



Introduction
Rhenopyrgids are an unusual, morphologically distinctive group of edrioasteroids that are rare but both geographically and temporally wide ranging (Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013). They are known from all continents except Asia and Antarctica, from Lower Ordovician to Middle Devonian age rocks. Rhenopyrgids are morphologically highly organized and characterized by a small domed oral surface composed of five short ambulacra with large cover plates, separated by interradial oral plates, an elongated stalk-like pedunculate zone, and a basal coriaceous sac.
The history of taxonomic assignment of rhenopyrgid edrioasteroids is complex, but was recently well summarized by Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) and is not repeated here. However, Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) firmly established rhenopyrgids as edrioasterid edrioasteroids that are most closely related to other highly turreted edrioasterids (cyathocystids and edrioblastoids) based on similarities in the structure of the oral surface. The recent description of Heropyrgus from the Hereford Lagerstätte by Briggs et al. (Reference Briggs, Siveter, Siveter, Sutton and Rahman2017) has increased the number of genera assigned to the family Rhenopyrgidae Holloway and Jell to two. The key skeletal difference being that Heropyrgus apparently lacks cover plates and has floor plates that gape and thereby do not floor the entire ambulacra; a very unusual feature in edrioasteroids and indeed all echinoderms (Sumrall, personal communication, 2018).
Rhenopyrgids have been variously interpreted as living in mud burrows, into which the oral surface may be withdrawn by supposed contraction of the pedunculate zone (Smith and Jell, Reference Smith and Jell1990; Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008), or as mudstickers or encrusters with a flexible pedunculate zone (Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013). However, there is a lack of direct evidence for any behavior; none have been found in-situ, either within burrows, affixed to muddy substrates, or encrusting hard or firm substrates.
New material presented here from the Silurian of Canada and various European localities (Ordovician of Girvan, Argyllshire, UK, and Montagne Noire, France) extends the paleogeographical and temporal distribution of this rare fossil group. The new material provides an insight into previously overlooked morphological variability and taphonomic observations, which in turn have led to new paleoecological inferences.
Geological setting
New specimens included in this study were collected from the lower Silurian of Anticosti Island, Québec, Canada (Fig. 1.1–1.3), and the Ordovician of Montagne Noire, France and the Girvan district of Scotland. The geological setting of each of these localities will be dealt with in turn, although only the newly erected species from Anticosti Island will be dealt with in detail.
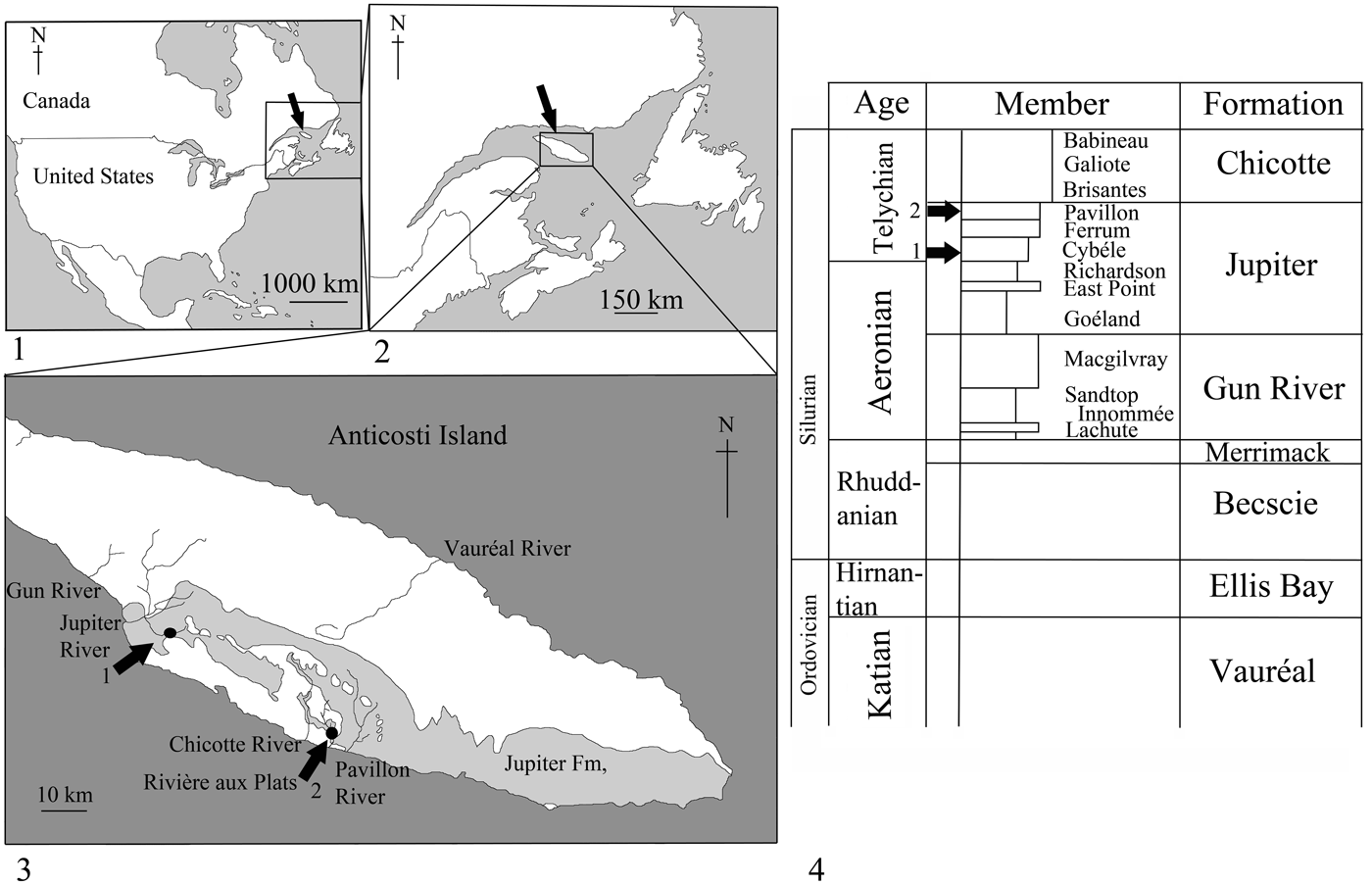
Figure 1. (1–4) Location and geology of Anticosti Island. (3) Map showing locations of collection sites on Anticosti Island: 1, Rock Pool Ledge, Jupiter River; 2, Pavillon River Section. (4) Stratigraphic chart of Anticosti Island; arrows show positions of rhenopyrgid fossils.
Anticosti Island
The geology of Anticosti Island comprises Upper Ordovician to lower Silurian strata (Chatterton and Ludvigsen, Reference Chatterton and Ludvigsen2004) that dips gently to the southeast (see Fig. 1.4). The units are dominated by shallow marine carbonate ramp deposits of limestones and shales (Long, Reference Long2007). During deposition of these rocks, the southern margin of Anticosti was most likely exposed to westerly directed tropical cyclones off the Iapetus Ocean, which deposited frequent tempestites (Wilde, Reference Wilde, Barnes and Williams1991). The rhenopyrgid edrioasteroids were found in two members of the lower Silurian (Telychian) Jupiter Formation. Almost all specimens of R. viviani n. sp. came from the Cybèle Member at collection site 1, but a single specimen came from the Pavillon Member at collection site 2 (see Fig 1.4). Within the published literature, the only other edrioasteroid known from Anticosti Island is Isorophusella pleiadae (Sinclair and Bolton, Reference Sinclair and Bolton1965) from the Upper Ordovician Vauréal Formation (Sinclair and Bolton, Reference Sinclair and Bolton1965; Bell, Reference Bell1976). Unpublished edrioasteroid material has been collected more recently, but none of this included rhenopyrgid edrioasteroids (W. Ausich personal communication, 2018).
The Cybèle Member comprises 32 m of gray, sparsely fossiliferous, lithographic limestone deposited on a broad, south-facing carbonate ramp. Fossils are relatively scarce (M. Martin, personal observation, 2015), but dominated by trilobites and highly endemic crinoids (Ausich and Copper, Reference Ausich and Copper2010). The specimens all came from a hard, medium-gray, tempestite limestone. Other fossils include current orientated tentaculitids, disarticulated trilobite fragments, and ostracod debris.
The Pavillon Member is composed of 10 m of thin recessive tempestite limestones that are interbedded within gray/green shales. The member is generally highly fossiliferous and rich in brachiopods and trilobites. Other fossils include large numbers of isolated crinoid columnals and several fairly long partially articulated stem fragments, along with disarticulated trilobite remains, bryozoan fragments, and numerous tentaculitids.
The specimens from Anticosti Island are very well preserved in calcite with some evidence of stereom microstructure present (Fig. 2.5, 2.10). Specimens are preserved either lying on the bedding plane and apparently similarly orientated to other tubular fossils (e.g., tentaculites) (Fig. 2.8), or nearly perpendicular to the bedding (such as Figs. 2.1, 4). All are preserved in a hard gray micritic limestone. Most specimens display partial crushing, but retain some three-dimensionality. The oral surface is usually partially collapsed, with disrupted cover plates (Figs. 2.1–2.3, 6, 7, 9), while others show disarticulation of the suboral constriction and pedunculate zone (Fig. 2.5). The specimens that are preserved perpendicular to bedding surface also suffer either minor crushing or have flattened oral areas (Figs 2.1–2.4) and loss of the distal pedunculated zone and coriaceous sac. None appears to be in life position or within a burrow. All specimens from the Cybèle Member are partially disarticulated and missing parts of the distal pyrgate zone and coriaceous sac. This suggests, in both members, short transportation with preservation occurring after rapid burial.

