INTRODUCTION
Schistosomiasis is an endemic tropical disease in many parts of the world, with 600 million people at risk due to inadequate hygiene, treatment and control programs. The pathology associated with hepatic schistosomiasis ranges from headache, lethargy and fever, to hepatic fibrotic obstruction, with the latter leading to complications such as portal hypertension, ascites and hepato-splenomegaly and, in extreme cases, death (Ross et al. Reference Ross, Bartley, Sleigh, Olds, Li, Williams and McManus2002). Recent meta-analysis of the disease indicates that, in terms of morbidity and mortality, the burden on individuals and communities due to schistosomiasis has increased worldwide with disability-adjusted life years (DALY) estimates rising approximately 20% from 40 to 48 per 100 000 in the past 20 years (Murray et al. Reference Murray, Vos, Lozano, Naghavi, Flaxman, Michaud, Ezzati, Shibuya, Salomon, Abdalla, Aboyans, Abraham, Ackerman, Aggarwal, Ahn, Ali, Alvarado, Anderson, Anderson, Andrews, Atkinson, Baddour, Bahalim, Barker-Collo, Barrero, Bartels, Basanez, Baxter, Bell and Benjamin2013).
The basis of the pathology arising from a schistosome infection is the host's immune response against parasite eggs trapped within tissues, leading to the formation of granulomas (Gryseels et al. Reference Gryseels, Polman, Clerinx and Kestens2006). The majority of disease pathology forms at the sites of egg deposition, which is mainly the liver in the case of Schistosoma mansoni and Schistosoma japonicum or the genitourinary tract in Schistosoma haematobium infections (Gryseels et al. Reference Gryseels, Polman, Clerinx and Kestens2006). Granuloma formation limits the damage caused by hepatotoxic compounds released by parasite eggs, by partitioning and/or elimination, but this process may also lead to extensive fibrogenesis within host tissues (Wilson et al. Reference Wilson, Mentink-Kane, Pesce, Ramalingam, Thompson and Wynn2007). Many of the general features of granuloma formation within the liver have been investigated, particularly with reference to the S. mansoni murine model (Burke et al. Reference Burke, Jones, Gobert, Li, Ellis and McManus2009). A CD4 + Th2 response drives the formation of hepatic granulomas leading to host pathology (Chuah et al. Reference Chuah, Jones, Burke, McManus and Gobert2014a ). Th2 cytokines, predominantly IL-4 and IL-13, promote this response, while IL-10, IFN-γ, IL13Rα2 and a subset of regulatory T-cells (T-Regs) and alternatively activated macrophages (AAM)ϕ, act to restrict the immune response and the pathology associated with schistosomiasis (Chuah et al. Reference Chuah, Jones, Burke, McManus and Gobert2014a ).
We have previously published studies on the transcriptional events that coincide with hepatic pathology using the schistosomiasis japonica murine model (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010a , Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert b ; Perry et al. Reference Perry, Burke, Stenzel, McManus, Ramm and Gobert2011; Chuah et al. Reference Chuah, Jones, Burke, Owen, Anthony, McManus, Ramm and Gobert2013). These studies present the analysis of egg-induced hepatic granuloma/fibrosis formation; with a focus on the chemokines and cytokines elevated during disease (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b ) and the events required for cellular recruitment to the liver and site of disease pathology (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010a ). Following studies provided a transcriptional framework to explain contrasting hepatic pathology in two inbred mouse strains infected with S. japonicum and indicated the importance of neutrophils in restricting pathology (Perry et al. Reference Perry, Burke, Stenzel, McManus, Ramm and Gobert2011). We then produced a gene atlas of the granuloma, increasing our resolution of the transcriptional responses associated with hepatic schistosomiasis (Chuah et al. Reference Chuah, Jones, Burke, Owen, Anthony, McManus, Ramm and Gobert2013). This latter study reaffirmed the importance of neutrophils in hepatic granuloma formation, and suggested two distinct roles for neutrophils extracellular traps in recruiting other immunological cells and in controlling fibrosis (Chuah et al. Reference Chuah, Jones, Burke, McManus, Owen and Gobert2014b ). These roles for neutrophils appear to be unique to the disease caused by S. japonicum and not S. mansoni. While our findings, and those of others, have provided a good appreciation of the cellular and molecular events that occur during hepatic schistosomiasis, they have been restricted to animal models and insights from clinical sources have been much more limited.
We have previously reported the collection and histological characterization of a large biopsy study in Hunan Province, The People's Republic of China, where patients underwent splenectomy and liver biopsy for the treatment of advanced schistosomiasis japonica (Li et al. Reference Li, Chen, Ross, Burke, Yu, Li, Zhou and McManus2011). Various clinical parameters were recorded including liver function tests, serology for hepatitis B/C/D, ultrasound diagnostic grading, and histology of liver biopsies. These cases represented a unique cohort of patients with advanced schistosomiasis in which the presence of S. japonicum eggs caused inflammation and distinct hepatic periportal fibrosis, leading to portal hypertension, hepatosplenomegaly and ascites (Li et al. Reference Li, Chen, Ross, Burke, Yu, Li, Zhou and McManus2011). Most patients were also sero-positive for hepatitis B. The degree of fibrosis ranged from grade II–III as determined by ultrasound and grade III–IV by histology. This work provided unique clinical data for advanced clinical schistosomiasis, with useful histological assessments of fibrosis. In the present paper we report the results of comprehensive transcriptional profiling of a subset of this unique clinical tissue collection, with the aim of providing some novel molecular insights on the aetiology of this chronic disease.
MATERIALS AND METHODS
Ethics statement
All work was conducted with approval from the human ethics committees of Hunan Institute of Parasitic Disease, China, and QIMR Berghofer Medical Research Institute, Australia (P919). Written informed consent was obtained from all subjects.
