


INTRODUCTION
Panopea clams are known for their extreme longevity among invertebrates. For instance, Panopea generosa Gould, 1850 has a record longevity of 168 years (Bureau et al., Reference Bureau, Hajas, Surry, Hand, Dovey and Campbell2002). Until recently, this species has incorrectly appeared in the literature as P. abrupta (Vadopalas et al., Reference Vadopalas, Pietsch and Friedman2010). They live burrowed aggregating in dense patches down to 1 m depth into soft sediments from the intertidal to 110 m (Jamison et al., Reference Jamison, Heggen and Lukes1984; Campbell et al., Reference Campbell, Yeung, Dovey and Zhang2004). Worldwide, geoduck clams have a temperate–subtemperate antitropical distribution with species generally circumscribed to major boundary currents. In the north-west Pacific, Panopea japonica Adams, 1850, the Japanese geoduck, is found in coastal waters of Korea, China and Japan (Bernard et al., Reference Bernard, Cai and Morton1993); whereas P. generosa, the Pacific geoduck, is found on the eastern coast from Alaska, USA to the Pacific coast of Baja California (Morris et al., Reference Morris, Abbott and Haderlie1980; Coan et al., Reference Coan, Scott and Bernard2000). A third species, the Cortes geoduck, Panopea globosa Dall, 1898, has been known hitherto to be endemic to the Gulf of California (GC), from San Felipe, Baja California, to Mazatlán, Sinaloa (Keen, Reference Keen1971; Hendrickx et al., Reference Hendrickx, Brusca and Findley2005; Arambula-Pujol et al., Reference Arambula-Pujol, García-Juárez, Alcántara-Razo and Aragón-Noriega2008). However, anecdotal evidence has suggested the presence of the Cortes geoduck on the Pacific coast of Baja California, at least in Bahía Magdalena (BM) on the Pacific coast of southern Baja California (E.A. Aragón-Noriega and L.E. Calderón-Aguilera, personal observations), a region potentially overlapping the range of the Pacific geoduck P. generosa. This situation has generated some confusion about the actual geographical boundaries of both north-east Pacific species (Rocha-Olivares et al., Reference Rocha-Olivares, Calderon-Aguilera, Aragon-Noriega, Saavedra-Sotelo and Moreno-Rivera2010). More recently, soon to be published (Leyva-Valencia et al., in press) and unpublished (Pérez-Valencia, Reference Pérez-Valencia2011; Leyva-Valencia, Reference Leyva-Valencia2012) reports have identified the presence of P. globosa in Bahía Magdalena through phenotypic and genetic analyses. However, the accurate range delimitation of P. generosa and P. globosa on the Pacific coast of Baja California remains elusive. Aside from the clear biological significance of clarifying the geographical range of both species, elucidating the causes of their structural and functional distinction is highly relevant, given their economic importance as target of a highly-valued burgeoning fishery in Mexico. For instance, official records indicate that in 2006 catches of both P. generosa and P. globosa, which are managed indistinctively, were in the order of 1300 t in Baja California, representing an income in excess of 82 million dollars (Diario Oficial de la Federación, 2012).