Figure 2. Rhenopyrgus viviani n. sp. from Jupiter Formation, Telychian of Anticosti Island, Québec, Canada. (1–9) From Cybéle Member, Rock Pool Ledge site on Jupiter River (1–3) (paratype MPEP 1126.1); (1) lateral view of entire specimen (note it is preserved perpendicular to bedding with flattened oral surface but undisturbed pyrgate zone); (2) detail of oral surface; white arrows point to ridges on adambulacral margin of the oral ossicles, which accommodated cover plates; black arrow points to the ridged ambulacral floor that is presumably an extension of the interadial oral ossicle (coated with ammonium chloride); (3) oral surface without ammonium chloride; again, black arrow points to ridged ambulacral floor; (4) oral view of cover plates showing vestige of 2-1-2 ambulacral arrangement (paratype EE 15752); (5) lateral view, note lack of any floor plate sutures along adambulacral margin of oral plates (paratype EE 15753). (6–8) Holotype (EE 16642); (6) note changing morphology of pyrgate zone ossicles; (7) detail of oral surface and sub-oral constriction; black arrow points to anal pyramid composed of rod-like ossicles; white arrow points to triangular cross section of cover plates; (8) holotype slab showing two individuals (holotype on right). (9) Lateral view with a well-preserved suboral constriction; note a change in morphology of pyrgate zone plates (paratype EE 15754). (10) From Pavillon Member at Rivière aux Plats; distal pyrgate zone and bulbous coriaceous sack-like holdfast comprised of small granular plates (paratype EE 15756). Abbreviations: CL = collar plates, CP = cover plates, O = oral plate. All scale bars represent 1 mm.
Ardmillan specimen (Girvan)
The precise locality of the specimen EE 16524 is simply labelled “Ardmillan” and does not have any further specific detail. Although it is different from the well-known Lady Burn Starfish Bed, it is reminiscent of equivalent rock of the Ardmillan Series of the Upper Ordovician (Hirnantian) exposed on the coast to the south of Girvan, Scotland (in the vicinity of a camp site called ‘Ardmillan’). It therefore seems likely to have come from this locality.
The fossil was preserved in an oval concretion comprised of a massive fine, dark gray, muddy sandstone, including numerous larger clasts of detrital calcite. Unusually for Paleozoic echinoderms, EE 16524 is preserved as a three-dimensional calcite cast (secondary calcite overgrowing primary stereom) composed of a long articulated pyrgate zone missing the adoral structures and basal coriaceous sac (Fig. 3.5). Thus, the imperfect preservation of the specimen and the massive nature of the sediment again suggest short transportation and rapid burial in a mass accumulation.
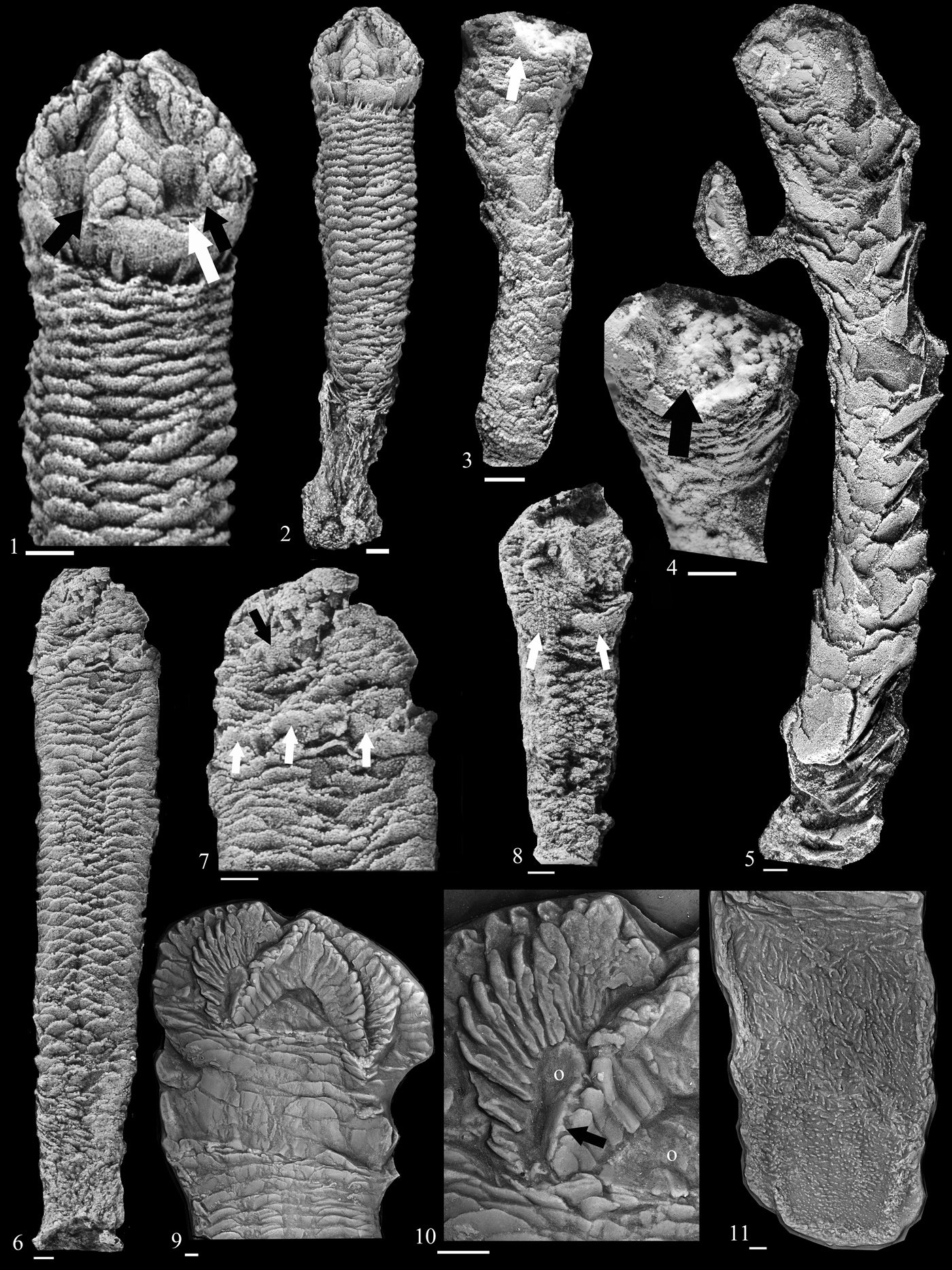
Figure 3. Rhenopyrgidae; all whitened with ammonium chloride. (1, 2) Rhenopyrgus grayae, Upper Ordovician, Lady Burn Starfish Bed, Girvan, Ayrshire, Scotland (holotype E23470): (1) details of oral surface with prominent collar plates and no suboral constriction apparent; black arrows highlight exposed floor plates in distal ambulacra; white arrow points to a small, exposed part of a plate of the suboral constriction, suggesting that suboral constriction could be contracted behind collar plates; (2) lateral view. (3, 4) Rhenopyrgus indet. 1, Silurian, Newland Formation, Newlands, Ayrshire, Scotland (E 62753): (3) lateral view of pyrgate zone, arrow points to enlarged plates interpreted here as collar plates; (4) oral view, black arrow highlights disarticulated large D-shaped oral ossicle. (5) Rhenopyrgus indet. 2, Ordovician, Drummuck Series, Ardmillan, Girvan District, Ayrshire, Scotland (EE 16254); lateral view of pyrgate zone; note difference in size and morphology of the pyrgate ossicles suggesting it is different than R. grayae, which is found in similar age rocks that are geographically close; also note disarticulated ridged deltoid-shaped plate closely associated with articulated pyrgate plates. (6–8) Rhenopyrgus indet. 3: (6, 7) Foulon Formation (middle Floian), La Croix de Roquebrun, Saint-Nazaire-de-Ladarez, Hérault, France (UCBL-FSL 713312); (6) lateral view of whole specimen; (7) enlargement of the oral surface, showing confused plate articulation of this region; black arrow highlights possible oral ossicle; white arrows highlight collar plate series; (8) late Tremadocian, beneath Saint-Chinian Formation, Saint-Chinian, SW of Donnadieu, Babeau-Bouldoux, Hérault, France (UCBL-FSL 713316), lateral view. (9–11) Rhenopyrgus coronaeformis Rievers, Reference Rievers1961, Lower Devonian, Emsian, Hunsrück Slate, Bavaria, Germany (holotype SNSB-BSPG 1958 XV 50): (9) detail of oral surface and proximal structures; (10) detail of oral surface; note complicated cover plate articulation surfaces; black arrow highlights grooved adambulacral margin of oral plate; (11) detail of coriaceous sac. Abbreviations, O = oral plate. All scale bars represent 1 mm.
Montagne Noire specimens
The Southern Montagne Noire (France) exhibits a rather complete Lower Ordovician succession. Rhenopyrgid material has been collected from the Saint-Chinian, Foulon, and Landeyran formations.
The Saint-Chinian Formation (<500 m thick) consists of monotonous dark-gray clayey siltstones intercalated by fine- to medium-grained sandstones, interpreted as outer-platform deposits (Álvaro et al., Reference Álvaro, González-Gómez and Vizcaïno2003). The specimen studied (UCBL-FSL713316) here was collected from the E. filacovi Zone (late Tremadocian) and is preserved as a mold within a gray calcareous concretion in three dimensions. The sediment is a fine-grained siltstone.
The Foulon Formation (80 m thick) corresponds to an alternation of shales, bearing siliceous fossiliferous concretions, and fine-grained sandstones deposited via storm-generated turbidity currents that transported sandy material to offshore settings (Noffke and Nitsch, Reference Noffke and Nitsch1994). The specimen studied here (UCBL-FSL713312) was collected from the transitional beds at the uppermost part of the formation at the top of the N. arenosus Zone (middle Floian) and is preserved as a crushed mold within a gray shale (i.e., not within a concretion) (see Fig. 3.6, 3.7).
