1. Introduction
The Ordovician–Silurian boundary interval was a remarkable period of climatic, environmental and sea-level changes. Rapid deglaciation of the Earth's Southern Hemisphere brought a dramatic rise in global sea level, weakened oceanic circulation and enhanced stratification of water masses. Profound environmental changes are documented by faunal and sedimentary turnover associated with conspicuous geochemical signatures. The Ordovician–Silurian boundary is marked by an anoxic event recorded worldwide (Melchin et al. Reference Melchin, Mitchell, Holmden and Štorch2013). In the majority of European sections, postglacial upper Hirnantian mudstones and shales, occasionally with the trilobite Mucronaspis and Hirnantia shelly brachiopod fauna, are sharply overlain by anoxic black shales with abundant graptolites. This scheme, however, varies depending on the palaeogeography and location relative to the shoreline and the local rate of sedimentation. In the offshore successions of Baltica, represented by sections and drill cores in Bornholm (Bjerreskov, Reference Bjerreskov1975; Koren’ & Bjerreskov, Reference Koren’ and Bjerreskov1997) and southern Sweden (Bergström et al. Reference Bergström, Huff, Korenʼ, Larsson, Ahlberg and Kolata1999; Koren’, Ahlberg & Nielsen, Reference Korenʼ, Ahlberg, Nielsen, Ortega and Aceñolaza2003), the lowermost part of the black shale succession has yielded low-diversity to moderately diverse graptolite faunas of the upper Hirnantian Metabolograptus persculptus Biozone. Graptolites of the persculptus Biozone have also been reported from northeastern Poland (Masiak, Podhałańska & Stempień-Sałek, Reference Masiak, Podhałańska and Stempień-Sałek2003; Podhałańska, Reference Podhalańska, Ortega and Aceñolaza2003; Trela et al. Reference Trela, Podhałańska, Smolarek and Marynowski2016). In the Avalonian Welsh Basin, the uppermost Ordovician strata contain several thin levels (Blackett et al. Reference Blackett, Page, Zalasiewicz, Williams, Rickards and Davies2009) of black shale or mudstone with abundant M. persculptus (Elles & Wood, Reference Elles and Wood1907) and M. parvulus (Lapworth, Reference Lapworth1900). Moderately diverse graptolite assemblages from the persculptus Biozone, including the earliest monograptid Atavograptus ceryx (Rickards & Hutt, Reference Rickards and Hutt1970), have been described from the Lake District, northern England (Rickards, Reference Rickards1970; Hutt, Reference Hutt1974). At the global boundary stratotype section and point (GSSP) for the base of the Silurian System in Dob's Linn, southern Scotland (Williams, Reference Williams1983; Melchin, Reference Melchin, Ortega and Aceñolaza2003; Fan, Melchin & Williams, Reference Fan, Melchin, Williams, Sheehan and Servais2005), continuous black shale sedimentation commenced in the persculptus Biozone. The graptolite assemblage recorded by Fan, Melchin & Williams (Reference Fan, Melchin, Williams, Sheehan and Servais2005) in the lowermost part of this black shale succession included several species (Normalograptus minor (Huang, Reference Huang1982), N. mirnyensis (Obut, Sobolevskaya & Nikolaev, Reference Obut, Sobolevskaya and Nikolaev1967), N. praetamariscus (Li, Reference Li1984), N. rhizinus (Li & Yang in Nanjing Institute of Geology and Mineral Resources, 1983), Korenograptus laciniosus (Churkin & Carter, Reference Churkin and Carter1970), N. skeliphrus (Korenʼ & Melchin, Reference Korenʼ and Melchin2000), N. ugurensis (Korenʼ & Melchin, Reference Korenʼ and Melchin2000) and Neodiplograptus shanchongensis (Li, Reference Li1984)) previously recognized in the low-latitude palaeobiogeographical province of China, Central Asia, Siberia and northern North America outlined by Melchin (Reference Melchin1989).
In peri-Gondwanan Europe, the graptolite record across the Ordovician–Silurian boundary is rather different, less abundant and less complete. Metabolograptus persculptus, locally accompanied by tentatively determined normalograptid rhabdosomes, Mucronaspis trilobites and Hirnantia brachiopod faunas, has been collected from Hirnantian mudstones of the Prague Synform in the Czech Republic (Nová Ves, Pankrác, Praha-Nové Butovice and Praha-Řepy sections; Štorch, Reference Štorch1986; Štorch & Loydell, Reference Štorch and Loydell1996), Saxony (Frankenberg section; Jaeger, Reference Jaeger1977), the Austrian Southern Alps (Cellon and Feistritsgraben sections; Jaeger, Havlíček & Schönlaub, Reference Jaeger, Havlíček and Schönlaub1975; Štorch & Schönlaub, Reference Štorch and Schönlaub2012) and western Bulgaria (Sachanski, Reference Sachanski1993; Lakova & Sachanski, Reference Lakova and Sachanski2004). Hirnantian graptolites are either missing (most sections in the Czech Republic, Thuringia, Sardinia, Brittany, Montagne Noire, Portugal and Spain), badly preserved (Bulgaria, Austrian Carnic Alps) or confined to a distinct level separated by a barren interval from the basal Silurian ascensus Biozone (Řepy, Nové Butovice, Nová Ves and Běchovice sections in the Czech Republic, Frankenberg section in Saxony). The lowermost samples of Silurian black shales exhibit rich graptolite faunas dominated by Akidograptus ascensus Davies, Reference Davies1929, Nd. lanceolatus Štorch & Serpagli, Reference Štorch and Serpagli1993 and normalograptids in all coeval sections from Portugal (Piçarra et al. Reference Piçarra, Štorch, Gutiérrez-Marco and Oliveira1995) and Spain (Jaeger & Robardet, Reference Jaeger and Robardet1979; Gutiérrez-Marco & Robardet, Reference Gutiérrez-Marco and Robardet1991; Roqué, Reference Roqué1999), through Brittany (Piçarra et al. Reference Piçarra, Robardet, Oliveira, Paris and Lardeux2009), Montagne Noire (Štorch & Feist, Reference Štorch and Feist2008), Sardinia (Štorch & Serpagli, Reference Štorch and Serpagli1993), Thuringia and Saxony (Schauer, Reference Schauer1971), northern Bavaria (Stein, Reference Stein1965), central Bohemia (Štorch, Reference Štorch1996), and eastern Serbia (Krstič, Maslarevič & Sudar, Reference Krstič, Maslarevič and Sudar2005) to western Bulgaria (Lakova & Sachanski, Reference Lakova and Sachanski2004). The biostratigraphy and graptolite assemblages of the A. ascensus–Parakidograptus acuminatus Biozone of peri-Gondwanan Europe were reviewed by Štorch (Reference Štorch1996). A rather different assemblage, including exotic graptolite species, was described by Štorch & Schönlaub (Reference Štorch and Schönlaub2012) from the Waterfall Section near Zollnersee in the Austrian Southern Alps. Undeterminable normalograptids occur throughout much of the latter black, cherty succession, while determinable specimens including Par. ʻacuminatusʼ (Nicholson, Reference Nicholson1867) and K. bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974) (= Rickardsograptus bifurcus (Ye, Reference Ye1978)) are confined to a rather thin interval 92–102 cm above the top of the thick uppermost bed of quartzite.
Some doubts about the high-resolution biostratigraphy of the Ordovician–Silurian boundary faunas of Europe, and about biostratigraphical and palaeobiogeographical interpretations of the Austrian fauna in particular, can be resolved as a result of the recent discovery of a richly fossiliferous Ordovician–Silurian boundary section near Estana village in the south-central Pyrenees of Spain (Fig. 1). A preliminary report on the Estana section was presented to the Geological Society of Spain by Roqué Bernal, Štorch & Gutiérrez-Marco (Reference Roqué Bernal, Štorch and Gutiérrez-Marco2017), who identified six graptolite species and provided an approximate placement of the Ordovician–Silurian boundary above the Bar Quartzite. Further study of the section and graptolite material has revealed that the M. persculptus of the latter authors belongs in M. parvulus which is a closely similar species, common in the upper persculptus Biozone and uncommon in the ascensus–acuminatus Biozone. The stratigraphically highest records are from Jordan, from strata tentatively referred to the lower Cystograptus vesiculosus Biozone by Loydell (Reference Loydell2007).

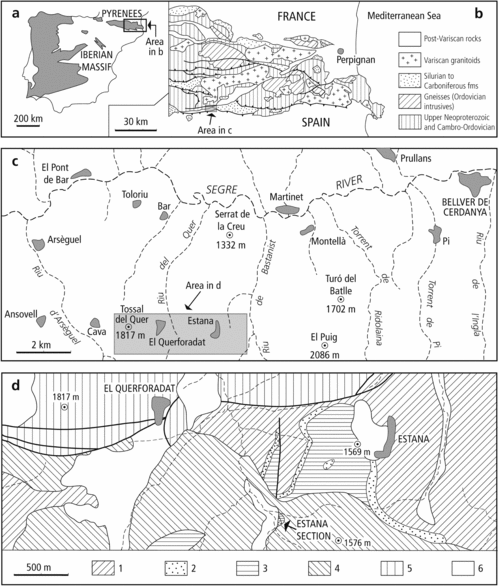
Figure 1. (a) Location map with insets showing Variscan outcrop areas (grey) in the Iberian Peninsula. (b) Geological sketch map of eastern part of the Axial Zone of the Pyrenees. (c) Geographical map of the studied area south of the Segre River valley. (d) Geological sketch map showing position of the Estana section, modified from the Atles Geològic de Catalunya (Institut Geològic de Catalunya and Institut Cartogràfic de Catalunya, 2010). 1, Ansovell Formation, dark shales with interbedded fine-grained sandstones; 2, Bar Formation, grey medium-grained sandstones; 3, black graptolitic shales; 4, dacitic and andesitic lavas of Carboniferous–Permian age; 5, undifferentiated Mesozoic rocks; 6, Quaternary cover.
