


Introduction
The number of penguin species (fourteen) and size classes (six) obtained from the late Eocene localities of Isla Marambio (La Meseta Formation, Elliot & Trautmant 1982), Antarctica, exceed the highest number of sympatric species (four) and size classes currently occurring in regional breeding from the cold sub-Antarctic to cool temperate waters south of the Subtropical Convergence. There may be palaeobiological or taphonomical reasons for this high percentage, but it is probable that it also reflects a real abundance of penguins, which appear to have been the dominant coastal birds in the late Eocene of Antarctica (Case Reference Case1996). Even though La Meseta Formation is one of the richest units in terms of diversity and abundance of penguins, no articulated or associated skeleton has previously been described. The Anthropornis nordenskjoeldi biozone defined within the Submeseta Allomember constitutes the unit with highest specific diversity of fossil penguins ever known, with fourteen recorded species (Tambussi et al. Reference Tambussi, Acosta Hospitaleche, Reguero and Marenssi2006, however see Jadwiszczak Reference Jadwiszczak2006b). This type of deposit, with high abundance of isolated elements, is not exclusive of Antarctic sediments, but it is also characteristic of most fossil penguin assemblages in the world.
As in other cases, the remains are largely accumulated close to nesting areas of the breeding colonies. These deposits are usually represented by isolated and sometimes fragmented bones. This taphonomic characteristic of the record has compelled palaeontologists to propose a systematic scheme entirely based on isolated skeletal remains. A consequence of this is that most species are known only through a single element, the tarsometatarsus in most cases, or the humerus in a few others. Only a small number of fossil species, whose diagnoses are recent (Slack et al. Reference Slack, Jones, Ando, Harrison, Fordyce, Arnason and Penny2006, Acosta Hospitaleche et al. Reference Acosta Hospitaleche, Tambussi, Donato and Cozzuol2007, Clarke et al. Reference Clarke, Ksepka, Stucchi, Urbina, Giannini, Bertelli, Narváez and Boyd2007), have been described from articulated elements.
In the present contribution, the first articulated skeleton from Antarctica is studied. An osteologic description of the recovered elements and a brief discussion about its systematic assignment are given. The significance of this finding in the context of the Anthropornis nordenskjoeldi biozone is also discussed.
Material and methods
During the 2008 field season on Isla Marambio one of the authors (M.R.) collected in situ a partially complete skeleton of a fossil spheniscid. The fossil studied here comes from the lower level of the Submeseta Allomember, La Meseta Formation (Marenssi et al. Reference Marenssi, Santillana and Rinaldi1998a) or TELM 7 (Elliot & Trautman Reference Elliot and Trautman1982), which outcrops at several localities. The penguin-bearing horizon is composed of thick shelly conglomerates, well-sorted sands and interlaminated sand/mud channel-fills with thin shelly conglomeratic intervals.
Specimen MLP 96-I-6-13 and the comparison materials are housed at the División Paleontología de Vertebrados of the Museo de La Plata (MLP), Argentina. Diagnostic characters for Antarctic penguin species are taken from Myrcha et al. (Reference Myrcha, Jadwiszczak, Tambussi, Noriega, Gazdzicki, Tatur and Del Valle2002). Osteological terminology follows Baumel & Witmer (Reference Baumel and Witmer1993). Measurements were taken with a Vernier Caliper (0.01 mm accuracy).
Geological and depositional setting
A series of prolific fossil sites and quarries are now known from La Meseta Formation in Isla Marambio, James Ross Basin (del Valle et al. Reference Del Valle, Elliot and Macdonald1992), Antarctic Peninsula, as a result of work conducted by various research groups since the mid 1980s (Fig. 1). The La Meseta Formation is composed of sandstones and mudstones with interbedded shell-rich conglomerates, organized into six erosionally-based internal units, named from base to top Valle de Las Focas, Acantilados, Campamento, Cucullaea I, Cucullaea II and Submeseta allomembers (Fig. 2). These units were deposited during the Eocene in deltaic, estuarine and shallow marine settings, mostly within a north-west–south-east trending valley (Marenssi et al. Reference Marenssi, Santillana and Rinaldi1998a, Reference Marenssi, Santillana and Rinaldi1998b).

Fig. 1 a. Map showing the location of Isla Marambio, Antarctic Peninsula. b. Sketch map of the northern part of Isla Marambio showing the distribution of the Submeseta Allomember and the fossil penguin-bearing locality cited in the text.