The Landeyran Formation (~300 m thick) is composed of brown, green, and gray homogeneous shales bearing siliceous concretions, interpreted as bioclastic storm deposits in upper offshore environments (Bonin et al., Reference Bonin, Nardin, Vennin, Vizcaïno, Vennin, Aretz, Boulvain and Munnecke2007). The specimens studied (UCBL-FSL 713313–713315) were all collected from the A. incisus Zone and are preserved both as a mold within a gray calcareous concretion in three dimensions (UCBL-FSL 713315) and as a flattened mold within a gray shale (UCBL-FSL 713313, 713314).
Materials and methods
The specimens new to science that were used in this study come from several locations. These are listed below:
Anticosti material
EE 16642 (holotype), EE 15752–15755 (paratypes), MPEP 1126.1 (paratype) all from Rock Pool Ledge, Jupiter River Section, Cybèle Member (Loc. 1 on Fig. 1.3) and EE 15756 (paratype), from the Pavillon River section, Pavillon Member (Loc. 2 on Fig. 1.3), Anticosti Island, Québec, Canada (Fig. 1). Specimens were found by surface collecting in 06/2011 by T. Vivian, M. Martin, and P. Isotalo.
Ardmillan material
EE 16254; “Ardmillan,” South of Girvan, Girvan District, Ayrshire, Scotland. Method of collection unknown. Found unregistered within the collections of the Natural History Museum London.
Montagne Noire material
UCBL-FSL 713312; La Croix de Roquebrun, Saint-Nazaire-de-Ladarez, Hérault; UCBL-FSL 713313–713314; Pont Supérieur, Saint-Nazaire-de-Ladarez, Hérault; UCBL-FSL713315, Vallée du Landeyran, UCBL-FSL 713316; SW of Donnadieu, Babeau-Bouldoux, Hérault. Specimens were collected by D. Vizcaïno and E. Monceret by surface collection and splitting of concretions.
Methods
Matrix surrounding the specimens from Anticosti Island was initially removed using an ME-9100 and a Micro Jack #4 air scribe and a Crystal Mark MV-241 Micro air abrasion unit with 40 μm dolomite powder blast media with final uncovering of plates done using a hardened steel needle so as to minimize damage to the surface texture/structure of the plates. Moldic specimens (from various sites) were cast in latex. All specimens or latex casts were then coated with ammonium chloride and imaged using a Canon SLR 340 digital camera.
An attempt was made to scan the oral surface of one Anticosti Island specimen (EE 15755) at the CERN light source, Switzerland to understand the internal structure. However, this proved to be unsuccessful owing to insufficient contrast between the matrix and fossil.
Rhenopyrgid morphological terminology follows that of Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013; see also Fig. 4.1). The term “pyrgate zone” is here used to refer only to the highly imbricated, scale-like plates distal to the collar plate circlet and proximal to the coriaceous sac; it differs from the term “pedunculated zone,” which includes these other structures.
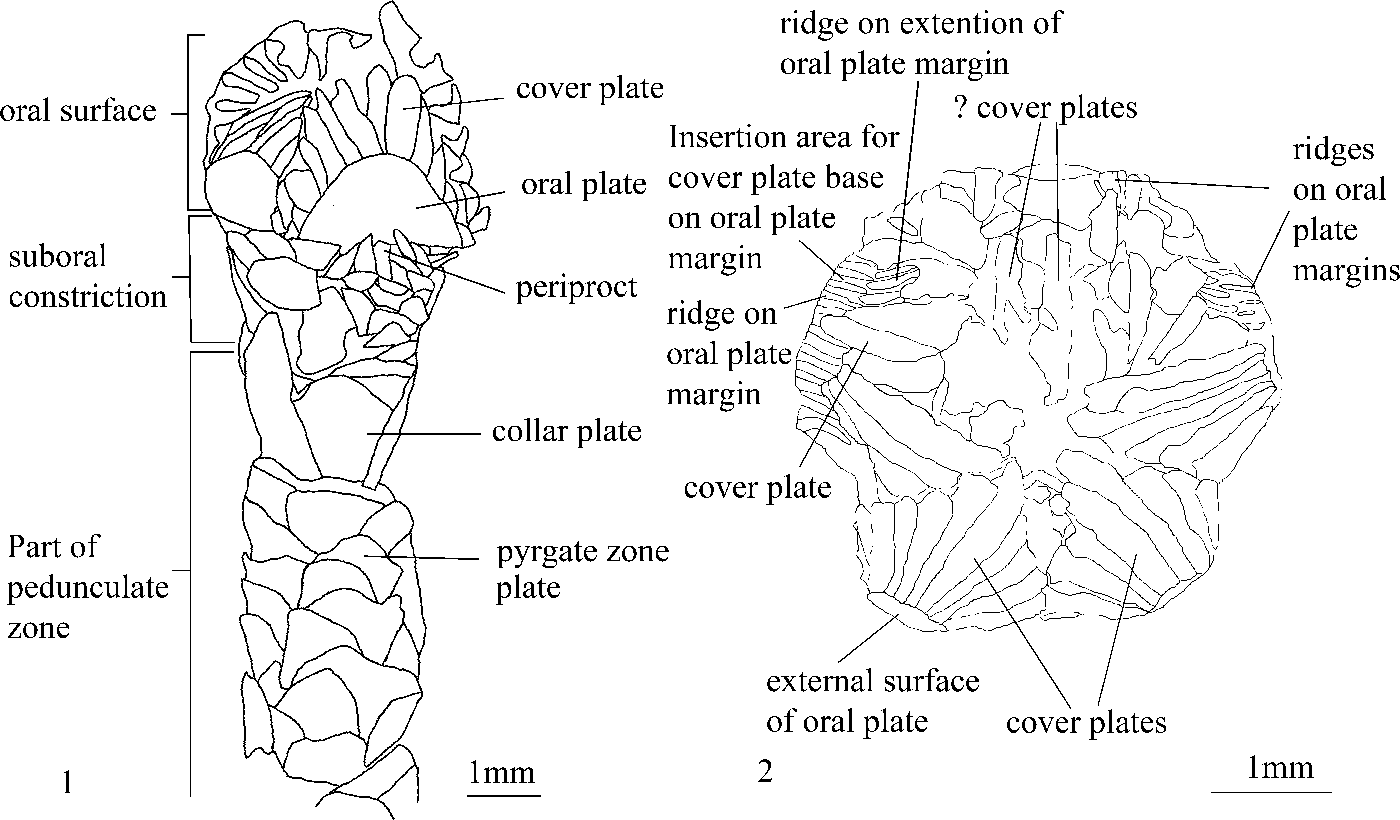
Figure 4. Rhenopyrgus viviani n. sp. Silurian (lower Telychian), Jupiter Formation, Jupiter River, Anticosti Island, Canada. (1) Camera lucida of lateral CD ambulacral of holotype of Rhenopyrgus viviani n. sp. (EE 16642); note periproct formed of small lath-shaped plates adjacent to edge of oral plate; also note lack of floor plates exposed along adambulacral margin of oral plate; (2) camera lucida view of oral surface of paratype MPEP 1126.1; note groove and ridge arrangement along adambulacral margin of oral plate that accommodated and held in place cover plates; also note ridged surface of fused floor plates, which may have accommodated tube feet.
Repositories and institutional abbreviations
Specimen prefixes are housed in the following institutions: E or EE are abbreviations of the full prefix NHMUK PI E/EE: Natural History Museum London, UK. MPEP: Musée de paléontologie et de l’évolution, Québec, Canada. SNSB-BSPG: Bayerische Staatssammlung für Paläontologie und Geologie, Germany. UCBL-FSL: University Claude Bernard 1, Lyon, France. USNM: National Museum of Natural History (Smithsonian Institution), Washington DC, USA.
Systematic paleontology
Phylum Echinodermata de Bruguière, Reference Bruguière and de1791 (ex. Klein, Reference Klein1734)
Class Edrioasteroidea Billings, Reference Billings1858
Order Edrioasterida Bell, Reference Bell1976
Suborder Edrioblastoidina Fay, Reference Fay1962
Family Rhenopyrgidae Holloway and Jell, Reference Holloway and Jell1983
Genera included
Rhenopyrgus Dehm, Reference Dehm1961; Heropyrgus Briggs et al., Reference Briggs, Siveter, Siveter, Sutton and Rahman2017.
Diagnosis (Emended)
Pyrgate edrioasteroids with relatively small oral surfaces bearing five, short, straight rapidly tapering ambulacra; interambulacra formed of single tessellate interradial oral plates and biserial and alternating floor plates that may or may not be fused; cover plates, where present, are biserial and tall proximally; sub oral constriction generally short, composed of circlets of poorly organized imbricate, scale-like plates and periproct, sometimes withdrawn behind collar plates; collar plates large, forming a single imbricate circlet at base of suboral constriction; pyrgate zone comprised of highly imbricated scale-like plates often in eight rows arranged in alternating circlets of four; holdfast formed of coriaceous sac comprised of small unorganized platelets.
Occurrence
North Africa (Morocco), Europe (Germany, UK, France, Russia, Sweden, ?Spain), North and South America. Tremadocian, Lower Ordovician to Eifelian, Middle Devonian.
Remarks
The description of R. viviani n. sp. and a detailed reassessment of previously described taxa (see discussion) with well-preserved oral surfaces have highlighted varying degrees of fusion of the interadial oral plate and the ambulacral floor plates (particularly the externally exposed parts). The ambulacral floor plates of the type species, Rhenopyrgus coronaeformis Rievers, Reference Rievers1961, have totally fused to each other and the interradial oral plate (Fig. 3.9, 3.10), while plate sutures are visible in Rhenopyrgus grayae Bather (Fig. 3.1). This highlights greater structural diversity than was previously thought, which, as well as broadening the familial diagnosis, also enables a re-evaluation of the relationships with other edrioasterids that have a similar oral arrangement (see below).
