


Introduction
Pliosaurid plesiosaurs (sensu Benson and Druckenmiller, Reference Benson and Druckenmiller2014) were Mesozoic macropredator sauropterygians that evolved a short neck and relatively large skull condition (Sassoon et al., Reference Sassoon, Noè and Benton2012; Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013; Fischer et al., Reference Fischer, Arkhangelsky, Stenshin, Uspensky, Zverkov and Benson2015). Pliosaurids had a global distribution and their biochron spans from the Early Jurassic to the early Late Cretaceous (Kear, Reference Kear2012; Sasoon et al., Reference Sassoon, Noè and Benton2012; Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013; Benson and Druckenmiller, Reference Benson and Druckenmiller2014; Fischer et al., Reference Fischer, Arkhangelsky, Stenshin, Uspensky, Zverkov and Benson2015; Páramo-Fonseca et al., Reference Páramo-Fonseca, Gómez-Pérez, Noé and Etayo-Serna2016).
Despite this long record, the best-known species come mostly from European Oxfordian and Kimmeridgian deposits (Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013). However, new records have recently improved the knowledge of pliosaurids from other Jurassic localities such as Spitsbergen (Knutsen et al., Reference Knutsen, Druckenmiller and Hurum2012), Mexico (Buchy et al., Reference Buchy, Frey, Stinnesbeck and López-Oliva2003; Barrientos-Lara et al., Reference Barrientos-Lara, Fernández and Alvarado-Ortega2015), and Patagonian Argentina (Gasparini and O’Gorman, Reference Gasparini and O’Gorman2014) as well as the Lower Cretaceous of the USA (Albright et al., Reference Albright, Gillette and Titus2007; Schumacher et al., Reference Schumacher, Carpenter and Everhart2013), Colombia (Hampe, Reference Hampe2005; Páramo-Fonseca et al., Reference Páramo-Fonseca, Gómez-Pérez, Noé and Etayo-Serna2016), and Australia (Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013). At the same time, new phylogenetic analyses based on the revision of classic taxa, including the genus Pliosaurus Owen, Reference Owen1842 and new specimens, have improved the knowledge of the relationships among pliosaurids (O’Keefe, Reference O’Keefe2001; Ketchum and Benson, Reference Ketchum and Benson2010; Knutsen et al., Reference Knutsen, Druckenmiller and Hurum2012; Sassoon et al., Reference Sassoon, Noè and Benton2012; Benson and Druckenmiller, Reference Benson and Druckenmiller2014). Additionally, morphofunctional aspects of pliosaurids have recently been discussed (Foffa et al., Reference Foffa, Cuff, Sassoon, Rayfield, Mavrogordato and Benton2014a, Reference Foffa, Sassoon, Cuff, Mavrogordato and Bentonb).
One of the main results of the revisions of Knutsen (Reference Knutsen2012) and Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) is clarification of the classic problem of definition of the genus Pliosaurus (previously discussed by Tarlo, Reference Tarlo1960 and Noè et al., Reference Noè, Smith and Walton2004). These last revisions agreed in the validity and definition of Pliosaurus although with differences regarding the validity of some species. The general conclusions of these authors are followed in this contribution.
Argentine pliosaurids come from the Vaca Muerta Formation, Neuquén, from the middle Tithonian (Pliosaurus patagonicus Gasparini and O’Gorman, Reference Gasparini and O’Gorman2014) and late Tithonian (Gasparini et al., Reference Gasparini, Spalletti and de la Fuente1997; this study). At the beginning of 1986, the brothers Sergio and Rafael Cocca, technicians of the Museo Provincial de Ciencias Naturales Prof. Dr. Juan A. Olsacher de Zapala, Neuquén Province, Argentina (MOZ), discovered part of a pliosaur postcranium in Cajón de Almanza, 10 km east of Loncopué village, in west-central Neuquén Province (Fig. 1). A subsequent field trip (1986) was carried out with the MOZ staff and ZG to determine the possible position of the skull. In February 1987, the MOZ staff collected the entire specimen (MOZ 3728P) and in January 1988 one of the authors (L.S.) determined the exact stratigraphic position of the specimen.

Figure 1 Location and geological maps of the study area: (1) graphic log of the Vaca Muerta Formation at Cajón de Almanza; (2) the measured section, showing an overall increase in grain size and carbonate content toward the top.
The main objective of this contribution is to describe MOZ 3728P, the holotype and only specimen of Pliosaurus almanzaensis new species from the upper Tithonian levels of the Vaca Muerta Formation, Patagonia. Pliosaurus almanzaensis n. sp., along with P. patagonicus, are the only Pliosaurus species from Gondwana.
Geological setting
The Neuquén Basin is located in the western margin of South America, between 36º and 40º south latitude (Fig. 1.1), in west-central Argentina and central Chile. It is characterized by a 2600 m thick Mesozoic and Cenozoic record and covers a surface of over 120,000 km2 (Yrigoyen, Reference Yrigoyen1991). The basin is limited to the east and south by wide cratonic areas, and to the west by the Pacific Ocean. Due to its location, it has been considered as a back arc-retroarc basin (Digregorio et al., Reference Digregorio, Gulisano, Gutiérrez Pleimling and Minitti1984; Legarreta and Uliana, Reference Legarreta and Uliana1991).
In most of the Neuquén Basin, the Upper Jurassic to Lower Cretaceous record is characterized by the dark bituminous shales and marls of the Vaca Muerta Formation (Weaver, Reference Weaver1931). These deposits concordantly overlie the clastic and mostly continental deposits of the Tordillo Formation (Fig. 1.2); this early Tithonian basal contact is an isochronous surface that marks the beginning of a widespread marine transgression (Leanza, Reference Leanza1981). The top of the Vaca Muerta Formation is roughly diachronic (Leanza et al., Reference Leanza, Marchese and Riggi1977) and progradational (Mitchum and Uliana, Reference Mitchum and Uliana1985; Legarreta and Gulisano, Reference Legarreta and Gulisano1989). It includes younger stages passing from the south sector (middle Tithonian) to the central and northern sectors of the basin (Valanginian).