Patients
A subset of 13 liver biopsies, from long-term residents of the schistosomiasis-endemic Dongting Lake area, Hunan Province, People's Republic of China, was selected based on a clinical diagnosis of advanced schistosomiasis. The methods used to assess the clinical, parasitological and serological status of these patients have been reported (Li et al. Reference Li, Chen, Ross, Burke, Yu, Li, Zhou and McManus2011). All schistosomiasis patients were symptomatic and had splenomegaly (Hackett criteria ⩾II) according to published classification criteria. Patients were classified as either ‘schistosomiasis only’ ([S]) or ‘schistosomiasis and hepatitis B’ ([SH]) on the basis of fibrosis grading, schistosomiasis history indicated by questionnaire and serology results (Li et al. Reference Li, Chen, Ross, Burke, Yu, Li, Zhou and McManus2011). All patients were subsequently hospitalized and underwent surgery at Xiang-Yue Hospital, Yueyang, an affiliated hospital of Hunan Institute of Parasitic Diseases, in the Dongting Lake region. Surgical intervention included splenectomy, left gastric vein ligature and devascularization of the great curvature of the stomach for treatment of portal hypertension as described (Li et al. Reference Li, Chen, Ross, Burke, Yu, Li, Zhou and McManus2011). A further four liver biopsies were collected from control patients ([C]) who were from areas non-endemic for schistosomiasis. These samples were obtained from individuals undergoing abdominal surgery (for removal of gallstones) or emergency abdominal surgery following a traffic accident. A medical questionnaire was used to obtain details of medical history and demographics for the 17 patients, all of whom provided written informed consent allowing the collection and use of the biopsied material.
Microarray analysis
Total RNA was extracted from liver tissue biopsies using Trizol (Life Technologies, Carlsbad, USA) and an RNeasy Mini Kit (Qiagen, Valencia, USA). Total RNA quantity was measured using a Nanodrop-1000 (Nanodrop Technologies) and quality was assessed using an Agilent Bioanalyzer (Agilent Technologies, Foster City, USA). cRNA was synthesized using the Illumina Total Prep RNA Amplification kit (Ambion, Austin, USA). Microarray analysis was performed using Illumina SingleColor Human HT-12_V3 Whole Genome Expression Chips (Illumina, San Diego, USA). All gene expression data are publically available (NCBI's Gene Expression Omnibus; Series Accession Number: GSE61376).
Data analysis
Quality control of microarray data involved examination of intensity histograms of hybridization efficiency and noise using BeadStudio, version 3 (Illumina, San Diego, USA). All subsequent analyses were performed using Genespring GX, version 12.6 (Agilent Technologies, Foster City, USA). Expression values were normalized to the median and 75th percentile. Values less than 0·01 were set to 0·01. The data were then filtered for significant signal on the basis of detection score (d ⩾ 0·95, which equates to a confidence value of P ⩽ 0·05). At least 15 of 17 hybridizations had to pass these filtering criteria for a probe (gene) to be accepted. Analysis of variance (ANOVA, P ⩽ 0·05 using Benjamini and Hochberg correction for multiple testing) identified genes whose expression differed between groups. Principle components analysis was used to examine the relatedness between groups. When multiple copies of the same gene are reported, this represents multiple probes to the same gene represented on the microarray platform. To further demonstrate the relationship between individual samples, clustering analysis was performed. This involved using the standard GeneSpring hierarchical clustering algorithm with the following settings and normalized intensity values: Euclidean distance metric, Ward's linkage rule and clustering on both entities (genes) and conditions (samples). A heatmap was compiled ranging from −2·0 as green to +2·0 as red, whereas black colouring indicated signal intensities close to the median control values.
Ingenuity pathway analysis (IPA)
IPA was used to identify diseases and biofunctions or canonical pathways that were over-represented by the differentially expressed genes (Jimenez-Marin et al. Reference Jimenez-Marin, Collado-Romero, Ramirez-Boo, Arce and Garrido2009). Network analysis was performed to create an overview of the main signalling processes occurring in the schistosome- or schistosome and hepatitis-affected liver. Up-stream regulator analysis was performed to identify common upstream factors (eg transcription factors or drugs) that could be regulating the observed response.
Comparison with other hepatic diseases and a schistosome murine model
We compared our current findings with our previously published whole genome microarray analysis of a S. japonicum infection time course in C57 mice (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b ). Additionally, a survey was performed of the literature and, more specifically, the NCBI website (http://www.ncbi.nlm.nih.gov) with complete GEO datasets (GDS) related to liver disease and disease models. Four additional datasets were selected including Alcoholic hepatitis (GDS4389), Concanavalin A-induced fulminant hepatitis model (GDS3752), Hepatitis B virus associated acute liver failure (GDS4387) and Hepatitis C Virus infection effect on Huh7 hepatoma cells (GDS4160). For all auxiliary datasets, intensity values were either presented as normalized to time point 0 for time course experiments or as fold changes relative to control tissues. Data were matched to the current results using either Gene Symbols or Gene Bank Accession Number for each feature.
Additional datasets used included Alcoholic hepatitis of Homo sapiens (Affo et al. Reference Affo, Dominguez, Lozano, Sancho-Bru, Rodrigo-Torres, Morales-Ibanez, Moreno, Millan, Loaeza-del-Castillo, Altamirano, Garcia-Pagan, Arroyo, Gines, Caballeria, Schwabe and Bataller2013), concanavalin A (Con A) model analysis of Balb/C mice injected with ConA to induce fulminant hepatitis (Chen et al. Reference Chen, Zhu, Zhou, Li, Zhao, Wu, Wang, Liu and Chen2010), Hepatitis B in an analysis of clinical samples (Nissim et al. Reference Nissim, Melis, Diaz, Kleiner, Tice, Fantola, Zamboni, Mishra and Farci2012), and an analysis of human Huh7 hepatoma cells infected with Hepatitis C Virus and sampled at 6, 12, 18, 24 and 48 h post-infection (Blackham et al. Reference Blackham, Baillie, Al-Hababi, Remlinger, You, Hamatake and McGarvey2010).