The discovery of a well-preserved anal pyramid in the holotype of R. viviani n. sp. confirms the suggestion of Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) and Holloway and Jell (Reference Holloway and Jell1983) that the periproct is located within the suboral constriction, adjacent to an oral plate in the CD interray. This location is similar to that seen in other edrioasterids (Bell, Reference Bell1976) and Heropyrgus (Briggs et al., Reference Briggs, Siveter, Siveter, Sutton and Rahman2017).
Briggs et al. (Reference Briggs, Siveter, Siveter, Sutton and Rahman2017) recently erected Heropyrgus within the Rhenopyrgidae because it shared many key characteristics of skeletal organization and differed only by lacking cover plates and that the floor plates gaped and did not completely floor the ambulacra, although these variations were not included in an emended diagnosis. However, the lack of cover plates and the unusual arrangement of the ambulacral floor are very curious, particularly because the ridged adambulacra margin of the oral and floor plates look very similar to those of other rhenopyrgid genera that clearly bear cover plates (particularly R. viviani n. sp.; Fig. 2.2). As such, taphonomic loss and/or disruption should be considered a possibility for cover plates lacking in specimens assigned to Heropyrgus and potentially the ambulacral gape. That said, in lieu of further evidence, this genus must currently be maintained and its variations included in the family diagnosis.
Following Holloway and Jell (Reference Holloway and Jell1983), Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) placed rhenopyrgid species into their own family; however, they questioned the validity of classifying specimens based solely on characters of the pyrgate and pedunculate zones or where they lacked well-preserved oral areas. As such, Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) removed all such examples from the genus Rhenopyrgus, but retained them within the Rhenopyrgidae Holloway and Jell (Reference Holloway and Jell1983) on account of the pyrgate zone being formed of eight rows of plates. We follow this approach here.
Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) also suggested the removal of generic assignation of rhenopyrgid taxa that lacked a suboral constriction, thereby suggesting Rhenopyrgus grayae (Bather, Reference Bather1915) and possibly R. flos Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008 should be placed in a separate genus. However, we believe that the lack of a sub-oral constriction is taphonomic in rhenopyrgids (see discussion section). As such, we retain both these species within Rhenopyrgus, which also include taxa with clearly preserved suboral constrictions.
The higher taxonomic relationship of rhenopyrgids was reviewed by Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) who, in agreement with previous workers, suggested that rhenopyrgids are edrioasterids united with edrioblastoids and cyathocystoids by the reduced number of interambulacral plates, the pseudo-fivefold symmetry of the ambulacral system, and the turreted thecal shape. These groups differ from one another primarily in the form of the pedunculate zone—organized flexible stalk in rhenopyrgids; fused cup in cyathocystids; and an elongate, rigid, multiplated stem in edrioblastoids. However, Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) left the specific relationships between these families open. Sprinkle and Sumrall (Reference Sprinkle and Sumrall2015) drew closer affinities between cyathocystids and derived edrioasterids because both shared a similar oral surface construction, formed by fusion of the floor plates to each other and the interadial oral plate. However, these comparisons were made on the assumption that all rhenopyrgid taxa did not possess this feature, which we here demonstrate is not the case. We cannot really add anything further to refine these relationships other than highlight that all three turreted edrioasterid families display similar ambulacral structures in various constituent species. As indicated by Sprinkle and Sumrall (Reference Sprinkle and Sumrall2015) for cyathocystids and derived edrioblastoids, this suggests some form of homoplasy/convergence, which was particularly suitable for turreted edrioasterid ecology.
Genus Rhenopyrgus Dehm, Reference Dehm1961
Type species
Pyrgocystis (Rhenopyrgus) coronaeformis Rievers, Reference Rievers1961; Hunsrück Slate, Lower Devonian, (Emsian) of Germany.
Other species
Rhenopyrgus coronaeformis Rievers, Reference Rievers1961; R. flos Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008; R. viviani n. sp.; R. sp. indet.1, formerly Pyrgocystis procera (Aurivillius) Bather, Reference Bather1915; R. sp. indet. 2, R. sp. indet. 3, R. sp. indet. 4.; R. grayae (Bather, Reference Bather1915); R. whitei Holloway and Jell, Reference Holloway and Jell1983; and R. piojoensis Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013.
Emended diagnosis
Rhenopyrgids with fused or unfused ambulacral floor and interradial oral plates; suboral constriction, formed of a zone of highly imbricate elongate plates formed between the oral surface and suboral collar, may or may not be exposed. Cover plates present. Pyrgate zone formed of eight rows of imbricated plates arranged in two circlets of four.
Occurrence
Lower Ordovician (Tremadocian) to Lower Devonian (Emsian); from France (Lower Ordovician, upper Tremadocian and Floian), UK (Katian, Upper Ordovician, and middle Llandovery, lower Silurian), Canada (Rhuddanian, lower Silurian), Argentina (lower Ludlow, upper Silurian), Australia (Lower Devonian), Morocco (Emsian, Lower Devonian), and Germany (Emsian, Lower Devonian).
Remarks
There is significant variation in the fusion of the interradial oral plates with the ambulacral floor plates in different species throughout the range of Rhenopyrgus. Some (e.g., Rhenopyrgus coronaeformis, R. flos, and R. viviani n. sp.) display complete fusion, giving the appearance of only a single D-shaped interradial plate with a broad curved ambital margin. It is tempting to use this character to subdivide the genus further, however there are few other characters that either unify or differentiate these taxa from those with unfused (or less completely fused) oral surfaces. As such, until detailed cladistic analyses are employed, we withhold from formerly erecting a new genus for these taxa with unfused floor plates (because the type species R. coronaeformis clearly displays fused interaidal and ambulacral floor plate elements; see Fig. 3.9, 3.10).
The fusion of the interradial and floor plate systems, however, means that isolated D-shaped oral plates, with either smooth or notched margins, that are found in close association with articulated sections of pyrgate zone comprised of eight rows of plates, which may or may not also lack clearly defined collar and suboral constriction plates (because these plates are determined to be moveable and therefore could have contracted within the pyrgate zone and be obscured, e.g., Fig. 2.3, 2.4), can now be used to imply classification to Rhenopyrgus. Owing to the presence of such associations, Rhenopyrgus sp. indet. 1, formerly Pyrgocystis procera (Aurivillius) Bather, Reference Bather1915, from the lower Silurian Newlands Beds of Newlands, Girvan, Ayrshire, Scotland (Fig. 3.3, 3.4); Rhenopyrgus sp. indet. 2 from the Ordovician of Ardmillan, Girvan, Ayshire, Scotland (see below); Rhenopyrgus sp. indet. 3 from the Montagne Noire, France (see below); R. flos (Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008) from the Devonian of Morocco; and Rhenopyrgus sp. indet. 4 (as illustrated by Grigo [Reference Grigo2000]) can all be assigned to this genus.
Holotype
EE 16642, paratypes EE 15752–15756 and MPEP 1126.1. Cybèle Member, Jupiter Formation, Telychian, upper Llandovery, lower Silurian from Rock Pool Ledge, Jupiter River, Anticosti Island, Québec, Canada.
Diagnosis
Rhenopyrgus with interambulacra composed of a single broad, relatively low D-shaped compound interadial plate with gently curving adambulacral margin. Distal cover plates distinctly square. Adoral constriction composed of 3–5 circlets. Plates of the pedunculate zone vary in morphology of exposed aboral margin, being curved adorally, but becoming distinctly rhombic (angular) aborally.
Occurrence
Cybèle Member to Pavillon Member, Jupiter Formation, Telychian, upper Llandovery, lower Silurian, Anticosti Island, Québec, Canada (see above for details).
Description
Oral surface domed with straight ambulacra, fairly evenly spaced, but with a vestige of 2-1-2 ambulacral arrangement (Sprinkle, Reference Sprinkle1973) apparent (Fig. 2.2–2.4). External oral surface composed of biserial, alternating, tessellated cover plates with zig-zag perradial suture and tessellated, D-shaped, compound interradial plates (formed of fused interradial oral plate and ambulacral floor plates) lacking in obvious sutures (in un-weathered specimens) and with curved adambulacral margins. Cover plates number 12–16 per interradial oral plate, 6–8 in each ambulacrum (Fig. 2.2–2.7). Proximal cover plates long thin rectangles with triangular or T-shaped cross sections (Fig. 2.7, 2.9), but grade distally into shorter, wider, more equi-dimensional (square) plates (Fig. 2.4, 2.5, 2.9). All cover plates articulate in grooves formed by ridges along the margin of the compound interradial plates (Figs. 2.2, 2.3, 4.2). Below cover plate articulation areas, the ridges anastomose or become off-set, forming a distinct set of ridges. Ambulacral floor not adequately seen in any specimen, but proximally appears to be formed by the shallow, ridged slope of the adambulacral margin of the compound interradial oral plate. Distally, floor plates appear to have a similar morphology (Figs. 2.2, 2.3, 4.2). Aboral margin of interradial oral plates straight and abut along oral surface ambitus with imbricate plates of suboral constriction zone.
Suboral constriction composed of at least 3–5 circlets of less-regularly aligned, highly imbricating, wedge-shaped plates that increase in size distally, but which are comparatively small compared to those of collar and pyrgate zone (Fig. 2.5–2.7, 2.9). Distally, these circlets are bordered by a circlet of larger, rectangular imbricated plates that form the collar (Fig. 2.5–2.7, 2.9). Periproct (Figs. 2.7, 4.1) located in CD interray adjacent to oral plate margin, composed of a single circlet of short imbricated rectangular plates. Gonopore and hydropore unknown.