The study area is located in the deepest (western sector) of the basin, where the Vaca Muerta Formation exceeds 500 m in thickness and is dominated by offshore facies. The measured section, where the remains of Pliosaurus almanzaensis n. sp. were found, represents the late Tithonian. The specimen was found in the ammonite Substeueroceras koeneni Biozone, the uppermost part of the upper Tithonian (H. Parent, A. Garrido, personal communication, 2016; Parent et al., Reference Parent, Garrido, Scherzinger, Schweigert and Fözy2015). The unit is essentially composed of two facies associations. The lower part is entirely dominated by fine-grained deposits, ranging from purely siliciclastic mudstones to hybrid siliciclastic/carbonate marls (Fig. 1.2), accumulated under anoxic bottom conditions that favored the preservation of organic matter. Toward the upper part, the succession is characterized by dark grey and greenish calcareous laminated and bioturbated mudstones showing thin intercalations of very fine- and fine-grained sandstones and tuffaceous sandstones, and a very conspicuous floatstone bed essentially composed of oyster shells (Fig. 1.2). Sandstones are essentially massive, although some beds show normal grading and only a few strata are characterized by ripple lamination. This facies association clearly represents a progradational arrangement and, as shown by Massaferro et al. (Reference Massaferro, Zeller, Giunta, Sagasti and Eberli2014) and Sagasti et al. (Reference Sagasti, Remírez and Spalletti2015), should be interpreted as the record of foreset-topset ramp deposits developed upon anoxic bottomset facies.
Materials and methods
The holotype specimen shown and examined in this study is deposited in the MOZ. All measurements were taken using a digital caliper with 0.01-mm accuracy. To test the phylogenetic position of Pliosaurus almanzaensis n. sp., we used a dataset based on Benson and Druckenmiller (Reference Benson and Druckenmiller2014) modified by Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) with the addition of P. almanzaensis n. sp. and P. patagonicus. The data set (87 taxa scored for 270 characters) was compiled using Mesquite Software (Maddison and Maddison, Reference Maddison and Maddison2011). The final data set (Supplementary data I) was analyzed using TNT 1.5 version software (Goloboff and Catalano, Reference Goloboff and Catalano2016) using a heuristic search (tree bisection reconnection, with 1000 random addition sequence replicates). To evaluate the grade of homoplasy, consistency (CI) and retention (RI) indices (Farris, Reference Farris1989) were calculated. Additionally, Bremer Support (Bremer, Reference Bremer1994) was considered to indicate clade support. IterPCR scipt (Pol and Escapa, Reference Pol and Escapa2009) was used to detect wildcard taxa and remove them from the reduced consensus.
Repositories and institutional abbreviations
BHN, Musée-sur-Mer, Boulogne, France; BRSMG, Bristol City Museum and Art Gallery, Bristol, UK; CAMSM, Sedgwick Museum of Earth Sciences, Cambridge, UK; DOKDM, Dorking & District Museum, Dorking, UK; DORCM, Dorset County Museum, Dorchester, UK; MLP, Museo de La Plata, La Plata, Argentina; MOZ, Museo Provincial de Ciencias Naturales Prof. Dr. Juan A. Olsacher, Zapala, Neuquén, Argentina; NHMUK, The Natural History Museum, London, UK; OXFUM, Oxford University Museum of Natural History, Oxford, UK; PIN, Paleontological Institute, Russian Academy of Sciences, Moscow, Russia; QM, Queensland Museum, Brisbane, Australia.
Systematic paleontology
Class Sauropterygia Owen, Reference Owen1860
Order Plesiosauria de Blainville, Reference de Blainville1835
Family Pliosauridae Seeley, Reference Seeley1874
Clade Thalassophonea Benson and Druckenmiller, Reference Benson and Druckenmiller2014
Genus Pliosaurus Owen, Reference Owen1842 (sensu Knutsen, Reference Knutsen2012; Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013)
Type species
Pliosaurus brachydeirus Owen, Reference Owen1841 by original designation. Lower Kimmeridgian, England.