The Estana section, discovered by JRB and described in detail in this paper (Fig. 2), is unique in a European context in having uniform, uninterrupted and somewhat rapid black shale sedimentation across the Ordovician–Silurian boundary interval, combined with a rich, continuous and moderately well-preserved graptolite fossil record. The sedimentation rate is considered to be comparable to that of deep-water black shale Ordovician–Silurian boundary sections at Dob's Linn, Scotland (Williams, Reference Williams1983) or the Röstanga core in southern Sweden (Korenʼ, Ahlberg & Nielsen, Reference Korenʼ, Ahlberg, Nielsen, Ortega and Aceñolaza2003). Black shales overlying the quartzites of the Bar Formation contain a high-diversity graptolite fauna with two new and several age-diagnostic taxa along with the biostratigraphical marker species M. parvulus and cosmopolitan biozonal-indices A. ascensus and Par. acuminatus.

Figure 2. Estana section: lithologies and graptolite fossil record.
2. Methods and materials
The section, repeatedly studied by the authors since 2013, was systematically sampled for its graptolite fossil record, lithologies and organic carbon geochemistry. Each 20 cm thick interval of the section was studied bed-by-bed and graptolites were collected from a rock volume of c. 0.03 m3 per sample. Some intervals were divided into two 10 cm thick parts and resampled. Graptolites are preserved as flattened impressions with periderm partly pyritized and, for the most part, covered by pale mineral overgrowths that make rhabdosomes clearly visible against the black shale. Common preservation of long nemata, long and complex virgellar structures as well as complete synrhabdosomes indicates a quiet depositional environment without significant post-mortem transport. Bedding planes covered by aligned rhabdosomes are rare. The effects of tectonic strain have changed proportions and measured parameters of some specimens by 5–10%. Illustrated and measured specimens are housed in the Museo Geominero, Instituto Geológico y Minero de España, Madrid and bear an MGM designation.
The section was sampled for total organic carbon (TOC) content and organic carbon isotope composition at the same regular intervals (20 cm and 10 cm) from the base of the black shale succession to the top of the measured section. Hand specimens were cut and rock powder was prepared from a few grams of a fresh sample. A few milligrams of rock powder were taken for TOC and isotope analyses. Before analyses, rock powders were decarbonatized then washed and dried. TOC content was analysed in the Geological Institute AS CR, Prague. Carbon isotope analyses were performed by GeoZentrum Nordbayern, Erlangen with a Flash EA 2000 elemental analyser connected online to ThermoFinnigan Delta V Plus mass spectrometer. Carbon isotope values are related to V-PDB. Accuracy and reproducibility of the analyses were checked by replicate analyses of laboratory standard CH4N2O calibrated to international standards USGS 40 and 41. Reproducibility was better than ±0.1‰ (1σ).
3. Geological setting
The studied section is located on the southern edge of the central Pyrenees Axial Zone, west of the La Cerdanya Neogene basin (Fig. 1). It is exposed adjacent to the old mountain track from Estana to Querforadat, on the left (southern) bank of the Segre River valley in the province of Lleida, NE Spain. The main graptolite-bearing section is within an isolated outcrop of uppermost Ordovician sandstones and lowermost Silurian black shales surrounded by weathered Carboniferous–Permian dacitic lavas and Holocene sediments. The locality lies on the hillslope above the right bank of the Riu del Quer (= Quer stream), c. 1200 m SW of the village of Estana, and 400 m south of fossil locality no. 68 of Dégardin (Reference Dégardin1988, fig. 203), from where this author listed 19 Aeronian graptolite species. Geographical coordinates for the base of the Silurian (base of the ascensus–acuminatus Biozone) in the Estana section are 42°18′51.65″N and 1°39′2.68″W.
The highest Ordovician succession in the central Pyrenees is very similar to those of the Canigó Massif in the axial part of the eastern Pyrenees. In both areas, above calcareous shales and limestones of the Estana Formation (up to 150 m thick) with Kralodvorian (late Katian–Ka3-4) brachiopods and conodonts (Hartevelt, Reference Hartevelt1970; Gil-Peña et al. Reference Gil-Peña, Barnolas, Villas, Sanz-López and Vera2004; Colmenar, Reference Colmenar2015), there are poorly bedded dark slates of the Ansovell Formation (20–300 m thick), followed by the Bar Quartzite (0–20 m of dark-grey, medium-grained sandstone) which is overlain by black shales with graptolites. The assumed record of Late Ordovician brachiopods near the base of the Bar Quartzite (Hartevelt, Reference Hartevelt1970), and of middle Rhuddanian – Aeronian graptolites in the overlying black shales (Roussel, Reference Roussel1904; Dalloni, Reference Dalloni1930; Boissevain, Reference Boissevain1934; Dégardin, Reference Dégardin1988, Reference Dégardin1990), has led to different interpretations about the placement of the Ordovician–Silurian boundary. For some authors it should be at the base of the quartzite unit (Schmidt, Reference Schmidt1931; Boissevain, Reference Boissevain1934), for others at the top (Hartevelt, Reference Hartevelt1970; Dégardin et al. Reference Dégardin, Alonso, Bessière, Bodin, Bouquet, Brula, Centène, Duran, Garces-Cocchio, García-López, García-Sansegundo, Guérangé, Julivert, Laumonier, Losantos, Palau, Paris, Pouit, Raymond, Richard, Sanz, Truyols-Massoni, Villas, Barnolas and Chiron1996; Casas, Reference Casas2010) or at an undetermined level within this unit (Gil-Peña et al. Reference Gil-Peña, Barnolas, Sanz-López, García-Sansegundo and Palau2001, Reference Gil-Peña, Barnolas, Villas, Sanz-López and Vera2004; Gil-Peña & Barnolas, Reference Gil-Peña and Barnolas2007; Puddu & Casas, Reference Puddu, Casas, Gutiérrez-Marco, Rábano and García-Bellido2011; Casas & Palacios, Reference Casas and Palacios2012; Margalef et al. Reference Margalef, Castiñeiras, Casas, Navidad, Liesa, Linnemann, Hofmann and Gärtner2016; Casas, Puddu & Álvaro, Reference Casas, Puddu and Álvaro2017; Puddu, Casas & Álvaro, Reference Puddu, Casas and Álvaro2017). Brachiopods found by Hartevelt (Reference Hartevelt1970) in the Bar Quartzite and reviewed by E. Villas (in Gil-Peña et al. Reference Gil-Peña, Barnolas, Villas, Sanz-López and Vera2004) represent reworked specimens derived from both the Estana and Cava formations, the latter of late Berounian (c. Ka2) age. Gil-Peña et al. (Reference Gil-Peña, Barnolas, Sanz-López, García-Sansegundo and Palau2001) referred to an important erosive unconformity at the top of the Ansovell Formation, probably related to the sea-level fall resulting from the Hirnantian glaciation, which is locally overlain by a discontinuous conglomerate containing pebbles of some older Upper Ordovician stratigraphic units. The Bar Quartzite, overlying the palaeorelief infilled by conglomerates or the Ansovell Formation, is interpreted as a transgressive unit attributed to the sea-level rise at the end of the glacial event. Recent dating of detrital zircons from the Bar Quartzite of the Rabassa dome (Pyrenean Axial Zone; Margalef et al. Reference Margalef, Castiñeiras, Casas, Navidad, Liesa, Linnemann, Hofmann and Gärtner2016) yielded a youngest peak age of 443 Ma, close to the Ordovician–Silurian boundary estimated at 443.8±1.5 Ma according to the latest edition (2017/02) of the International Chronostratigraphic Chart. Detrital zircons from Ordovician sediments of Pyrenees and SW Sardinia studied by Margalef et al. (Reference Margalef, Castiñeiras, Casas, Navidad, Liesa, Linnemann, Hofmann and Gärtner2016) indicate that the two terranes shared the same source area. Both terranes were situated near the northern Gondwana margin, adjacent to the Arabian–Nubian Shield between the Arabian–Nubian drainage system in the east and present-day Libya and Algeria in the west.
The discovery of the new, complete and richly fossiliferous Estana section has revealed the true position of the Ordovician–Silurian boundary in the south-central Pyrenees (Roqué Bernal, Štorch & Gutiérrez-Marco, Reference Roqué Bernal, Štorch and Gutiérrez-Marco2017). In the Riu de Quer area (Estana section), the Bar Quartzite shows a sharp and well-defined upper conformable contact with the black graptolitic shales that have been unanimously considered to be Silurian by all preceeding authors. Our study extends over the first 4.1 m of the unnamed formation, known in the literature as the ʻlower graptolitic shalesʼ, ʻblack shales with graptolitesʼ or ʻblack graphitic shalesʼ. In these strata we found a specific graptolite assemblage with an abundant occurrence of M. parvulus and other taxa indicating the upper Hirnantian persculptus Biozone. The uninterrupted black shale succession continued into the lower Rhuddanian ascensus–acuminatus Biozone (Fig. 2). The Ordovician–Silurian boundary, marked by the first appearance datum (FAD) of A. ascensus, is located 2.2 m above the top of the Bar Quartzite.
4. Graptolite record, biostratigraphy and correlation
Silty-micaceous black shales conformably overlying the thin-bedded quartzites of the Bar Formation are rich in relatively diverse graptolites (Fig. 3) tentatively assigned to the upper part of the persculptus Biozone. True M. persculptus is missing from the Estana section but the closely related M. parvulus, which is a common species of the upper persculptus Biozone (see Melchin, McCracken & Oliff, Reference Melchin, McCracken and Oliff1991; Loydell et al. Reference Loydell, Mallett, Mikulic, Kluessendorf and Norby2002; Blackett et al. Reference Blackett, Page, Zalasiewicz, Williams, Rickards and Davies2009), is abundant in association with N. ajjeri (Legrand, Reference Legrand1977), N. minor, N. angustus (Perner, Reference Perner1895) and N. baridaensis sp. nov. The parvulus Biozone, correlatable with the upper part of the persculptus Biozone, is defined in the section as the interval between the FAD of M. parvulus and the FAD of A. ascensus.