RESULTS
Patient parameters for selection in the study
We selected 17 liver biopsy samples for RNA isolation and subsequent microarray analysis. This subgroup consisted of 4 controls [C], 6 with schistosomiasis only [S], and 7 with schistosomiasis and viral hepatitis B [SH] histories. A summary of the clinical and histological features of these patients is presented in Table 1. The term ‘Class’ indicates the broad classification of samples into Control [C] with no features indicating hepatitis or schistosomiasis history, schistosomiasis [S] indicating a diagnosed schistosomiasis fibrosis grading and positive schistosomiasis history, and schistosomiasis/hepatitis [SH] were patients presenting with both; schistosomiasis fibrosis grading and positive schistosomiasis history, and indications of a current (as antigen positive) or past hepatitis infection (antibody positive). ‘ID’ is a patient specific coding. ‘Egg’ indicates if the patient was positive or not for schistosome eggs in faecal samples by the Kato Katz diagnostic technique (Katz et al. Reference Katz, Chaves and Pellegrino1972). ‘HBV’ is indicative of either antibodies or antigen to hepatitis B. ‘Fibrosis Grade’ was determined by ultrasound. ‘Drinking History’ of alcohol intake consisting of the weekly consumption of more than two standard drinks was noted, ‘Sex’ was coded and ‘Age’ recorded, ‘Schisto history’ and ‘Schisto Treatment’ indicates whether each patient was aware of a past schistosome infection and the number of praziquantel treatments he/she had received in the previous 5 years. ‘Current HBV’ due to hepatitis B infection was also noted. No patients were antibody positive for hepatitis C or D.
Table 1. Pathological features and clinical history of patients whose liver biopsies were used in the microarray analysis

a Class shows the broad classification of samples into Control [C] with no features indicating any prior hepatitis or schistosomiasis history; Schistosomiasis [S] indicates a history of the disease and S1-S4 represents the schistosomiasis fibrosis grading obtained by ultrasound; Schistosomiasis/Hepatitis B [SH] were patients presenting with both diseases, with schistosomiasis fibrosis grading (S2-S4) and schistosomiasis history, and indications of a current (as antigen positive) or past (antibody positive) hepatitis infection.
b ID is the specific identification code for each patient.
c Egg indicates whether the patient was faecal egg-positive or egg-negative by the Kato Katz technique.
d HBV is indicative of either antibodies or antigen to Hepatitis B, HBsAb+ (antibody positive for Hep B surface protein); HBcAb+ (antibody positive for Hep B core protein); HBeAb+ (antibody positive for Hep B extracellular antigen).
e Fibrosis grading determined by ultrasound.
f Presentation with an enlarged liver by ultrasound and by palpation.
g Sex is coded.
h Age at time of liver biopsy.
i Drinking history of alcohol.
j Schistosome infection history determined by questionnaire.
k Schistosome treatment indicates if the patient was aware of a past schistosome infection and the number of praziquantel treatments they had received in the previous 5 years.
l Current Hepatitis B virus infection determined by the detection of hepatitis B surface antigen in liver sections.
Samples were selected on the basis of clinical history and RNA quality (RIN ⩾ 6) required to proceed to microarray analysis.
Transcriptional findings
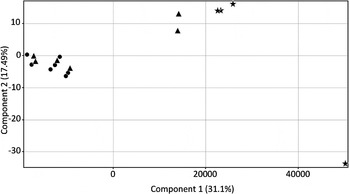
Filtering of microarray data for the 17 samples commenced with the complete 48 803 probes contained within the microarray platform which were then filtered for flags (in at least 15/17 samples) thereby reducing the number to 20 964. We then applied filters for detection score (d ⩾ 0·95 equivalent of P-value ⩽0·05, in at least 15/17 samples), reducing the gene list to 12 611 probes, and then differential expression (1-way ANOVA with multiple testing correction P ⩽ 0·05 (Benjamin-Hochberg), reducing the list to 3067 probes (online Supplementary Table 1). Of these 3067 probes, 1248 showed at least a ±2-fold change in expression in [S] and/or [SH] samples compared with the control [C] biopsy samples. Using the ±2-fold change cut-off, compared with [C], for [S], 12 genes were upregulated and 478 genes were downregulated, while for [SH] 52 genes were upregulated and 1167 genes were downregulated A total of 46 probes showed at least a 2-fold difference in expression between the [SH] and [S] groups. The relative transcriptional profiles of each of the samples are presented in Fig. 1; Control [C] samples were distinct from the other two disease groups (schistosomiasis only [S] and the combination schistosomiasis and hepatitis B [SH]) which had closer transcriptional profiles to each other. Principal components analysis (PCA) demonstrated that all of the [SH] samples clearly clustered together with three of the four [C] samples also clustering together. However, the [S] samples were divided between a close grouping with the [SH] samples (four samples), and two that were located between the [SH] and [C] groupings. The one outlier sample for [C] was from Patient ID XY204, who was older (61 years) than two of the other [C] samples but younger than the other (73 years). XY204 was the same sex as three other patients and could not be differentiated on the basis of drinking history or HBV status. Similarly the two outlier [S] samples (XY202, XY213) could not be separated by fibrosis grade, sex, age, drinking history, or schistosome treatment history. The diversity of the gene expression profiles demonstrated both control and diseased patients, shown in the PCA (Fig. 1) likely reflects the diversity of the clinical population sampled.

Fig. 1. Principal Component Analysis of the 17 liver biopsy samples utilizing 12 611 genes that passed quality control filtering. The controls [C] (stars), schistosomiasis [S] only patients (triangles) and the combination of schistosomiasis and hepatitis B [SH] (circles) are presented.
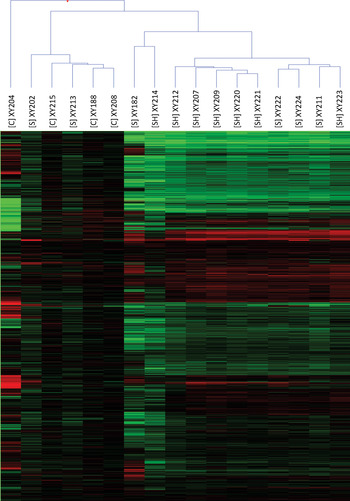
Hierarchical clustering of the 12 611 genes was performed to further present the gene profiles of individual samples (Fig. 2). The outlier samples for each group identified earlier were again evident as were the similarities of the [S] and [SH] samples in general. There was a clear general downregulation of genes of the disease samples (represented by the green colouration), whereas the upregulation of genes (in red) was less evident.