Pliosaurus almanzaensis new species
url:lsid:zoobank.org:pub:D2F274C4-B584-42AD-B3E9-E37C203B6EB6
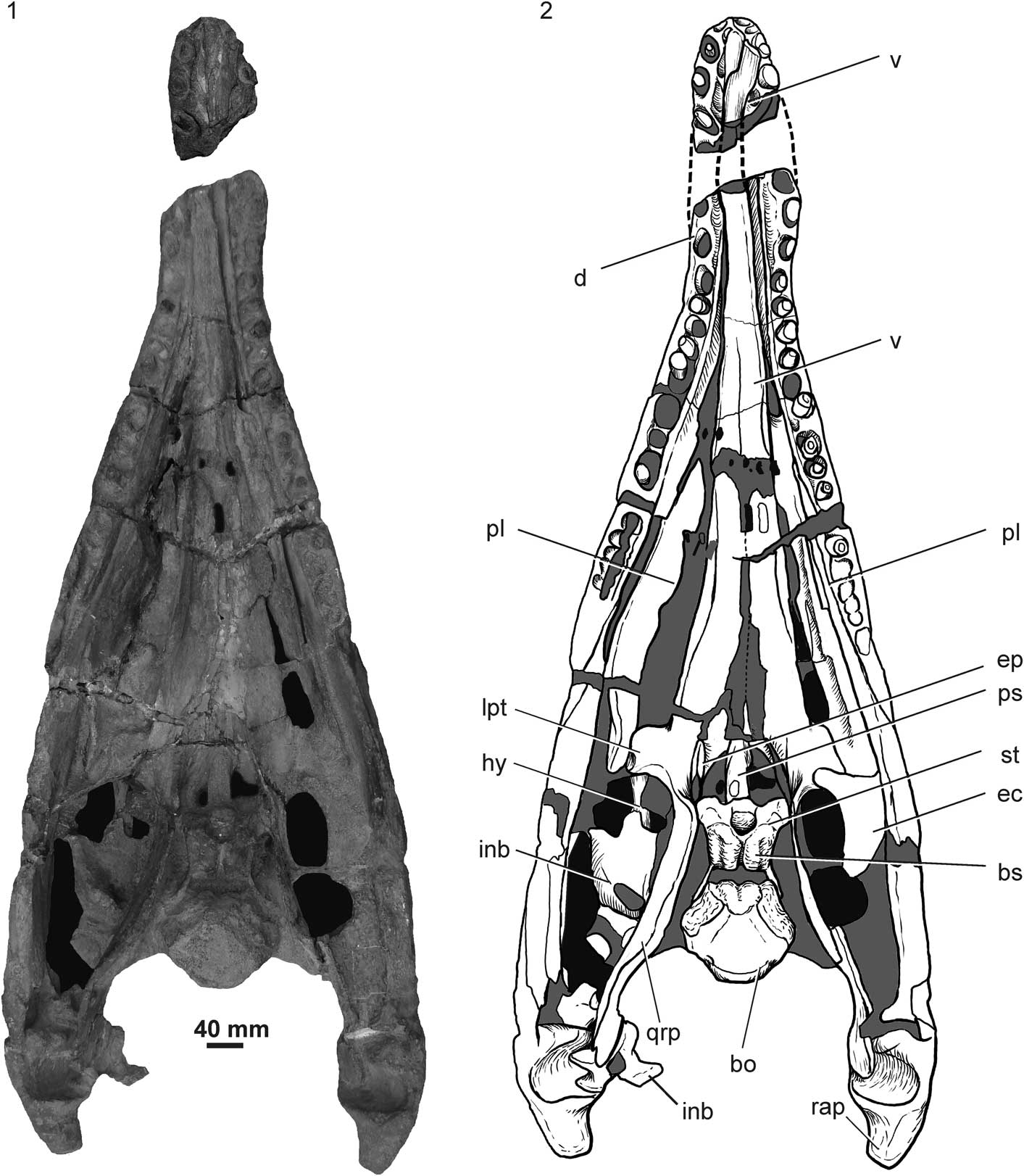
Figure 2 Pliosaurus almanzaensis n. sp. (MOZ 3728P), Vaca Muerta Formation, Cajón de Almanza, Neuquén Province, skull and mandible in dorsal view: (1) photograph; (2) interpretive drawing. bo=basioccipital; bs=basisphenoid; d=dentary; ec=ectopterygoid; ep=epipterygoid; hy=hyoid element; inb=indeterminate bone; lpt=lateral ramus of pterygoid; pl=palatine; ps=parasphenoid; qrp=quadrate ramus of pterygoid; rap=retroarticular process; st=sella turcica (pituitary fossa); v=vomer.

Figure 3 Pliosaurus almanzaensis n. sp. (MOZ 3728P), Vaca Muerta Formation, Cajón de Almanza, Neuquén Province, skull and mandible in ventral view: (1) photograph; (2) interpretive drawing. an=angular; ar=articular; bo=basioccipital; c=coronoid; d=dentary; ec=ectopterygoid; hy=hyoid element; in=internal naris; inb=indeterminate bone; mx=maxilla; pl=palatine; ps=parasphenoid; pt=pterygoid; sp=splenial; v=vomer.

Figure 4 Pliosaurus almanzaensis n. sp. (MOZ 3728P), Vaca Muerta Formation, Cajón de Almanza, Neuquén Province, skull and mandible in occipital view: (1) photograph, (2) interpretive drawing; (3–5) anterior part of the mandibular symphysis, in anterior (3), right lateral (4), and dorsal (5) view; (6) detail of mandibular symphysis showing the paradental plate; (7) detail of trihedral teeth of the right mandibular ramus. btu=basioccipital tuber; ft=functional tooth; oc=occipital condyle; ppl=paradental plate; pt=pterygoid.

Figure 5 Pliosaurus almanzaensis n. sp. (MOZ 3728P), Vaca Muerta Formation, Cajón de Almanza, Neuquén Province: (1, 2) detail of the opening of the Meckelian canal, in anterior (1) and posterior (2) view; (3) detail of braincase in dorsal view; (4) right femur; (5, 6) right forelimb in dorsal view, as photograph (5) and interpretive drawing (6). Afo=adductor fossa; bo=basioccipital; bpp=basipterygoid process; dc=distal carpal; eof=exoccipital-opisthotic facet; int=intermedium; Maop=Meckelian anterior opening; mf=middle furrow; mt=metatarsal; pre=prearticular; prg=prootic groove of basisphenoid; qrp=quadrate ramus of pterygoid; ra=radius; rad=radial; st=sella turcica (pituitary fossa); ul=ulna. Roman numerals indicate carpal numbers.
Holotype
MOZ 3728P, basicranium, complete palate, mandible and postcranial remains comprising a posterior cervical centrum, dorsal centra, humera and proximal elements of the forelimb and femora; upper Tithonian (Jurassic), Vaca Muerta Formation, Neuquén Province, Patagonia.
