


INTRODUCTION
Changes in natural ecosystems are not easily predictable and legacies of previous land use and disturbances can persist for centuries, conditioning ecosystem responses (Foster et al. Reference Foster, Swanson, Aber, Burke, Brokaw, Tilman and Knapp2003). Recent studies on different biomes worldwide stressed that different trajectories of land use change strongly influence forest structure, diversity and composition (Foster Reference Foster1992; Hermy & Verheyen Reference Hermy and Verheyen2007; Walck et al. Reference Walck, Hidayati, Dixon, Thompson and Poschlod2011; Puerta-Piñero et al. Reference Puerta-Piñero, Espelta, Sánchez-Humanes, Rodrigo, Coll and Brotons2012). This paper further develops similar studies on dry tropical forest systems (Chazdon Reference Chazdon2003; Molina-Colón & Lugo Reference Molina-Colón and Lugo2006; Romero-Duque et al. Reference Romero-Duque, Jaramillo and Pérez-Jiménez2007; Álvarez-Yépiz et al. Reference Álvarez-Yépiz, Martínez-Yrízar, Búrquez and Lindquist2008; Molina-Colón et al. Reference Molina-Colón, Lugo and González Ramos2011), where ecological conservation and local livelihoods have been jointly exposed to land use changes (Chima et al. Reference Chima, Adedire, Aduradola and Agboola2012).
Tropical dry forests (hereafter TDF) cover up to 42% of tropical vegetation worldwide (Murphy & Lugo Reference Murphy, Lugo, Bullock, Mooney and Medina1995). Owing to their favourable climatic and edaphic conditions, TDF ecosystems have historically supported high human population densities and productive activities (Tosi & Voertman Reference Tosi and Voertman1964) and as a result TDFs are one of the most threatened major tropical ecosystems (Janzen Reference Janzen and Wilson1988; Bullock et al. Reference Bullock, Mooney and Medina1995; Maass Reference Maass, Bullock, Mooney and Medina1995; Quesada and Stoner Reference Quesada, Stoner, Frankie, Mata and Vinson2004; Hoekstra et al. Reference Hoekstra, Boucher, Ricketts and Roberts2005). According to Bawa et al. (Reference Bawa, Kress, Nadkarni and Lele2004), pressures over TDFs may further increase with economic globalization processes and climate change.
Despite the urgency for TDF conservation and management, this ecosystem has received only recent attention (Barreda-Bautista et al. Reference Barreda-Bautista, Lopez-Caloca, Couturier, Silvan-Cardenas and Carayannis2011). An increasing number of studies is being carried out in the dry Central America corridor (Griscom & Ashton Reference Griscom and Ashton2011; and specifically for Nicaraguan TDFs, see Sabogal Reference Sabogal1992; Sabogal & Valerio Reference Sabogal, Valerio, Dallmeier and Comiskey1998; Castro-Marín et al. Reference Castro-Marín, Nygard, Rivas and Oden2005; Gonzalez Rivas et al. Reference Gonzalez Rivas, Tigabu, Gerhardt, Castro-Marín and Oden2006; Tarrasón et al. Reference Tarrasón, Urrutia, Ravera, Herrera, Andrés and Espelta2010), demonstrating that, at present, TDF ecosystems have been reduced to fragmented patches covering less than 2% of their original area (Sabogal Reference Sabogal1992; Laurance & Bierregaard Reference Laurance and Bierregaard1997). However, the impacts of gradual long-lasting disturbance (namely chronic disturbance, as defined by Janzen Reference Janzen1986) versus acute disturbance events on vegetation structure and composition have been overlooked (Gillespie et al. Reference Gillespie, Grijalva and Farris2000).
This paper strengthens existing knowledge of TDF responses to different pressures in dry Central America corridor via a new perspective considering interdisciplinary dialogue between ecology and history, which may enable better-informed environmental management (Szabó Reference Szabó2010).