Fig. 2. Heatmap of the hierarchical clustering of all 17 liver biopsy samples utilizing 12 611 genes that passed quality control filtering. The controls [C], schistosomiasis [S] only patients and the combination of schistosomiasis and hepatitis B [SH] are presented. The patient ID is also included to allow cross-referencing with Table 1. Gene intensities normalized to controls are shown, ranging from downregulated to −2·0 in green, upregulated to 2·0 in red, with control intensities shown in black.
While different fibrosis grades were present with both the [S] and [SH] groups, comparisons between different grades was not possible due to the limited number of replicated samples available. This would be a useful approach for the future if larger numbers of samples can be obtained.
Ingenuity pathway analysis and keyword searches
Ingenuity pathway analysis of the 3067 identified genes by 1 way ANOVA for the [S] and [SH] groups revealed a similar functional profile (online Supplementary Table 2). Overall, the transcriptional profiles of these two patient cohorts were consistent with chronic liver disease of infectious aetiology in which significant immunomodulation has occurred. Over-represented biofunctions included ‘protein synthesis’, ‘cell cycle’ and ‘energy metabolism’. The most prominent feature was identification of EIF2 (Eukaryotic Initiation Factor 2) signalling pathway as an enriched pathway for both [S] and [SH] (P-value 6·6 × 10−8), with key gene components downregulated in both disease groups when compared with controls. The downregulation of genes in this pathway and others associated with the ‘protein synthesis’ biofunction indicates likely impairment of protein synthesis in the liver of patients with [S] and/or [SH]. This is a common feature of chronic liver disease and is an indicator of reduced liver function (Krastev, Reference Krastev1998).
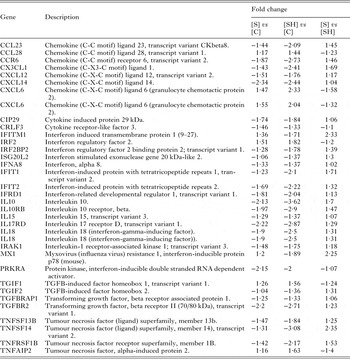
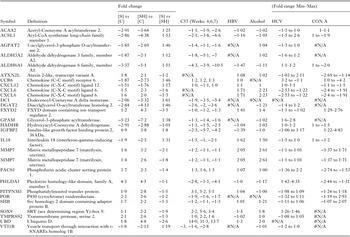
Most immune associated genes were downregulated or unchanged in both [S] and [SH] disease groups (See Table 2). There was no change in the expression of Th2 cytokines compared with controls while there was significant downregulation of IL-10 and IL18. Due to the immune-mediated nature of schistosome-induced pathology, we specifically examined the expression of genes associated with ‘inflammation’ and ‘immune responses’ using IPA keyword based searches. Genes associated with inflammation are presented in Table 3 and generally were downregulated in the [S] and [SH] samples. In a murine model of acute hepatic schistosomiasis japonica (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b ) (up to 7 weeks post cercarial challenge) none of these genes were generally differentially expressed, with the exception of IL10 which approached upregulation at 1·9-fold. A set of three genes associated with granulocytes, MMP7, CLDN7 and CXCL6 ([S] 1·55, 1·54, 1·55, [SH] 3·21, 2·17, 2·04), were upregulated in [SH] samples, but a lower level of upregulation, less than a 2-fold cut-off, was noted for the [S] samples. Similarly, functions and genes related to the extracellular matrix (FERMT2, SDC2 and VHL, [S]: −2·20, −1·96, −1·61; [SH] −3·23 −3·02 −2·19) were downregulated in both the [S] and [SH] disease groups, as were genes associated with connective tissue cells (Table 4). Antiviral genes IFITM1 (1·4, −1·7) and MX1 (1·2, −1·9) were also identified in annotation searches and were differentially expressed in [S] and [SH] patients.
Table 2. Immune associated genes, identified from genes shown to be differentially expressed using keyword searches for ‘interleukin’, ‘chemokine’, ‘cytokine’, ‘interferon’ or ‘tumour necrosis factor’

Fold change is presented as schistosomiasis only [S] or schistosomiasis and viral hepatitis B [SH] relative to controls [C], and schistosomiasis only [S] relative to schistosomiasis and viral hepatitis B [SH].
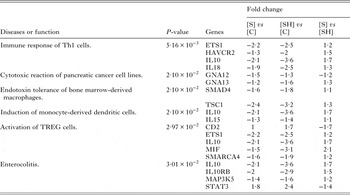
Table 3. Diseases or functions annotation associated with ‘Inflammation’ identified by Ingenuity Pathway Analysis of the 3065 differentially expressed genes

Fold change is presented as schistosomiasis only [S] or schistosomiasis and viral hepatitis B [SH] relative to controls [C], and schistosomiasis only [S] relative to schistosomiasis and viral hepatitis B [SH].
Table 4. Differentially expressed genes of novel biological functions selected from Ingenuity Pathway Analysis

Fold changes are presented as schistosomiasis only [S] or schistosomiasis and viral hepatitis B [SH] relative to controls [C], and schistosomiasis only [S] relative to schistosomiasis and viral hepatitis B [SH]
Comparison of [S] and [SH] profiles
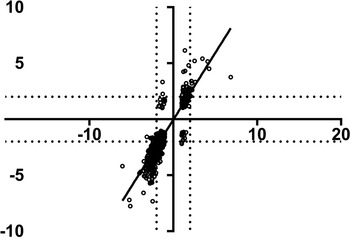
Final interrogation of the transcriptional dataset was undertaken to differentiate between [S] and [SH] patients. This involved consideration of all differentially expressed genes with a ±2-fold change between the two disease groups [S] and [SH], which yielded 46 candidates (online Supplementary Table 1). No genes were upregulated in one group and downregulated in the other by more than the ±2-fold cut-off. Correlation of the two groups generally presented a high degree of similarity with r = 0·85 (P value <0·0001) (Fig. 3). For example, TSPAN33 (tetraspanin 33), which ensures normal erythropoiesis, through the differentiation of progenitor cells (Heikens et al. Reference Heikens, Cao, Morita, Dehart and Tsai2007), was downregulated in both samples although downregulation of the gene was more pronounced in [S] (−1·73-fold) than in [SH] (−3·9-fold). Other examples included genes such as TACSTD1, LEPR, UBD, LGALS4 and FXYD2, which were moderately up-regulated in the [S] group (1·2–2·6-fold) but exhibited increased upregulation in the [SH] group (4·3–6·1-fold). Similarly, genes that were downregulated in the [S] group, such as RPS15A, NDUFB10, BLVRB and PCOLCE2 (−2·5 to −2·8-fold), were more down-modulated in the [SH] group (−5·7 to −7·3-fold)

Fig. 3. Correlation between the relative fold changes for 3067 genes of schistosomiasis only [S] samples (x-axis) compared with schistosomiasis and viral hepatitis B [SH] samples (y-axis) relative to control samples. A ±2-fold change is marked with a dotted line and used to identify genes with a similar differential expression patterns between the [S] and [SH] sample groups. The linear regression is plotted with an R 2 = 0·85 and P value <0·0001.