Pyrgate zone highly organized into alternating circlets of four, highly proximally imbricating plates. Plates form alternate circlets aligned into straight longitudinal columns of at least 52 (in the only known specimen with a complete pedunculate zone) such that eight peduncular ridges are present. Four plates from each circlet meet beneath the more distal circlet to form a ring with a fairly large lumen. Plates are squamose proximally with broadly rounded outline becoming distinctly more angular distally in the holotype and smaller specimens (Fig. 2.6, 2.9), but variation less apparent in large individuals (e.g., in EE 15756). Coriaceous sac bulbous at base, approximately a quarter of length of pedunculate zone and slightly wider. It is formed of numerous irregularly placed platelets (Fig. 2.10).
Etymology
Named for the initial discoverer Mr. Travis Vivian.
Materials
EE 16642 (holotype), EE 15752–15756 (paratypes), MPEP 1126.1 (paratype), MPEP1143.1.
Remarks
This taxon is differentiated from most other species of Rhenopygus by the fusion of interradial oral plates and the external portions of the floor plates, thereby giving the impression of cover plates being supported by a single interradial oral ossicle. Of the other Rhenopygus species that also display this fusion, the oral surface of R. viviani n. sp. is not so highly domed, the oral plates are lower, wider, and more rounded adorally, with narrower distal ambulacra. The pedunculate zone is also much more highly variable along the length than in other taxa.
The low ridges or walls between floor plates and along the ambulacral margin of the oral plates (producing a series of grooves) are remarkably similar to the higher ridges seen in Heropyrgus, where they also accommodated the tube feet (Briggs et al., Reference Briggs, Siveter, Siveter, Sutton and Rahman2017). Therefore, it seems reasonable to assume these ridges served a similar function in R. viviani n. sp. and/or are muscle-/ligament-attachment scars for closing the cover plates.
The less-formal arrangement of the plates of the suboral constriction, their high degree of imbrication coupled with a rather inconsistent exposure of plates, and number of circlets present in various individuals from the same locality suggest they were capable of vertical extension and contraction. This agrees with observations of R. piojoensis (Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013).
Despite the oral surface being crushed in specimen EE 15756 from the Pavillon Member, it differs from specimens of the Cybèle Member only in there being a lesser amount of change in the morphology of the pedunculate zone plates, although there is still some discernible change. Subtle changes in pedunculate plate morphology were also noted between larger and smaller individuals by Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) in a population of 84 R. piojoensis, but this variation was not as obvious as in R. viviani n. sp.
Rhenopyrgus grayae (Bather, Reference Bather1915)
Figure 3.1, 3.2
- Reference Bather1915
Pyrgocystis grayae Bather, p. 58, pl. 3, figs. 1, 2.
- Reference Holloway and Jell1983
Rhenopyrgus grayae; Holloway and Jell, p. 1004.
- Reference Smith1985
Rhenopyrgus grayae; Smith, p. 732, text-fig. 11.
- Reference Sumrall2013
Pyrgocystis grayae; Sumrall et al., fig. 1.
- Reference Sumrall, Heredia, Rodríguez and Mestre2013
Rhenopyrgus grayae; Sumrall et al., p. 773.
Holotype
E 23470. Upper Ordovician, Katian, Ardmillan Series, Drummuck Group, South Threave Formation, Farden Member, Lady Burn Starfish Bed, part of the Craighead inlier.
Diagnosis (Emended)
Rhenopyrgus with highly domed oral surface. Oral plates narrow and high with acutely curved adambulacral margin. Ambulacral zone prominent with long proximal cover plates and wide distal ambulacral zone.
Occurrence
Known only from the Upper Ordovician, Katian, Ardmillan Series, Drummuck Group, South Threave Formation, Farden Member, Lady Burn Starfish Bed, Lady Burn, near Girvan, Scotland, UK.
Description
Rhenopyrgus with well-preserved oral surface comprised of long proximal and distal cover plates arranged in fairly symmetrical pattern, but with a vestige of 2-1-2 pattern discernible (Sprinkle, Reference Sprinkle1973), oral plates fairly rectangular with a narrow ambital margin and acutely curved adoral margin. Three discernible floor plates form distal part of ambulacra and together give the interradial oral area a deltoid shape. Only part of a single plate of the sub-oral constriction is visible between proximal margins of collar plates. Collar plates large and rectangular with curved proximal margins. Two ridges on distal external surface of collar plates run down behind proximal plates of pyrgate zone. Pyrgate zone distinctly tapering and comprised of squamate imbricate plates with curved proximal margins arranged in circlets of four, forming eight rows; 58 circlets in only complete specimen (holotype). Coriaceous sac comprised of a granular membrane with bulbous base.
Materials
E 29570, E 23674.
Remarks
This taxon was erected by Bather (Reference Bather1915) with the diagnosis focused primarily on the morphology of the pyrgate zone. Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) questioned the validity of erecting taxa based solely on pedunculate zone morphology, and thus we here emend the diagnosis to include characters of the oral surface.
Although Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) regarded this taxon as a member of the Rhenopyrgidae, the apparent lack of a suboral constriction led them to suggest that it should be placed into a new, undescribed genus. However, we believe that the apparent lack of a suboral constriction is a feature of taphonomy and that the sub-oral constriction has been withdrawn behind the collar plates. Because this specimen is preserved as a mold, the plates of the sub oral constriction are no longer clearly visible. However, a clue to its presence can be seen in Figure 3.1 (highlighted by a white arrow), where the distal portion of a plate is just visible between two collar plates, exactly where one would expect to see such a plate if the sub-oral constriction zone were withdrawn. As such, we feel that there is no need to erect a new genus specifically for this and other rhenopyrgid taxa apparently also lacking a sub-oral constriction.
Rhenopyrgus grayae displays suture lines between the plates of the oral surface, particularly between the interradial oral plates and the external portions of floor plates, distinguishing it from R. viviani n. sp., R. coronaeformis, and R. flos. Of the Rhenopyrgus taxa that also display suture lines between the oral and floor plates, R. grayae can be distinguished by its high, narrow, oral plates with a relatively gentle apical curve and its broad distal ambulacra and cover plates. Both R. whitei and R. piojoensis have more acute apical angles, with the former having distal ambulacra that narrow more abruptly, whilst the latter has narrower distal ambulacral and cover plates.
Rhenopyrgus sp. indet. 1
Figure 3.3, 3.4
- Reference Bather1915
Pyrgocystis procera (Aurivillius) Bather, p. 59.
- Reference Sumrall2013
Rhenopyrgus? procera; Sumrall et al., fig.1C, p. 764.
Occurrence
Known only from the lower Silurian, middle Llandovery, Saugh Hill Group, Newlands Subgroup, Newlands Farm Formation, Newlands Beds. Newlands Farm, Argyle, Scotland, UK, but may extend into the Ardmillan group of the Upper Ordovician (see remarks).
Description
Only known from one relatively complete specimen, E 62753 (Fig. 3.3, 3.4), others comprise only sections of pyrgate zone. This specimen has standard Rhenopyrgus construction, with turreted theca divided proximally to distally into an oral surface, collar, pedunculate zone, and coriaceous sac. Suboral constriction not apparent, but this is assumed to be a feature of taphonomy and that it is obscured by collar plates.
Oral surface only known from one crushed specimen (E 62753) displaying a single large, D-shaped, compound, interradial plate pressed down into top of pyrgate zone (Fig. 3.4). Cover plating and ambulacra unknown. Suboral constriction plating unknown. Collar plates indistinct, but appear wider than other pedunculate plates and form a circlet at the top of the peduncle. Pyrgate zone, slightly curved and composed of ~48 circlets in longest specimen (E 62753). Circlets comprised of fairly narrow squamate plates, with curved proximal margin, although many are broken giving a more angular look (Fig. 3.3). Circlets are arranged at highly inclined angle. Coriaceous sac comprised of numerous small platelets.
Materials
E 62751–62753.
Remarks
The material from the Newlands beds was originally assigned to Pyrgocystis procera (Aurivillius) by Bather (Reference Bather1915) owing to similarities in the morphology of the pedunculate zone. However, Bather (Reference Bather1915) expressed concern about the validity of erecting a species solely on the morphology of the pedunculate zone. Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) suspected that this species was actually a rhenopyrgid on the basis of the pyrgate zone plating and coriaceous sac, but did not formally transfer this species to the genus, owing to uncertainty surrounding the nature of the oral surface and suboral contraction.
On reanalysis of the material, we interpret a large D-shaped plate within the top of the pyrgate zone in specimen E 62753 as a compound interradial plate (Fig. 3.3) seen in other species of Rhenopyrgus with fused floor and interradial oral plates. The position and morphology of this D-shaped plate are similar to the disarticulated oral surfaces of R. flos (see Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008, pl. 16.4, text-fig. 25), although they lack the prominent ridges on the margins. Specimen E 62753 also displays many other features compatible with Rhenopyrgus, including an enlarged circlet of proximal pedunculate plates, which we interpret as collar plates, while the lack of a discernible suboral contraction may be a feature of taphonomy (i.e., it was withdrawn behind the collar plates—see comments in Functional Morphology section). Thus, we tentatively assign these specimens to the genus Rhenopyrgus. However, we agree with Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) and with Bather (Reference Bather1915, in part) that species should not be solely based on morphology of the pedunculate zone. Because the type material of Aurivillius (Reference Aurivillius1892) consists only of pedunculate plates, we consider Pyrgocystis procera a nomen dubium, and owing to the imperfect preservation precluding adequate comparison with other Rhenopyrgus species, we assign this material to Rhenopyrgus sp. indet. until better material is found.