Additionally, this paper focuses on integrating scientific and local knowledge, and exploring human values and social perceptions, which may drive policy decisions (Foster et al. Reference Foster, Swanson, Aber, Burke, Brokaw, Tilman and Knapp2003). First, we assume that the involvement of local people in biodiversity assessment is justified, since conservation is a social goal that should voice opinions from all of society (Sheil & Lawrence Reference Sheil and Lawrence2004). Furthermore, the involvement of local people in the initial stages of selecting and prioritizing indicators encourages a more transparent decision-making process (Lawrence et al. Reference Lawrence, Paudel, Barnes and Malle2006; Lawrence Reference Lawrence2010). Second, what local people perceive and value as ‘important’ in a forest may not necessarily correspond to what ecologists identify as the most biologically diverse forest (Lawrence et al. Reference Lawrence, Paudel, Barnes and Malle2006). Indeed, the implementation of participatory or collaborative approaches for identifying and understanding local knowledge, values and key concerns on tropical forest conservation and management may improve quality of information and clarify opportunities for future intervention (Sheil et al. Reference Sheil, Puri, Wan, Basuki, van Heist, Liswanti, Rachmatika and Samsoedin2005). Participatory assessment methods can, however, be energy- and time-consuming. Indicators may reflect subjective evaluations of local people and be influenced by power relationships (Lawrence et al. Reference Lawrence, Paudel, Barnes and Malle2006). For this reason, we recommend the application of a two-way dialogue process for knowledge exchange, facilitated by scientists, in order to create a more robust assessment.
A previous attempt to involve local people and scientists in jointly evaluating TDF resources can be found in Tarrasón et al. (Reference Tarrasón, Urrutia, Ravera, Herrera, Andrés and Espelta2010). Recent approaches and methods in quantitative ethno-botany, however, suggest a need to distinguish between potential appreciation and concern for conservation (Hoffman & Gallaher Reference Hoffman and Gallaher2007). This research thus explored the factors behind prioritization of biodiversity conservation by local people, analysing whether ‘social importance’ reflects the value ascribed to TDF biodiversity by local communities, or simply local concern over the threat of loss and degradation of dry forest species and populations.
We sought to: (1) investigate whether and how the diversity, floristic composition and vegetation structure of TDF remnants differ according to historical land use change trajectories, and (2) integrate ecological and social information in a socioecological index that was evaluated in terms of its sensitivity to different land use trajectories.
We selected a protected area of dry northern Nicaragua for our research. During the second half of the 20th century, diverse forces have driven dry forest change in Nicaragua. From the 1950s onwards, land and natural resource use intensified significantly, encouraged by a growing population and land colonization programmes, expansion of the beef industry and agricultural modernization policies. As a result, TDFs experienced the most rapid and intense clearing (Kaimowitz Reference Kaimowitz1996). Ecological crisis, poverty gaps and social tensions increased during the 1970s, anticipating the regime shift towards a socialist system that implemented a cooperative management of production and natural resources. However, ten years later, the decline of beef prices and the economic crisis during the war promoted progressive land abandonment and vegetation recovery in some areas (Kaimowitz Reference Kaimowitz1996). In early 1990s, with peace restored and the failure of the cooperative system, a process of land reacquisition and degradation of natural resources strongly accelerated by the pressure of displaced people, the structural adjustment policies (Baumeister Reference Baumeister and Ghimire2001) and globalization (Kull et al. Reference Kull, Ibrahim and Meredith2007). Since the mid-1990s, marginal rural areas in the Central America region have been reconfigured as spaces for promoting conservation and management of remaining forest patches (Hecht et al. Reference Hecht, Kandel, Gomes, Cuellar and Rosa2006) through the creation of comanaged protected areas (Ravnborg Reference Ravnborg2008) and the introduction of payments for environmental services (Pagiola et al. Reference Pagiola, Ramirez, Gobbi, De Haan, Ibrahim, Murgueitio and Ruiz2007). Despite potentially inducing a local increase in social conflicts and marginalization, this new regulatory and institutional framework has been driving incipient pathways of forest recovery (Bray Reference Bray, Nagendra and Southworth2010).
METHODS
Study area and sampling
The study area is located in the Miraflor-Moropotente Terrestrial Protected Landscape (MMTPL; IUCN [International Union for Conservation of Nature] 1994) in the Nicaraguan department of Estelí (between 13° 19′ 30″–13° 60′ 30″ and 86° 11′ 00″–86° 22′ 00″).
The core area and the buffer zone cover an area of 290 km2 and consist of three ecological zones: dry lowlands (c. 30% of the territory), a semi-arid plateau (c. 25%), and mountainous cloud forests (c. 45%) (Tarrasón et al. Reference Tarrasón, Urrutia, Ravera, Herrera, Andrés and Espelta2010). Forest areas in the MMTPL occupy 11.3 % of the area (V. García-Milán & G. Moré, unpublished data 2009), and are characterized by high fragmentation and low connectivity between fragments (Do Carmo Correa Reference Do Carmo Correa2000). We selected three sites (mean 25730 km2) of TDF remnants within the dry and semi-arid zone (Fig. 1). The altitude in the study area ranges from 900 m to 1375 m above sea level. Mean annual temperature is 21°C and mean rainfall 800–1200 mm yr−1. Within the sites, we randomly sampled a total of 36 plots, each 0.1 ha in area. According to local people's accounts, all the selected plots were covered by old-growth dry forests prior to 1954, and are currently occupied by forest remnants.