Figure 3. Graptolite fauna of the parvulus Biozone. (a–c) Normalograptus lubricus (Chen & Lin, Reference Chen and Lin1978): (a) MGM-8100-O, sample EST 6–7; (b) MGM-8101-O, EST 6–7; (c) MGM-8102-O, EST 6–7. (d, g–i) Metabolograptus parvulus (Lapworth, Reference Lapworth1900): (d) MGM-8074-O, EST 4; (g) MGM-8075-O, EST 8; (h) MGM-8076-O, EST 2; (i) MGM-8077-O; EST 7. (e, f, l) Normalograptus angustus (Perner, Reference Perner1895): (e) MGM-8093-O, EST 3a; (f) MGM-8094-O, EST 7; (l) MGM-8095-O, EST 8. (j, k, m) Normalograptus baridaensis sp. nov.: (j) holotype, MGM-8096-O, EST 2b; (k) MGM-8099-O, EST 2b–3a; (m) MGM-8098-O, EST 2b. (n, q) Normalograptus rhizinus (Li & Yang in Nanjing Institute of Geology and Mineral Resources, 1983): (n) MGM-8106-O, EST 10b; (q) MGM-8107-O, EST 10b. (o, p) Normalograptus aff. rhizinus (Li & Yang in Nanjing Institute of Geology and Mineral Resources, 1983): (o) MGM-8104-O, EST 7; (p) MGM-8105-O, EST 7. (r–u) Normalograptus minor (Huang, Reference Huang1982): (r) MGM-8088-O, EST 3; (s) MGM-8089-O, EST 7; (t) MGM-8091-O, EST 4a; (u) MGM-8090-O, EST 5. (v, w) Normalograptus ajjeri (Legrand, Reference Legrand1977): (v) MGM-8082-O, EST 7; (w) MGM-8083-O, EST 9. All specimen in the same magnification, black bar represents 1 mm.
Normalograptus minor was first described from the uppermost Hirnantian strata of Tibet, China and later recorded from the same level in the Yangtze region of China (Chen et al. Reference Chen, Fan, Melchin and Mitchell2005) and from the Dob's Linn GSSP section in Scotland (Fan, Melchin & Williams, Reference Fan, Melchin, Williams, Sheehan and Servais2005). Koren’, Ahlberg & Nielsen (Reference Korenʼ, Ahlberg, Nielsen, Ortega and Aceñolaza2003) reported N. minor from their pre-ascensus avitus Fauna of southern Sweden. A single specimen from the lower ascensus–acuminatus Biozone of southwestern Sardinia, reported as Normalograptus sp. B by Štorch & Serpagli (Reference Štorch and Serpagli1993), extends its range into the lowermost Rhuddanian strata.The species is particularly common 0.7–1.0 m above the top of the sandstone unit. Species richness further increased in the middle part of the local range of M. parvulus. Normalograptus minor disappears 1.9 m above the quartzite while N. lubricus (Chen & Lin, Reference Chen and Lin1978), N. aff. rhizinus (Li & Yang in Nanjing Institute of Geology and Mineral Resources, 1983) and Glyptograptus dufkai Štorch, Reference Štorch1992 joined the assemblage. Higher up the section, N. ajjeri becomes abundant whereas most other taxa made their last occurrences. Synrhabdosomes of M. parvulus (Fig. 4a) are common, along with a few synrhabdosomes of N. ajjeri, at several levels from 0.3 to 1.9 m above the base of the black shale succession (Fig. 2). Relatively common uniserial abnormalities in the astogenetic development of biserial rhabdosomes (Fig. 4b–e) suggest some environmental stress and/or phylogenetic experimentation.

Figure 4. (a) Radially arrayed synrhabdosome of M. parvulus (Lapworth, Reference Lapworth1900), MGM-8081-O, sample EST 3. (b) Normalograptus ajjeri (Legrand, Reference Legrand1977) with long uniserial portion, MGM-8084-O, EST 8–9 (1.6–1.8 m above the top of the Bar Formation). (c–e) Abnormal normalograptid rhabdosomes distally forked into two uniserial stipes: (c) MGM-8085-O, EST 8–9; (d) MGM-8086-O, EST 8–9; (e) MGM-8087-O, EST 8–9. Black and white scale bars represent 5 mm.
Normalograptus lubricus was reported by Chen & Lin (Reference Chen and Lin1978) from a combined ascensus–bicaudatus Biozone of northern Guizhou Province, China in association with A. ascensus. Climacograptus sp. (cf. Cl. miserabilis), figured by Williams (Reference Williams1983) from the lower acuminatus Biozone (= ascensus–acuminatus Biozone as used here) of Dob's Linn, Scotland, also belongs to N. lubricus. In Arctic Canada, Melchin, McCracken & Oliff (Reference Melchin, McCracken and Oliff1991) distinguished a specific madernii–lubricus Subzone in the lower part of the acuminatus Biozone despite the absence of akidograptid graptolites in this lower subzone. In Uzbekistan, Korenʼ & Melchin (Reference Korenʼ and Melchin2000) recognized a N. lubricus Subzone in the lower part of their ascensus Biozone, again below the lowest occurrence of A. ascensus. In Saudi Arabia, Williams et al. (Reference Williams, Zalasiewicz, Boukhamsin and Cesari2016) recorded N. lubricus in association with N. parvulus and N. bifurcatus Loydell, Reference Loydell2007 in a distinct lubricus Biozone recognized by them below the lowest akidograptid occurrences. It is obvious that, in some regions at least, the appearance of N. lubricus preceded the formal base of the ascensus Biozone defined by the FAD of the nominal taxon. In a strict biostratigraphical sense, N. lubricus therefore occurs in the uppermost Hirnantian strata in the Estana section, 0.3–1.0 m below the lowest A. ascensus and below the FADs of K. bifurcus and N. rhizinus. The graptolite fauna of the lower part of the Estana section can be recognized as a distinct, presumably time-specific, assemblage (Fig. 5, Assemblage 2) tentatively correlated with the upper persculptus Biozone assemblage in Scotland (Dob's Linn) and avitus Fauna reported by Koren’, Ahlberg & Nielsen (2003) from the interval between the highest M. persculptus and lowest A. ascensus in southern Sweden (Röstanga core). Assemblage 2 (see Fig. 6 for details) comprises four cosmopolitan species: N. minor, thought to be confined in a low-latitude realm; G. dufkai of north Gondwanan distribution; and two ʻlocal speciesʼ recorded for the first time. This assemblage has not been found in the Ordovician–Silurian boundary sections elsewhere in peri-Gondwanan Europe.

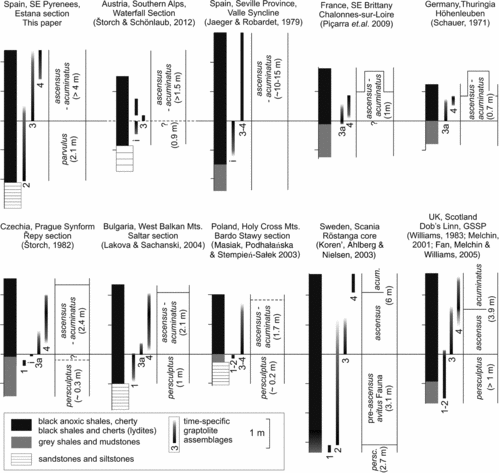
Figure 5. Correlation of the most complete graptolite-bearing Ordovician–Silurian boundary sections of Europe. Time-specific graptolite assemblages discussed in this paper: 1, upper Hirnantian fauna including M. persculptus; 2, graptolite assemblage of the parvulus (upper persculptus) Biozone; lowermost ascensus–acuminatus Biozone characterized by M. parvulus, N. minor, N. rhizinus and Nd. shanchongensis, which are accompanied either by N. avitus s.s., N. avitus of Williams, Reference Williams1983 and K. laciniosus in low-latitude sections (avitus Fauna of Korenʼ, Ahlberg & Nielsen, Reference Korenʼ, Ahlberg, Nielsen, Ortega and Aceñolaza2003) or by N. lubricus and K. bifurcus in the peri-Gondwanan realm; 3, graptolite assemblage of the lowermost ascensus–acuminatus Biozone marked by the appearance of A. ascensus and Par. praematurus in association with either taxa inherited from the previous assemblage or with Nd. lanceolatus (Assemblage 3a); 4, classical assemblage of the lower–middle ascensus–acuminatus Biozone characterized by the co-occurrence of A. ascensus and Par. acuminatus along with Nd. lanceolatus, N. trifilis and Cyst. ancestralis; i, graptolite assemblage limited to indeterminable or long-ranging normalograptids.

Figure 6. Time-specific graptolite assemblages of the parvulus (upper persculptus) and lower–middle ascensus–acuminatus biozones recognized in the Estana section. The species are grouped according to their palaeogeographical distribution: cosmopolitan species, species previously known only from low-latitude palaeoplates, species confined to peri-Gondwanan Europe (i.e. northwestern peri-Gondwana) and ʻlocal speciesʼ recorded solely from the Estana Section.