Our previous work has revealed significant morphometric and genetic differentiation of the Pacific and Cortes geoduck clams from northern Baja California and the upper Gulf of California (Rocha-Olivares et al., Reference Rocha-Olivares, Calderon-Aguilera, Aragon-Noriega, Saavedra-Sotelo and Moreno-Rivera2010). Here we extend our studies of Mexican geoducks both geographically and genetically. Using polymerase chain reaction–restriction fragments length polymorphisms (PCR-RFLPs) of nuclear ribosomal DNA (rDNA) and mitochondrial DNA (mtDNA) barcoding data, we ascertain the taxonomic identity of organisms collected in Bahía Magdalena. Our genetic results show unequivocally that these clams are P. globosa and, hence, help to establish its range extension northward to BM on the Pacific coast of southern Baja California and refute its status as a species endemic to the GC. On the other hand, morphometric data reveal that Pacific P. globosa differ from conspecifics from the Gulf and an ecological niche model indicates a very low probability that a species with the currently known distribution of P. globosa would successfully reside in Bahía Magdalena. These results lead us to suggest that although P. globosa is still a single evolutionary unit, the local deme on the Pacific coast of Baja California may be adapted to different conditions and thus is likely isolated. However, the combined effect of overlapping generations, the longevity of up to 60 years (Pérez-Valencia, Reference Pérez-Valencia2012; Pérez-Valencia & Aragón-Noriega, in press) and the long generation time of these clams may contribute to the lack of genetic differences discernible with our data, perhaps through a genetic storage effect (Ellner & Hairston, Reference Ellner and Hairston1994). This is the first comparative assessment of Mexican geoducks incorporating habitat analysis and our results are of consequence for the implementation of biologically sound and sustainable management and conservation measures for these commercially important clams.
MATERIALS AND METHODS
Geoduck clams Panopea sp. (N = 34) were sampled in February 2011 from the local fishery catch harvested inside Bahía Magdalena lagoon (24°39′10″N 112°01′47″W), on the Pacific coast of southern Baja California, Mexico. The right shell of each clam was measured (shell length (SL) and shell height (SH)) to the nearest mm as previously described (Rocha-Olivares et al., Reference Rocha-Olivares, Calderon-Aguilera, Aragon-Noriega, Saavedra-Sotelo and Moreno-Rivera2010). Siphon muscle tissue samples were dissected, preserved in 96% ethanol, and maintained at room temperature until laboratory analyses. For comparative purposes, we also analysed organisms (N = 20 for genetic analyses; N = 58 for morphometric analyses) of Panopea generosa from Santa Rosaliíta (28°40′00″N 114°15′57″W) and Islas Coronado (32°25′00″N 117°15′00″W) (Pacific coast, northern Baja California), and specimens (N = 19 for genetic analyses; N = 50 for morphometric analyses) of Panopea globosa from San Felipe, Baja California (31°01′30″N 114°50′27″W), and Puerto Peñasco, Sonora (31°19′05″N 113°32′12″W) (northern Gulf of California).
Genomic DNA was extracted using Chelex 100 (Bio-Rad, Hercules, CA). Based on our previous work (Rocha-Olivares et al., Reference Rocha-Olivares, Calderon-Aguilera, Aragon-Noriega, Saavedra-Sotelo and Moreno-Rivera2010), we used single-enzyme (Rsa I) PCR-RFLP of nuclear rDNA to genotype clams from Bahía Magdalena and reference organisms. This enzyme provides easily distinguishable diagnostic electrophoretic patterns for each species (P. generosa and P. globosa). PCR-RFLP patterns were resolved in 2% agarose gels stained with ethidium bromide (0.5 mg/ml). Further methodological details can be found in Rocha-Olivares et al. (Reference Rocha-Olivares, Calderon-Aguilera, Aragon-Noriega, Saavedra-Sotelo and Moreno-Rivera2010).
In addition, we PCR-amplified and sequenced the barcoding fragment of the mitochondrial cytochrome oxidase subunit I (COI) gene with the primers LCOI (5′- GGT CAA CAA ATC ATA AAG ATA TTG G-3′) and HCOI (5′- TAA ACT TCA GGG TGA CCA AAA AAT CA-3) (Folmer et al., Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994). PCR reactions (25µl) contained 0.2mM dNTPs, 1X PCR (Tris10mM, KCl 50mM, 1.5mM MgCl2), 0.4 µM each primer and 0.5U Taq (New England Biolabs, Ipswich, MA). Thermal cycling consisted of 2 minutes at 94°C, 35 cycles of 1 minute at 95°C, 1 minute at 35°C, and 1.5 minutes at 72°C, followed by a final extension of 7 minutes at 72°C. PCR products were purified using exoSAP-IT (USB, Cleveland, OH) before cycle-sequencing using Big Dye v.3.1 and electrophoresed in an automated Genetic Analyzer ABI-PRISM 3730 (Applied Biosystems, Foster City, CA) using the manufacturer's protocols.