Although the incomplete preservation of this occurrence precludes detailed comparison of all features with other named Rhenopyrgus species, useful comparisons can still be made. The pyrgate zone most closely resembles in both size and morphology those of Rhenopyrgus indet. 2, from the nearby Girvan site of “Ardmillan,” but which is probably from slightly older strata (see below). As such, it is described separately, but may in fact belong to the same, albeit indeterminable, taxon. The apparent fused nature of the interradial oral plating discounts affinities to R. grayae (Bather), which is also found from slightly older rocks nearby, or to Heropygus Briggs et al. (Reference Briggs, Siveter, Siveter, Sutton and Rahman2017). Otherwise, it is stratigraphically and morphologically most similar to R. viviani n. sp., but has slightly higher and narrower oral plates.
Rhenopyrgus sp. indet. 2
Figure 3.5
Occurrence
Part of the Craighead inlier, Katian, Upper Ordovician. “Ardmillan” [?Girvan], Scotland, UK, Ardmillan Series.
Description
Single specimen, displaying long portion (31 mm) of pyrgate zone comprised of at least 38 alternating circlets in eight columns (Fig. 3.5). Pyrgate zone plates large, steeply inclined, widely spaced and highly imbricating. Shape of plate margins unknown because all are damaged. Oral surface, suboral contraction, collar plates, and corriaceous sac are either not preserved or not exposed. Oral plating known only from a single disarticulated D-shaped ossicle with a ridged margin.
Materials
EE 16254.
Remarks
Only a single, poorly located specimen is known, however a curious disarticulated D-shaped ossicle, found in close association to the articulated portion pyrgate zone (Fig. 3.5), closely resembles a fused interradial plate of Rhenopyrgus. On this rather tentative basis, we assign this specimen to Rhenopyrgus sp. indet. Although the shape of the interradial plate and the size and morphology of the pedunculate zone distinguish it from the stratigraphically similar R. grayae, it instead resembles that of R. sp. indet. 1, formerly described as Pyrgocystis procera Bather, Reference Bather1915 (see above). Owing to the incompleteness of the material from both localities and the apparent stratigraphical differences, we describe it separately and await the discovery of better preserved material for further comparison.
Rhenopyrgus sp. indet. 3
Figure 3.6–3.8
- Reference Vizcaïno and Lefebvre1999
?Pyrgocystis sp. (Bather, Reference Bather1915) Vizcaïno and Lefebvre, p. 356.
- Reference Vizcaïno, Álvaro and Lefebvre2001
?Pyrgocystis sp. (Bather, Reference Bather1915) Vizcaïno et al., p. 218.
- Reference Sumrall, Heredia, Rodríguez and Mestre2013
Rhenopyrgidae Sumrall et al., p. 773.
Occurrence
(UCBL-FSL 713316) SW of Donnadieu, Babeau-Bouldoux, Hérault, France, E. filacovi Zone, Saint-Chinian Formation, late Tremadocian, Lower Ordovician; (UCBL-FSL 713312) la Croix de Roquebrun, Saint-Nazaire-de-Ladarez, Hérault, France, N. arenosus Zone, Foulon Formation, middle Floian, Lower Ordovician; (UCBL-FSL 713313–713315) Pont supérieur, Saint-Nazaire-de-Ladarez, Hérault, France, A. incisus Zone, Landeyran Formation, late Floian, Lower Ordovician;
Description
Known from several specimens with well-preserved portions of pyrgate zone and coriaceous sac, but disrupted or obscured portions of oral surface and sub-oral constriction. Oral surface in UCBL-FSL 713312 is disrupted and it is difficult to discern any structures with confidence, precise arrangement or morphology of cover plates difficult to determine; however, suspected interradial oral plates appear D-shaped (see black arrow on Fig. 3.7). Suboral constriction in UCBL-FSL 713312 and UCBL-FSL 713316 obscured by a prominent circlet of rectangular collar plates, with curved proximal edges (white arrows in Fig. 3.7, 3.8). Pyrgate zone long, either distinctly tapering or parallel sided; formed of small, low, squamate, imbricated plates arranged in eight rows of four alternating circlets, all with uniform curved proximal margins (64 circlets in only complete specimen); broader proximally, transitioning abruptly into coriaceous sac composed of small, irregularly placed, granular platelets distally.
Materials
UCBL-FSL 713312–713316. Specimens are found both flattened in shale and as three-dimensional molds in sideritic concretions.
Remarks
The specimens from Montagne Noire are the same as those referred to by Vizcaïno and Lefebvre, Reference Vizcaïno and Lefebvre1999 and Vizcaïno et al., Reference Vizcaïno, Álvaro and Lefebvre2001 as ?Pyrgocystis Bather (Reference Bather1915), but which are actually rhenopyrgids (Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013). The D-shape of the suspected interradial oral plates in UCBL-FSL 713312 and similarities in the pygate zone mean we tentatively suggest these specimens belong to the genus Rhenopyrgus. They are here described together because there is little to justify their division into different taxa apart from minor variations in stratigraphy. However, pyrgate zone morphology is not a satisfactory taxonomic character, and thus this grouping is provisional.
Despite UCBL-FSL 713312 being relatively complete, the oral surface is highly disrupted making further detailed taxonomically relevant observations impossible. While several specimens display a distinctly tapering pyrgate zone, such morphology is observed in several other rhenopyrgid taxa, particularly R. grayae, and cannot be used to differentiate it as a new species. Thus, because key taxonomic characters to differentiate this taxon from other rhenopyrgid species are not satisfactorily exposed, it seems prudent to wait for better preserved material to become available before erecting a new species. They are recorded here for completeness and are perhaps notable as being the oldest Rhenopyrgus/rhenopyrgid yet identified.
Discussion
The morphology of rhenopyrgids has long been thought to be fairly conservative (Sprinkle and Sumrall, Reference Sprinkle and Sumrall2015). However, description of the new taxon here and the re-evaluation of previously described material have resulted in the identification of wider morphological variation. This has implications for supra-familial relationships with other edrioasterids and function.
Variation in the oral surface and its implications
Specimens of Rhenopyrgus viviani n. sp. have some of the better-preserved external oral surfaces of any rhenopyrgid taxa yet described. This, along with the partial disarticulation of several R. viviani n. sp. specimens, the re-examination of previously described material, particularly the re-imaging of the genotypic specimen Rhenopyrgus coronaeformis Rievers (Fig. 3.9–3.11) and the description of Heropyrgus disterminus by Briggs et al. (Reference Briggs, Siveter, Siveter, Sutton and Rahman2017), have enabled novel observations, particularly of the organization of the oral surface and the position of the periproct.
Externally, there are no apparent sutures visible between the interradial oral plate and floor plates in R. viviani n. sp. (Fig. 2.2–2.9), R. coronaeformis (Fig. 3.10 black arrow), and R. flos (Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008, text-fig. 25, pl.16 8). Instead, the interradial area on the oral surface appears to be occupied by a single deltoid or D-shaped plate, which supports cover plates articulating in grooves along the entire length of the adambulacral margin (Fig. 2.2). However, in R. grayae, R. piojoensis and, H. disterminus the suture lines are more visible and distally the floor plates can be distinguished and seen to support individual cover plates, which is an arrangement and geometry identical to that seen in Edrioaster (Bell, Reference Bell1976) and other primitive edrioblastoids (Smith and Jell, Reference Smith and Jell1990). Thus, it appears that in R. viviani n. sp. and R. coronaeformis the interradial oral plate and the floor plates have become completely fused into a single compound element as seen in derived edrioblastoids (such as Lampteroblastus Guensburg and Sprinkle, Reference Guensburg and Sprinkle1994 and Porosublastus Sprinkle and Sumrall, Reference Sprinkle and Sumrall2015) and cyathocystids (Sprinkle and Sumrall, Reference Sprinkle and Sumrall2015). Fusion of these elements into a single compound plate, rather than loss of floor plates, is supported by the presence of faint suture lines in the distal radial parts of the interradial oral plate in a weathered specimen of R. viviani n. sp. (MPEP1143.1) and evidenced by sweeping extinction of this element in thin sections of edrioblastoids (C. Sumrall, personal communication, 2018).
In specimen MPEP1126.1 of R. viviani n. sp., the disturbed oral surface suggests the ambulacra are floored by an inclined projection of the proximal margin of this compound plate margin. A row of oval depressions occurs in areas just internally of the cover plate grooves, where the walls that maintain the cover plates in position appear to anastomose or become offset (Fig. 2.2, black arrows). Again, this is similar to what is observed in derived edrioblastoids (such as Lampteroblastus Guensburg and Sprinkle, Reference Guensburg and Sprinkle1994) and cyathocystids (Guensburg and Sprinkle, Reference Guensburg and Sprinkle1994, fig. 17 D; Sprinkle and Sumrall, Reference Sprinkle and Sumrall2015). Previously, such an oral arrangement was suggested to only occur in cyathocystids and independently in derived edrioblastoids, although this was used to imply a closer relationship to each other than either to rhenopyrgids by Sprinkle and Sumrall (Reference Sprinkle and Sumrall2015). Thus, the occurrence of this feature in some rhenopyrgids suggests convergence (homoplasy). This may have been part of a suite of adaptations to form an oral surface better able to resist compressional forces (further discussed below).
The ambulacral structure of Heropyrgus (Briggs et al., Reference Briggs, Siveter, Siveter, Sutton and Rahman2017) differs only in that it lacks cover plates and that, internally, the floor plates do not meet medially and instead appear to gape. What is similar between Heropyrgus and Rhenopyrgus (as well as cyathocystids and derived edrioblastoids) is the ridged nature of both the adambulacral margin of the oral plates and fused floor plates. It is curious that Heropyrgus should retain ridged adambulacral oral plate margins given the apparent lack of cover plates, but perhaps these ridges may have also supported and shielded the tube feet when they were extended and serve a similar function in Rhenopyrgus when the cover plates were opened. It is unknown if the ambulacra gaped internally in Rhenopyrgus, but we think that this is unlikely as there is no evidence to suggest that it did and that other turreted edrioasterids, and indeed all other echinoderms, display conventionally organized ambulacra floor plates (Mintz, Reference Mintz1970; Guensburg and Sprinkle, Reference Guensburg and Sprinkle1994; Sprinkle and Sumrall, Reference Sprinkle and Sumrall2015). Instead, as suggested by C. Sumrall (personal communication, 2019), an alternative explanation for the loss of cover plates and gaped ambulacra in Heropyrgus, other than as an original taxonomic character, is warranted. Deformation of the oral structures prior to fossilization is perhaps the most obvious potential cause, but more detailed work to explain the observed organization is desirable.