Normalograptus rhizinus, with its gradually tapering rhabdosome and long virgella widening into a small distal vane, K. bifurcus, with a robust bifurcating virgella, and the lowest N. mirnyensis appeared in a c. 0.3 m thick interval (Fig. 2, samples EST 10, 10a, 10b, 11a) between the last M. parvulus and first A. ascensus. The morphologically and stratigraphically significant N. rhizinus and K. bifurcus do not occur in the avitus Fauna of Koren’, Ahlberg & Nielsen (Reference Korenʼ, Ahlberg, Nielsen, Ortega and Aceñolaza2003). Records of N. rhizinus have been confined to the upper persculptus Biozone in both China (Chen et al. Reference Chen, Fan, Melchin and Mitchell2005) and Scotland (Fan, Melchin & Williams, Reference Fan, Melchin, Williams, Sheehan and Servais2005). Korenograptus bifurcus, which is common in the lowermost Silurian deposits of Sichuan Province, China, was reported by Štorch & Schönlaub (Reference Štorch and Schönlaub2012) from the ascensus–acuminatus Biozone of the Waterfall Section in the Austrian Southern Alps.
The base of the Silurian System is marked in the Estana Section by the lowest occurrence of A. ascensus 2.2 m above the top of the sandstone unit (Fig. 2, sample EST 11b). It coincides, almost precisely, with the stratigraphically lowest Par. praematurus (Davies, Reference Davies1929) and Nd. shanchongensis. The species richness of the graptolite fauna considerably increased as illustrated by Figures 7 and 8. The black shale is markedly less silty at this level than in the parvulus Biozone. A new assemblage (Figs 5, 6; Assemblage 3) comprised cosmopolitan A. ascensus, Par. praematurus, early Par. acuminatus, Nd. shanchongensis, N. mirnyensis and some other, long-ranging normalograptids along with four species widespread in low-latitude palaeoplates of Asia and North America (K. bicaudatus (Chen & Lin, Reference Chen and Lin1978), K. lanpherei (Churkin & Carter, Reference Churkin and Carter1970), K. bifurcus and N. rhizinus) and the newly decribed N. ednae sp. nov. Peri-Gondwanan elements represent G. dufkai and N. crassus Štorch & Feist, Reference Štorch and Feist2008. This assemblage is similar to that described by Štorch & Schönlaub (Reference Štorch and Schönlaub2012) from Austria in having K. bifurcus together with Par. praematurus (identified as Par. acuminatus in that paper), early forms of Par. acuminatus, N. ajjeri and some Glyptograptus. Nd. lanceolatus and cosmopolitan N. trifilis (Manck, Reference Manck1923) are absent from both the Austrian section and in the lower part of the ascensus–acuminatus Biozone in the Estana section. The joint occurrence of K. bicaudatus and A. ascensus allows for correlation of this level with the lowermost Silurian ascensus–bicaudatus Biozone defined in China by Chen & Lin (Reference Chen and Lin1978).

Figure 7. Graptolite fauna of the ascensus–acuminatus Biozone. (a) Normalograptus ajjeri (Legrand, Reference Legrand1977): MGM-1703-S, sample EST 13b. (b) Normalograptus rhizinus (Li & Yang in Nanjing Institute of Geology and Mineral Resources, 1983): MGM-1719-S, EST 16. (c, k) Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974): (c) MGM-1714-S, EST 12; (k) MGM-1715-S, EST 12. (d, aa) Parakidograptus acuminatus (Nicholson, Reference Nicholson1867), early form: (d) MGM-1756-S, EST 20; (aa) MGM-1757-S, EST 16. (e, i) Parakidograptus praematurus (Davies, Reference Davies1929): (e) MGM-1730-S, EST 13a; (i) MGM-1731-S, EST 13a. (f) Normalograptus mirnyensis (Obut, Sobolevskaya &Nikolaev, 1967): MGM-1721-S, EST 16. (g, q, bb) Normalograptus ednae sp. nov.: (g) MGM-1760-S, EST 16; (q) holotype, MGM-1761-S, EST 16; (bb) MGM-1762-S, EST 16. (h) Korenograptus lanpherei (Churkin & Carter, Reference Churkin and Carter1970): MGM-1749-S, EST 16. (j) Normalograptus angustus (Perner, Reference Perner1895): MGM-1708-S, EST 13b. (l, m) Neodiplograptus lanceolatus Štorch & Serpagli, Reference Štorch and Serpagli1993: (l) MGM-1773-S, EST 19; (m) MGM-1774-S, EST 19. (n, r, s) Glyptograptus dufkai Štorch, Reference Štorch1992: (n) MGM-1709-S, EST 16; (r) MGM-1710-S, EST 16; (s) MGM-1711-S, EST 12. (o, y, z) Akidograptus ascensus Davies, Reference Davies1929: (o) MGM-1738-S, EST 16; (y) MGM-1739-S, EST 15; (z) MGM-1740-S, EST 16. (p, x) Neodiplograptus shanchongensis (Li, Reference Li1984): (p) MGM-1722-S, EST 16; (x) MGM-1723-S, EST 16. (t, u) Normalograptus crassus Štorch & Feist, Reference Štorch and Feist2008: (t) MGM-1754-S, EST 15; (u) MGM-1755-S, EST 14. (v, w) Neodiplograptus sp.: (v) MGM-1758-S, EST 16; (w) MGM-1759-S, EST 15. All specimen in the same magnification, black bar represents 1 mm.

Figure 8. Graptolite fauna of the ascensus–acuminatus Biozone, continued. (a, ?i, r) Korenograptus bicaudatus (Chen & Lin, Reference Chen and Lin1978): (a) proximal part of 52 mm long specimen, MGM-1745-S, sample EST 12; (?i) rhabdosome in profile with slightly arcuate branches of the bifurcated virgella, MGM-1746-S, EST 12; (r) MGM-1747-S, EST 12. (b, p) Parakidograptus praematurus (Davies, Reference Davies1929): (b) MGM-1732-S, EST 11b; (p) MGM-1733-S, EST 11b. (c, m) Normalograptus trifilis (Manck, Reference Manck1923): (c) MGM-1781-S, EST 20; (m) MGM-1782-S, EST 20. (d) Rickardsograptus sp.: MGM-1771-S, EST 16. (e) Hirsutograptus sp.: MGM-1786-S, EST 20. (f, l) Korenograptus lanpherei (Churkin & Carter, Reference Churkin and Carter1970): (f) MGM-1750-S, EST 14; (l) MGM-1751-S, EST 12. (g, ?j) Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974): (g) MGM-1716-S, EST 13a; (?j) MGM-1717-S, EST 12. (h, o) Metabolograptus? aff. parvulus (Lapworth, Reference Lapworth1900): (h) MGM-1767-S, EST 16; (o) MGM-1768-S, EST 16. (k) Normalograptus ajjeri (Legrand, Reference Legrand1977): MGM-1704-S, EST 11b. (n) Cystograptus ancestralis Štorch, Reference Štorch1985: MGM-1778-S, EST 20. (q) Normalograptus ednae sp. nov.: MGM-1763-S, EST 16. (s) Normalograptus medius (Törnquist, Reference Törnquist1897): MGM-1772-S, EST 16. All specimen in the same magnification, black bar represents 1 mm.
About 3 m above the base of the black shale succession (Fig. 2, 2.7–3.5 m, samples EST 14–17) the graptolite fauna further diversified. Parakidograptus praematurus was gradually replaced by an early morphotype of Par. acuminatus, distinguished from Par. praematurus by its later point of origin of theca l¹ (th11), generally more protracted proximal part of the rhabdosome and more acuminate metathecae with triangular outline and flowing genicula. Korenograptus bifurcus with its substantial forked virgella is replaced by K. lanpherei with its simple, long and thinner virgella. This interval, which is dominated by abundant N. ajjeri and characterized by N. crassus and apparently short-ranging N. ednae sp. nov., with its short and robust triple basal spines (virgella and two lateral spines), also incorporates the highest occurrences of G. dufkai, N. rhizinus, N. mirnyensis and Nd. shanchongensis, rare Metabolograptus? aff. parvulus (Lapworth, Reference Lapworth1900) and single specimens of N. medius (Törnquist, Reference Törnquist1897) and Rickardsograptus sp.
A graptolite assemblage dominated by Nd. lanceolatus and A. ascensus associated with N. trifilis, N. ajjeri, N. angustus, Cyst. ancestralis Štorch, Reference Štorch1985 and Par. acuminatus (Assemblage 4 in Figs 5, 6) appears nearly 4 m above the base of the black shale succession in the Estana section (samples EST 18–20, Fig. 2). Early representatives of Par. acuminatus are difficult to distinguish from its ancestor Par. praematurus which made its lowest occurrence much lower, at about the same level as the earliest A. ascensus. The co-occurrence of the three species has led us to use the combined ascensus–acuminatus Biozone in the sense of Štorch (Reference Štorch1996) rather than separate ascensus and acuminatus biozones. The ascensus–acuminatus Biozone is defined as the interval between the FAD of A. ascensus and the FAD of Cyst. vesiculosus (Nicholson, Reference Nicholson1868), the index species of the succeeding biozone which is also present in a higher part of the Estana section.