Fig. 2 a. Stratigraphic section of the La Meseta Formation, Isla Marambio, Antarctic Peninsula (modified from Reguero et al. Reference Reguero, Marenssi and Santillana2002). b. Measured section from the eastern flank of the plateau (stratotype of the Anthropornis nordenskjoeldi biozone) showing stratigraphic levels of the Cucullaea II and Submeseta Allomembers (modified from Marenssi Reference Marenssi1995). Shaded rectangular area delimits the Anthropornis nordenskjoeldi biozone.
The fossil presented here comes from the upper-level Submeseta Allomember in the south-western slope of the plateau (meseta) of the island (Fig. 1). Almost all fossil vertebrate specimens from the Submeseta Allomember were found in a stratigraphic interval that is easily distinguishable by the massive occurrence of penguin bones and the phosphatic brachiopod “Lingula”, and stratigraphically located 30–35 m below the top of the 145 m thick Submeseta Allomember, within the Anthropornis nordenskjoeldi biozone (Tambussi et al. Reference Tambussi, Acosta Hospitaleche, Reguero and Marenssi2006, but see Jadwiszczak Reference Jadwiszczak2006b). The locality DPV 10/84 is located 70–75 m below this rich vertebrate-bearing horizon (Fig. 2b), and approximately 20 m above the contact between the Cucullaea II and Submeseta allomembers. The base of the Submeseta Allomember is correlated with the main lowstand at 36 Ma, and the first part of this unit is characterized by estuarine heterolithic sediments followed by a set of aggradational-retrogradational shoreface parasequences where this specimen was deposited (Marenssi Reference Marenssi2006). The age of the fossiliferous interval is constrained by the age of the basal unconformity (Marenssi Reference Marenssi2006, 36 Ma) and the Sr-derived age of Dingle & Lavelle (Reference Dingle and Lavelle1998) at 34.2 Ma.
Marine vertebrates such as the gadiform “Mesetaichthys”, a few sharks (Pristiophorus and Carcharias), and a primitive mysticete whale (Llanocetus denticrenatus) as well as archaeocetes are also present in this zone (personal observations). A small number of penguin bones assigned to Anthropornis and Delphinornis were found close to the specimen described here.
Sedimentologically, the locality DPV 10/84 is included in a facies association (facies association II of Marenssi et al. Reference Marenssi, Santillana and Rinaldi1998b), and is characterized by conglomeratic beds to mudstones with a diverse and abundant macrofauna that correspond to a valley-confined estuary mouth/inner estuary complex. Tidal channels and mixed flats, tidal inlets and deltas, and washover and beach environments represent the interfingering of high and low energy environments (Marenssi et al. Reference Marenssi, Santillana and Rinaldi1998b). Penguin bones recovered are usually disarticulated; some are broken; most are complete albeit with various degrees of abrasion. Most if not all of them were transported at least for a short time before burial and therefore their accumulations represent parauthoctonous assemblages.
The Palaeeudyptes dilemma
Before dealing with the descriptive analysis of the specimen under study, we need to introduce the taxonomic problem that pervades this genus. Four species have been nominated for this genus. Palaeeudyptes antarcticus Huxley, 1859 happened to be the first fossil species of penguin to be described, and consequently became the type species of Palaeeudyptes.
A partial associated skeleton that included elements of the wing and leg was described by Hector (Reference Hector1872) and assigned to P. antarcticus. Subsequently, other remains, which were originally attributed to this same individual (Hector Reference Hector1873), were reviewed by Oliver (Reference Oliver1930) and assigned to Pachydyptes ponderosus Oliver, Reference Oliver1930. However, the presence of an associated tarsometatarsus among these remains is not mentioned in these later works. The subsequent references to this skeleton made by Lambrecht (Reference Lambrecht1933) and Marples (Reference Marples1952) confirm the absence in the collections of nearly half of the skeletal elements that had been described by Hector (Reference Hector1872, Reference Hector1873).
Other materials from New Zealand collections, assigned on repeated occasions to P. antarcticus (see details in Marples Reference Marples1952), are not unquestionably associated with tarsometatarsal elements comparable with the type materials. Also, the provenance attributed by Lambrecht (Reference Lambrecht1933) to this skeleton, was not stated by Hector (Reference Hector1872, Reference Hector1873) in his original contributions and it was later said to be unknown by Simpson (Reference Simpson1971). Additional materials detailed in Marples (Reference Marples1952) and probably assigned to the same species acknowledged by Hector (Reference Hector1872), cannot be conclusively assigned to P. antarcticus at present.