In general, when normalized to controls [C] enrichment of disease, biofunctions and canonical pathways were more pronounced in the [SH] group compared with the [S] group. Increased downregulation of genes associated with, for example, EIF2 Signalling (P-value 1·58E-13), Protein Ubiquitination (3·09 × 10−8), Regulation of eIF4 and p70S6K Signalling (2·45 × 10−7), and mTOR Signalling (P value 1·02 × 10−6) (online Supplementary Table 2). This suggests that liver function was more compromised in the [SH] compared with the [S] patients a feature consistent with the increased pathology in this group of patients.
Ubiquitin has been reported as a biomarker for non-alcoholic Steatohepatitis (NASH) in humans (Banner et al. Reference Banner, Savas, Zivny, Tortorelli and Bonkovsky2000). The identification of Post-Translational Modification: ‘ubiquitination of protein’ from IPA (P-value 1·92 × 10−7) showed that almost all of the 68 genes in the pathway were moderately downregulated in both the [S] and [SH] groups with only UBD (ubiquitin D) being upregulated 1·8-fold for [S] and 4·8-fold for [SH] (See online Supplementary Tables 1 & 2).
The most upregulated gene in [S] patients was IGFBP2 (6·9-fold) and this was also upregulated in [SH] but to a lesser extent (3·8-fold).
Comparison with the mouse model of schistosomiasis japonica
A comparison of the transcriptional profiles of human hepatic schistosomiasis and our previously published study of acute S. japonicum infection in the livers of C57 BL/6 mice (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b ) indicated few similarities. Of the 3067 differentially expressed genes in the human samples, 1625 were also differentially expressed in the mouse model (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b ). A Spearman's correlation of expression of these genes in the [S] or [SH] samples compared with the livers from mice 7 weeks post cercarial challenge (the latest time point analysed) produced an R 2 of [S]/C57 = 0·1 (P value <0·001) and [SH]/C57 = 0·07 (P value = 0·0034) (Fig. 4). A subset of genes did show similar gene expression levels in human hepatic and murine schistosomiasis (online Supplementary Table 3). Genes that were: upregulated in both [S] and mice (0 genes), upregulated in both [SH] and mice (3 genes, UBD, PITPNM1, SOX9, see Table 4), downregulated in both [S] and mice (54 genes) and downregulated in both [SH] and mice (109 genes). Common genes that were downregulated in [S] and [SH] compared with [C], and also downregulated in murine schistosomiasis included those associated with fatty acid metabolism (ACSL1, ALDH6A1, DCI, HADHB and ACAA2), vesicle transport (VTI1B), and proinflammatory IL18, the only gene related to immunology.

Fig. 4. Correlation between the relative fold changes of schistosomiasis japonica [S] (left) or schistosomiasis and viral hepatitis B [SH] (right) liver biopsy samples on the y-axis and C57 (7 weeks post cercarial challenge from (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b )) on the x-axis, for 1625 genes. Regions of similar gene expression are boxed; a ±2-fold change is marked with a dotted line and used to identify genes with a similar expression pattern in the human and murine studies.
Comparison with other hepatic diseases
Distinct differences were apparent on comparing transcriptional responses demonstrated in the schistosomiasis datasets to other human hepatic diseases. The transcriptional profiles of genes that were upregulated in either [S] or [SH] were compared with other hepatic disease datasets (see Table 5). For example, the neutrophil chemokine CXCL6 (upregulated 1·5 to 2·3-fold in [S] or SH) is downregulated in HCV (−2·5-fold) and Con A (−2·4-fold) disease states, but is upregulated in Alcohol (2·2). MMP7 was upregulated in [S] and [SH] (1·4–3·2-fold) to a similar extent compared with some other hepatic diseases examined (up-regulated in HBV, 2·0-fold; Con A,1·7-fold; and alcohol,2·6-fold). In addition a further 283 genes that were downregulated in [S] or [SH], compared with normal controls [C] are presented in online Supplementary Table 4. Of these downregulated genes, some (see online Supplementary Table 4 and Table 5) were related to Ribosome Function (MRPL13, RPL21, RPL23, RPL3, RPS15, RPS15A, RPS16, RPS3A), Fatty Acid Metabolism (ACSL1, ALDH3A2, DCI, HADHB, ACAA2) and Glycolipid Function (AGPAT2, ALDH3A2, DGAT2, GPAM). The downregulation of these genes is not mirrored in other hepatic diseases (HBV, HCV, Con A and Alcohol) examined (See online Supplementary Table 4) but has been observed in murine models of schistosomiasis reported by our group (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b ).