Figure 1 Study location within the Miraflor-Moropotente Terrestrial Protected Landscape. The white line indicates the separation between mountainous cloud forests and dry and semi-arid forest zones (area of study). Sites: (1) the semi-arid plateau, (2) the dry southern lowlands, and (3) the dry eastern lowlands.
Land-use change analysis
We first analysed land-use change in the area by using black and white aerial photographs from the Instituto Nicaragüense de Estudios Territoriales (INETER) to compile land-use maps for the area for the years 1954, 1971, 1988 and 1996. The photographs were taken at scales between 1:25000 and 1:40000 in summer. Raw images were digitized and an ortho-photograph was created by assembling the various photographs of different parts of the area. Two principal methods of geometric correction were applied: first- and second-order polynomial functions, and orthorectification using ground control points. We used MiraMon GIS (Pons Reference Pons2005) to perform geometrical correction, photo-interpretation and digitization of the images. We first obtained a map of vectors from the interpretation, which we converted into raster format (5 m resolution).
The current forest cover classes were assigned by photo-interpretation of Google Earth images and verified during field work in March 2010. Based on the previous analysis and a pre-defined list of attributes, we identified the following land-cover categories: (1) agriculture (even texture, uniform colour, no trees, plough lines); (2) pasture and pasture with trees (dominated by grasses with sparse or no woody vegetation, < 30% tree cover); (3) shrub and secondary vegetation (areas covered by shrubs or tree saplings); (4) low-density forest (30–70% tree cover with uneven canopy); and (5) high-density forest (> 70% tree cover with even-structured canopy).
In order to check uncertainty and misclassification, two researchers were asked to independently and randomly photo-interpret subsets of 250 points in the study area per combination of year and land cover aspect, using 1000 bootstrap replications. The total area of each land-use class was calculated for each year and all land-cover maps were analysed to assess land-use change by using an overlay method.
According to Mena (Reference Mena2008), a land use change trajectory is the temporal sequence of land-cover classes at a location, described through a time series of classified aerial photographs. Similar to Álvarez-Yépiz et al. (Reference Álvarez-Yépiz, Martínez-Yrízar, Búrquez and Lindquist2008), we classified the main trajectories of land use change according to different intensities and frequencies of disturbance: (1) the acute-disturbance (AD) trajectory corresponds to a catastrophic intervention on the forest (such as clearing, conversion to cropland or pastureland) experienced between 1954 and 1996; (2) the chronic-disturbance (CD) trajectory corresponds to long-lasting high-intensity disturbance of the original old-growth forest through fire, grazing and selective thinning, from 1954 to the present, and (3) the low-disturbance (LD) trajectory corresponds to high-density forest, existing for at least the last 20–30 years, meaning that land forest has been maintained as old-growth forest from 1954 to the present (Appendix 1, Fig. S1, see supplementary material at Journals.cambridge.org/ENC). Twelve key informants were selected based on their recognition by local researchers and local people for their traditional ecological knowledge and historical expertise. We finally validated the trajectory of each plot with the key informants. This validation helped to avoid omissions and errors of classification, given the key informants’ knowledge of changes occurring in each plot over time.
Field sampling and ecological data analysis
Forest inventories were conducted in 2007 and 2010. In each plot, all species at the tree layer (dbh [diameter at breast height] ≥ 7.5 cm) were identified. We inventoried all trees and used the data to calculate tree density and basal area, species richness and Shannon-Wiener diversity index (H) and the Shannon species evenness measure (Eh). We also calculated the relative frequency and dominance per species for every land use change trajectory to explore differences in floristic composition. We tested the similarity of tree species composition within and among land-use trajectories by means of the Sørensen index.