The discovery of two specimens from New Zealand showing associated skeletal elements began to change this systematic panorama (Ksepka et al. Reference Ksepka, Bertelli and Giannini2006, Acosta Hospitaleche et al. Reference Acosta Hospitaleche, Tambussi, Donato and Cozzuol2007), which was up to that point based on isolated tarsometatarsi. A humerus associated with a tarsometatarsus (no. C43-80) from the Burnside marl (Kaiatan, Upper Eocene) from Burnside, near Dunedin and curated at the Museum of Otago was described by Marples (Reference Marples1952) and determined as P. antarcticus. In the same way, other articulated elements (no. C47-17) from Burnside greensland (Waitakian, middle Oligocene) were taxonomically assigned by Marples (Reference Marples1952) to P. antarcticus, and they are morphologically comparable to the ones published by Hector (Reference Hector1872). These discoveries represented the first opportunity to jointly study the features and proportions of the tarsometatarsus and humerus in Palaeeudyptes.
Subsequently, the species Palaeeudyptes gunnari (Wiman, 1905) was named, based on an incomplete tarsometatarsus, which had originally been assigned to Eospheniscus Wiman, 1905 and then placed into Palaeeudyptes by Simpson (Reference Simpson1971).
The third species to be erected was Palaeeudyptes klekowsii Myrcha, Tatur & del Valle, 1990 on the basis of a fragmented tarsometatarsus, larger than the other known species of Palaeeudyptes (Myrcha et al. Reference Myrcha, Tatur and Del Valle1990).
Finally, Palaeeudyptes marplesi Brodkorb, Reference Brodkorb1963 is the last described species of the genus, founded on the basis of the reinterpretation of elements previously assigned to P. antarcticus (see Marples Reference Marples1952) and subsequently to Palaeeudyptes cf. antarcticus (see Simpson Reference Simpson1957), which Brodkorb (Reference Brodkorb1963) reallocated into this new species. A revised diagnosis was provided by Simpson (Reference Simpson1971) when reviewing the New Zealand pre-Pliocene material. His results show only size differences with respect to the other species of the genus. No other remains were assigned to this taxon by Simpson (Reference Simpson1971) or any subsequent author.
According to this assessment, each of these four species is represented by tarsometatarsi, and only two (P. antarcticus and P. marplesi) have other associated elements. Consequently, it is currently impossible to carry out a systematic assignment of any element other than a tarsometatarsus to P. klekowskii and P. gunnari, at least until other articulated remains are found. However, Jadwiszczak (Reference Jadwiszczak2006a) assigns different skeletal elements based on their size and robustness.
A thorough assessment of the characters present in the tarsometatarsus of the Antarctic representatives was performed by Myrcha et al. (Reference Myrcha, Jadwiszczak, Tambussi, Noriega, Gazdzicki, Tatur and Del Valle2002) on the occasion of the review of the Polish and Argentine penguin collections acquired on Isla Marambio (Myrcha et al. Reference Myrcha, Jadwiszczak, Tambussi, Noriega, Gazdzicki, Tatur and Del Valle2002). According to them, P. gunnari and P. antarcticus are present in Antarctic sediments, together with another twenty specimens assigned to genus Palaeeudyptes but too incomplete to be taxonomically assigned to any species.
Jadwiszczak (Reference Jadwiszczak2006a) published another systematic reference work in which the diagnosis of these species was amended. As in other recent contributions (Kanfeder Reference Kanfeder1994, Tambussi et al. Reference Tambussi, Acosta Hospitaleche, Reguero and Marenssi2006), he follows the criterion of Wiman (Reference Wiman1905b) regarding the “size groups”.
Wiman’s “size groups”
The first collections of Antarctic fossil penguins, collected on Isla Marambio by the Swedish expedition in 1901–1903, was studied by Wiman (Reference Wiman1905a, Reference Wiman1905b), who proposed a new approach to deal with the nature of fossil penguin assemblages. Due to the fragmentary state of the bones, he did not classify them all in a systematic scheme. Instead, he proposed grouping them in eight categories according to their size and robustness; the third, fifth and seventh of these show some degree of intragroup variability. Each category includes several remains that belong to different individuals but are morphologically similar and could be conspecific.
Group 1 was established for the largest synsacrum (holotype of Orthopteryx gigas, Wiman Reference Wiman1905b), an element known only from a few fossil penguin species, and no other elements were subsequently assigned to it.
Group 2 remained unnamed and also consists of a synsacrum, but in this case incomplete and not assigned to any particular species.
Group 3 was identified as Anthropornis nordenskjoeldii Wiman, 1905, on the basis of a tarsometatarsus and comprises several elements (tarsometatarsus, coracoid, humerus, ulna, carpometacarpus, femur, tibiotarsus, synsacrum) as well.