Table 5. Selected genes from [S] or [SH] liver biopsies with relative fold changes with corresponding transcriptional changes reported in other hepatic diseases and a murine model of schistosomiasis

Current study comparison of fold change of schistosomiasis only [S], schistosomiasis and viral hepatitis B [SH] and [C] controls. C57 is a previously published murine study of S. japonicum infection time course (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b), four additional datasets were selected including Alcohol, alcoholic hepatitis, Con A, concanavalin A-induced fulminant hepatitis model, HBV, hepatitis B virus associated acute liver failure and HCV, hepatitis C virus infection effect on Huh7 hepatoma cells
We compared our data with those obtained by Nissim and colleagues (Nissim et al. Reference Nissim, Melis, Diaz, Kleiner, Tice, Fantola, Zamboni, Mishra and Farci2012) who examined the gene profile of clinical cases of acute liver failure attributed to HBV. A number of genes in the [S] and/or [SH] patients correlated with genes up or downregulated in the HBV study (online Supplementary Table 4). Some downregulated genes in the [S] subjects were more downregulated in [SH], similar to the pattern of downregulated genes reported by Nissim et al. (Reference Nissim, Melis, Diaz, Kleiner, Tice, Fantola, Zamboni, Mishra and Farci2012), possibly reflecting the differences between the [S] and [SH] patients. These genes included (fold change [S]|[SH]|HBV): cysteine dioxygenase, type I (CDO1; −2·17|−3·17|−1·71); thyroid hormone responsive (THRSP; −3·30|−4·46|−1·67); procollagen C-endopeptidase enhancer 2 (PCOLCE2; −2·46|−7·32|−1·64); aldehyde dehydrogenase 6 family, member A1 (ALDH6A1; −3·37|−5·10|−1·47); and growth hormone receptor (GHR; −2·19|−2·71|−1·45). These genes represent biological function related to metabolism, tissue remodelling and hormone regulation, While a small number of genes were upregulated in the HBV study could contribute to upregulation seen in our [S] and to a great extent [SH], these included the tissue remodelling gene matrix metallopeptidase 7 (MMP7; 1·55|3·21|2·05), and chemokine (C-X-C motif) ligand 6 (CXCL6; 1·55|2·04|1·71).
DISCUSSION
Pathway analysis and gene ontology analysis of moderately differentially expressed genes presented in either in [S] or [SH] patients compared with controls, demonstrated many potentially important biological functions reflecting the transcriptional profile of human hepatic schistosomiasis. Data generated from murine schistosomiasis studies are generally obtained from short-term acute infections, of 2–3 months duration only. This is direct contrast to the results obtained here from a clinical cohort with patients having been infected/cured multiple times over a long period, in some instances, up to several years at a time. Consequently, direct comparisons between experimental murine schistosomiasis and the natural progression of the human disease are problematical and somewhat artificial but the current study emphasizes the value and novelty of the clinical data we describe. The limitations and difficulties in undertaking animal studies for extended periods make progress in our understanding of chronic hepatic schistosomiasis challenging.
mTOR signalling (mechanistic target of rapamycin) was one of the prominent pathways identified from examination of differentially expressed genes in the livers of schistosomiasis only [S] or schistosomiasis and viral hepatitis B [SH] patients, compared with controls [C]. It is known that mTOR signalling regulates basic metabolic functions in the liver and is disrupted in both metabolic (diabetes, obesity) and non-metabolic (aging, neurodegeneration, cancer) disease states in a wide range of tissues (Laplante and Sabatini, Reference Laplante and Sabatini2012). Furthermore, mTOR signalling is significant in promoting fibrosis via macrophages and myofibroblasts in both kidney and intestinal tissues (Whaley-Connell et al. Reference Whaley-Connell, Habibi, Panfili, Hayden, Bagree, Nistala, Hyder, Krueger, Demarco, Pulakat, Ferrario, Parrish and Sowers2011; Chen et al. Reference Chen, Chen, Wang, Peng, Sun, Liu and Liu2012; Jiang et al. Reference Jiang, Xu, Mao, Li, Fang, Zhou, Liu, He, Zhao, Yang and Dai2013). Therefore, in our study it is noteworthy that in [S] and even [SH] patients, with a history of both viral hepatitis B and schistosomiasis, this pathway was depressed, with 45 of 50 genes in the pathway downregulated, which may reflect a lack of active gene transcription for fibrosis in these chronic [S] and [SH] patients. This is in direct contrast to reports of the upregulation of mTOR in the livers of subjects with both hepatitis C (Bose et al. Reference Bose, Shrivastava, Meyer, Ray and Ray2012) and hepatitis B (Yen et al. Reference Yen, Lin, Yen, Tsai, Tsai, Chang, Huang, Lin, Chiang and Chang2012), and the correlation of this elevated expression with the increased likelihood of hepatocellular carcinoma (Wang et al. Reference Wang, Jin, Jin and Wang2014). In this respect, it has been suggested that the risk of hepatocellular carcinoma development is elevated in chronic schistosomiasis associated with HBV infection, compared with HBV infections solely (Ross et al. Reference Ross, Bartley, Sleigh, Olds, Li, Williams and McManus2002). A possible mechanism explaining the downregulation of the mTOR pathway in the current study could involve the upregulation of DNA damage response genes, such as caspase 9 (CASP9, [S] 1·2-fold, [SH] 1·5-fold), or genes associated with oxidative stress such as FXYD2 (FXYD domain containing ion transport regulator 2) and POR (P450 (cytochrome) oxidoreductase) (See Table 5), as well as the reduction in gene expression of multiple ATPases (Adenosine triphosphate) and ATP synthases (see online Supplementary Table 1). It is known that reduced metabolism, leading to reduced ATP levels and increased hypoxia and DNA damage causes constitutive downregulation of the PI3K-mTORC1 axis (reviewed in (Laplante and Sabatini, Reference Laplante and Sabatini2012)). Furthermore, apoptosis in hepatocarcinoma cells has been linked to telomerase activity and the downregulation of mTOR signalling (Bu et al. Reference Bu, Jia, Wang, Guo, Wu and Wei2007), which may mirror the situation reported here. Another possible mechanism that could be responsible for the downregulation of the mTOR pathway could involve the dis-regulation of growth hormone in the liver, as has been suggested for the Ames dwarf mouse model (Sharp and Bartke, Reference Sharp and Bartke2005). Here, [S] patients had reduced levels of growth hormone receptor (GHR −2·7-fold), which would reflect reduced growth hormone action in this system and may impact on the mTOR pathway as a whole.
Interferon-induced transmembrane (IFITM1) is an interferon-induced protein which has known antiviral activity as demonstrated in HCV-infected hepatocytes (Raychoudhuri et al. Reference Raychoudhuri, Shrivastava, Steele, Kim, Ray and Ray2011). Despite its nominated antiviral activity this gene was surprisingly downregulated (−1·7-fold) in [SH] but upregulated (1·4-fold) in [S] patients (Table 2). Similarly, another antiviral gene MX1 (or MxA: myxovirus resistance 1), which is also interferon-inducible, was similarly modulated ([S] 1·2-fold, [SH] −1·9-fold). A direct effect of interferon on these genes can be discounted as there was a similar expression pattern of other interferon genes or related genes in both pathology groups (Table 2). The potential effect of antiviral genes in response to a schistosome infection is intriguing area for future research.