The graptolite assemblage from the uppermost samples of the Estana section has been reported from almost every graptolite-bearing Ordovician–Silurian boundary section of peri-Gondwanan Europe by Štorch (Reference Štorch1996). It is typically found in the lower and lower–middle parts of the combined ascensus–acuminatus Biozone in other European and Spanish sections (Štorch, Reference Štorch1996) and was recently recorded in Saudi Arabia (Williams et al. Reference Williams, Zalasiewicz, Boukhamsin and Cesari2016). Faunal elements from Estana that are new to peri-Gondwanan Europe are few, but K. bicaudatus, K. lanpherei and Hirsutograptus sp. enhance correlation of this assemblage with the middle–upper acuminatus Biozone or the middle–upper part of a combined ascensus–acuminatus Biozone in Asia and northern North America. Korenograptus bicaudatus has been employed as a lowermost Silurian biozonal index taxon in the ascensus–bicaudatus Biozone in China (Chen & Lin, Reference Chen and Lin1978), K. lanpherei is confined to the acuminatus Biozone in Alaska and northern Canada (Loxton, Reference Loxton2017) and Hirsutograptus Koren’ & Rickards, Reference Koren’ and Rickards1996 is another taxon of particular biostratigraphical significance outside Europe. A H. sinitzini Subzone is distinguished in the upper part of the acuminatus Biozone in Arctic Canada (Melchin, McCracken & Oliff, Reference Melchin, McCracken and Oliff1991) and in the uppermost part of the ascensus–acuminatus Biozone in Uzbekistan (Korenʼ & Melchin, Reference Korenʼ and Melchin2000) where its lowest occurrence coincides with the FAD of typical Par. acuminatus. Other hirsutograptids appear at the same stratigraphical level also in China (Chen et al. Reference Chen, Rong, Mitchell, Harper, Fan, Zhan, Zhang, Li and Wang2000), Tien Shan (Chaletskaya, Reference Chaletskaya and Markovsky1960), Russian northern Siberia (Obut, Sobolevskaya & Nikolaev, Reference Obut, Sobolevskaya and Nikolaev1967; Gogin et al. Reference Gogin, Koren’, Pegel’ and Sobolevskaya1997) and the southern Urals of western Kazakhstan (Koren’ & Rickards, Reference Koren’ and Rickards1996). Melchin (Reference Melchin2001) identified Hirsutograptus in the middle acuminatus Biozone at Dob's Linn, southern Scotland.
5. TOC and organic carbon geochemistry
Little fluctuating but generally increasing TOC values of 1.8–2.8 wt% in the parvulus Biozone and 2.3–3.2 wt% in the lower ascensus–acuminatus Biozone (Fig. 9) conform with the rather uniform black shale lithology and may also indicate stable, slightly increasing palaeoproductivity and stable or slightly decreasing sedimentation rate. The interpreted relatively high rate of black shale sedimentation in the Estana section would have the potential to reveal short-term carbon isotope excursions across the Hirnantian–Rhuddanian (Ordovician–Silurian) boundary interval much better than the more condensed, and possibly incomplete, sedimentary record preserved in many other sections of peri-Gondwanan Europe. The δ 13Corg values, varying from –29.84‰ to –30.60‰ with a general shift to lower values (Fig. 9), are consistent with values and trends recorded from other fully anoxic Ordovician–Silurian boundary sections (Underwood et al. Reference Underwood, Crowley, Marshall and Brenchley1997; Fan, Peng & Melchin, Reference Fan, Peng and Melchin2009; Melchin et al. Reference Melchin, Mitchell, Holmden and Štorch2013). These values suggest that black shale sedimentation commenced in the Estana section well after the end of the Hirnantian positive carbon isotope excursion (HICE), which is consistent with the biostratigraphical dating of the lowermost black shale samples to the upper part of the persculptus Biozone. The minor decline in the δ 13Corg values below the base of the ascensus–acuminatus Biozone (samples EST 8–11a) resembles the minor negative excursion recorded at the same stratigraphic level in the Dob's Linn (Underwood et al. Reference Underwood, Crowley, Marshall and Brenchley1997) and Wangjiawan (Fan, Peng & Melchin, Reference Fan, Peng and Melchin2009) sections. However, significant negative correlation between TOC and δ 13Corg using three correlation coefficients (Pearson's r = –0.62, Spearman's rs = –0.49 and Kendall's τ = –0.40) indicates partial post-diagenetic opening of the carbon isotope system, possibly related to Carboniferous–Permian dacitic lavas exposed nearby. The δ 13Corg data must be considered with reservation in the Estana section, despite the fairly normal values matching other coeval sections.

Figure 9. δ 13Corg isotopic record and TOC in the Estana section. See Figure 2 for lithology explanations and abbreviations.
6. Discussion
The Ordovician–Silurian boundary strata exposed in the Estana section not only yield a diverse and unique graptolite fauna with several species previously unknown from Europe, but have also revealed some deviations in stratigraphical occurrences of the graptolite taxa.
Since no M. persculptus s.s. has been found in the Estana section, the exact biostratigraphical correlation of Assemblage 2 (Fig. 5) recovered from the black shale interval below the lowest occurrence of akidograptids is based upon the co-occurrence of other biostratigraphically useful species. The closely similar but smaller M. parvulus, common in the uppermost part of the persculptus Biozone all around the world, continues into the ascensus–acuminatus Biozone in some sections (Loydell, Reference Loydell2007; Blackett et al. Reference Blackett, Page, Zalasiewicz, Williams, Rickards and Davies2009). It is abundant in the lower part of the Estana section. It is accompanied by both long-ranging normalograptids and N. minor which has been described from the upper Hirnantian part (persculptus Biozone) of the Lungmachi Formation (Yangtze Platform, China). Normalograptus rhizinus, which also occurs in the upper persculptus Biozone in China and southern Scotland, made its lowest occurrence two samples below the lowest occurrence of A. ascensus in the Estana section and ranges well into the lower part of the ascensus–acuminatus Biozone. Normalograptus lubricus, described by Chen & Lin (Reference Chen and Lin1978) from the lowermost ascensus–bicaudatus Biozone of China and recorded from the same level in southern Scotland (Fan, Melchin & Williams, Reference Fan, Melchin, Williams, Sheehan and Servais2005) and Arctic Canada (Melchin, McCracken & Oliff, Reference Melchin, McCracken and Oliff1991), gives its name to a subzone in Uzbekistan (Koren’ & Melchin, Reference Korenʼ and Melchin2000) and a biozone in Saudi Arabia (Williams et al. Reference Williams, Zalasiewicz, Boukhamsin and Cesari2016). It is confined to the uppermost part of the newly established parvulus Biozone, still below the lowest occurrence of akidograptids. We suppose that at least the upper part of the parvulus Biozone, c. 1.1–2.2 m above the base of the black shale succession, can be correlated with the lubricus Biozone of Koren’ & Melchin (Reference Korenʼ and Melchin2000) and Williams et al. (Reference Williams, Zalasiewicz, Boukhamsin and Cesari2016) as well as with the rather different graptolite fauna of the pre-ascensus interval recognized by Koren’, Ahlberg & Nielsen (2003) in Sweden. Considering that this nearly cosmopolitan fauna has not yet been recorded in any other section in peri-Gondwanan Europe, we could speculate that such a unique graptolite assemblage spread under specific environmental conditions enabling immigration of distinct elements of low-latitude faunas (sensu Melchin, Reference Melchin1989) in the course of post-glacial climate amelioration and major rise of sea level. Indeed, specific and unusual environmental conditions in the Ordovician pre-ascensus part of the Estana section could be inferred from the abundant occurrence of graptolite synrhabdosomes (Fig. 4a, see also Gutiérrez-Marco & Lenz Reference Gutiérrez-Marco and Lenz1998 for discussion) and from abnormal astogeny with uniserial development in biserial taxa (Metabolograptus and Normalograptus, Fig. 4b–e). Such abnormalities had to be more widespread, although not ubiquitous in the Ordovician–Silurian boundary interval as indicated by synrhabdosomes of Normalograptus ex gr. normalis and M. persculptus s.l. from the persculptus Biozone of Mauritania (Underwood, Deynoux & Ghienne, Reference Underwood, Deynoux and Ghienne1998) and uni-biserial rhabdosomes recorded in a notably similar interval at Dob's Linn, the Ordovician–Silurian boundary GSSP (Williams, Reference Williams1983, text-figs 3b, 7d, 8h; Muir, Reference Muir2011, fig. 1). It is worth noting that the present author (PŠ) recorded numerous synrhabdosomes of Nd. africanus (Legrand) in the lower–middle Rhuddanian strata of Mauritania, which indicates that synrhabdosomes were perhaps linked to a regional environment rather than being entirely specific to a particular time.
Mature septate biserial rhabdosomes split distally into two long uniserial stipes, each furnished with a nema, suggests that abnormal colonies were able to grow until full maturity, typical of other specimens of the respective species. Specimens from the Estana section support Muirʼs (Reference Muir2011) assumption that fine-tuning of rhabdosome hydrodynamics was not essential for further growth and survival of the graptolite colony. Common uniserial growth in the distal part of biserial graptolite taxa due to post-mortem breakage during transport of the rhabdosomes, suggested by Williams (Reference Williams1983), can be excluded in the Estana section because of the common co-occurrence of unbroken long and complex virgellar structures, nemata and complex synrhabdosomes.
The common occurrence of abnormal specimens in the uppermost Hirnantian black shales of the Estana section may be considered as evidence of stressful environmental conditions. Synrhabdosomes have been interpreted by Gutiérrez-Marco & Lenz (Reference Gutiérrez-Marco and Lenz1998) as temporary structures, perhaps formed in relatively restricted water masses with low food supply. ʻTeratological eventsʼ, recognized by Delabroye et al. (Reference Delabroye, Munnecke, Servais, Vandenbroucke and Vecoli2012), Munnecke et al. (Reference Munnecke, Delabroye, Servais, Vandenbroucke and Vecoli2012) and Vandenbroucke et al. (Reference Vandenbroucke, Emsbo, Munnecke, Nuns, Duponchel, Lepot, Quijada, Paris, Servais and Kiessling2015) in other planktic groups near the Ordovician–Silurian boundary, have also been related to diverse causes that generated a high level of environmental stress such as upwelling of anoxic/dysoxic waters, heavy metal pollution or acidification due to explosive volcanism. Better understanding of causal relations among environmental changes near the Ordovician–Silurian boundary and their biotic response will require further consideration; both abnormal graptolite colonies and synrhabdosomes appeared in the Estana section in the same stratigraphical interval that witnessed the incipient adaptive radiation of the graptolite faunas after the latest Katian – early Hirnantian mass extinction event, including the origination of the uniserial monograptid colony.