DNA sequences were verified using Codon Code Aligner v3.7.1 (Codon Code Corporation, Dedham, MA, USA) before their alignment with the program MEGA v 5.05 (Tamura et al., Reference Tamura, Peterson, Peterson, Stecher, Nei and Kumar2011). The same program was used to reconstruct a neighbour-joining phylogenetic tree using the Kimura two parameter model of sequence evolution (K2P: Kimura, Reference Kimura1981) and branch support was obtained from 1000 bootstrap pseudo-replications. We performed an analysis of molecular variance with the COI DNA sequences of P. globosa using Arlequin v3.5.1.2 (Excoffier et al., Reference Excoffier, Laval and Schneider2005) to partition genetic variance within and between Bahía Magdalena and the Gulf of California and between pairs of geographical samples. Significance was adjusted for multiple testing using the sequential Bonferroni correction (Rice, Reference Rice1989). Morphometric variables (SL, SH and SH/SL ratio) were statistically and graphically compared among groups of geoducks (P. generosa from the Pacific coast, P. globosa from the Gulf of California and putative P. globosa from Bahía Magdalena).
We applied a maximum entropy (MaxEnt) species distribution model to map the probability of finding P. globosa off the west coast of the Baja California peninsula based on the characteristics of the habitat occupied by populations in the Gulf of California. MaxEnt is a machine learning method particularly suited to model species distributions based on presence-only data (Phillips et al., Reference Phillips, Anderson and Schapire2006). In statistical terms, the procedure uses environmental data (i.e. covariates) from the occurrence records (i.e. presence sites) and from a random sample from the geographical area being modelled (i.e. background landscape data) to estimate the ratio of the conditional probability density and the covariates at the presence sites and the unconditional (i.e. marginal) probability density of the covariates across the studied landscape. The model is optimized fitting a penalized maximum likelihood model to minimize the distance between conditional and unconditional densities and the ratio is used to estimate the conditional probability that the species is present given the environment (i.e. logarithmic output: see Dudik et al. (Reference Dudik, Phillips and Schapire2007) for machine learning and, Elith et al. (Reference Elith, Phillips, Hastie, Dudik, Chee and Yates2011) for statistical explanations of MaxEnt). In this case, distributional data of the species (N = 45 geo-referenced populations) came from the Invertebrate Macrofauna of the Gulf of California database (www.desertmuseum.org/center/seaofcortez/database.php), reports from the scientific literature (Aragón-Noriega et al., Reference Aragón-Noriega, Chavez-Villalba, Gribben, Alcantara-Razo, Maeda-Martínez, Arambula-Pujol, Garcia-Juarez and Maldonado-Amparo2007; Calderon-Aguilera et al., Reference Calderon-Aguilera, Aragón-Noriega, Reyes-Bonilla, Paniagua-Chávez, Romo-Curiel and Moreno-Rivera2010), and field data from the authors. Environmental background covariate data from the Gulf of California and the northern Mexican Pacific consisted of 11 variables (values at 1° lat long resolution), temperature (annual mean, maximum and minimum sea surface temperature; ± 1°C), chlorophyll-a concentration (mgm−3), total particulate organic carbon (μg.l−1) (source: MODIS-Aqua satellite, available at ftp.cdc.noaa.gov/datasets/noaa.oisst.v2/sst.mmean.mc), salinity (psu), nutrient concentrations (phosphate, silicate and nitrates; μM), dissolved oxygen (ml/l) (source: World Ocean Atlas available at www.nodc.noaa.gov), and bathymetry (m) (source: General Bathymetric Chart of the Oceans available at www.gebco.net). The rationale behind covariate choice for the model was to include a set of variables that would fulfil two conditions: (1) characterize the basic physical, chemical and ecological attributes of the environment; and (2) be synoptically available for the entire region. Admittedly, relevant variables for infaunal organisms such as substrate type and composition were not included mainly because of lack of data. The compromise between available and relevant data is a common concern and has been discussed by Tittensor et al. (Reference Tittensor, Mora, Jetz, Lotze, Ricard, Berghe and Worm2010) for the marine realm, among others.