Group 4, recognized as Pachypteryx grandis Wiman, 1905, was based on a tarsometatarsus and also contains others appendicular elements (coracoid, radius, carpometacarpus, tibiotarsus). In the current systematic scheme (Ksepka et al. Reference Ksepka, Bertelli and Giannini2006, Acosta Hospitaleche et al. Reference Acosta Hospitaleche, Tambussi, Donato and Cozzuol2007), this species is considered a synonym of Anthropornis grandis (Wiman Reference Wiman1905a).
In a later revision, Simpson (Reference Simpson1946) proposed that the four above mentioned groups be sorted in the same category, probably assigned to Anthropornis nordenskjoeldii, an idea later supported by Marples (Reference Marples1953).
Group 5 was associated with Eospheniscus gunnari Wiman, 1905 - currently Palaeeudyptes gunnari (Wiman, Reference Wiman1905a) Simpson, Reference Simpson1971 - on the basis of a tasometatarus, type of the species. This group also comprises many other elements (coracoid, humerus, ulna, femur, tibiotarsus, synsacrum). It is worth repeating that none of these bones are associated and that they belong to different specimens. However, this was not taken into account in subsequent works, and the allocations made by Wiman (Reference Wiman1905a) were considered in a systematic context.
Group 6 was also innominate and was based on fragments of a humerus, coracoid, scapula, and femur. In a later review, Notodyptes wimani Marples, Reference Marples1953 was also included in this category. This species was erected on the basis of an incomplete tarsometatarsus from the same penguin assemblage in Isla Marambio (Marples Reference Marples1953) and placed later within Archaeospheniscus wimani (Marples, Reference Marples1953) Simpson, Reference Simpson1971.
Group 7 includes a single tarsometatarsus that was the basis for the creation of Delphinornis larsenii Wiman, Reference Wiman1905a.
Finally, group 8 represents the smallest species, identified as Ichtyopteryx gracilis Wiman, Reference Wiman1905a. This species is based on a fragmentary tarsometatarsus that Brodkorb (Reference Brodkorb1963) considered belonging to a distinct species. Meanwhile, Marples (Reference Marples1953) and Myrcha et al. (Reference Myrcha, Jadwiszczak, Tambussi, Noriega, Gazdzicki, Tatur and Del Valle2002) completely disregarded it due to its poor state of preservation.
Systematic Palaeontology
AVES
SPHENISCIFORMES Sharpe, 1891
Family SPHENISCIDAE Bonaparte, 1831
Palaeeudyptes Huxley, 1859
Type species. Palaeeudyptes antarcticus Huxley, 1859
Included species. P. antarticus Huxley, 1859, P. gunnari (Wiman, Reference Wiman1905a), P. marplesi Brodkorb, Reference Brodkorb1963, P. klekowsii Myrcha, Tatur & del Valle, 1990.
Palaeeudyptes gunnari (Wiman, Reference Wiman1905a)

Fig. 3 Pectoral elements of Palaeeudyptes gunnari (MLP 96-I-6-13) from Isla Marambio, locality DPV 10/84, Antarctic Peninsula. a. & b. right humerus, anterior and posterior views. c. Incomplete right carpometacarpus, posterior view. d. Incomplete right ulna, posterior view. e. Incomplete right radius, anterior view. f. Digiti minoris phalanx, distal view. g. Proximal digiti majoris phalanx, distal view. h. Distal digiti majoris phalanx, distal view. i. Proximal end of right scapula, dorsal view. j. Distal end of right left coracoids, posterior view. k. Omal portion of right coracoids, proximal view. l. Incisura intercostalis of sternum, lateral view. Scale bar: 1 cm.
Referred material. MLP 96-I-6-13 (complete right humerus, almost complete right carpometacarpus, four fragments of left carpometacarpus, fragmentary left and right ulnae and radii, three digital phalanges of the wing, two tiny fragments of a scapula, fragmentary distal end of omal portion of coracoid, fragment of sulcus carinae of sternum, three articulated dorsal vertebrae and several other fragments of indeterminate vertebrae, synsacrum, lateral portion of proximal end and condylus lateralis of right femur, proximal end and condylus lateralis of right tibiotarsus, condylus medialis of left tibiotarsus, left fibula, distal portion of left tarsometatarsus preserved in two pieces, eight pedal phalanges and two ungual phalanges).
Provenance. The locality DPV 10/84 is located on the south-western slope of the Isla Marambio plateau (Fig. 1b). Stratigraphically is located within the Submeseta Allomember, on the upper levels of La Meseta Formation (late Eocene, Fig. 2b).