Diagnosis
Species of Pliosaurus that differs from others by two autapomorphies: angular participating in mandibular symphysis and occipital condyle without notochordal pit or irregularly-arranged grooves. Following combination of characters differentiates P. almanzaensis from other species: (1) trihedral teeth (differs from P. kevani Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013), (2) nine or more symphyseal teeth (differs from P. rossicus Novozhilov, Reference Novozhilov1948 and P. patagonicus), and (3) 15–17 post-symphyseal alveoli and (4) parasphenoid without ventral keel (differs from P. kevani).
Occurrence
Cajón de Almanza, 10 km east of Loncopué, Neuquén. Upper Tithonian levels of the Vaca Muerta Formation, Substeueroceras koeneni Biozone (Parent et al., Reference Parent, Garrido, Scherzinger, Schweigert and Fözy2015).
Description
The new specimen (MOZ 3728P) is preserved partially articulated. It comprises the mandible, part of the palate, and basicranium. Additionally, part of the postcranium including a posterior cervical, dorsal centra, limbs, and girdle fragments were collected. When the specimen was collected, the postcranium was exposed and therefore it was severely affected by recent weathering. However, the cranium and mandible were still buried and therefore less affected by weathering.
The cranium preserves a fragment of the left maxilla, complete palate, complete basicranium, hyoid elements, and an almost complete mandible. The mandible is triangular and measures ~900 mm from the posterior limit of the mandibular symphysis to the retroarticular process and 430 mm between the glenoid cavities. The palate and basicranium are located between the mandibular rami, whereas the cheek, rostrum, and skull roof are almost absent. Although the palate is well preserved, it shows slight displacement, mostly affecting the palatines.
Rostrum
Only part of the left maxillary is preserved (Fig. 3) attached to the palate. This fragment is rectangular and bears one tooth. There are small depressions and foramina in lateral view and near the alveolar margin.
Palate
The conjoined vomers are well preserved and located above the mandibular symphysis (Fig. 3). The vomers are fused with each other in the anterior part but a midline suture is observed posteriorly (Fig. 3). The dorsal surface of the vomer becomes strongly concave at the same level. The posterior contact with the pterygoid is not well preserved in dorsal view, but a transverse interdigitated suture between the elements is clearly visible in ventral view (Fig. 2). A lateral gently concave area is identified as part of the internal naris. Two foramina-like openings occur between the vomer and pterygoids, but close observation revealed that they were produced by breakage.
The internal nares are not completely preserved, however the right one preserves its medial border formed by a slightly concave border in the vomer; a posterior part of the palatine appears to close it posteriorly (Fig. 3).
Both palatines are preserved although with displacement, the right one almost in vertical position. The right palatine projects anteriorly and participates in the posterior margin of the internal naris (Fig. 3). It has pits and small sulci on the ventral surface. The suborbital fenestra is not well defined (Fig. 2).
As in other pliosauroids, the pterygoids form a central plate with anterior, lateral, and quadrate rami. No anterior interpterygoid vacuity is present. The space between the pterygoids is a consequence of breakage during preparation. In palatal view, the contact between the pterygoid and vomer forms a deep interdigitate suture (Fig. 2). Lateral to the central plate the pterygoids form two square lateral rami that contact the ectopterygoids (Fig. 2.2). Caudally to the central plate, the pterygoids form a posterior interpterygoid vacuity that is longer than wide (110 mm long x 40 mm wide; Fig. 2). The lateral margin of the posterior interpterygoid vacuity forms sharp convex margins (Fig. 2). The quadrate rami of the pterygoids are relatively long and contact the quadrate posteriorly (Fig. 2). In dorsal view, the quadrate rami form relatively tall walls that enclose the basicranium (Fig. 2). The pterygoids meet each other caudally to the posterior interpterygoid vacuity and form a long posterior pterygoid symphysis that totally covers the ventral surface of the basioccipital (Fig. 3). The posterior pterygoid symphysis shows a midline groove following the middle suture (Fig. 3). The posterior pterygoid symphysis is separated from the general palatal surface by a marked step that generates a lateral ridge (Fig. 3). Posterior to the posterior interpterygoid vacuity, the parasphenoid is covered ventrally by the pterygoid (Fig. 3).
Only the right ectopterygoid is preserved. It is plate-like with a concave medial margin. The natural position and exact relation with the pterygoid is unknown due to displacement (Fig. 2).
Braincase
The braincase is partially preserved in its natural position. The epipterygoids are at both sides of the posterior interpterygoid vacuity, although the left one is better preserved. The epipterygoid is low and is located anteriomedially to the high walls formed by the pterygoids that enclose the basicranium (Fig. 2).
The parasphenoid is a long element that divides the posterior interpterygoid vacuity into two portions ventrally covering the basisphenoid and basioccipital (Fig. 3). The anterior limit is affected by breakage, but it shows a cultriform process that becomes thinner and overlaps the pterygoid for a short distance. Posteriorly, the parasphenoid is covered ventrally by the posterior interpterygoid symphysis (Fig. 3). No sharp ventral medial keel is observed but the parasphenoid shows a triangular cross section with the apex facing ventrally. Additionally, there is a small convexity located on the posterior third (Fig. 3). In ventral view, the posterior end shows a small pit limited caudally by the posterior interpterygoid symphysis (Fig. 3).