The most significantly upregulated gene in [S] patients was insulin-like growth factor binding protein 2 (IGFBP2) which was increased 6·9-fold relative to controls. IGFBP2 is involved in many important biological functions such as synthesis of proteins and lipids, increasing the mass and size of the liver, migration of vascular smooth muscle cells and proliferation of fibroblasts (Firth and Baxter, Reference Firth and Baxter2002). It should be noted that all of the [S] and [SH] patients presented with enlarged livers (Table 1), and this may reflect the elevated levels of IGFBP2. Increased IGFBP2 has also been reported to be associated with hepatitis C-associated osteosclerosis (HCAO), a rare disorder characterized by a marked increase in bone mass during adult life, a feature not normally seen in either control Hepatitis C or Hepatitis B patients (Khosla et al. Reference Khosla, Hassoun, Baker, Liu, Zein, Whyte, Reasner, Nippoldt, Tiegs, Hintz and Conover1998). However, hepatic engorgement is more likely due to portal hypertension which is a key feature of chronic schistosomiasis (Li et al. Reference Li, Chen, Ross, Burke, Yu, Li, Zhou and McManus2011). It is noteworthy that two other IGF-binding proteins – IGFBP3 and IGFBP5 ([S] −1·6, −4·0, [SH] −2·6, −4·8) – were downregulated in both [S] and [SH] samples. While IGFBP2 was also upregulated in [SH] (3·8-fold), the gene most highly upregulated (6·1-fold increase relative to controls) in these samples was tumour-associated calcium signal transducer 1 (TACSTD1, also known as EpCAM), whose expressed protein is known to be associated with cellular adhesion (Balzar et al. Reference Balzar, Winter, de Boer and Litvinov1999). TACSTD1 is a hepatic stem cell marker previously studied in carcinomas (Yamashita et al. Reference Yamashita, Budhu, Forgues and Wang2007), that is increased in Hepatitis B and hepatic carcinoma (Kimura et al. Reference Kimura, Kondo, Kogure, Kakazu, Ninomiya, Iwata, Morosawa and Shimosegawa2014) as well as in alcoholic hepatitis (see GEO accession: GDS4389 and (Affo et al. Reference Affo, Dominguez, Lozano, Sancho-Bru, Rodrigo-Torres, Morales-Ibanez, Moreno, Millan, Loaeza-del-Castillo, Altamirano, Garcia-Pagan, Arroyo, Gines, Caballeria, Schwabe and Bataller2013)).
The most downregulated gene in [S] was the uncharacterized C10orf116 (−6·0-fold) followed by GPAM (glycerol-3-phosphate acyltransferase, mitochondrial) at −5·2-fold. C10orf116, also known as Adipose most abundant gene transcript 2 (APM2), is known to be expressed in adipose tissue and over expression of the encoded protein leads to increased cellular proliferation and increased glucose transport but its full functional significance, particularly in the liver, is unknown (Chen et al. Reference Chen, Zhou, Zhou, Zhu, Ji, Shi, Qiu and Guo2013). GPAM, associated with many aspects of fatty acid metabolism and the synthesis of glycerolipids (Wendel et al. Reference Wendel, Lewin and Coleman2009), was also downregulated in [SH] (−7·20-fold). The gene with the lowest expression in [SH] was TXNIP (−7·9-fold) which was also highly downregulated in [S] (−5·1-fold). TXNIP is related to lipid metabolism and is reported to be necessary for the replication of Hepatitis C (Blackham et al. Reference Blackham, Baillie, Al-Hababi, Remlinger, You, Hamatake and McGarvey2010).
Relative to controls, most cytokines and chemokines were downregulated in [S] and [SH] patients. We have previously reported that neutrophils are an important cell type recruited to the site of granuloma formation in a murine model of schistosomiasis japonica (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b ; Perry et al. Reference Perry, Burke, Stenzel, McManus, Ramm and Gobert2011; Chuah et al. Reference Chuah, Jones, Burke, Owen, Anthony, McManus, Ramm and Gobert2013, Reference Chuah, Jones, Burke, McManus, Owen and Gobert2014b ). One chemokine that was upregulated in the livers of [S] and [SH] patients was CXCL6, which recruits neutrophils, a prominent feature of the murine S. japonicum model. CXCL6 is also upregulated in alcoholic cirrhosis of the liver and HBV (see Table 5). A decrease in the serum levels of other chemokines, such as granulocyte associated CXCL5, has been reported in patients with chronic liver disease (Tacke et al. Reference Tacke, Zimmermann, Trautwein and Schnabl2011) but our observations represent a decrease in the liver tissue itself rather than in the peripheral blood.
A comparison of our findings with a previous study examining concanavalin A-induced liver damage in an acute murine model (Cao et al. Reference Cao, Chen, Li, Wu, Wang and Chen2010) identified many genes in common. These included upregulated genes such as IGFBP2 and downregulated genes such as B2M. Genes such as Granulin (GRN) were downregulated in [S] (−1·9) and [SH] (−3·3), but these remained unchanged in mice with concanavalin A-induced hepatitis. An early study showed that beta-2-microglobulin (B2M) () was as a serum marker for many types of liver diseases including acute viral hepatitis, chronic persistent or active hepatitis and liver cirrhosis (Revillard, Reference Revillard1980), but this was not apparent in the [S] and [SH] patients (downregulated [S] −2·7 and [SH] −4·8) or the results of other hepato-pathologies we correlated in this study, all which assessed gene expression directly in liver tissue. This might be expected since tissue gene profiling studies indicate that gene expression in liver tissue is relatively low, with higher expression levels being apparent in many peripheral blood immune cells (Wu et al. Reference Wu, Macleod and Su2013). GRN had decreased expression in both [S] and [SH], but its levels were generally unchanged in other hepatic disease conditions we considered (See online Supplementary Table 4), and in our published murine schistosomiasis studies (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010b ). GRN is a well charactered growth factor associated with many types of cancer including hepatocellular carcinomas (Cheung et al. Reference Cheung, Wong, Leung, Chen, So, Ng and Fan2004). The expression of a GRN-like molecule by the human liver fluke, Opisthorchis viverrini, has been linked to the development of cholangiocarcinoma (Smout et al. Reference Smout, Laha, Mulvenna, Sripa, Suttiprapa, Jones, Brindley and Loukas2009) and tissue repair (Bateman and Bennett, Reference Bateman and Bennett2009). However, there is limited information as to whether reduced expression of the mammalian gene may lead to liver pathology; of the studies reporting a reduction in GRN levels, all have been associated with neurological conditions, rather than hepatic-related pathologies (Capell et al. Reference Capell, Liebscher, Fellerer, Brouwers, Willem, Lammich, Gijselinck, Bittner, Carlson, Sasse, Kunze, Steinmetz, Jansen, Dormann, Sleegers, Cruts, Herms, Van Broeckhoven and Haass2011; Dopper et al. Reference Dopper, Seelaar, Chiu, de Koning, van Minkelen, Baker, Rozemuller, Rademakers and van Swieten2011). Consequently, the reduction in GRN expression we recorded is a unique finding and worthy of future investigation to determine the role of this protein in the livers of advanced schistosomiasis patients.