The model was run with the software MaxEnt v3.3.1 (Phillips et al., Reference Phillips, Dudik and Schapire2004), using a maximum iteration value of 1000, default parameters (convergence threshold of 10−5 and automatic regularization of 10−4) and logistic output. The geographical output ranges from 0 to 1, and indicates the probability of occurrence of the species in a given region; as recommended in the literature (Franklin, Reference Franklin2009; Peterson et al., Reference Peterson, Soberón, Pearson, Anderson, Martínez-Meyer, Nakamura and Araújo2011) any value >0.5 is taken as adequate for the establishment of a reproductive population of the species. To assess the model discriminatory power, a statistical evaluation was performed with a threshold-independent receiver operating characteristic analysis (ROC: Phillips et al., Reference Phillips, Anderson and Schapire2006), using 75% of the occurrence locations for model training and 25% for testing (Franklin, Reference Franklin2009). Finally, variables were jackknifed to determine their contribution to the presence of P. globosa in the Gulf of California (Elith et al., Reference Elith, Phillips, Hastie, Dudik, Chee and Yates2011).
RESULTS AND DISCUSSION
All clams collected in Bahía Magdalena produced the same rDNA RsaI PCR-RFLP banding pattern. Comparisons with reference organisms of bona fide Panopea generosa from northern Baja California and Panopea globosa from the upper Gulf of California revealed that the RFLP pattern of Bahía Magdalena clams was characteristic of P. globosa and clearly distinguishable from that of P. generosa clams collected from Santa Rosaliíta (Figure 1). Further evidence of the identity of Bahía Magdalena clams as P. globosa was obtained from the mitochondrial COI. All five mtDNA haplotypes found in Bahía Magdalena clams grouped with those of P. globosa from the upper Gulf of California. A predominant haplotype was found in 66% of the clams and was shared among the three localities. The 13 P. globosa haplotypes were very divergent from the 11 P. generosa haplotypes from the Pacific coast of northern Baja California (uncorrected p-distance = 0.18). These results corroborate the high genetic divergence between these north-east Pacific Panopea species previously reported by Rocha-Olivares et al. (Reference Rocha-Olivares, Calderon-Aguilera, Aragon-Noriega, Saavedra-Sotelo and Moreno-Rivera2010) (Figure 2).

Fig. 1. Polymerase chain reaction–restriction fragment length polymorphism RsaI digestions of rDNA amplicons from Panopea sp. from Bahía Magdalena (lanes 3–19 and 22–27), P. generosa (lanes 1 and 2) and P. globosa (lanes 20 and 21) run on a 2% agarose gel. Lane M denotes size standard (50–10000 base pairs ladder).

Fig. 2. Neighbour-joining phylogram based on Kimura 2-parameter distances of mitochondrial cytochrome c oxidase subunit I and rDNA sequences from Mexican geoduck clams. Numbers refer to non-parametric bootstrap support values (>50%). The nuclear tree is modified from Rocha-Olivares et al. (Reference Rocha-Olivares, Calderon-Aguilera, Aragon-Noriega, Saavedra-Sotelo and Moreno-Rivera2010). Note: the basal branches leading to terminal haplotypes of the COL tree are broken. See text for levels of interspecific divergence.