In addition to the comments by Sprinkle and Sumrall (Reference Sprinkle and Sumrall2015) regarding comparison of the oral surface construction among cyathocystids, edrioblastoids, and rhenopyrgids, a distinct difference between the former two families and rhenopyrgids is the presence of large “primary peristomal cover plates” (sensu Kammer et al., Reference Kammer, Sumrall, Ausich, Deline and Zamora2013) that support adjacent cover plates, as seen in Cyathocystis (Bell, Reference Bell and Sprinkle1982) and Astrocystites (see Fay, Reference Fay and Moore1968, fig. 174.1; Smith and Jell, Reference Smith and Jell1990, fig. 34; Kammer et al., Reference Kammer, Sumrall, Ausich, Deline and Zamora2013; Zhu et al., Reference Zhu, Zamora and Lefebvre2014, fig. 2B for this feature in other basal edrioblastoids). However, this feature is lost in derived edrioblastoids (e.g., Lampteroblastus Guensburg and Sprinkle, Reference Guensburg and Sprinkle1994) and is yet to be observed in any rhenopyrgid taxa. This may suggest a selective pressure for this feature to be lost independently in two distinct lineages, rather than reflect any phylogenetic relationships, but this needs to be tested further.
The phylogenetic trichotomy of rhenopyrgids, cyathocystids, and edrioblastoids (Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013, fig. 2) cannot be resolved without further observations. Some of the similarities observed in the three groups (cover plates articulating directly on the oral plates and floor plates fused into a compound plate) may have appeared independently in each lineage. It does, however, challenge the suggestion of Sprinkle and Sumrall (Reference Sprinkle and Sumrall2015) that cyathocystids and derived edrioblastoids are more closely related because they share a similar ambulacral structure; this has now been observed in rhenopyrgids also.
The holotype of R. viviani n. sp. displays a well-preserved short periproct composed of thin elongate rod-like plates surrounded by several small square-shaped plates (Fig. 2.7). It is significant because it confirms the tentative observations of Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) and Briggs et al. (Reference Briggs, Siveter, Siveter, Sutton and Rahman2017) that the periproct is located in the suboral constriction adjacent to the interradial oral plate in rhenopyrgids. This construction looks very similar to that seen in Cyathocystis americanus Bassler, Reference Bassler1936 (USNM 144878) and, to a lesser extent, that seen in Edrioaster bigsbyi Billings, Reference Billings1857 and Edriophus lavis (Bather, Reference Bather1914). In edrioasteroids, the periproct is located in the C-D interray. This is significant because it suggests the location of other structures, particularly the hydropore, which is usually located either within the C-D interradial oral plate or along its distal margin (Bell, Reference Bell1976). In Cyathotheca, the hydropore has been tentatively located within the interradial oral (“deltoid”) plate, although this is only identified in one specimen (Bockelie and Paul, Reference Bockelie and Paul1983). Unfortunately, the periproct obscures much of the central and distal part of the C-D interradial oral plate and its margin in the R. viviani n. sp. holotype (Fig 2.7). Furthermore, this feature also was not recorded by Briggs et al. (Reference Briggs, Siveter, Siveter, Sutton and Rahman2017) during their study of Heropyrgus. Thus, the location of the hydropore remains obscure in rhenopyrgids.
Rhenopyrgid functional morphology
Specimens of rhenopyrgids show great variability in the nature of preservation and the manner of skeletal deformation and disarticulation. This has led to a variety of interpretations of functional morphology. The cover plates have been interpreted as either opening individually (Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013) or simultaneously along the entire length of the ambulacra because they are interlinked (Bartels et al., Reference Bartels, Briggs and Brassel1998). The suboral constriction and collar have been identified as flexible (Smith, Reference Smith1985; Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013), whereas the whole pedunculate zone has been suggested to have been contractile so as to withdraw the animal into a burrow (Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008) or that the whole pedunculate zone remained in a burrow with only the oral surface exposed (Smith and Jell, Reference Smith and Jell1990, fig. 53). The coriaceous sac has been long identified as an attachment structure (Bather, Reference Bather1915; Rievers, Reference Rievers1961); however, it has been variously identified as only attaching at the very base to a hard/firm substrate or inserted into mud (Bather, Reference Bather1915; Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013), or as being inserted into mud, along with some or all of the pyrgate zone (Holloway and Jell, Reference Holloway and Jell1983; Smith and Jell, Reference Smith and Jell1990; Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008). The new material described here enables a review of these previous suggestions and new observations to be made.
Functional morphology of the oral surface
The oral surface of rhenopyrgids is frequently disarticulated or missing (Fig. 3.3–3.8), with disarticulation of the cover plates taking the form of individuals out of place (Fig. 2.2), the entire ambulacra opened like a fan (Figs. 2.7, 2.9, 3.9), or cover plates absent with just disarticulated oral plates remaining (Fig. 3.4; Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008). This has led to the notion that the oral surface was not such a robust structure when compared to the more commonly articulated plates of the pedunculated zone (Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013). While true, there are several apparent adaptations to strengthen parts of the oral surface. Primarily, the tessellation of oral plates, but the cover plates also display many examples of strengthening. They are generally robust, tessellate without gaps or pores, have an off-set triangular cross section with a flat external surface, and a proximally positioned internal keel (e.g., Fig. 2.7), which presumably acted both to strengthen the plate and provide a wider attachment site for ligaments and muscles to close them. The bases of the cover plates in some species of Rhenopyrgus were held in place by grooves in the adambularcral marginal wall of the interradial oral plates (e.g., Figs. 2.2, 3.9). Furthermore, in R. coronaeformis (Fig. 3.9, 3.10) the cover plates display a complex system of teeth and sockets that enabled the cover plates to interlock together. This interlocking seems very effective because the partially disarticulated cover plates are frequently preserved in a manner resembling a fan and have opened along the ambulacral midline (e.g., Fig. 3.9, 3.10; Bartels et al., Reference Bartels, Briggs and Brassel1998, fig. 174). Similar fan-like arrangements are also suggested in several R. viviani n. sp. specimens (Fig. 2.2, 2.3, 2.6, 2.7, 2.9), as well as in cyathocystids (Bockelie and Paul, Reference Bockelie and Paul1983). The fan-like opening of the ambulacra of R. coronaeformis was suggested by Bartels et al. (Reference Bartels, Briggs and Brassel1998) as evidence that the cover plates opened simultaneously along the midline of the ambulacra via articulation along the aboral margin of the oral plates, in a manner identical to that proposed by Bockelie and Paul (Reference Bockelie and Paul1983) for cyathocystids. We prefer to interpret the cover plates being able to open individually, as well as simultaneously, by articulation along the adambulacral margin of the oral plates owing to the cross-sectional morphology of the cover plates and that each was inserted individually within grooves on oral plate margins in Rhenopyrgus. What is apparent in these taxa though, is that the cover plates had various adaptations to strengthen and/or resist forces that would expose the ambulacra. The oral surface might have disarticulated more rapidly than other parts of the skeleton in rhenopyrgids (particularly the pyrgate zone) because the elements it is comprised of were, in part, moveable in life or supported by plates of the sub-oral constrictions, which was also moveable (see below).
Functional morphology of the pedunculate zone (suboral constriction, pyrgate zone, and coriaceous sac)
The suboral constriction is composed of several circlets of highly imbricated plates, that vary in size and shape. This zone has frequently been invoked as being flexible (Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008; Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013) and/or able to contract, along with the pyrgate zone (Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008). The new material described here and a review of previously published literature enables us to review these suggestions.
A single slab containing 84 specimens of R. piojoensis was described by Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013), where it was noted that several individuals had greatly extended zones whereas others lacked the plates of the suboral constriction, and interpreted them as being obscured by the collar plates (see Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013, fig. 6D, 6G). This is precisely what would be expected if the suboral constriction was contractile. Indeed, variation in the extension of the suboral constriction is seen in several other taxa (e.g., R. viviani n. sp., Fig. 2.5–2.7), although not to the extent as has been observed in R. piojoensis. Furthermore, in R. grayae, while the majority of the sub-oral constriction is absent, a small proximal part of a plate is visible between the margins of two collar plates, distal to the oral surface (see Fig. 3.1 white arrow). This seems likely to be the most proximal part of a proximal plate of the sub-oral constriction and appears precisely where we would predict it would be if the zone was preserved in the contracted position. If we assume that this zone is contractile in all rhenopyrgids, then it is of taxonomic significance, particularly for the holotype of R. grayae (see Fig. 3.1, 3.2), but also R. flos Klug et al. (Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008, pl.16, figs. 1–14), where an apparent lack of a suboral constriction was suggested by Sumrall et al. (Reference Sumrall, Heredia, Rodríguez and Mestre2013) to be grounds for placing such occurrences in new genera. However, if this feature is just a taphonomic artefact, it is not a reliable taxonomic feature and should not be used, which is a suggestion we follow here in the systematic section. Accordingly, it also enables the inclusion of material that otherwise displays all other rhenopyrgid morphological characters to named genera, such as Rhenopyrgus indet. 1 and 3 (see Fig. 3.3, 3.4, 3.6, 3.7).