Participatory evaluation of social value and concern over threat, and development of an integrated index
To explore the social importance of TDFs, we applied and refined the social simplified importance value index (SsIVI), as it summarizes both ecological indicators and local people's perceptions of the importance of conservation of the tree species present (Tarrasón et al. Reference Tarrasón, Urrutia, Ravera, Herrera, Andrés and Espelta2010). We performed a first series of semi-structured interviews (n = 82) for a preliminary free-listing, and, in 2007, a focus group with people aged between 50 and 89 years old was organized to investigate the history of land-use changes and species conservation. During a second series of field visits in 2010, we conducted in-depth interviews with the key informants (n = 12) based on the participatory methodology ‘walk-in-the-woods’ (Phillips & Gentry Reference Phillips and Gentry1993).
We triangulated the information from semi-structured interviews, participant observation and focus-group discussion, and interpreted data with the interdisciplinary staff, composed of western researchers and local technicians with long-standing experience in the area, in order to enable the identification of the multiple values associated with local species diversity. The biodiversity social values were subsequently grouped into four classes, and species were appraised for their direct or indirect use, their importance in specific ecological processes, and for intangible or existence values (cultural and aesthetic) (Appendix 1, Table S1, see supplementary material at Journals.cambridge.org/ENC). Key informants were later asked to rank the top ten species they would like to see in the local dry forests, and to express the associated values. The informants assigned a final score between 0 and 1 to each species (weighted by the number of informants).
Secondly, the interviewer showed key informants the historical series of aerial photographs, provided previously as support for the interview. They recognized the forest covers and used their historical knowledge to evaluate the degree of threat for each species, based on population size trends in the area. Next, the research staff translated and classified the linguistic expressions into the five degrees of threat adopted by the IUCN (2001): (1) present in the past, but already or almost extinct (endangered or critically endangered); (2) present in the past, but greatly declined over time (vulnerable); (3) more or less abundant than previously, but declined over time (between vulnerable and near threatened); (4) abundant and declined over time causing some concern (near threatened), or (5) very abundant and declined over time, but of least concern (least concern). To translate qualitative information with fuzzy nature by means of linguistic variables, we formally adopted the framework of fuzzy set theory (Munda Reference Munda1995). We validated the tolerance levels of their own evaluation with key informants.
To compare the forest remnants in terms of their conservation status, we modified the SsIVI index, recalculating this as the value SsIVI and the threat SsIVI by using the species prioritized after analysing social value and concern over threat:
 \begin{equation*}
{\rm SsIVI} = \Sigma \,w_i \, {\it SsIVI}_i = \Sigma \,w_i [(\% {\rm }N_i + \% G_i )/2]
\end{equation*}
\begin{equation*}
{\rm SsIVI} = \Sigma \,w_i \, {\it SsIVI}_i = \Sigma \,w_i [(\% {\rm }N_i + \% G_i )/2]
\end{equation*}
where %Ni is the percentage of individuals and Gi is the percentage in basal area of the target species, prioritized for (1) social value or (2) concern over threat. To calculate the SsIVI (concern over threat), we assigned a weight (wi ) in accordance with (1) the final score assumed by prioritized species for their social value and (2) the five degrees of threat.
Statistical analysis
We tested the pooled plot data by category of land use change trajectories for normality and constant variance. The influence of the land use change trajectories on the ecological variables (basal area, tree density and species richness) and the value and threat SsIVI indices was tested by means of a one-way nested ANOVA with land use change trajectories nested within site to account for differences among the three sites sampled in the study area. When the factor ‘land use change trajectory’ was significant, we applied Student Tukey-HSD post-hoc tests to perform pairwise comparisons of means among the different trajectories. Tree density, Shannon-Wiener H and Shannon Eh data were not normally distributed, thus for those variables, we used the Kruskal-Wallis non parametric ANOVA test. We performed all statistical analyses with Statistica version 6.0.
Assuming that geographical distance between plots and land use trajectories were the two possible explanatory variables of vegetation composition, we used a simple Mantel test to first verify whether community composition differences depended on distance between plots. We computed, using the statistical package R, the Euclidean distance matrices of all pair-wise distances among plots for species similarity (the Sørensen index), calculated according to the presence/absence and abundance data for all observed species and land-use trajectories. To recalculate the correlation between land-use trajectories and species composition, after checking the geographical distance between plots, a partial Mantel test was applied. The significance of simple and partial Mantel correlations between distance matrices was assigned by randomly permuting the order of elements within one matrix. This process was repeated 999 times for each analysis, and a correlation coefficient was calculated per iteration. A significant result indicates that the data are spatially autocorrelated. The Mantel and partial Mantel test were computed using the software package PASSAGE (version 2.0.11.6).