The basisphenoid is a stocky element preserved in its natural position, forming the floor of the braincase (Fig. 2). All posterior dorsal and anterior surfaces are rugose, indicating the presence of chondral tissue covering the ossified preserved sector (Figs. 2.1, 5.3). However, the lateral surface and the posterior lateral processes show smooth bone surfaces. The lateral surfaces of the basisphenoid are anteroposteriorly concave and seem to bear a foramen for the internal carotid, but it is covered by matrix that prevents observation. The anterodorsal area bears a deep transversely elongated depression interpreted as the sella turcica, for reception of the hypophysis (Figs. 2, 5.3). The floor of this cavity shows a medial keel and lateral depressions, probably the foramina for the internal carotids. A deep medial groove is observed along the dorsal surface of the basisphenoid. This groove generates the posterior notch in the clivus (basisphenoid body). This notch is observed in other plesiosaurs such as Thalassiodracon hawkinsi Owen, Reference Owen1840 and Eurycleidus arcuatus Owen, Reference Owen1840 (O’Keefe, Reference O’Keefe2001, fig. 4; Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013). Lateral and posterior to the sella turcica, there is a depression on the dorsal surface of the basisphenoid, probably for the anterior process of the prootic (Fig. 5.3). The lateroventral surface of the basisphenoid projects into a pair of basipterygoid processes that contact the pterygoids (Fig. 2). The basisphenoid has two posterolateral wall-like processes that lateroventrally embrace the basioccipital and laterally limit a wide unossified space between the basioccipital and basisphenoid (Figs. 2, 5.3).
The basioccipital is visible in dorsal, ventral, and posterior views (Figs. 2, 3, 4.1, 4.2). In dorsal view, the floor of the neural canal is subtriangular (Fig. 2). The exoccipital facets are strongly laterally inclined (Fig. 2). Each facet is divided into anterior and posterior parts by a slightly narrow bridge (Figs. 2, 5.3). The posterior one probably receives the exoccipital whereas the anterior receives the opistothic. The entire anterior surface of the basioccipital shows a lobulate texture, indicating the presence of chondral tissue. The contact between basioccipital and basisphenoid is limited to a lateral contact with the posterolateral processes of the basisphenoid (Figs. 2, 5.3). The occipital condyle is formed only by the basioccipital. The condyle is strongly convex, with a slight central depression, but without a marked notochordal pit (Fig. 4.1, 4.2). The condyle is not limited by a marked neck, but the entire ventral surface shows a wide and diffuse concave area that limits the condylar articular surface. The basioccipital tubera are ventrolaterally directed, but they do not contact the dorsal surface of the pterygoid of the posterior pterygoid symphysis (Fig. 4.1, 4.2).
Indeterminate bones
Two indeterminate bones are probably part of the cranium. The largest is located on the left subtemporal fossa and shows a dorsal projection cut by a fracture; the same element is concave in palatal view. Other elements located close to the glenoid cavity seem to be part of the opistothic or prootic, but poor preservation impedes a secure identification (Fig. 2).
Mandible
The mandible is almost complete, missing only part of the symphysis and the coronoid eminences; because the palate is enclosed between the mandibular rami, some parts of the medial surface of the mandibular rami are not visible (Figs. 2, 3). The mandibular symphysis is relatively long, with straight lateral borders. Several teeth are partially preserved, but almost all of them are severely damaged and without enamel.
The dentaries are preserved in two parts. One is the anterior part of the symphysis that includes the first four alveoli for functional teeth of the right side and five of the left (Fig. 4.3–4.5). The anterior sector of the symphysis is wider than high and shows no lateral expansion. In lateral view, the ventral margin of the symphysis shows a marked change of inclination between the third and fourth alveoli (Fig. 4.4). This sector has no ventral keel. The ventral and lateral surfaces are pierced by foramina. Because the medial area of the symphysis is lost, the total number of symphyseal alveoli cannot be determined, but there are at least nine pairs. In lateral view, the mandible shows unclear sutures and therefore there is no way to determine the posterior limit of the dentary.
The coronoid, visible in medial view, is partially covered by the vomers. It projects anteriorly as an elongate sheet of bone that dorsally covers the medial surface of the mandible. The coronoid participates in the posterior part of the mandibular symphysis. No coronoid eminence is visible, probably because this area is damaged. The space between the coronoid eminence and the posteriormost alveolus is equivalent at least to the extension occupied by the posteriormost nine alveoli.
The splenial is visible in ventral view where contact with the coronoid is evident (Fig. 3). The anterior end of the splenial enters into the mandibular symphysis. Posterior to the symphysis, the splenial limits the ventral edge of the anterior Meckelian opening (Fig. 5.1).
The angular is well preserved. The posterior part of this element is not clearly distinguished because no sutures are visible, whereas the anterior part is visible clearly entering into the mandibular symphysis (Fig. 3).
The dorsal part of the surangular is not preserved, and the sutures between the surangular and the coronoid are not clear. No lateral external concavity is observed in the preserved part. The suture with the prearticular is visible in medial view (Fig. 5.2). The prearticular is a blade-like bone that medially limits the Meckelian canal and forms part of the anterior limit of the posterior opening of the Meckelian canal.
The articular forms the glenoid cavity and the retroarticular process. The glenoid cavity is deep and transversely expanded facing medially. No lateral expansion of the glenoid area is observed (Fig. 2). The retroarticular process is short and robust with a rounded end. The glenoid cavity is located slightly below the alveolar line.
Both hyoid elements are preserved. They are long, gracile and gently curved (Fig. 3).
Alveoli
The anterior sector of the symphysis bears five alveoli on the left side and four on the right side; the posteriormost alveoli are incomplete on both sides (Fig. 2). The anteriormost pair of alveoli is smaller, circular, and cranially oriented. The second pair is larger than the first. The third, fourth, and fifth alveoli are even larger and posteromedially inclined (Fig. 4.5). In the posterior part of the mandibular symphysis, there are four alveoli on the right side and three on the left. Considering the lost part, at least nine symphyseal alveoli are present. There are 17 alveoli on the right mandibular ramus (post-symphysis), whereas the left mandibular ramus shows 15 alveoli, but it is incomplete. Therefore, the right mandible shows 26+ alveoli and the left 24+ alveoli. The interalveolar spaces are short, but those between the third and fourth and between the fourth and fifth alveoli are about half as long as the anteroposterior length of the fourth alveoli. Clear paradental plates are observed between the third and fourth pairs of alveoli (Fig. 4.6).
