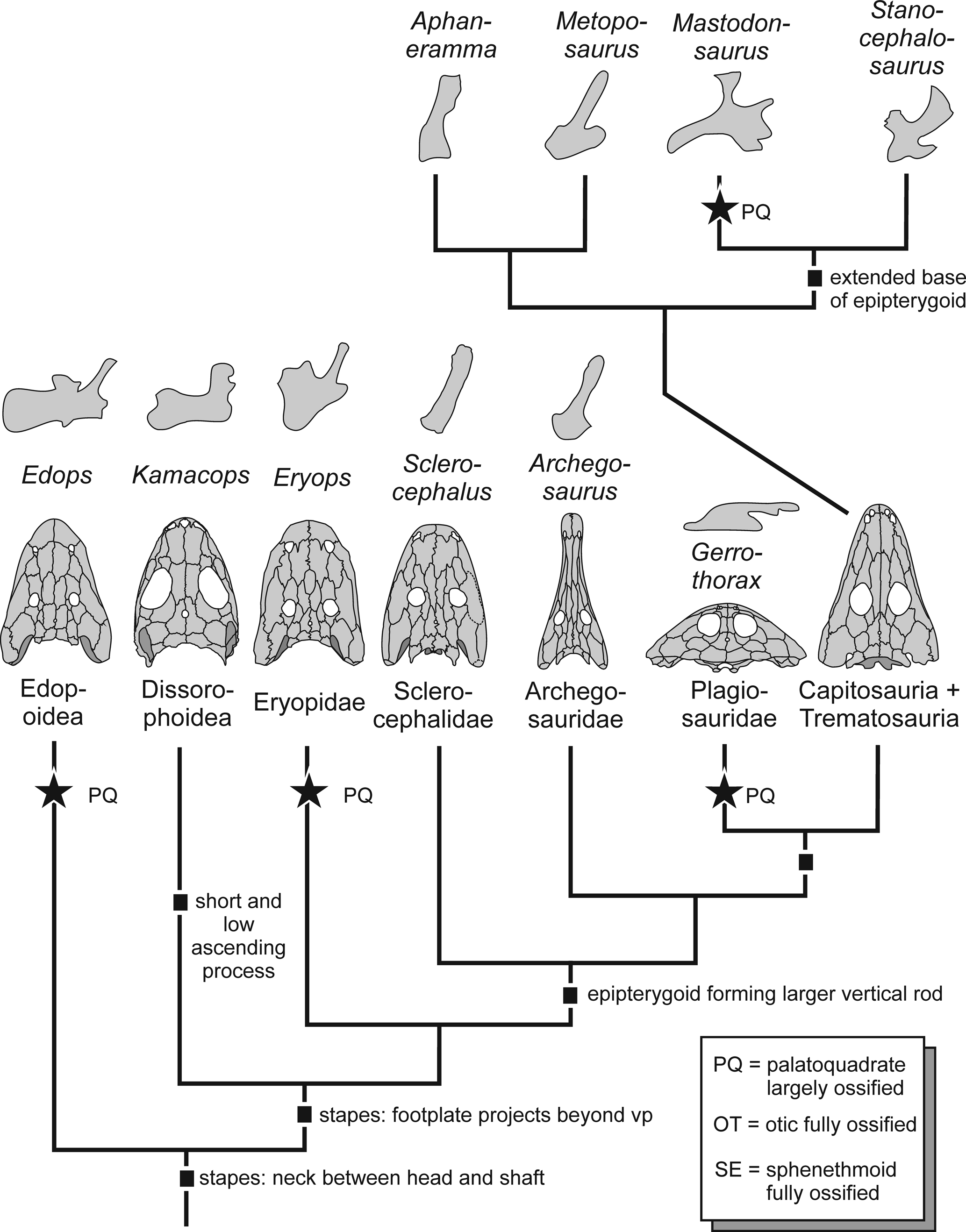
Figure 6. Evolution of endocranium in temnospondyls. Topology based on Schoch (Reference Schoch2013). Complete ossification of the palatoquadrate occurred repeatedly in the group, correlating with large size and/or heavy ossification of the skeleton. The morphology and complexity of the epipterygoid evolved in various different directions.