RESULTS
Land use change trajectories
Overall, forests cover decreased by c. 13% (between 1954 and 1971). Between 1971 and 1988, high- and low-density forested areas remained stable, while from 1988 to 1996 they increased by up to 26%, including forest patches that increased in area c. 8%. By exploring the trajectories of all plots included in the forest inventory, we found different combinations of changes in land use: 17% of plots showed low disturbance since 1954, while 44% exhibited signs of chronic disturbance. The rest of the plots (39%) corresponded to sites with acute intervention (Appendix 1 gives the type of uses associated with each category, see supplementary material at Journals.cambridge.org/ENC).
Forest structure, diversity and species composition
There were no significant differences in tree density or basal area between land-use trajectories: chronic and low-disturbance trajectories of land use showed similar values, while data for acute intervention sites showed high variability (Table 1). Species richness was significantly lower in forests that had experienced acute disturbance, especially in comparison with chronically disturbed sites (F = 2.4; df = 6; p < 0.05) Similarly, Shannon-Wiener H was significantly lower values in acute disturbed sites than in chronically or low disturbed sites (χ2 = 7.2; df = 2; p < 0.01). Sites with low or chronic disturbance trajectories not only had a greater number of species and diversity, but individuals in the community were distributed significantly more equitably (Shannon Eh) among all species (F = 2.7; df = 6; p < 0.01)
Table 1 Ecological characteristics (mean ± SE) of the sites according to the land-use change trajectories (AD = acute disturbance, CD = chronic disturbance, and LD = low disturbance). *Characteristics that differ significantly according to the ANOVA or Kruskal-Wallis tests. For the ANOVA tests, different letters indicate significant differences among trajectories as identified by Tukey-HSD tests (n = 36).
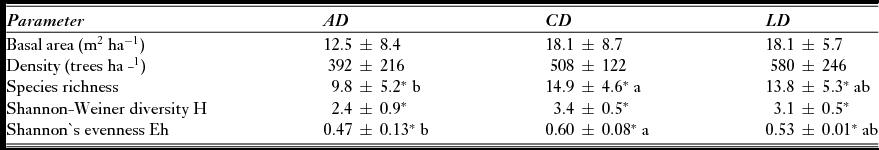
We found a total of 42 families, 77 genera and 89 species across all sites. All plots with an acute intervention trajectory were dominated in terms of frequency and abundance by species in the early stages of forest succession, common in degraded areas, for example c. 20% of the individuals were Acacia pennatula (Table 2). Only one species of high value and threat degree (Diphysa americana) was found exclusively in one acute disturbed trajectory (Appendix 1, Table S2, see supplementary material at Journals.cambridge.org/ENC). Bursera simaruba was the only tree species perceived as threatened among the five most frequent and abundant species in all land use change trajectories, even though it is only the dominant species in low disturbed sites. The plots with chronic disturbance trajectory showed the presence of various species of secondary vegetation among the most frequent and abundant. In contrast, low disturbed forests were more frequently and abundantly composed of species highly prioritized for their value and degree of threat (for more information see Appendix 1, Table S2, see supplementary material at Journals.cambridge.org/ENC).
Table 2 List of the five most frequent species for each land use change trajectory (AD = acute disturbance, CD = chronic disturbance, and LD = low disturbance) and their ecological attributes: RF = relative frequency; RA = relative abundance; H&E = habitat and ecology [sd (common in gallery semi-deciduous forest); d (common in deciduous forest), sv (common in secondary vegetation)]; Succ state = successional state [e (early), i (intermediate), or l (late)]; DA = common in degraded areas; DR = drought resistant; FR = fire resistant; Veg = vegetative propagation; Disp = secondary dispersal of seed [a (by ants), ca (by cattle), w (by wind), bi: (by birds), b (by bat), r (by rodents), and m (by monkey)]. Sources: Stevens et al. (Reference Stevens, Ulloa, Pool and Montiel2001), Tarrasón et al. (Reference Tarrasón, Urrutia, Ravera, Herrera, Andrés and Espelta2010), Kalacska et al. (Reference Kalacska, Sánchez-Azofeifa, Calvo-Alvarado, Quesada, Rivard and Janzen2004), Griscom & Ashton (Reference Griscom and Ashton2011), and our own personal observations and local knowledge.