Teeth
There are no complete teeth, but several partially preserved ones show a trihedral cross sections (Fig. 4.6, 4.7). The flat surface is labial and without striae. All of these features are observed in several teeth, but most clearly on the left fourth symphyseal tooth.
Postcranium
The poorly preserved vertebral remains comprise a posterior cervical centrum and several dorsal centra.
The four limbs are preserved. The femora are larger than the humeri (Fig. 5.4, 5.5). The right anterior limb is the best preserved (Fig. 5.5). The right humerus is preserved in two parts. The capitulum is convex as is the tubercle. Both capitulum and trochanter are almost completely confluent, but there is an anterior wide concave area between the capitulum and the trochanter on the posterior surface. The distal end shows two distal facets. The radial is short and widely rectangular (65 mm long x 35 mm wide). The ulna shows a posterior convex limit and two distal facets. The radial is also rectangular. The intermedium is not well preserved. Distal carpal IV shows a proximal triangular end. Metacarpal V is completely shifted to the distal line. The distal carpal II+III is rectangular. Several phalanges are preserved (Fig. 5.5, 5.6).
The femora are larger but more poorly preserved than the humeri. The femoral capitulum is less circular than the humeral one. The distal end is expanded and similar to the humeral distal end (Fig. 5.4, 5.5).
Etymology
After Cajón de Almanza, where MOZ 3728P, the holotype and only specimen, was collected.
Phylogenetic results
The analysis recovered +20000 most parsimonious trees of 1338 steps (CI=0.294; RI=0.683). Pliosaurus almanzaensis n. sp. was recovered as part of a polytomy including other Pliosaurus species, the Brachaucheninae Williston, Reference Williston1925, and Gallardosaurus iturraldei Gasparini, Reference Gasparini2009. This result is congruent with that obtained by Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013; Fig. 6.1). The IterPCR script identified the pliosaurid wildcard taxa Pliosaurus rossicus, DOKDM G/1–2, QM_F51291, Pliosaurus sp. (CAMSM J.35991), ‘P. irgisensis’ (Novozhilov, Reference Novozhilov1964), and P. patagonicus; these taxa were removed from the consensus and a reduced consensus was generated (Fig. 6.2). Pliosaurus almanzaensis n. sp. was recovered nested within the other Pliosurus species other than ‘P. andrewsi’ Tarlo, Reference Tarlo1960 (Fig. 6.2). The monophyly of Pliosaurus in the reduced consensus is supported by character 139 (0-1): trihedral cross section of teeth.

Figure 6 Cladogram showing the relationships of Pliosaurus almanzaensis n. sp., Vaca Muerta Formation, Cajón de Almanza, Neuquén Province: (1) strict consensus; (2) reduced consensus (excluding P. rossicus, P. patagonicus, DOKDM G/1–2, QM_F51291, Pliosaurus sp. (CAMSM J.35991), and ‘P. irgisensis’). Bremmer support indicated below selected nodes.
Remarks
To establish the validity of P. almanzaensis n. sp., it is compared with the other species of Pliosaurus. Pliosaurus brachydeirus and P. kevani have long alveolar lines: post-symphyseal alveoli (P. brachydeirus, 24; P. kevani, >22). Therefore, they differ from P. almanzaensis n. sp. with 15 or 16 post-symphyseal alveoli (Knutsen, Reference Knutsen2012; Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013). Additionally, P. kevani has subtrihedrical teeth with only a slightly flattened labial surface that differs from P. almanzaensis n. sp. with trihedral teeth. The lateral surface of the mandible of P. kevani is concave (not observed in P. almanzaensis n. sp.) and the parasphenoid of P. kevani shows a sharp ventral keel that is absent in P. almanzaensis n. sp. Finally, the angular can be seen to penetrate into the symphysis in ventral view in both P. almanzaensis and P. kevani.
Pliosaurus funkei Knutsen, Druckenmiller, and Hurum., Reference Knutsen, Druckenmiller and Hurum2012, in which the number of symphyseal alveoli is unknown, has the interalveolar wall thicker than in P. almanzaensis n. sp. Additionally, P. funkei shows a ‘retroarticular fossa’ in the lateral surface of the retroarticular process (Knutsen et al., Reference Knutsen, Druckenmiller and Hurum2012) that is absent in P. almanzaensis n. sp.
Sassoon et al. (Reference Sassoon, Noè and Benton2012) studied two specimens (BRSMG Cc332 and Cd6172) from the lower Kimmeridge Clay, Westbury, Wiltshire, and referred them to Pliosaurus sp. Later Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) named the species P. westburyensis Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013 based on BRSMG Cc332, and P. carpenteri Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013 based on BRSMG Cd6172. These species coincide with P. almanzaensis n. sp. because they show more than nine pairs of mandibular symphyseal alveoli. However, in both P. carpenteri and P. westburyensis, the angular does not penetrate in the symphysis in ventral view, whereas it does so in P. almanzaensis n. sp.
Pliosaurus patagonicus and P. rossicus each have a short symphysis with only six alveoli, differing from P. almanzaensis n. sp., which has at least nine symphyseal alveoli (Novozhilov, Reference Novozhilov1964, fig. 329; Halstead, Reference Halstead1971, text-fig. 1; Storrs et al., Reference Storrs, Arkhangelskii and Efimov2000, fig. 11.1B, C). Pliosaurus almanzaensis n. sp. differs from P. patagonicus in the following characters: (1) the symphysis of P. almanzaensis n. sp. is relatively more flattened than that of P. patagonicus; (2) the distance between the coronoid eminence and the posteriormost alveoli of P. patagonicus is equivalent to the length of the last fourteen alveoli, whereas in P. almanzaensis n. sp., this distance is shorter, approximately nine alveoli; and (3) the angular does not participate in the mandibular symphysis in P. patagonicus, but it does participate in P. almanzaensis n. sp.