In the Estana section, the relatively low-diversity graptolite assemblage of the uppermost samples of the upper Hirnantian parvulus Biozone was succeeded by an assemblage comprising N. rhizinus, N. mirnyensis, N. crassus, K. bicaudatus, K. bifurcus, K. lanpherei, Nd. shanchongensis, M.? aff. parvulus, A. ascensus, Par. praematurus and long-ranging normalograptids, later joined by N. ednae and an early form of Par. acuminatus. This high-diversity assemblage, which is confined to the lower part of the ascensus–acuminatus Biozone in the Estana section (Figs 5, 6; Assemblage 3), is unique in a European context, with the partial exception of the Austrian Southern Alps (having K. bifurcus and Par. praematurus) and of southern Scotland having Par. praematurus, N. mirnyensis, N. rhizinus and Nd. shanchongensis. A few elements of this assemblage associated with Nd. lanceolatus (Fig. 5, Assemblage 3a) ranged up to a slightly higher stratigraphical level in the ascensus–acuminatus Biozone. Some species have also been reported from Montagne Noire (N. crassus; Štorch & Feist, Reference Štorch and Feist2008), Sardinia (N. crassus, determined as N. medius by Štorch & Serpagli, Reference Štorch and Serpagli1993), Bohemia (G. dufkai; Štorch Reference Štorch1996), Bulgaria (Par. praematurus; Lakova & Sachanski, Reference Lakova and Sachanski2004) and Saudi Arabia (N. crassus; determined as N. medius by Williams et al. Reference Williams, Zalasiewicz, Boukhamsin and Cesari2016, fig. 11C).
Abundant occurrences of N. trifilis, Nd. lanceolatus, Cyst. ancestralis, A. ascensus and Par. acuminatus in the uppermost part of the Estana succession (samples EST 18 – EST 20) belong to Assemblage 4 (Figs 5, 6) which is known from the lower–middle part of the combined ascensus–acuminatus Biozone in the majority of lower Rhuddanian black shale sections in peri-Gondwanan Europe (see Štorch, Reference Štorch1996 and Štorch & Feist, Reference Štorch and Feist2008 for summary), north-eastern Morocco (PŠ, personal observation), Jordan (Loydell, Reference Loydell2007), Saudi Arabia (Williams et al. Reference Williams, Zalasiewicz, Boukhamsin and Cesari2016), Scotland (Williams, Reference Williams1983 and M.J. Melchin, pers. comm.), eastern Poland (Masiak, Podhałańska & Stempień-Sałek, Reference Masiak, Podhałańska and Stempień-Sałek2003) and southern Sweden (Korenʼ, Ahlberg & Nielsen, Reference Korenʼ, Ahlberg, Nielsen, Ortega and Aceñolaza2003). In this interval, Par. praematurus has been entirely replaced by Par. acuminatus. The varied, not always favourable mode of preservation and co-occurrence of the two parakidograptid morphotypes is the reason that we prefer a combined ascensus–acuminatus Biozone instead of separate, but imperfectly distinguished ascensus and acuminatus biozones.
The appearance of typical ʻperi-Gondwananʼ faunal assemblages relatively high in the Estana black shale succession and the absence of the earlier O–S boundary assemblages with N. lubricus, N. rhizinus, N. aff. rhizinus, N. minor, N. baridaensis, N. ednae, K. bifurcus, K. bicaudatus, K. lanpherei and Nd. shanchongensis in other stratigraphically relevant black shale sections of northern peri-Gondwana may be alternatively explained by either immigration of exotic graptolite taxa under specific local environmental conditions or widespread omission of corresponding graptolitic black shale strata in many peri-Gondwanan sections. The absence of these taxa is in notable conjunction with condensed, discontinuous or oxic sedimentation, whereas their presence, although usually incomplete, is linked with less condensed and presumably continuous black shale sedimentation that commenced well before the appearance of the first akidograptids.
The faunal affinity of a single interval with well-determinable graptolite taxa in the Austrian Southern Alps to the lower part of the ascensus–acuminatus Biozone in the Estana section matches this assumption, although poor graptolite preservation in other levels of the Austrian section precludes full evidence. Determinable graptolites found in siliceous black shale c. 1 m above the top of the underlying quartzite bed in Austria included K. bifurcus, Par. praematurus and simple normalograptids, whereas Nd. lanceolatus, N. trifilis, Cyst. ancestralis and other elements of Assemblage 4 are missing. Black shales and lydites below this fossiliferous level may be correlative with the upper persculptus (parvulus) Biozone of the Estana section; black shales above the fossiliferous level could be assigned to the middle part of the ascensus–acuminatus Biozone, although direct faunal evidence is not preserved. Condensed sedimentation, disconformities and an incomplete fossil record observed in many other sections can be explained by sediment starvation associated with a rapid and culminating post-glacial rise in sea-level (see Melchin et al. Reference Melchin, Mitchell, Holmden and Štorch2013 for review). In the Prague Synform of central Bohemia, for instance, condensed sedimentation or non-deposition at the interface between upper-Hirnantian mudstones and lowest Silurian black shales has been presumed by Štorch (2006), prior to the indirect evidence provided by the Estana section. In central Bohemia the uppermost Hirnantian oxic mudstones with Hirnantia fauna and M. persculptus, Assemblage 1 in Figure 5, are topped by a firmground (Štorch, 2006) overlain by black shale with abundant Nd. lanceolatus, A. ascensus, N. angustus, G. dufkai (Assemblage 3a), 0.1–0.2 m higher joined by N. trifilis, Cyst. ancestralis, Nd. parajanus (Štorch, Reference Štorch1983) and Par. acuminatus (Assemblage 4). A black shale succession from the parvulus and lowermost ascensus–acuminatus biozones correlative with those of the Spanish Estana and Austrian Waterfall sections is missing from the Prague Synform.
The record of a single specimen of Hirsutograptus, a genus previously unknown from Europe (other than Scotland), within a graptolite assemblage characteristic of the lower–middle ascensus–acuminatus Biozone in northern Gondwana, suggests that this level may be tentatively correlatable with the H. sinitzini Subzone recognized high in the ascensus–acuminatus Biozone of Asia and northern North America and with the lower part of the acuminatus Biozone of Yukon Territory, Canada (sensu Loxton, Reference Loxton2017). The poorly preserved Hirsutograptus sp. from the Estana section is similar to relatively small multispinous hirsutograptids including H. comantis (Chaletskaya, Reference Chaletskaya and Markovsky1960), H. sinitzini (Chaletskaya, Reference Chaletskaya and Markovsky1960) and H. villosus Korenʼ & Rickards, Reference Koren’ and Rickards1996.
7. Conclusions
The Ordovician–Silurian boundary section near Estana village in the south-central Pyrenees of Spain exposes richly fossiliferous black shales little affected by faulting or tectonic strain and sheds new light on high-resolution biostratigraphy and the global correlation of the boundary interval worldwide. Increasing and little fluctuating TOC content (1.8–3.2 wt%), δ 13Corg values decreasing with some fluctuation from –29.84‰ to –30.60‰, and gradual replacement of graptolite assemblages conforms with presumably uninterrupted anoxic sedimentation within a single graptolite biofacies.
We have recognized and defined a distinct M. parvulus Biozone correlatable with the uppermost part of M. persculptus Biozone, including the so-called pre-ascensus interval with N. avitus Fauna recognized by Korenʼ, Ahlberg & Nielsen (Reference Korenʼ, Ahlberg, Nielsen, Ortega and Aceñolaza2003) below the FADs of A. ascensus and Par. praematurus.
A combined A. ascensus–Par. acuminatus Biozone has been adopted in the lowermost part of the Silurian succession instead of separate ascensus and acuminatus biozones because the two index taxa overlap for a large part of their stratigraphical ranges. In addition, Par. praematurus, which is difficult to distinguish when unfavourably preserved from early specimens assigned to Par. acuminatus, has its lowest occurrence in the same sample as A. ascensus.
High-diversity graptolite faunas of the parvulus and lower–middle ascensus–acuminatus biozones comprise 27 species, including two new taxa (N. baridaensis sp. nov. and N. ednae sp. nov.) and several taxa of particular biostratigraphical significance in Siberia, central Asia, China and northern North America, that is, in lower latitudes according to the early Llandovery Earth palaeogeographical reconstruction by Torsvik & Cocks (Reference Torsvik and Cocks2017, fig. 7.1a).
A succession of four graptolite assemblages is recognized in the Ordovician–Silurian boundary interval comprising persculptus (here parvulus) and lower–middle ascensus–acuminatus biozones (Fig. 5). The lowest assemblage marked by the occurrence of M. persculptus (Assemblage 1), is missing from the black shale Estana section. Only the fourth assemblage, recorded in the uppermost part of the studied succession, is fully represented in all relevant sections in peri-Gondwanan Europe. The high number of graptolite taxa recorded in the Estana section for the first time in peri-Gondwanan Europe (in particular N. minor, N. lubricus, N. mirnyensis, N. rhizinus, N. aff. rhizinus, M. parvulus, Hirsutograptus sp., K. bicaudatus, K. bifurcus, K. lanpherei, Nd. shanchongensis and Par. praematurus) and the presence of two new species (N. ednae, N. baridaensis) might be explained by the local occurrence of specific environmental conditions which, along with indigenous speciation, allowed for the immigration of several truly exotic taxa. Although the black shale facies is closely similar to those elsewhere in peri-Gondwanan Europe, somewhat specific conditions during latest Hirnantian time may be inferred from the presence of abnormal split and uniserial rhabdosomes and the common occurrence of synrhabdosomes.
However, many of the ʻexoticʼ graptolite taxa previously known only from low-latitude palaeoplates are actually cosmopolitan; their general absence from other sections of peri-Gondwanan Europe leads us to assume that the lower 2–3 m of the black shale succession studied in the Estana section, with their highly distinctive graptolite assemblages of the parvulus and lowermost ascensus–acuminatus biozones (Assemblage 2 and fully developed Assemblage 3), represent strata which are either oxic and barren of graptolites, condensed or even missing elsewhere from peri-Gondwanan Europe.