Matrix metallopeptidase 7 (MMP7) was upregulated in [S] with even higher expression in [SH] patients (Table 5), a situation not replicated in chronic murine schistosomiasis (Chuah et al. Reference Chuah, Jones, Burke, Owen, Anthony, McManus, Ramm and Gobert2013). MMP7 degrades proteoglycans, fibronectin, elastin and casein and has been implicated in wound healing and tissue remodelling (Huang et al. Reference Huang, Chuang, Chou, Wu, Chen, Wang, Chen, Chen and Tai2005) and in the fibroproliferative process in the liver during chronic hepatitis C (Lichtinghagen et al. Reference Lichtinghagen, Michels, Haberkorn, Arndt, Bahr, Flemming, Manns and Boeker2001). Our results suggest that MMP7 could be a potential candidate biomarker for chronic human schistosomiasis, although the induction of this gene in cirrhosis is also seen in other aetiologies (see Table 5) which would limit its diagnostic specificity. The suggestion that MMPs, and more specifically MMP7, may prove useful as biomarkers of fibrosis has been made previously for pulmonary disease (Rosas et al. Reference Rosas, Richards, Konishi, Zhang, Gibson, Lokshin, Lindell, Cisneros, Macdonald, Pardo, Sciurba, Dauber, Selman, Gochuico and Kaminski2008; Song et al. Reference Song, Do, Jang, Colby, Han and Kim2013), and the results presented here support their potential value as biological markers of hepatic fibrosis in schistosomiasis. In support of these findings MMPs, measured in SEA stimulated peripheral blood mononuclear cells (PBMCs) as well as serum, have been related to S. japonicum infection of humans (Fabre et al. Reference Fabre, Wu, PondTor, Coutinho, Acosta, Jiz, Olveda, Cheng, White, Jarilla, McGarvey, Friedman and Kurtis2011; McDonald et al. Reference McDonald, Cheng, Jarilla, Sagliba, Gonzal, Amoylen, Olveda, Acosta, Baylink, White, Friedman and Kurtis2014).
Further comparison of the transcriptional findings in subjects with schistosomiasis and other disease states resulted in the identification of individual genes associated with key biological hepatic functions. Three chemokine receptors or ligands that were differentially expressed in [S] or [SH] patients are associated generally with liver disease, including chemokine (C-C motif) receptor 6 (CCR6), CXCL6 and chemokine (C-X-C motif) ligand 12 (CXCL12). CXCL6 was the only gene upregulated over 2-fold in [SH] (2·3-fold, 1·5-fold in [S]), while the others were downregulated (see Table 5). These genes were also shown to be differentially expressed in the C57 murine model of schistosomiasis and in other liver diseases we examined. Upregulation of CXCL6 is a feature of Alcohol disease and HBV, whereas it is downregulated in concanavalin A-induced hepatitis. The functional roles of these chemokines in liver disease have been reviewed (Sahin et al. Reference Sahin, Trautwein and Wasmuth2010), with CCR6 shown to be associated with T cells, B cells and dendritic cells; CXCL6 is associated with neutrophils and monocytes; and CXCL12 is expressed in most resident hepatic cells including hepatocytes, vascular endothelial cells and stellate cells, as well as in immunological cells including monocytes, basophils and B cells. The upregulation of neutrophil chemokines is a well-known feature of chronic schistosomiasis japonica, but not chronic schistosomiasis mansoni, in mice (Chuah et al. Reference Chuah, Jones, Burke, Owen, Anthony, McManus, Ramm and Gobert2013, Reference Chuah, Jones, Burke, McManus and Gobert2014a ). In this human study of chronic schistosomiasis, the changes in neutrophil gene expression we observed may be reflective of tissue remodelling rather a role in inflammation.
Conclusions
Our previous studies with the murine model of schistosomiasis japonica increased understanding of the transcriptional and cellular events that regulate S. japonicum-induced granuloma formation and fibrosis (Burke et al. Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert2010a , Reference Burke, McManus, Ramm, Duke, Li, Jones and Gobert b ; Perry et al. Reference Perry, Burke, Stenzel, McManus, Ramm and Gobert2011; Chuah et al. Reference Chuah, Jones, Burke, Owen, Anthony, McManus, Ramm and Gobert2013). Here we provide some insight on the molecular events that occur in Chinese patients with chronic hepatic schistosomiasis japonica. The results provide a comprehensive transcriptional profile of human schistosomiasis and demonstrate similarities and differences with other liver diseases. The unique features we report of gene expression in human liver, in conjunction with previous reports of the formation and recovery and the cellular composition of schistosome-induced granulomas, improve our understanding of the molecular immunopathology and general physiological status underlying the aetiology of schistosomiasis.
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0031182015000682.
ACKNOWLEDGEMENTS
Y. S. L is an ARC Future Fellow; D. P. M is a NHMRC Senior Principal Research Fellow and Senior Scientist at QIMR Berghofer.
FINANCIAL SUPPORT
This work was supported by the National Health and Medical Research Council (NHMRC) of Australia.