Thus, the validity of Pliosaurus almanzaensis n. sp. from southwestern Gondwana is supported by two autapomorphies: (1) the angular entering into the mandibular symphysis and the occipital condyle without notochordal pit or grooves, and (2) by a combination of trihedral teeth, the symphysis with more than nine pairs of alveoli, the total number of alveoli approximately +26, and the parasphenoid without a ventral keel.
Discussion
The systematics of Pliosauridae is complex and under constant revision (e.g. Druckenmiller and Russell, Reference Druckenmiller and Russell2008; Ketchum and Benson, Reference Ketchum and Benson2010, Reference Ketchum and Benson2011; Knutsen, Reference Knutsen2012; Sassoon et al., Reference Sassoon, Noè and Benton2012; Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013; Fischer et al., Reference Fischer, Arkhangelsky, Stenshin, Uspensky, Zverkov and Benson2015). However, at least the definition of the genus Pliosaurus is now reaching a consensus. Both Knutsen (Reference Knutsen2012) and Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) indicated the importance of the presence of trihedral or subtrihedral teeth. Fischer et al. (Reference Fischer, Arkhangelsky, Stenshin, Uspensky, Zverkov and Benson2015) also recognized the presence of trihedral teeth in the basal brachauchenine Makhaira rossica Fischer et al., Reference Fischer, Arkhangelsky, Stenshin, Uspensky, Zverkov and Benson2015, but in this case the trihedral teeth show distinctive serrated carinae.
The recent revisions of Pliosaurus by Knutsen (Reference Knutsen2012) and Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) generated a new diagnosis of the genus. The diagnosis given by Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) includes seven features. Among these is the trihedral to subtrihedral (in P. kevani) teeth with a flat, smooth labial surface and convex lingual surface with longitudinal enameled ridges, a character also pointed by previous authors (e.g., Noè, Reference Noè2001; Sassoon et al., Reference Sassoon, Noè and Benton2012). MOZ 3728P shows trihedral teeth with crowns triangular in cross section, and thus, it is referred to Pliosaurus. The diagnosis of Pliosaurus given by Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) considered six additional characters in addition to tooth morphology. Five of these cannot be observed in MOZ 3728P. Only the third character, “occipital condyle lacking notochordal pit, but scored by several, irregularly-arranged grooves” (Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013, p. 4), could be scored. MOZ 3728P effectively lacks a notochordal pit but no irregular grooves are observed, although the condylar surface shows some irregularities.
Within the genus Pliosaurus there are also some systematic issues regarding the validity of the species included. Knutsen (Reference Knutsen2012) recognized four valid species of Pliosaurus plus four others of questionable validity, and, following Noè et al. (Reference Noè, Smith and Walton2004), remarked that the number of pairs of alveoli for functional teeth in the mandibular symphysis is a relevant character. Later, Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) confirmed most of the conclusions of Knutsen (Reference Knutsen2012), but retained P. rossicus as a valid species and added three new species. Additionally, the issues about the validity of the classic P. macromerus (Philips, Reference Phillips1871) and P. brachyspondylus (Owen, Reference Owen1840), both from the upper Kimmeridgian of England have not been solved. Both species are based on type material that is currently considered undiagnostic (Knutsen et al., Reference Knutsen, Druckenmiller and Hurum2012; Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013). Knutsen (Reference Knutsen2012) proposed the designation of neotypes for these two species but to date the request has not been submitted to the commission of the ICNZ. Therefore, they are here considered nomina dubia following Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013). The more recently nominated Pliosaurus species, P. patagonicus, was added by Gasparini and O’Gorman (Reference Gasparini and O’Gorman2014). This is the general background that we follow here. The species considered valid are listed in Table 1.
Table 1 Valid species of Pliosaurus (following Knutsen et al., Reference Knutsen, Druckenmiller and Hurum2012, Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013, and Gasparini and O’Gorman, Reference Gasparini and O’Gorman2014).

Phylogenetic results
The reduced consensus indicates some interesting relationships obscured in the complete consensus. First, Pliosaurus is recovered as a monophyletic group, including all Pliosaurus species considered other than ‘P. andrewsi,’ a result previously obtained by Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013). The position of ‘P. andrewsi’ outside the Pliosaurus clade is related to the absence of trihedral teeth (character 139, 0) and other features that differ from those recorded for other Pliosaurus species, such as mandibular glenoid fossa just posterior to the occipital condyle (character 10, 1), midpoint of the posterior interpterigoid vacuity posterior to anterior margin of the subtemporal fossa (character 104, 0), and anterior cervical neural spines curved posterodorsally (character 157, 0). Therefore ‘P. andrewsi’ needs referral to another genus.
The results regarding the Pliosaurus clade indicate that the plesiomorphic state of character 63 (condyle surface) is the lack a well-marked notochordal pit (which differs from those in Gallardosaurus Gasparini, Reference Gasparini2009 and Liopleurodon), but also the lack of grooves mentioned by Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) as diagnostic feature of Pliosaurus. Additionally, the topology indicates that the plesiomorphic state of character 83 (parasphenoid ventral surface) is the absence of a distinctive ventral keel on the parasphenoid, a feature shared by P. almanzaensis n. sp. and P. wesburyensis.
Basicranium comparisons
MOZ 3728P shows the brasicranial elements, usually covered by the skull roof or poorly preserved in other specimens, and thus enables improvement of the knowledge of this element in Pliosaurus.