In the primitive condition of tetrapods, the palatoquadrate had five processes (Goodrich, Reference Goodrich1930; de Beer, Reference de Beer1937): pterygoid (anterolateral), ascending (dorsomedial), otic (dorsoposterior), quadrate (posterolateral), and basipterygoid (ventral). The basipterygoid process forms the base or footplate of the element, which is ossified in most taxa. In temnospondyls, the epipterygoid primarily connects the pterygoid with the parietal, and the element involved is the ascending process. This is most apparent in stereospondyls, where it forms a fully established, well-ossified strut (Schoch, Reference Schoch2002b), whereas in dvinosaurs and pedomorphic dissorophoids, it remained almost entirely unossified (Boy, Reference Boy1972, Reference Boy1995; Shishkin, Reference Shishkin1973). The ascending process usually extends from ventrolateral to dorsomedial, with the angle depending on the overall structure of the skull.
Tracing the morphology of the epipterygoid across the major clades of temnospondyls makes it apparent that (irrespective of size) well-ossified forms share a massive footplate (Edops craigi, Kamacops acervalis Gubin, Reference Gubin1980, Eryops megacephalus, stereospondyls) that occupied a substantial area on the dorsal side of the pterygoid. In these taxa, the footplate also forms the largest portion of the epipterygoid, whereas in other taxa the ascending process dominates (Dendrerpeton acadianum, Trimerorhachis insignis Cope, Reference Cope1878, basal stereospondylomorphs).
Most consistent with the primitive tetrapod condition, Edops craigi retained all processes, but the quadrate process was short, leaving a gap between the epipterygoid and a separate quadrate ossification. This taxon has a straight, dorsally unexpanded ascending process and a well-offset, rod-like otic process. Dendrerpeton acadianum has a much less extensive epipterygoid, comprising only a slender, Edops-like ascending process and a short footplate (Robinson et al., Reference Robinson, Ahlberg and Koentges2005).
In dissorophoids there is a substantial footplate, and the otic process is rudimentary (Peryella olsoni Carlson, Reference Carlson1987) or absent (Kamacops acervalis), and the ascending process is slender and aligned at an angle of 45° toward the braincase (Schoch, Reference Schoch1999; Ruta and Bolt, Reference Ruta and Bolt2006). Cross sections in Tersomius texensis revealed that the ascending process is much thinner than the ascending lamella of the pterygoid (Carroll, Reference Carroll1964).
In Eryops megacephalus, the otic process is massive, and unlike the rod-like outgrowth in Edops, it forms a tall wall-like structure paralleling the otic ossification that is located posterior to it. In stereospondylomorphs, the epipterygoid is not as extensive as in Eryops megacephalus, which is consistent with the lesser ossification throughout the skeleton. Archegosaurus decheni has a rod-like ascending process borne by a convex, poorly ossified footplate that lacks any other distinct processes. In Sclerocephalus concordiae, the footplate is unexpanded with an elongate oval articular surface, confined to the anterior margin of the pterygoid. In these relatively high skulls, the ascending process is much taller than in most Triassic taxa.