A striking feature of the contraction of the suboral constriction in articulated rhenopyrgids is that the oral surface is not withdrawn behind the collar plate circlet, only plates of the suboral contraction are obscured. This small retraction amounts to only 2–3 mm. Smith and Jell (Reference Smith and Jell1990, fig. 53) reconstructed rhenopyrgids buried up to the oral surface; however, such a small retraction of the sub-oral constriction, on its own, seems insufficient to withdraw the oral surface far enough into a burrow to make this mode of life a feasible suggestion, although its movement may have deterred some predators. It seems more likely that this contraction served to protect and cover the periproct (and other pores located within the suboral constriction). This must have been a particularly useful adaptation for deterring organisms entering these orifices if rhenopyrgids were epifaunal mudstickers with only the coriaceous sac inserted into the substrate (see below). The collar plates may also have been able to be partially withdrawn, as evidenced by the presence of processes on the aboral portions of these plates in R. grayae (Smith, Reference Smith1985) and R. flos (Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008, pl. 16, figs. 1–14). Flexibility of the suboral constriction is apparent in numerous specimens and may have facilitated the redirection of the oral surface into more favorable positions within changeable currents (e.g., facing directly into a current to enhance feeding, disperse gametes or remove waste), as is observed in crinoids (Hess et al., Reference Hess, Ausich, Brett and Simms1999). There is, though, little evidence to suggest the mechanism by which the suboral constriction was contracted or extended. Based on analogies with some holothurians and evidenced by the large periproct, Bell (Reference Bell1977) and later Sumrall (Reference Sumrall1993) proposed that discocystinid edrioasteroids could contract or extend their pedunclate zones by inflation and deflation of respiratory trees via an anal pumping mechanism. Such a mechanism is possible in rhenopyrgids, however the periproct appears different from those of discocystinids and was located within the zone of contraction. As such, perhaps contraction was controlled by muscular or ligament action, but we await the discovery of better-preserved material.
The pyrgate zone elevates the feeding structures higher into the water column for more efficient filter feeding, a trend seen in other edrioasteroids (Sumrall, Reference Sumrall1993; Sprinkle and Sumrall, Reference Sprinkle and Sumrall2015). In addition to this, Klug et al. (Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008) speculated that the imbricate nature of the pyrgate zone in rhenopyrgids enabled it to contract, withdrawing the animal into the safety of a burrow. Indeed, this withdrawal would be made even more effective when combined with the contraction of the suboral constriction outlined above. To support this view, Klug et al. (Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008, text fig. 9a–9c) illustrated two end members: one end-member was short and broad with shallowly inclined plates; the other elongate, thinner, with steeply inclined plates, and a series of intermediate specimens from the Emsian of Morocco. They speculated that the short, broad variety were contracted individuals and applied this same logic to specimens of P. ansticei Bather, Reference Bather1915 (here classified as Rhenopyrgidae indet.). However, these specimens all show several features of diagenetic crushing (during sediment compaction). These are: (1) all specimens have disarticulated oral zones with only disarticulated elements (cover and oral plates) pushed into the top of the pyrgate zone (see Klug et al., Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008, pl. 16, figs. 3, 4, 8, 9); (2) no plates of the suboral constriction are exposed, and only the tips of the collar plates are visible; (3) many ‘contracted’ specimens are curved whereas extended ones are not; and (4) the plates of the pyrgate zone are not able to change inclination in life because they are fused in circlets of four plates (Sprinkle and Sumrall, Reference Sprinkle and Sumrall2015). Crushing is thus a far more likely explanation for the observations in R. flos and other indeterminate rhenopyrgid specimens reported by Klug et al. (Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008).
There is little further evidence of stem contraction in other rhenopygid taxa. Despite localized contraction being apparent in the proximal and distal parts of the pyrgate zone of specimen E 62753 (R. indet. 1, Fig. 3.3, 3.4), which is preserved perpendicular to the sediment bedding, evidence for taphonomic crushing rather than an ability to contract is suggested by the oral surface being crushed and the suboral constriction being contracted. There is very little variation in the structure of the pyrgate zone of R. piojoensis specimens irrespective of position with respect to bedding or, if the suboral constriction is contracted or not (see Sumrall et al., Reference Sumrall, Heredia, Rodríguez and Mestre2013, fig. 6). Furthermore, several specimens of R. viviani n. sp. (MPEP1126.1, EE 16642b, EE 15755) preserved perpendicular to the bedding have crushed oral surfaces and contracted suboral constrictions, but unaffected pyrgate zones, as one would expect if the latter could not contract (Fig. 2.1, 2.8).
Although the arrangement and external morphology of the pyrgate zone plates in rhenopyrgids resemble those of discocystinid edrioasteroids, which were obviously capable of extensive extension and contraction (Sumrall, Reference Sumrall1993, Reference Sumrall1996), they are different. The key structural difference is that the circlets of plates are unfused in discocystinids and could extensively slide past one another to telescopically extend or collapse the stem in an orderly manner, as demonstrated by numerous well-preserved individuals in both states (see Sumrall, Reference Sumrall1996, fig. 2.11, 2.14). Such orderly contraction in well-preserved material is only evident in the suboral constriction in rhenopyrgids, but never in the pyrgate zone, because the plates of this structure are fused into circlets, which could not slide past each other.
Thus, no specimens of rhenopyrgids unequivocally show a capability of being able to contract the plates of the pyrgate zone. If the pyrgate zone could not contract, it implies that rhenopyrgids would be unable to sufficiently retract the oral surface into a burrow by only the action of the suboral constriction, thereby making a burrow dwelling lifestyle, as reconstructed by Klug et al. (Reference Klug, Kröger, Korn, Rücklin, Schemm-Gregory, de Baets and Mapes2008) and Smith and Jell (Reference Smith and Jell1990), unfeasible.
The pyrgate zones of almost all rhenopyrgid taxa, however, display some degree of flexibility. Some suggest a great deal of flexibility (e.g., R. coronaeformis, R. flos, and R. sp. indet. 1), while others less so. This flexure of the pyrgate zone probably facilitated movement of the oral surface to more favorable positions within variable currents. We therefore conclude that the imbricate nature of the pyrgate zone was to facilitate flexure rather than contraction, as has been proposed for other echinoderms, such as the proxistele of solutans (Lefebvre et al., Reference Lefebvre, Derstler, Sumrall, Kroh and Reich2012).
No rhenopyrgid specimens have been found in-situ, but the coriaceous sac of several taxa from widely differing environments of preservation (e.g., R. cornaeformis from the relatively deep black shales of the Hunsrück Slate to the shallow water limestones of the Jupiter Formation) at different geological times and paleogeographical provinces frequently show entire membranous sacs, which may be lobed and/or textured. Heropyrgus disterminus and possibly some others even had spine-like extensions. The fact they are often complete suggests that they were not cemented to a hard substrate. The morphology is consistent with a mud-sticking strategy, where the lobed morphology and textured surfaces would have anchored the organism firmly within soft to firm substrates. It is perhaps worth reiterating that we propose only the coriaceous sac was buried, not the pyrgate zone.
Based on all these observations, we created the idealized reconstruction in Figure 5 to illustrate how we believe these animals would have appeared and behaved.

Figure 5. Idealized reconstruction of Rhenopyrgus viviani n. sp. Silurian (lower Telychian), Jupiter Formation, Jupiter River, Anticosti Island, Canada. Note individuals with extended and contracted suboral constrictions and with only the coriaceous sac and very distal part of the pyrgate zone buried in the substrate.
Conclusions
There is broader morphological variation in the construction of the oral surface within the Rhenopyrgidae than previously thought, although this variation is similar to that seen in the other stalked edrioasterid families Cyathocystidae and Edrioblastidae. As such, it would seem that stalked edrioasteroids have all converged on a similar morphological solution and that currently this feature does not help elucidate any clear interrelationships among these edrioasterids.
Oral surface construction was generally robust in rhenopyrgids, particularly so in certain taxa where the interradial plates and ambulacral floor plates were fused into a single compound plate (with equivalents seen in cyathocystids and derived edrioblastoids). More frequent loss of the oral structures during preservation, when compared to the pyrgate zone, is attributed to the fact that they were comprised of many moveable parts and were also supported by moveable plates of the suboral constriction.
The suboral constriction could contract behind the collar plates and this was done to protect the periproct and related orifices, while the small movement of the oral surface may have also disturbed potential predators. However, this contraction was insufficient to withdraw the oral surface behind the collar plates, and thus protection of the oral surface was not the primary function of the contraction.
The pyrgate zone was flexible, but not contractile. This served to raise the oral surface into the water column and potentially re-orientate it (in conjunction with movement within the suboral constriction) to more advantageous positions. Rhenopyrgids were epibenthic, firm or soft substrate stickers, not burrow dwellers or encrusters.
Acknowledgments
We would like to thank C. Sumrall, E. Nardin, A. Kroh, W.I. Ausich, and A.B. Smith for helpful discussions, comments, and supply of images of specimens; I. Rahman for scanning a specimen at the CERN light source; M. Graham and D. Comfort for specimen preparation; M. Cournoyer for making specimens available from the Musée de paléontologie et de l’évolution, Québec, Canada; M. Reich for making the type specimen of Rhenopyrgus coronaeformis Rievers and images of it available from the Bayerische Staatssammlung für Paläontologie und Geologie (Bavarian State Collections of Paleontology and Geology); H. Taylor for photography of other specimens; D. Vizcaïno and E. Monceret for making specimens from the Montagne Noire, France available for study and their donation and discussion of stratigraphy; D. Clarke for donation of a specimen from Anticosti Island; V. Tanasa for creating the reconstruction in Figure 5; the associate editor B. Lefebvre for pertinent comments, help with locating additional specimens, and resolving the lodging of existing specimens from the Montagne Noire; and reviewers J. Sprinkle, C. Sumrall, and third anonymous reviewer, who made helpful suggestions for improvements without which this paper would be poorer. This work was supported by the Spanish Ministry of Economy and Competitiveness [grant number CGL2017-87631], co-financed by the European Regional Development Fund.