The occipital condyle of Pliosaurus almanzaensis n. sp. lacks a notochordal pit as in Kronosaurus spp., but differs from other pliosauroids (Gasparini, Reference Gasparini2009; Benson et al., Reference Benson, Ketchum, Noè and Gómez‐Pérez2011, Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013). The presence of a deep notch in the posterior margin of the dorsal surface of the body of the basisphenoid [‘clivus’] as observed in P. almanzaensis n. sp. is present in some pliosaurids such as Peloneustes Lydekker, Reference Lydekker1889 and Liopleurodon Sauvage, Reference Sauvage1873 (Benson and Druckenmiller, Reference Benson and Druckenmiller2014), but is difficult to assess among the Brachaucheninae and other Pliosaurus spp. because the dorsal surface of basisphenoid is usually covered in most-complete specimens.
The parasphenoid shows the plesiomorphic triangular condition observed in other pliosaurids (O’Keefe, Reference O’Keefe2001), however, the morphology of the posterior end of the parasphenoid varies among pliosaurids. Some of them, such as Rhomaleosaurus cramptoni (Carte and Baily, Reference Carte and Bailey1863) and Peloneustes philarchus Seeley, Reference Seeley1869, show a short parasphenoid not ventrally covering the basioccipital in the midline (Smith and Dyke, Reference Smith and Dyke2008; Ketchum and Benson, Reference Ketchum and Benson2011). In contrast, among species of Pliosaurus, in P. kevani, the parasphenoid reaches the level of the basioccipital (Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013). Additionally, in P. kevani, the parasphenoid has a posterior notch (Benson et al., Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013, fig. 5), and in P. almanzaensis n. sp., there is a small depression in the posteriormost visible area of the parasphenoid. Therefore, the posterior notch, similar to that of P. kevani, is probably present although it is mostly covered by the posterior interpterygoid symphysis. The rugose surface of the basisphenoid has been mentioned for Peloneustes philarchus (see O’Keefe, Reference O’Keefe2001), suggesting a poor state of ossification. The same feature in Pliosaurus almanzaensis n. sp. reinforces the observation of O’Keefe (Reference O’Keefe2001) about poor ossification of the basisphenoid in pliosaurids. The sella turcica of Pliosaurus almanzaensis n. sp. is located in a caudoventrally directed depression, differing from the caudally directed depression recorded in several Plesiosauroidea such as Brancasaurus brancai Wegner, Reference Wegner1914, Dolichorhynchops Williston, Reference Williston1902 (see Sato et al., Reference Sato, Wu, Tirabasso and Bloskie2011), and Alexandronectes zealandiensis Otero et al., Reference Otero, O’Gorman, Hiller, O’Keefe and Fordyce2016.
Symphyseal alveolar account and morphological disparity
According to Knutsen (Reference Knutsen2012), the Upper Jurassic species of Pliosaurus ranging from the Kimmeridgian to the Tithonian show a reduction in the total number of alveoli in the mandible as a whole, especially those of the symphyseal sector, and particularly during the Tithonian, there are no forms with eight or nine symphyseal tooth pairs. This suggests an evolutionary trend of the cranium of Pliosaurus toward a relative decrease of symphyseal alveoli. This model agrees with the condition in P. patagonicus (six symphyseal alveoli pairs; Gasparini and O’Gorman, Reference Gasparini and O’Gorman2014) from the middle Tithonian levels of the Vaca Muerta Formation, Neuquén. However, the condition of the upper Tithonian P. almanzaensis n. sp. (with at last nine symphyseal alveoli pairs) shows that during the last part of this stage, Pliosaurus species with symphysis longer (in terms of alveoli count) than in P. patagonicus and P. rossicus were present in Patagonia. The upper Tithonian represents the moment just prior to the faunal turnover that affected the plesiosaurs at the end of the Jurassic (Benson and Druckenmiller, Reference Benson and Druckenmiller2014). Pliosaurus almanzaensis n. sp. indicates that the decrease in morphological disparity regarding the morphology of the mandibular symphysis is not as marked as previously supposed (Fig. 7) and could be relevant in discriminating between various models (with or without a previous decrease of diversity) of the plesiosaur faunal turnover at the end of the Jurassic.

Figure 7 Plot of symphyseal alveoli number and stratigraphic position of various Pliosaurus species. Data from Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) and Gasparini and O’Gorman (Reference Gasparini and O’Gorman2014).
Acknowledgments
The authors thank J.J. Moly (Museo de La Plata) for preparation of part of the specimen, J. González for the drawing, and C. Deschamps for reviewing the English grammar. Thanks to A. Garrido and H. Parent for comments regarding the age of the stratigraphic levels where the material was collected and M.S. Fernández for reading and commenting on an earlier version of the manuscript. Author ZG also thanks the curators of the vertebrate paleontological collections of the following institutions during visits: CAMSM; MOZ; NHMUK; OXFUM; Bayerische Staatssammlung für Paläontologie und Historische Geologie, München, Germany; Paläontologische Sammlung der Eberhard Karls, Universität Tübingen, Germany; Muséum National d’Histoire Naturelle, Paris, France; Peterborough City Museum and Art Gallery, Peterborough, UK; Staatlische Museum für Naturkunde, Stuttgart, Germany; and Universidad Autónoma de Nueva León, Linares, México. The authors thank the Willi Hennig Society for access to TNT software. We also thank the financial support of research grants PICT 2008-0261, PICT 2012-0748, PIP 433, UNLP N 607, and National Geographic Society Grant 5178. Finally, the authors thank T. Sato and one anonymous reviewer for comments that improved this contribution.
Supplementary Material
Data available from the Dryad Digital Repository: http://dx.doi.org/10.5061/dryad.jc6r4.