Within stereospondyls, the epipterygoid underwent further modification. In the highly apomorphic plagiosaurid Gerrothorax pulcherrimus, which has an extremely flattened skull, the epipterygoid is aligned nearly horizontally, forming a slender strut with a delicate, slightly curved shaft and well-expanded ventral and dorsal ends; as in Eryops megacephalus, there is a robust otic process (Witzmann et al., Reference Witzmann, Schoch, Hilger and Kardjilov2012). By contrast, trematosaurids and metoposaurids have a slender ascending process borne by a rounded footplate (Säve-Söderbergh, Reference Säve-Söderbergh1936, Reference Säve-Söderbergh1944; Sulej, Reference Sulej2007), much like in Archegosaurus decheni. In Wetlugasaurus angustifrons Riabinin, Reference Riabinin1930, Benthosuchus sushkini Efremov, Reference Efremov1929, and capitosaurs, the epipterygoid is generally large and more differentiated (Bystrow and Efremov, Reference Bystrow and Efremov1940; Schoch and Milner, Reference Schoch and Milner2000). They all share a large footplate extending from the basipterygoid region dorsally along the ascending lamella of the pterygoid. Furthermore, the shaft of the ascending process is flattened, more like a blade than a rod, and its dorsal end is expanded with two distinct facets. In Wetlugasaurus angustifrons and Benthosuchus sushkini, the more posterior of these facets attaches to the parietal, whereas the more anterior one contacts the sphenethmoid (Schoch and Milner, Reference Schoch and Milner2000). In Benthosuchus uralensis Ochev, Reference Ochev1966, there is a tiny additional process pointing anteriorly, which is much larger in some capitosaurs where it forms a plate extending parallel to the sphenethmoid (Eryosuchus garjainovi Ochev, Reference Ochev1966, Mastodonsaurus giganteus). In Mastodonsaurus giganteus, paralleled only by Gerrothorax pulcherrimus, the epipterygoid varies in extent with ontogeny, extending along the dorsal surface of the pterygoid both anteriorly (pterygoid process) and posteriorly (quadrate process) in large specimens.
The most striking difference in the osteology of the endocranium in stereospondyls is between trematosauroids and capitosauroids, with the former group having poorly ossified braincases and the epipterygoid confined to a delicate ascending process and the latter with extensively ossified, sometimes confluent braincase regions and a massive epipterygoid with a differentiated ascending process and an anterior extension contacting the sphenethmoid.
Paleoecology
Species of Sclerocephalus are known from a wide range of localities and horizons in the Saar-Nahe Basin that represent various different types of water bodies (Branco, Reference Branco1887; Ammon, Reference Ammon1889; Boy, Reference Boy1987; Boy et al., Reference Boy, Haneke, Kowalczyk, Lorenz, Schindler, Stollhofen, Thum, Lützner and Kowalczyk2012). The interplay between ontogeny and paleoecology in this taxon was analyzed by Schoch (Reference Schoch2009), who reported divergent ontogenetic trajectories among time-averaged samples (“populations”) of S. haeuseri in different lake deposits.
Boy et al. (Reference Boy, Haneke, Kowalczyk, Lorenz, Schindler, Stollhofen, Thum, Lützner and Kowalczyk2012) recognized the Sclerocephalus-bearing horizon at Lake Concordia as preserving the deepwater facies of a small lake. In Lake Concordia, Sclerocephalus was much more massively ossified than in all other lakes, and the adults lacked lateral line sulci and were probably capable of terrestrial locomotion, as indicated by the size and morphology of the robust humerus, which is quite consistent with that of Onchiodon labyrinthicus Geinitz, Reference Geinitz1862. Although morphologically most similar to the Jeckenbach (M6) sample, S. concordiae has extremely thickened dermal bones, a feature consistent with the samples from small lakes, such as Niederkirchen (M6) and Kappeln (M9), contrasting strongly with the weakly ossified specimens from large and deepwater bodies, such as from Odernheim (M8) and Humberg (M10), as analyzed in detail by Schoch (Reference Schoch2009). Therefore, the morphology and high extent of ossification in S. concordiae indicate that adults likely left the water, probably because the lake did not offer sufficient food resources. This is also supported by the find of a large bone referable to Sclerocephalus sp. in a channel-fill horizon below the lake sequence at the type locality (Boy, Reference Boy1987).
Acknowledgments
We thank A. Gehler for access to the Göttingen collection and loan of material and I. Rosin for skillfully preparing the specimen. The very helpful reviews by R. Werneburg and H.-D. Sues are gratefully acknowledged.
Data availability statement
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.ngf1vhhtf.