Our genetic results clearly show that the clams we analysed from the harvest in Bahía Magdalena are unequivocally Panopea globosa. This evidence alone helps to extend the range of this species outside of the Gulf of California to this new latitude (24°39′N). Two recent studies provide concurring evidence for the genetic identity of Bahía Magdalena clams with bona fide P. globosa specimens from the Gulf of California: Pérez-Valencia (Reference Pérez-Valencia2011) using nuclear microsatellite loci specifically designed for Cortes geoducks and Leyva-Valencia (Reference Leyva-Valencia2012) using DNA sequences of mitochondrial COI and nuclear rDNA 18S and 28S genes. Additional sampling effort over a wider area should be devoted to establish the distributional limits of the Pacific and the Cortes geoducks on the Pacific coast of Baja California.
Clams from Bahía Magdalena were clearly distinguishable from the other Cortes geoducks from the Gulf of California and Pacific geoducks from the western coast of northern Baja California (Figure 3A). This phenotypic distinction was significant after correcting for size, as shown by the higher SH/SL ratio (t = −16, df = 66, P < 0.001, Figure 3B), which begs the question of whether this distinction has been a source of confusion regarding the identity of clams from this locality. Shell shape plasticity has been recognized as a potential source of taxonomic confusion in the identification of P. generosa and P. globosa from Mexico (Leyva-Valencia et al., in press). Interestingly, our rather crude morphometric assessment of these clamshells concurs with more elaborate geometric analyses of Leyva-Valencia et al. (in press), involving peripheral and inner shell landmarks, in revealing the phenotypic distinction between Pacific and gulf specimens of P. globosa. On the other hand, DNA sequence variation at the mitochondrial COI gene revealed no evidence of significant genetic differentiation between Gulf of California and Pacific Cortes geoducks. A marginally non-significant 4.7% (ΦST = 0.047, P = 0.06) of the genetic variance was partitioned between the geographical samples and 95.3% was found within them. These results concur with those of Pérez-Valencia (Reference Pérez-Valencia2011) who found a non-significant ΦST between clams from the Pacific (Bahía Magdalena) and the Gulf of California (San Felipe, Puerto Peñasco and Guaymas), using two polymorphic microsatellite loci. However, variation in those nuclear loci revealed small (<4%) but significant pairwise differentiation between Bahía Magdalena and San Felipe and between Bahía Magdalena and Puerto Peñasco, whereas our COI data revealed comparable but not statistically significant levels of differentiation (Table 1). Both studies coincide in finding null levels of genetic differentiation among Gulf of California localities in contrast to a higher percentage of the genetic variance partitioned between gulf and Pacific samples in pairwise comparisons, lack of significance adjusted for multiple testing notwithstanding. This is not unexpected given the contrasting environments between the regions.

Fig. 3. Morphometric analyses of Mexican geoducks: (A) bivariate plot of shell measurements; (B) box-and-whisker plots of shell height/shell length ratios of north-east Pacific Panopea clams (box: median bound by 25 and 75 percentiles; whiskers: 10 and 90 percentiles; black circles: outliers). Comparative data of P. generosa and P. globosa are from Rocha-Olivares et al. (Reference Rocha-Olivares, Calderon-Aguilera, Aragon-Noriega, Saavedra-Sotelo and Moreno-Rivera2010).
Table 1. Analysis of molecular variance pairwise ΦST (below diagonal) and corresponding P values (above diagonal) based on mitochondrial cytochrome c oxidase subunit I sequences of Panopea globosa (BM, Bahía Magdalena; PP, Puerto Peñasco; SF, San Felipe).

BM versus PP not significant after Bonferroni correction.