8. Palaeontological notes
Normalograptus lubricus (Chen & Lin, Reference Chen and Lin1978)
Specimens assigned to N. lubricus (Figs 3a–c) are confined to a narrow stratigraphical interval below the lowest occurrence of akidograptids in the Estana section. The proximal end of the rhabdosome is furnished with a rather short virgella and one or two basal spines growing from the upwards turning point of th11. The maximum width of 0.95–1.05 mm, attained at the fifth thecal pair, is less than that measured by Korenʼ & Melchin (Reference Korenʼ and Melchin2000) in specimens from Uzbekistan but still greater than the maximum width of N. lubricus from Dobʼs Linn (see Cl. sp. (aff. Cl. miserabilis) of Williams, Reference Williams1983). The number of basal spines documented by other authors (2–4 including virgella) depends on burial and flattening of the rhabdosome; many specimens are preserved in sub-scalariform view and spines are projected in various directions, not simply parallel to the rhabdosome profile view.
Normalograptus minor (Huang, Reference Huang1982)
Abundant specimens of N. minor (Figs 3r–u, 10e, p) attain a maximum length of 34 mm, excluding the long and robust, 0.1–0.18 mm wide nema and 3–5 mm long virgella which splits into multiple, up to 1.3 mm long, densely packed root-like branches. The proximal end is sub-rounded, slightly asymmetrical and 0.9–1.15 mm wide at the level of th1¹–th1² apertures. The rhabdosome gradually widens until the maximum width of c. 1.5–1.6 mm is attained by the 9th–10th thecal pair. Extreme values range from 1.3 mm to 1.95 mm, depending on rhabdosome orientation relative to the tectonic strain. Thecae are strongly geniculated; narrow thecal excavations occupy about one-third of the rhabdosome width. Supragenicular walls are slightly inclined in most specimens. Two thecae repeat distance (2TRD) is 1.5–1.75 mm at th2 and 1.8–2.0 mm at th10. Distal thecae number 10–11 in 10 mm. Relatively short specimens in the same bedding plane assemblages exhibit lesser maximum width (1.1 mm), slightly more densely packed thecae (12 in 10 mm) and shorter virgellae, although terminated by an identical root-like furcation. The type specimens of N. minor from Tibet are rather small, with 6 thecae in 5 mm of rhabdosome length and a maximum width of 1.2–1.3 mm (Huang, Reference Huang1982). Their virgella forked 3.5–5.5 mm below the sicular aperture into several isolated branches. All specimens are characterized by a long and forked virgella, long nema and slight and gradual widening of the rhabdosome. The scalariform specimen of Normalograptus sp. B, recorded by Štorch & Serpagli (Reference Štorch and Serpagli1993, text-fig. 7P) from the ascensus–acuminatus Biozone of south-western Sardinia, may also be tentatively assigned to N. minor. Climacograptus radicatus Chen & Lin, Reference Chen and Lin1978 and Diplograptus coremus Chen & Lin, Reference Chen and Lin1978, other species with long multifurcated virgella which occur in Ordovician–Silurian boundary beds, can be readily distinguished from N. minor by their robust rhabdosomes, widening from 0.6–0.7 mm at th1 to a distal maximum of 2.2–2.4 mm, bluntly geniculated thecae with markedly inclined supragenicular walls, wider thecal spacing with a 2TRD of 2.3–2.4 mm, and virgella branched into few, relatively long branches (see also Chen & Lin, Reference Chen and Lin1978). Chen et al. (Reference Chen, Fan, Melchin and Mitchell2005) regarded Cl. radicatus as a junior synonym of Nd. coremus (Chen & Lin, Reference Chen and Lin1978).
Normalograptus aff. rhizinus (Li & Yang in Nanjing Institute of Geology and Mineral Resources, 1983)
Normalograptus aff. rhizinus (Figs 3o, p, 10r) matches N. rhizinus in having a rhabdosome widening gradually from 0.75–0.8 mm at the first thecal pair to a maximum of 1.55–1.7 mm which is attained by the 10th–14th thecal pair. The long virgella widens distally into a vane-like structure. However, the virgella of N. aff. rhizinus is slender and the vane structure, when preserved, is confined to its distal part. Distal thecae are long and widely spaced in N. aff. rhizinus with a 2TRD at th10 of 2.6–3.0 mm as opposed to the 1.8–2.5 mm recorded in N. rhizinus.
Metabolograptus parvulus (Lapworth, Reference Lapworth1900)
Abundant material assigned to M. parvulus (Figs 3d, g–i, 10c–d) differs from the closely similar M. persculptus as shown by Štorch & Loydell (Reference Štorch and Loydell1996), Loydell et al. (Reference Loydell, Mallett, Mikulic, Kluessendorf and Norby2002), Loydell (Reference Loydell2007) and Blackett et al. (Reference Blackett, Page, Zalasiewicz, Williams, Rickards and Davies2009) in possessing a narrower rhabdosome, less overlapping thecae and closer thecal spacing. The maximum width of the present flattened rhabdosomes, including those oriented parallel or perpendicular to the tectonic strain, varies between 1.5 mm and 1.75 mm, 2TRD at th2 is 1.2–1.35 mm and 2TRD at th10 is 1.3–1.55 mm.
Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974)
The abundant large rhabdosomes of K. bifurcus (Figs 7c, k, 8g, ?j, 10a) possess a long and robust virgella bifurcating 3.0–6.5 mm below the sicular aperture into two equally robust, 10–15 mm long branches. The absence of rhabdosomes preserved in scalariform or sub-scalariform view suggests that the plane of bifurcation was parallel to the rhabdosome profile view. Thecae are gently geniculated, acuminate in profile view, having a convex supragenicular wall and aperture perpendicular to the rhabdosome axis. The proximal end is bluntly triangular, 1.0–1.15 mm wide at the level of th1¹–th1² apertures. A maximum width of 2.4–2.65 mm is attained by the 10th–15th thecal pair. In several specimens a robust nema splits into a large, irregular, net-like, probably membranous structure. Similar irregular branching developed on the forked virgella of the specimen questionably assigned to this species illustrated in Figure 8j. Material from the Austrian Southern Alps was identified as Rickardsograptus? bifurcus (Ye, Reference Ye1978) by Štorch & Schönlaub (Reference Štorch and Schönlaub2012), but further examination of the Chinese material suggests that ʻGlyptograptus bifurcusʼ Ye, Reference Ye1978 as well as ʻOrthograptusʼ lonchoformis Chen & Lin, Reference Chen and Lin1978 and ʻOrthograptus furcatusʼ Ye in Jin et al. Reference Jin, Ye, He, Wan, Wang, Zhao, Li, Xu and Zhang1982 are conspecific with the senior synonym ʻDiplograptus bifurcusʼ Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974. The pattern H (Mitchell, Reference Mitchell1987) early astogeny, largely glyptograptid thecae with little change of geniculation throughout the relatively slowly widening rhabdosome, lack of genicular thickening and rather late origin of the median septum refer this and related species to the genus Korenograptus defined by Melchin et al. (Reference Melchin, Mitchell, Naczk-Cameron, Fan and Loxton2011).
Korenograptus bicaudatus (Chen & Lin, Reference Chen and Lin1978)
Specimens assigned to this species (Figs 8a, r, ?i, 10s) match the type material of ʻClimacograptusʼ bicaudatus figured by Chen & Lin (Reference Chen and Lin1978, pl. 5, figs 10, 11) and refigured by Mu et al. (Reference Mu, Li, Ge, Lin and Ni2002, pl. 176, figs 3, 4). Korenograptus bicaudatus is characterized by its short virgella forked 0.25–0.5 mm from the sicular aperture into two long, ventrally directed and only slightly curved branches. The rhabdosome widens gradually from 0.9–1.1 mm at th1 to a maximum of 2.1–2.3 mm in profile view attained by th16–20. Scalariform specimens barely attain 2.0 mm in width. The 2TRD increases from 2.0 mm at th2 to 2.2–2.5 mm in distal thecae of mature, more than 30 mm long specimens. The short virgella and relatively narrow and deeply incised apertural excavations enable K. bicaudatus to be distinguished from other taxa with a bifurcated virgella described from the same level (ascensus–acuminatus Biozone) in China. The species, formerly assigned to Climacograptus, is commonly preserved in sub-scalariform view, but a specimen preserved in profile view (Fig. 8r) reveals its thecal geniculation and inclined supragenicular walls resembling other similar species here assigned to Korenograptus.

Figure 10. (a) Korenograptus bifurcus (Mu et al. in Nanjing Institute of Geology and Palaeontology, 1974): MGM-1716-S, sample EST 13a. (b, f) Normalograptus ednae sp. nov.: (b) holotype, MGM-1761-S, EST 16; (f) MGM-1762-S, EST 16. (c, d) Metabolograptus parvulus (Lapworth, Reference Lapworth1900): (c) MGM-8075-O, EST 8; (d) MGM-8074-O, EST 4. (e, p) Normalograptus minor (Huang, Reference Huang1982): (e) MGM-8090-O, EST 5; (p) MGM-8091-O, EST 4a. (g, o) Neodiplograptus shanchongensis (Li, Reference Li1984): (g) MGM-1722-S, EST 16; (o) MGM-1724-S, EST 11b. (h) Akidograptus ascensus Davies, Reference Davies1929: MGM-1739-S, EST 15. (i) Hirsutograptus sp.: MGM-1786-S, EST 20. (j) Cystograptus ancestralis Štorch, Reference Štorch1985: MGM-1778-S, EST 20. (k) Parakidograptus acuminatus (Nicholson, Reference Nicholson1867), early form: MGM-1757-S, EST 16. (?l, q) Normalograptus baridaensis sp. nov.: (?l) MGM-8097-O, EST 6; (q) holotype, MGM-8096-O, EST 2b. (m) Korenograptus lanpherei (Churkin & Carter, Reference Churkin and Carter1970): MGM-1749-S, EST 16. (n) Parakidograptus praematurus (Davies, Reference Davies1929): MGM-1731-S, EST 13a. (r) Normalograptus aff. rhizinus (Li & Yang in Nanjing Institute of Geology and Mineral Resources, 1983): MGM-8104-O, EST 7. (s) Korenograptus bicaudatus (Chen & Lin, Reference Chen and Lin1978): MGM-1748-S, EST 12. All specimen in the same magnification, black bar represents 1 mm.