Remarks. A revised list of tarsometatarsal features was given by Myrcha et al. (Reference Myrcha, Jadwiszczak, Tambussi, Noriega, Gazdzicki, Tatur and Del Valle2002) in their generic and specific revised diagnosis. Posteriorly, others characters of the coracoid, humerus, ulna and carpometacarpus were included in the generic diagnosis by Jadwiszczak (Reference Jadwiszczak2006a), and some others features of the synsacrum, humerus, ulna, femur and tibiotarsus were incorporated in the specific diagnosis (Jadwiszczak Reference Jadwiszczak2006a).
Additionally, in his thorough taxonomic revision, Jadwiszczak (Reference Jadwiszczak2006a) assigned several different skeletal elements to this species, based mostly on their size.
Description of MLP 96-I-6-13 (Figs 3 & 4)
A brief description of the osteological characters present in this specimen is provided below. Detailed comparisons with other species have already been provided by other authors (Marples Reference Marples1952, Simpson Reference Simpson1971 and literature cited therein), and will not be repeated here.
Humerus (Fig. 3a & b). It is medium-sized, compared to the Eocene Antarctic penguin species, smaller than those of P. antarcticus and P. klekowskii. The diaphysis is sigmoid and robust, with a slight angulus preaxialis. The caput humeri is sturdy and the undivided fossa tricipitalis is comparatively small. The edges of this fossa are weak, except in the area of the well developed tuberculum ventrale.
The sulcus ligamentosus transversus is wide and continues to the incisura capitis. There is a weak and incomplete gap between these structures. Both are deep, as previously described by Marples (Reference Marples1953). The sulcus ligamentosus transversus is oblique to the axis of the diaphysis and its most proximal end is shallower than the other extreme connected to the incisura capitis. The former is developed toward the facies cranialis reaching almost the facies caudalis. Two muscular impressions are developed on the facies musculi supracoracoideus. One of them is elongated and parallel to the diaphysis, and the other one, which is closer to the fossa tricipitalis, is deeper and rounded. The impressio coracobrachialis is large and limited by a narrow crista deltopectoralis.
The distal epiphysis is especially robust and the shaft of the trochlear angle is small (see Acosta Hospitaleche Reference Acosta Hospitaleche2004, Acosta Hospitaleche et al. Reference Acosta Hospitaleche, Tambussi, Donato and Cozzuol2007).
The condylus dorsale continues toward the extremitas distalis humeri constituting a radial condyle like a wide platform. The condylus ventrale is also strong and elongated. The incisura intercondylaris is wide and deep, limiting with a prominent tuberculum supracondylare ventrale. There is also a deep and conspicuous fossa supracondylare dorsale. The extremitas distalis humeri are of a non-Spheniscus type; i.e. the most caudal one is distally projected beyond the corpus humeri (see Acosta Hospitaleche Reference Acosta Hospitaleche2004, Acosta Hospitaleche et al. Reference Acosta Hospitaleche, Tambussi, Donato and Cozzuol2007).
Measurements: total maximun length 137 mm, proximal width of the head taken latero-medially 43 mm, latero-medially width of the distal end 31 mm, anteroposterior width of the head 27 mm, anteroposterior width at diaphyseal axis 12 mm, fossa tricipitalis maximum diameter 12 mm.
Carpometacarpus (Fig. 3c). It is a robust and straight bone with the proximal end rounded and flat. The fovea carpalis caudalis is shallow and sub-rounded. The sulcus tendoneus is slightly marked, and rounded toward the distal end. The trochlea carpalis is poorly developed and the fossa supratrochlearis is shallow.
The facies articularis digitalis major is represented by two sharp prominences.
Radius (Fig. 3e). Only the proximal half is preserved. It presents a sub-triangular and slightly concave cotyla humeralis. The tuberculum occipitalis radialis is strong and robust.
Ulna (Fig. 3d). It is flat and compact, subtriangular in shape. The proximal end is almost complete and exhibits a pointed crista intercotylaris dividing two elongated cotyla.
Phalanges of the wing (Fig. 3f–h). The distal and proximal phalanges of digiti majoris and the phalanx of digiti minoris are preserved with slightly fragmented distal ends. It is not possible to determine which wing they belong to.
Scapula (Fig. 3i). The small portion of the extremitas cranialis displays a tuberculum coracoideum rounded and not very prominent.
Coracoid (Fig. 3j & k). Only two small fragments are preserved. On of them is a proximal end preserving the facies articularis clavicularis and circular cotyla scapulae next to the flat facies articularis humeralis. The other fragment corresponds to a tiny portion of the sternal end.
Sternum (Fig. 3l). A small piece is preserved corresponding to a portion of the incisura intercostalis is preserved; it does not display any particular features.