Bahía Magdalena is recognized as a very distinct habitat from the Gulf of California. As a biogeographical transition zone between temperate and tropical waters (Garcia-Rodriguez & Perez-Enriquez, Reference Garcia-Rodriguez and Perez-Enriquez2006), it shows great spatial and temporal oceanographic variability, being influenced by the California Currents system flowing southwardly featuring low temperature and salinity water rich in oxygen and nutrients, and the Coastal Mexican Current flowing northwardly with oligotrophic, more saline and warmer water (Lavin et al., Reference Lavin, Fiedler, Amador, Ballance, Farber-Lorda and Mestas-Nunez2006). The shallow, protected waters of the Bahía Magdalena–Almejas complex, are also typically warmer, more quiescent, and more productive than adjacent offshore waters (Bizarro, Reference Bizarro2008). On the other hand, habitat conditions in the Gulf of California are dominated by tidal currents, upwelling, high insulation, and mesoscale oceanographic features such as cyclonic eddies (Alvarez-Borrego, Reference Alvarez-Borrego and Brusca2010). These features enhance productivity that surpasses the one found in Bahía Magdalena. For instance, chlorophyll-a concentration in the gulf can exceed 20 mg.m−3 in winter, and the annual mean reaches over 8 mg.m−3 (Millán-Núñez & Lara-Lara, Reference Millán-Núñez, Lara-Lara, González-Farías and de la Rosa-Vélez1995; Alvarez-Borrego, Reference Alvarez-Borrego and Brusca2010). In contrast, peak chlorophyll-a concentration in Bahía Magdalena reaches only 15 mg.m−3 in spring and the annual average is below 5 mg.m−3 (Cervantes-Duarte et al., Reference Cervantes-Duarte, López-López and González-Rodríguez2007). These differences were also reflected in the data used in the MaxEnt model (Table 2).
Table 2. Mean (± SE) values of environmental covariates used in the MaxEnt model characteristic for Bahía Magdalena (BM) in the Pacific coast of Baja California Sur and for the regions occupied by Panopea globosa in the Gulf of California (GC).

SE, standard error; MaxEnt, maximum entropy; SST, sea surface temperature; Chl-a, chlorophyll-a; POC, particulate organic carbon; psu, practical salinity units; DO, dissolved oxygen; GEBCO, General Bathymetric Chart of the Oceans; † 1° × 1° quadrants include deep water, means do not reflect the depth at which clams were collected.
The MaxEnt model was statistically very robust (area under curve = 0.995), and indicated that the main variables influencing the distribution of P. globosa are chlorophyll-a (38% of explained variance; presence at >2.5 mg.m−3), nitrates (31% of variance; presence at 1.5–3.0 µM) and bathymetry (18.2% of variance; presence at depths <40 m); the remaining variables were of lesser importance. According to the modelled niche distribution of the Cortes geoduck, the most likely habitat for the species may be found in relatively shallow and productive areas restricted to the Gulf of California, with particularly high suitability in the upper Gulf and at the southern coast of the State of Sonora, even though the extent of potential habitat supporting reproductive populations is more widespread (i.e. green areas in Figure 4). The probability of occurrence of the species in BM is predicted to be low (<0.1), mostly because of variables relating to differences in productivity such as chlorophyll-a and nutrient concentrations, as stated above (Table 2). For instance the mean (±SE) nitrate concentration in the lagoon was almost 18 times lower (0.155 ± 0.011 µM) than the one found in the regions occupied by P. globosa in the gulf (2.762 ± 0.539 µM).
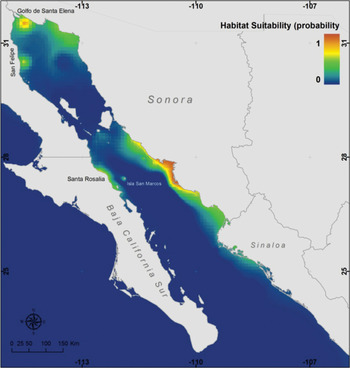
Fig. 4. Predicted habitat suitability for Panopea globosa using a maximum entropy model (MaxEnt).