Across all plots, simple Mantel tests revealed some spatial structuring of both the land use change trajectories and species composition. Plots that were closer together were more similar in species composition than they were to distant plots. The same correlation was also found between geographical distance between plots and land use trajectories. Nevertheless, after checking for the effects of geographical distance between plots, land use change trajectories still consistently explained most of the variation in the similarity of species composition among plots (r2 = 0.99, p = 0.001; Fig. 2), indicating that species composition has been influenced by history, and was therefore more similar among plots that had experienced the same land use change trajectory.

Figure 2 Simple Mantel paths between ‘space and tree species composition’ and ‘space and land-use trajectories’ and partial Mantel path between ‘land-use trajectories and tree species composition’ calculated for all plots in the area (n = 36).
Social simplified importance value indices (SsIVI) under different land-use change trajectories
Forty-one per cent of species inventoried in our plots were prioritized on account of their value. The four major categories recognized according to the information gathered by local people were related to: goods provision, such as timber and housing (62% of prioritized species), firewood use (21% of prioritized species) and extraction of non-timber products, such as medicines, fodder for domestic animals and fruits for human use (31% of prioritized species); indirect uses, such as trees for live fences and shade (50% of prioritized species); ecological values, such as pollination, refuge and fodder for wild fauna, epiphytes support and regulation services (50% of prioritized species); and existence values, such as aesthetic and cultural values (40% of prioritized species) (Appendix 1, Table S1, see supplementary material at Journals.cambridge.org/ENC).
Regarding the threat degree of forest species, a small percentage (7%) was not a cause for any concern. Certain species (11%) prioritized for their high degree of threat, were not listed as priority for their social value. Six species were prioritized by key informants as endangered or critically endangered, but they were not found in the plots (Cordia dentata, Ocotea veraguensis, Guaiacum sanctum, Swietenia humilis, Tabebuia rosea, Ruprechtia costata, and Bombacopsis quinata). Some of the threatened species are on the IUCN (2001) Red List (Appendix 1, Table S2, see supplementary material at Journals.cambridge.org/ENC).
Both the SsIVI (social value) and the SsIVI (concern over threat) were significantly sensitive to the land use change trajectories (Fig. 3), as both showed significantly higher values in low or chronically disturbed sites than in sites that had undergone acute disturbance (one-way ANOVA F = 2.85, p < 0.05 and F = 5.68, p < 0.001, respectively).

Figure 3 Mean ± SD of the SsIVI (V = value) and SsIVI (T = threat) for different land use change trajectories (AD = acute disturbance, CD = chronic disturbance, and LD = low disturbance). Differing letters indicate significant differences between trajectories identified by the Tukey-HSD tests (significant at p < 0.05).
DISCUSSION
Different land use change trajectories may influence species diversity, composition and social value prioritization, but not the structure of TDF remnants. While the study enhances knowledge on TDF ecology, the dialogue between local and scientific knowledge in evaluating the past and present status of TDFs may also facilitate development of more appropriate management and conservation guidelines.
Land-use trajectories and ecological forest response
The lack of differences in forest structure among contrasting land-use trajectories suggests that TDF species have very fast recovery ability after disturbance. As suggested by previous studies in Mesoamerican dry forests (Álvarez-Yépiz et al. Reference Álvarez-Yépiz, Martínez-Yrízar, Búrquez and Lindquist2008), in Central and Caribbean dry forest and rainforest (Guariguata et al. Reference Guariguata, Chazdon, Denslow, Dupuy and Anderson1997; Aide et al. Reference Aide, Zimmerman, Pascarella, Rivera and Marcano-Vega2000) and in Neotropical rainforests (Finegan Reference Finegan1996), the structural characteristics of early succession sites are similar to those of the oldest sites. This fast recovery may occur through resprouting (McLaren & McDonald Reference McLaren and McDonald2003; Vieira & Scariot Reference Vieira and Scariot2006; Peguero & Espelta Reference Peguero and Espelta2011) or seedling establishment, often after seed dispersal by cattle (Gillespie et al. Reference Gillespie, Grijalva and Farris2000; Peguero et al. Reference Peguero, Lanuza, Savé and Espelta2012). These results suggest that, despite forest structure being usually prioritized by scientists and managers when listing key functions of TDF (such as carbon sequestration), in itself it does not provide reliable information on land use change trajectories.