Vertebrae and synsacrum (Fig. 4a & b). Three thoracic vertebrae still joined are completely preserved. The processus transversus are slightly caudally projected. The fragment of synsacrum is much deteriorated and no particular osteologic character can be observed.
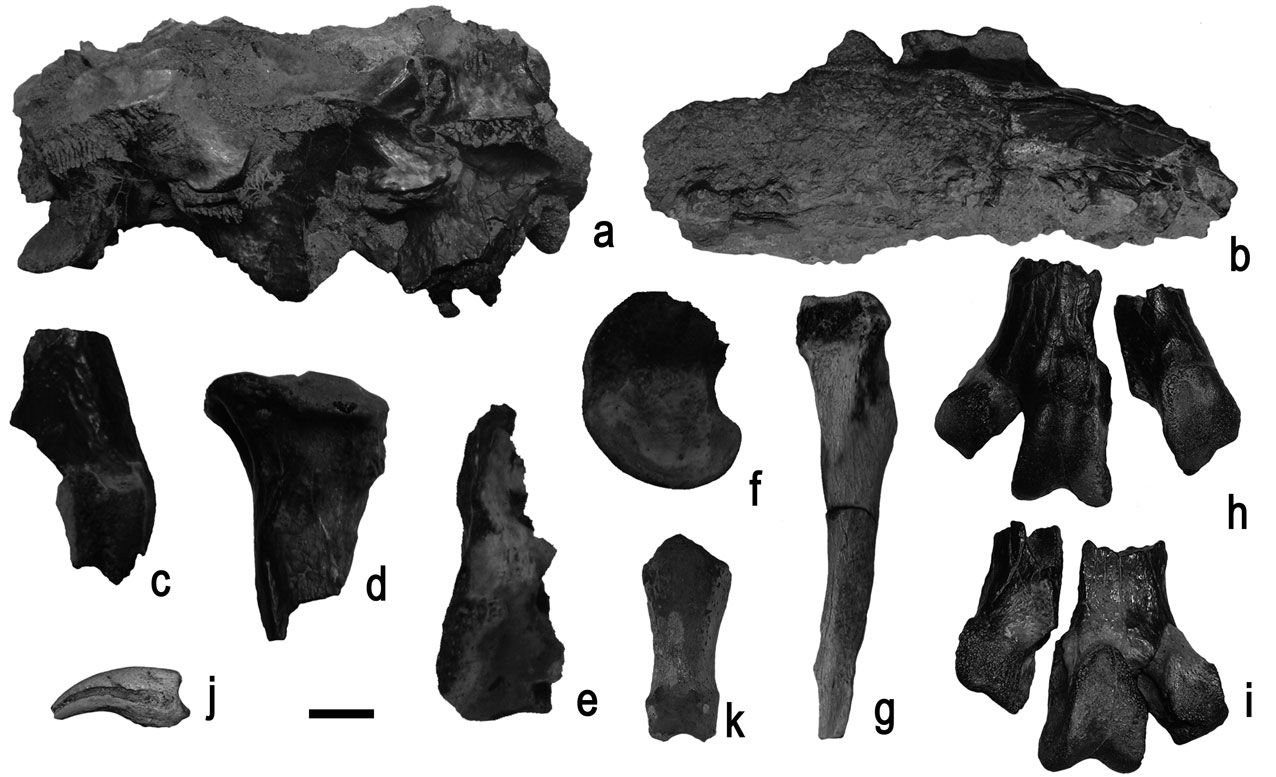
Fig. 4 Vertebrae and pelvic elements of Palaeeudyptes gunnari (MLP 96-I-6-13) from Isla Marambio, locality DPV 10/84, Antarctic Peninsula. a. thoracic vertebrae, lateral view. b. Synsacrum, lateral view. c. Distal end of right femur, anterior view. d. Proximal end of right tibiotarsus, anterior view. e. Distal end of right tibiotarsus, anterior view. f. Medial condyle of left tibiotarsus, medial view. g. Right fibula, anterior view. h. & i. Distal end of left tarsometatarsus in anterior and posterior views. j. Ungual phalanx, anterior view. k. Pedal phalanx, anterior view. Scale bar: 1 cm.
Femur (Fig. 4c). It is preserved in several fragments, and it was not possible to reconstruct it in a single piece. The impressiones illiotrochantericas are deep and well developed. A portion of the caput femoris shows a facies articularis acetabularis with no particular characters.
The condylus medialis is rounded with a small depression in the centre of the medial surface. The condylus lateralis is eroded and the trochlea fibularis is broken at its distal end. The crista tibiofibularis is damaged but it seems to have been rounded.
Tibiotarsus (Fig. 4d–f). Three badly damaged fragments were preserved. The proximal end shows a well elevated and rounded area interarticularis. The fossa flexoria is deep and laterally expanded.