These MaxEnt results are at odds with the presence of P. globosa in BM, since the model suggests that given the present conditions it would not be able to colonize the western coast of the peninsula (Figure 4). Reconciliation of these findings requires looking at the historical biogeography of the region. It is well known that temperate and sub-temperate coastal species have experienced range expansions and episodes of radiation at the time of Miocene and Pleistocene oceanographic changes (e.g. peaks of upwelling) as well as during Pleistocene and Holocene climate changes (Jacobs et al., Reference Jacobs, Haney and Louie2004), which may have favoured the presence of Panopea on both coasts of the Baja California peninsula as supported by palaeontological records (Dall, Reference Dall1918). The poleward expansion of subtropical waters during subsequent interglacial warming during the Holocene may have acted as an ecological barrier helping to isolate eastern and western Baja Californian coastal populations, producing disjunct distributions also present in other taxa (Terry et al., Reference Terry, Bucciarelli and Bernardi2000; Stepien et al., Reference Stepien, Rosenblatt and Bargmeyer2001; Bernardi et al., Reference Bernardi, Findley and Rocha-Olivares2003; Rocha-Olivares et al., Reference Rocha-Olivares, Leal-Navarro, Kimbrell, Lynn and Vetter2003). This hypothesized isolation should be conducive to the genetic and morphological distinction of clams, particularly in the face of contrasting selective pressures.
The genetic differentiation of geoducks in Bahía Magdalena from conspecifics in the Gulf of California needs further assessment. The genetic data available on this species, including those of Pérez-Valencia (Reference Pérez-Valencia2011), point to a shallow differentiation between clams from these regions. However, we argue that these data sets have insufficient statistical power to reliably settle this question both in terms of number of loci analysed and in terms of levels of polymorphism. It is also possible that owing to their late maturity and protracted life cycle (Aragón-Noriega et al., Reference Aragón-Noriega, Chavez-Villalba, Gribben, Alcantara-Razo, Maeda-Martínez, Arambula-Pujol, Garcia-Juarez and Maldonado-Amparo2007; Calderon-Aguilera et al., Reference Calderon-Aguilera, Aragón-Noriega, Reyes-Bonilla, Paniagua-Chávez, Romo-Curiel and Moreno-Rivera2010) favouring a genetic storage effect (Ellner & Hairston, Reference Ellner and Hairston1994), clams in BM have not diverged enough to be genetically distinguishable from the other conspecifics living inside the GC. Nevertheless, we find that the concurrence of our morphometric, genetic and ecological results with those of others (Pérez-Valencia, Reference Pérez-Valencia2011; Leyva-Valencia, Reference Leyva-Valencia2012; Leyva-Valencia et al., in press) pertaining to the differentiation of Cortes clams from the Pacific and the gulf strongly suggests high levels of local adaptation implying that there may not be a frequent (or continuous) and successful larval exchange between BM and the GC populations, and that the former is isolated from the rest.
As a corollary, our results extend the range of P. globosa outside of their initial distributional range inside the Gulf of California, hence contradicting its status as a species endemic to the Gulf of California as previously stated (Keen, Reference Keen1971; Hendrickx et al., Reference Hendrickx, Brusca and Findley2005; Arambula-Pujol et al., Reference Arambula-Pujol, García-Juárez, Alcántara-Razo and Aragón-Noriega2008; Rocha-Olivares et al., Reference Rocha-Olivares, Calderon-Aguilera, Aragon-Noriega, Saavedra-Sotelo and Moreno-Rivera2010), and also shed light on the ecological and evolutionary consequences of inhabiting Bahía Magdalena, a presumably disjunct and marginal habitat relative to the one occupied by conspecifics in the Gulf of California.
ACKNOWLEDGEMENTS
We thank Cristina Hernández, for help in sample collection, Doña Lili, for allowing sampling of her catch, fishers of Puerto San Carlos for facilitating field operations, and Sara Melo, David Petatán and Cecilia Mozqueda for helping with data analyses. We also thank two anonymous referees for their constructive remarks. This research was supported by CICESE grant 625112 to A.R.O.