Interestingly, in addition to their similar structure, chronically disturbed and low disturbed (old-growth forest) sites had similar species richness, diversity and composition. The observed differences in species richness and diversity between long-term land-use trajectories with different intensities of disturbance (such as chronically versus acutely disturbed sites) corroborate purported differences persisting for a long period (Álvarez-Yepiz et al. Reference Álvarez-Yépiz, Martínez-Yrízar, Búrquez and Lindquist2008; Powers et al. Reference Powers, Becknell, Irving and Pérez-Aviles2009). Conversely, in sites with an acute disturbance trajectory, forest recovery appears a more complex process, and a simplification and homogenization of tree species is observed (see also Harvey et al. Reference Harvey, Villanueva, Villacís, Chacón, Munoz, López, Ibrahim, Taylor, Martínez, Navas, Sáenz, Sánchez, Medina, Vílchez, Hernández, Pérez, Ruiz, López, Lang, Kunth and Sinclair2005; Esquivel et al. Reference Esquivel, Harvey, Finegan, Casanoves and Skarpe2008; Wirth et al. Reference Wirth, Meyer, Leal and Tabarelli2008; Peguero & Espelta Reference Peguero and Espelta2011). In fact, as intensity of disturbance increases, a slower recovery in terms of species richness or diversity is expected due to changes such as soil compaction, propagule dispersal limitation (see Griscom & Ashton Reference Griscom and Ashton2011), disruptions of nutrient cycles (Lawrence et al. Reference Lawrence, D’Odorico, Diekmann, DeLonge, Das and Eaton2007), and invasion of pioneer species. Lugo (Reference Lugo1988) termed these delayed effects of degradation by acute disturbance a ‘time tax’ because organisms need time to overcome the impacts of degradation. Land use change trajectories were also partially responsible for the presence of common or rare species (evaluated in terms of relative frequency). Among other factors, management by farmers may also play a key role in the establishment of particular tree species. For instance, in chronically disturbed sites, singular trees (structural legacies) protected by local farmers for multiple purposes (from fruit production to aesthetic values) may ultimately act as nuclear foci to facilitate and accelerate the recovery of forests once the disturbance ceases (Janzen Reference Janzen and Wilson1988; Harvey & Haber Reference Harvey and Haber1999).
Usefulness of the integrated socioecological index in monitoring TDF changes
The integration of historical information, ecological data and social values, explored through participatory methods, illustrates how the importance that local communities ascribed to TDF is expressed indistinctly in terms of both the value associated with biodiversity and the concern over biodiversity threat.
Both of the integrated indices were sensitive to land use change trajectories. Forests in a chronic disturbance trajectory showed SsIVI results similar to forests in a low disturbance trajectory. This indicates a positive selection of valued and/or threatened species by farmers and confirms the ecological and social relevance of these remnants for future forest recovery, even after chronic disturbance. Conversely, the integrated indices’ values decreased as the degree of disturbance increased. As Tarrasón et al. (Reference Tarrasón, Urrutia, Ravera, Herrera, Andrés and Espelta2010) suggested, forest loss and degradation in these areas may be triggered by a ‘vicious cycle’ process: the more a forest system is degraded, the lower its social importance is, and thus the higher the chance of it being further degraded. This point is also confirmed by the absence of species considered to be valuable in acute disturbed plots (Appendix 1, Table S2, see supplementary material at Journals.cambridge.org/ENC).
Overall, these results suggest that the recovery of tropical forests may follow many distinct alternative paths, relating mainly to ecosystem resilience, landscape conditions and human disturbance (Holl & Aide Reference Holl and Aide2011). In contrast to ecological factors, knowledge of socioeconomic drivers is surprisingly limited in defining a predictable pattern for the recovery process (Lugo & Helmer Reference Lugo and Helmer2004; Lugo Reference Lugo2009). However, public understanding of the problem and the environment-human relationship, and social decisions on different management approaches are recognized as critical factors in successful conservation and management efforts (Molina-Colón et al. Reference Molina-Colón, Lugo and González Ramos2011). Undoubtedly, the ideologies and social interests of local people are factors that influence management priorities and, consequently, conditions for TDF recovery. Local farmers’ perceptions and behaviour shape local forests, influenced by the discourses around development versus conservation diffused by international agencies. Using farmers’ views and concerns to create indicators for biodiversity monitoring may help to increase the value they give to forest and their awareness of the implications of their behaviour (Lawrence et al. Reference Lawrence, Paudel, Barnes and Malle2006). Therefore, the social distinction between valued and threatened species observed in our study, provides both local people and policymakers with valuable information and a tool for monitoring changes in these ecosystems. It also gives them a means to devise a more integrated approach to TDF land use, conservation and management practices (Harvey et al. Reference Harvey, Villanueva, Esquivel, Gómez, Ibrahim, Lopez, Martinez, Muñoz, Restrepo, Saénz, Villacís and Sinclair2011).