Korenograptus lanpherei (Churkin & Carter, Reference Churkin and Carter1970)
Robust rhabdosomes (Figs 7h, 8f, l, 10m) attaining a length of c. 60 mm in full maturity match material described by Loxton (Reference Loxton2017) in most characters and measurable parameters including width and 2TRD. Greater maximum width, up to 2.9 mm, occurs in mature specimens flattened in full profile. The sub-triangular proximal end is furnished with a robust, more than 12 mm long virgella without any bifurcation. The ascending part of its th11 is inclined at an angle of 17–19° to the rhabdosome axis. Subsequent thecae possess rounded, glyptograptid genicula and straight or slightly convex supragenicular walls inclined at 23–38°. Low values occur in slightly sub-scalariform specimens and/or specimens oriented more nearly parallel to the principal elongation. Thecal apertures are horizontal, and each occupies one-quarter of the rhabdosome width. Two thecae repeat distance attains 2.2–2.55 mm in the distal part of particularly long rhabdosomes. Korenograptus lanpherei differs from its stratigraphic predecessor K. bifurcus in having a very long but simple virgella. Korenograptus bicaudatus possesses a short bifurcated virgella, narrower rhabdosome width and more geniculated thecae. The plane of virgellar bifurcation is probably oblique or perpendicular to the rhabdosome profile. The long and robust virgella, longer ascending part of th11, less geniculated thecae and greater width of its rapidly widening rhabdosome differentiate K. lanpherei from the otherwise similar K. lungmaensis (Sun, Reference Sun1933), although a rhabdosome from the base of the ascensus Biozone, assigned to the latter species by Chen, Rong & Fan (Reference Chen, Rong and Fan2003, fig. 1H), also possesses a long virgella, triangular proximal end and robust nema. This form also resembles Nd. parajanus in general rhabdosome appearance and many measurable parameters (rhabdosome width, early astogeny of pattern H, thecal spacing, long robust undivided virgella and nema) but can be readily distinguished by its sub-triangular proximal end and uniform glyptograptid thecae with flowing genicula throughout the rhabdosome. The thecae of Nd. parajanus, by comparison, are markedly biform: strongly geniculated, almost climacograptid in the proximal part of the rhabdosome and nearly orthograptid distally.
Neodiplograptus shanchongensis (Li, Reference Li1984)
Neodiplograptus shanchongensis from the persculptus and lower ascensus–acuminatus biozones of China (Li, Reference Li1984; Chen et al. Reference Chen, Rong, Mitchell, Harper, Fan, Zhan, Zhang, Li and Wang2000, Reference Chen, Fan, Melchin and Mitchell2005), Arctic Canada (Melchin, McCracken & Oliff, Reference Melchin, McCracken and Oliff1991), southern Scotland (Fan, Melchin & Williams, Reference Fan, Melchin, Williams, Sheehan and Servais2005), Jordan (Loydell, Reference Loydell2007) and Saudi Arabia (Williams et al. Reference Williams, Zalasiewicz, Boukhamsin and Cesari2016) can be differentiated from the rather similar Nd. lanceolatus by its thecal geniculation developed until the distal part of the rhabdosome, less steeply inclined supragenicular walls, and more parallel-sided rhabdosome widening from 1.3–1.4 mm at the first thecal pair to a maximum of 2.1 mm attained by th8–10. New records from the Estana section (Figs 7p, x, 10g, o) suggest that Nd. shanchongensis may be the stratigraphical praecursor of Nd. lanceolatus which appears in abundance in the higher and lower, but apparently not the lowermost, part of the ascensus–acuminatus Biozone.
Parakidograptus praematurus (Davies, Reference Davies1929)
Specimens of Par. praematurus (Figs 7e, i, 8b, p, 10n), which are fairly common in the lower part of the ascensus–acuminatus Biozone, are distinguished from early populations of Par. acuminatus (Figs 7d, aa, 10k) by their less-exposed sicula, lower position of thl1 bud on the sicula (˂ 0.2 mm from the sicular aperture), usually lesser maximum width (1.5 mm) and generally less protracted proximal end. Thecae are more geniculated throughout the rhabdosome when compared with flattened Par. acuminatus. Parakidograptus praematurus also lacks the acuminate thecal apertural margins typical of Par. acuminatus, including its stratigraphically lowest specimens which are marked by a less-protracted proximal part and lesser rhabdosome width (˂ 2.0 mm) than the stratigraphically higher specimens recorded from this locality above the measured section. Akidograptus ascensus can be distinguished from our specimens assigned to Par. praematurus by its less-inclined supragenicular thecal walls, usually lesser maximum width (0.8–1.2 mm), and more protracted proximal end with long exposed sicula and turning point of th11 situated c. 0.5 mm above the sicular aperture.
9. Systematic part
Normalograptus baridaensis sp. nov.
Figures 3j–k, m, 10 ?l, q
Derivation of name. After el Baridà, the small region of the upper Segre valley in which the Estana section is located and where the Late Ordovician stratigraphical units of the central Pyrenees were defined.
Holotype. MGM-8096-O (Figs 3j, 10q) from sample EST 2b, in the uppermost Hirnantian M. parvulus Biozone of the Estana section, central Pyrenees, Lleida Province, Spain.
Material. Twelve flattened specimens with proximal end furnished with virgella and secondary basal spine.
Diagnosis. Slender, 0.9–1.1 mm wide Normalograptus with long virgella and single straight secondary spine projecting with low angle of divergence from base of 0.4–0.6 mm long robust part of virgella.
Description. The slender, only 0.9–1.1 mm wide, parallel-sided rhabdosome of N. baridaensis is furnished with a 4–6 mm long virgella with a considerably thickened 0.4–0.6 mm long basal part and a single, straight 2–4 mm long secondary spine projecting from the end of the thickened part of the virgella with an angle of divergence usually of 4–5°. Alternating normalograptid thecae are strongly geniculated with gently inclined supragenicular walls, asymmetrical apertural excavations and a 2TRD ranging from 2.1 mm to 2.6 mm at the level of th10.
Remark. This uncommon form from the uppermost part of the parvulus (i.e. upper persculptus) Biozone can be distinguished from N. bifurcatus Loydell, Reference Loydell2007 by its wider thecal spacing and lack of any bifurcated antivirgellar spine.
Normalograptus ednae sp. nov.
Figures 7g, q, bb, 8q, 10b, f
Derivation of name. After the feminine name Edna, the second author's granddaughter.
Holotype. MGM-1761-S (Figs 7q, 10b) from sample EST 16, lowermost Rhuddanian lower ascensus–acuminatus Biozone of the Estana section, central Pyrenees, Lleida Province, Spain.
Material. Twelve mature specimens with proximal end, virgella and lateral spines and several incomplete or immature rhabdosomes.
Diagnosis. Normalograptus gradually widening from 0.7 mm at first thecal pair to distal maximum of 1.5 mm, with strongly geniculated thecae, parallel-sided supragenicular thecal walls, short and robust virgella and two claw-like lateral spines originating from the upward turning point of th11.
Description. Rhabdosome attaining length of c. 20–25 mm widens gradually from 0.7 mm at the level of the first thecal pair apertures, through 0.95–1.05 mm at th5, and 1.2–1.3 mm at th10 to a maximum of 1.5 mm attained by the 13–17th theca. The slightly asymmetrical proximal end is furnished with a 0.2–1.5 mm long virgella and two equally short and robust, somewhat claw-like curved lateral spines. Spines grew from the upward turning point of th11, probably parallel to the ventral margins of the rhabdosome. The upward growing part of th12 is relatively long (1.0–1.2 mm) compared with the narrow proximal end of the rhabdosome. Alternating climacograptid thecae exhibit parallel-sided supragenicular walls, sharp genicula and slightly asymmetrical apertural excavations. Two thecae repeat distance ranges from 1.8 mm to 2.5 mm throughout the rhabdosome, depending on the tectonic strain.
Discussion. Normalograptus ednae can be distinguished from N. trifilis by its narrower rhabdosome, gradually widening to a lesser maximum width of 1.5 mm compared with 2.0 mm in the latter species. The basal spines of N. ednae are short, robust and slightly curved, whereas both the virgella and basal spines of N. trifilis are long, slender and almost straight. In the Estana section, N. ednae is confined to a narrow stratigraphical interval in the lower part of the ascensus–acuminatus Biozone, below the FAD of N. trifilis which is a typical faunal component of the lower–middle ascensus–acuminatus Biozone.
Acknowledgements
PŠ appreciates the financial support provided by the Czech Science Foundation (grant no. 14–16124S) and in-house support received from the Institute of Geology of the Czech Academy of Sciences (no. RVO 67985831). JRB paid his own expenses. Work carried out by JCGM was funded by the Spanish Ministry of Economic Affairs, Industry and Competitivity (grant no. CGL2017-87631-P). We are indebted to David Loydell for valuable comments and for improving the English. Štěpán Manda, Sun Zongyuan and Mercè Secall are thanked for their assistance in the field. This is a contribution to the IGCP project 653 (IUGS-UNESCO).