The condylus medialis presents a strong edge more pronounced at the distal end. The outline is elongated and it encloses a prominent epicondylus medialis in the centre. A fragment of the trochlea cartilaginis tibialis with no particular characters has also been preserved.
Fibula (Fig. 4g). The corpus fibulae is complete and very well preserved. The spina fibulae is robust and not as straight as in living penguins. The caput fibulae is wide and the facies articularis fibularis is concave, while the lateral opposite surface is flat.
Tarsometatarsus (Fig. 4h & i). The corpus tarsometatarsi is relatively massive, smaller than that of P. klekowski, and the margo medialis is slightly concave.
The trochlea metatarsi II extends medially in a considerable way compared to living penguins, and the incisura intertrochlearis medialis is unusually deep, as in the holotype. Besides, the trochlea metatarsi II is caudally oriented with respect to the trochlea metatarsi III. Its edges are not well marked, and it is subrounded in medial view.
The trochlea metatarsi III is larger in size and it has strong edges. Consequently the trochlea has a triangular shape, wider at the proximal end. It is rounded in both lateral and medial views.
The fossa supratrochelaris plantaris is wide and there is also a shallow fossa proximal to the trochlea metatarsi III on the facies cranialis, which appears typically in the Antarctic species of Palaeeudyptes.
The trochlea metatarsi IV is intermediate in size compared to the other trochleae and elongated in lateral view. Unfortunately it is broken and it was not possible to reconstruct its exact location on the tarsometatarsus.
Measurements: distal lateral width of trochlea metatarsi II c. 9 mm (it is eroded), anteroposterior width of trochlea metatarsi II 13 mm, distal lateral width of trochlea metatarsi III 13 mm, anteroposterior width of trochlea metatarsi III 16 mm, (and 18 mm over trochlear edge), distal lateral width of trochlea metatarsi IV 10 mm, anteroposterior width of trochlea metatarsi IV 15 mm.
Pedal phalanges (Fig. 4j & k). Eight phalanges proximalis et intermediae are preserved. On the two ungual phalanges, the sulcus neurovascularis that runs on each side of the corpus phalangis, is curved and does not reach the basis phalangis. Unfortunately it is not possible to identify which digit they correspond to due to the absence of diagnostic characters in all phallanges in general (Mosto et al. unpublished) and only when more complete materials are available will it be possible to reconstruct the position of each one (e.g. Acosta Hospitaleche et al. Reference Acosta Hospitaleche, Tambussi, Donato and Cozzuol2007, but see Jadwiszczak Reference Jadwiszczak2006a).
Discussion and conclusions
The systematic paradigm has changed over the years (Ksepka et al. Reference Ksepka, Bertelli and Giannini2006, Acosta Hospitaleche et al. Reference Acosta Hospitaleche, Tambussi, Donato and Cozzuol2007), and the taxonomic studies of penguin are not immune to such changes. Along with this evolution of systematic analyses, the repeated findings of associated and sometimes articulated remains enable a new approach completely different to that of classical studies. Herein lies the importance of specimen MLP 96-I-6-13, which represents the first articulated skeleton known of Palaeeudyptes gunnari, a species that was previously only known from isolated tarsometatarsi.
Although the species of a genus can be differentiated from their tarsometatarsus sizes, this is not necessarily true for other elements of the skeleton such as the humerus, and the use of the size criterion introduces high uncertainty.
Comparative studies have been carried out in modern penguins, with the conclusion that the species of a same genus show allometric differences when appendicular elements of the wing and leg are considered. For example, the three species of Pygoscelis have been thoroughly described by Acosta Hospitaleche (Reference Acosta Hospitaleche2004), and a preliminary analysis of these relationships has been performed by Balseiro et al. (Reference Balseiro, Halpern and Sferco2004).
MLP 96-I-6-13 presents all the characters mentioned by Wiman (Reference Wiman1905a, Reference Wiman1905b) in his original diagnosis and validated by Myrcha et al. (Reference Myrcha, Jadwiszczak, Tambussi, Noriega, Gazdzicki, Tatur and Del Valle2002) in their emended diagnosis of Palaeeudyptes gunnari. Several tarsometatarsi from the late Eocene of Antarctica were assigned to P. gunnari and P. klekowskii (Myrcha et al. Reference Myrcha, Jadwiszczak, Tambussi, Noriega, Gazdzicki, Tatur and Del Valle2002, Jadwiszczak Reference Jadwiszczak2006a) and a single incomplete humerus was determined as P. antarcticus (Tambussi et al. Reference Tambussi, Acosta Hospitaleche, Reguero and Marenssi2006, but see Jadwiszczak Reference Jadwiszczak2006b, p. 296). Additionally, many other fragmentary and eroded remains were assigned to Palaeeudyptes sp. in the above mentioned contributions.