Implications for TDF management and conservation
This local case may serve as an ideal representation of linkages between the historical patterns of policy and land use change, and the current conservation and recovery status of forests in the Central American Dry Corridor. Since the mid of the 1990s, within the scope of conservation policies, promising TDF regrowth has been observed across Central America. Examples of this process come from Costa Rica (Arroyo-Mora et al. Reference Arroyo-Mora, Sanchez-Azofeifa, Rivard, Calvo-Alvarado and Janzen2005; Calvo-Alvarado et al. Reference Calvo-Alvarado, McLennan, Sánchez-Azofeifa and Garvin2009), El Salvador (Hecht & Saatchi Reference Hecht and Saatchi2007), Honduras and Guatemala (Tucker et al. Reference Tucker, Munroe, Nagendra and Southworth2005), Panama (Sloan Reference Sloan2008) and across Mesoamerica and the Caribbean (Grau et al. Reference Grau, Aide, Zimmerman, Thomlinson, Helmer and Zou2003; Klooster Reference Klooster2003). These experiences of forest recovery associated with a change in the structure of society are examples of ‘land use transition’; they create a new set of conditions in which farmers take into account the costs and benefits of alternative land use and management (Lambin et al. Reference Lambin, Geist and Lepers2003). The present study provides two key messages for enhancing distinct pathways of sustainable management for the conservation and recovery of TDF remnants in Nicaragua, as well as in the Central America dry corridor. First, the low ecological and social value rank of acute disturbed forests suggests that recovery interventions should interact with rural development policies involving rural institutions and social actors to actively restore species diversity and composition and featuring new cultural agropastoral and forested landscapes. The final purpose may be to create a multi-stakeholder network of knowledge and experiences exchange along the corridor. Second, given their similarity and the urgency of action to preserve them, remaining areas of both chronically disturbed and undisturbed forests can act as important sources for the maintenance of tropical dry ecosystem diversity. Therefore, in this case, combined national and regional programmes and projects should focus on natural regeneration and conservation of these structural legacies which may act as sources of propagules.
Conservation policies are, however, just one of the structural dynamics contributing to forest trends. These forests are today exposed to new and more recurrently interlinked forces of change than before (such as climate and policy changes, demographic shifts and cultural changes). Data concerning historical trends and previous study of the likely climate and socioeconomic scenarios for the area (Ravera et al. Reference Ravera, Tarrasón and Simelton2011) indicate further research may help envisage how overlapping global forces such as environmental change and sudden economic collapse may have unexpected impacts on land use trajectories and, hence, pressure on future forest conservation status.
CONCLUSIONS
This interdisciplinary and participatory study explores how natural disturbance and human pressure on the remaining dry forests in a Central American protected area have affected ecological characteristics over time, and how local people expressed their views and concerns over forest conservation. Integrating history and ecology in forestry study, and using participatory biodiversity assessment, may help to inform decisions and support a ‘land-use transition’ towards the recovery of forest areas.
The social simplified importance value index, calculated via the integration of information on forest structure, species diversity and composition, with local people's perceptions on socially valued and threatened species, has provided an index that is sensitive to land-use trajectories. Our study suggests that there is a feedback loop between ecological conditions, social value and awareness of conservation that may influence forest recovery through positive selection, creating novel dry forests. For this reason, forestry assessment and conservation strategies should involve people who are engaged in local forest management in order to harness their ecological knowledge of sustainable use, and perceptions on ecological changes and local extinction.
However, given the continuing interaction of multiple and multi-scale drivers, the future pathways of novel dry forests after land abandonment are unpredictable. Further research to widen the scope of the SsIVI to other contexts in TDFs would allow better understanding of the similarities and differences in conservation and the conditions for the recovery of TDFs, providing greater insight into the interaction between concomitants factors of present conservation and future change.
ACKNOWLEDGEMENTS
We are very grateful to the local people from the Miraflor-Moropotente area who participated in this research. We specially acknowledge the local staff of the FAREM-Estelí (UNAN). We also thank three anonymous reviewers who provided useful comments. This study has benefited from discussion and information exchange in the IBERO-REDD+ network (P411RT0559-CYTED). Funding was provided by AECID project D/7592/07. Federica Ravera was funded by the Alliance for 4Universities.
Supplementary material
To view supplementary material for this article, please visit Journal.cambridge.org/ENC