A slight difference in size appears when the new material is compared with other Antarctic remains assigned to this species. MLP 96-I-6-13 is just a little bit smaller than other remains recently assigned to this species by Myrcha et al. (Reference Myrcha, Jadwiszczak, Tambussi, Noriega, Gazdzicki, Tatur and Del Valle2002) and Jadwiszczak (Reference Jadwiszczak2006a). However, these differences can be dismissed by considering the intra-specific size variation recorded in the metatarsus of modern (Acosta Hospitaleche & Gasparini Reference Acosta Hospitaleche and Gasparini2007) and Miocene penguins (Acosta Hospitaleche Reference Acosta Hospitaleche2004).
Once the precise systematic assignment has been established using tarsometatarsal characters, it is interesting to compare the humerus of this specimen to those assigned by other authors to the different Palaeeudyptes species.
The humerus under study exhibits all the characters of Palaeeudyptes and it is morphologically very similar to those assigned by previous authors (e.g. Jadwiszczak Reference Jadwiszczak2006a, Tambussi et al. Reference Tambussi, Acosta Hospitaleche, Reguero and Marenssi2006) to Palaeeudyptes gunnari. All of them have a sigmoid shaft and an undivided fossa tricipitalis. The head is proportionally robust, the condylus ventralis and dorsalis are robust and the extremitas distalis caudalis is projected beyond the edge of the dyaphysis.
This is not the first time that the discovery of an articulated skeleton expands the knowledge of the skeletal anatomy of a species previously known from isolated bones. Thus, it is possible to validate or not the allocations previously made from any counterparts prior to the discovery of articulated skeletons. In this contribution, the study of a partially articulated skeleton assigned (from the characters present in the tarsometatarsus) to Palaeeudyptes gunnari enabled the detailed description of other bones. Particularly, it was possible to study the morphology of the humerus, and re-evaluate previous assignments made by other authors from such remains. As in the case of Palaeospheniscus patagonicus (Moreno & Mercerat, 1891), systematic determinations made from morphological criteria such as robustness and size have been re-evaluated and accepted after analysing the complete skeleton (Acosta Hospitaleche et al. Reference Acosta Hospitaleche, Castro, Tambussi and Scasso2008). This implies that previous generic and sometimes specific allocations of these remains to known taxonomic schemes, made in a partially intuitive manner, such as the case of the determinations made by Wiman (Reference Wiman1905b) in his proposed size groups, are maintained after detailed analysis of additional osteologic characters.
Palaeeudyptes gunnari is a typical Antarctic penguin. This species is widely represented in Antarctica and is the most numerous in Argentine and Polish collections (Myrcha et al. Reference Myrcha, Jadwiszczak, Tambussi, Noriega, Gazdzicki, Tatur and Del Valle2002). This might be the reason for Palaeeudyptes having been repeatedly chosen by palaeoartists to represent the penguin fauna of Antarctica during Eocene times (e.g. Mikołuszko Reference Mikołuszko2007, Jadwiszczak Reference Jadwiszczak2009 and http://www.teara.govt.nz/en/penguins accessed October 2009).
The stratigraphic distribution of this species includes numerous remains coming from the Submeseta Allomember and a few others from the Middle Eocene Cucullaea I Allomember. Palaeeudyptes gunnari was probably a medium sized penguin, living exclusively in cool temperate settings during the Middle and Late Eocene in Isla Marambio. The highest levels (Priabonian, late Eocene, Submeseta Allomembers, c. 34–36 m.y.a.) document a major taxonomic and body size diversity with 14 species co-occurring sympatrically. The Eocene/Oligocene transition was characterized by sharp climatic deterioration linked to the progressive separation of South America and Antarctica and the strengthening of the Antarctic Circum Polar Current (Kennett Reference Kennett1977). The effect of these changes on the Antarctic marine fauna is unknown, but the disappearance (extinction?) of P. gunnari, together with several other Eocene penguins, e.g. Anthropornis nordenskjoeldi, broadly coincides with these events.
Acknowledgements
This is a contribution to the projects PICT 26219, PICT 499 and PICTA 1/2008. Instituto Antártico Argentino provided logistic support in Antarctica. The authors wish to thank Conicet for constant support and Paula Bona and Nadia Haidr from Museo de La Plata for their help in the osteological reconstruction. We also thank P. Jadwiszczak and S.A. Marenssi for suggestions on the manuscript.




