



Introduction
Fossil sponges are a well-known faunal component of the Upper Ordovician (Sandbian) Lenoir Limestone (lower Chickamauga Group) of the Valley and Ridge Provenance of east Tennessee, USA. Although mentioned in several papers dealing with lower Paleozoic sponges (Raymond and Okulitch, Reference Raymond and Okulitch1940; Finks and Rigby, Reference Finks, Rigby and Kaesler2004) and stratigraphy (Benedict and Walker, Reference Benedict and Walker1978; Walker and Benedict, Reference Walker, Benedict, Walker, Broadhead and Keller1980; Shanmugam and Walker, Reference Shanmugam and Walker1983), the fossil sponge fauna from the Ordovician Lenoir Limestone of East Tennessee was never studied in detail (Myers et al., Reference Myers, Sumrall and Johns2009).
Occurrences of sponges in the Lenoir Limestone and other localities in eastern North America were among the first Ordovician fossil sponge localities to be described from the continent, including the Trenton Limestone at Dixon, Illinois (Ulrich and Everett, Reference Ulrich and Everett1890), Mingan Island, Quebec, Canada, and Valcour Island, New York (Raymond and Okulitch, Reference Raymond and Okulitch1940, later revised by Rigby and Desrochers, Reference Rigby and Desrochers1995). Sponges from western North America were first described from the Pogonip Group in Nevada (Bassler, Reference Bassler1941).
Raymond and Okulitch (Reference Raymond and Okulitch1940) investigated Lower to Middle Ordovician sponges from the Mingan Islands of the Champlain Valley, Tennessee and Virginia. From Tennessee, they mentioned Hudsonospongia cyclostoma Raymond and Okulitch, Reference Raymond and Okulitch1940 (Lenoir Limestone, Knoxville, Tennessee), Rhopalocoelia regularis Raymond and Okulitch, Reference Raymond and Okulitch1940 (Sevier Shale near Neubert, Tennessee), Allosacus sp. (Ottosee and Lenoir Limestone), and Zitelella pannosa Raymond and Okulitch, Reference Raymond and Okulitch1940 (Otosee Limestone).
The aim of this contribution is to describe the diverse sponge fauna collected from the Lenoir Limestone, east of Lenoir City, Loudon County, Tennessee. Furthermore, we will discuss the paleobiogeographical implications of this association in the context of the North American sponge distribution.
Geology and stratigraphy
In the study area, the middle and lower parts of the Chickamauga Group are a thick (~450 m) succession of limestones and shales in the Valley and Ridge provenance of East Tennessee (Fig. 1). The unit thickens and becomes more clastic to the southeast. The base of the Lenoir Limestone suggests restricted peritidal deposition with cryptagal laminated micrites, and a sparse fauna including large leperditid ostracodes and gastropods (Walker and Benedict, Reference Walker, Benedict, Walker, Broadhead and Keller1980). The upper part of the unit shows deeper subtidal enviroments and becomes more open marine with a rich and diverse fauna. Regionally, the Lenoir Limestone overlays and interfingers with the Athens Formation and is overlain by the Holston Limestone.
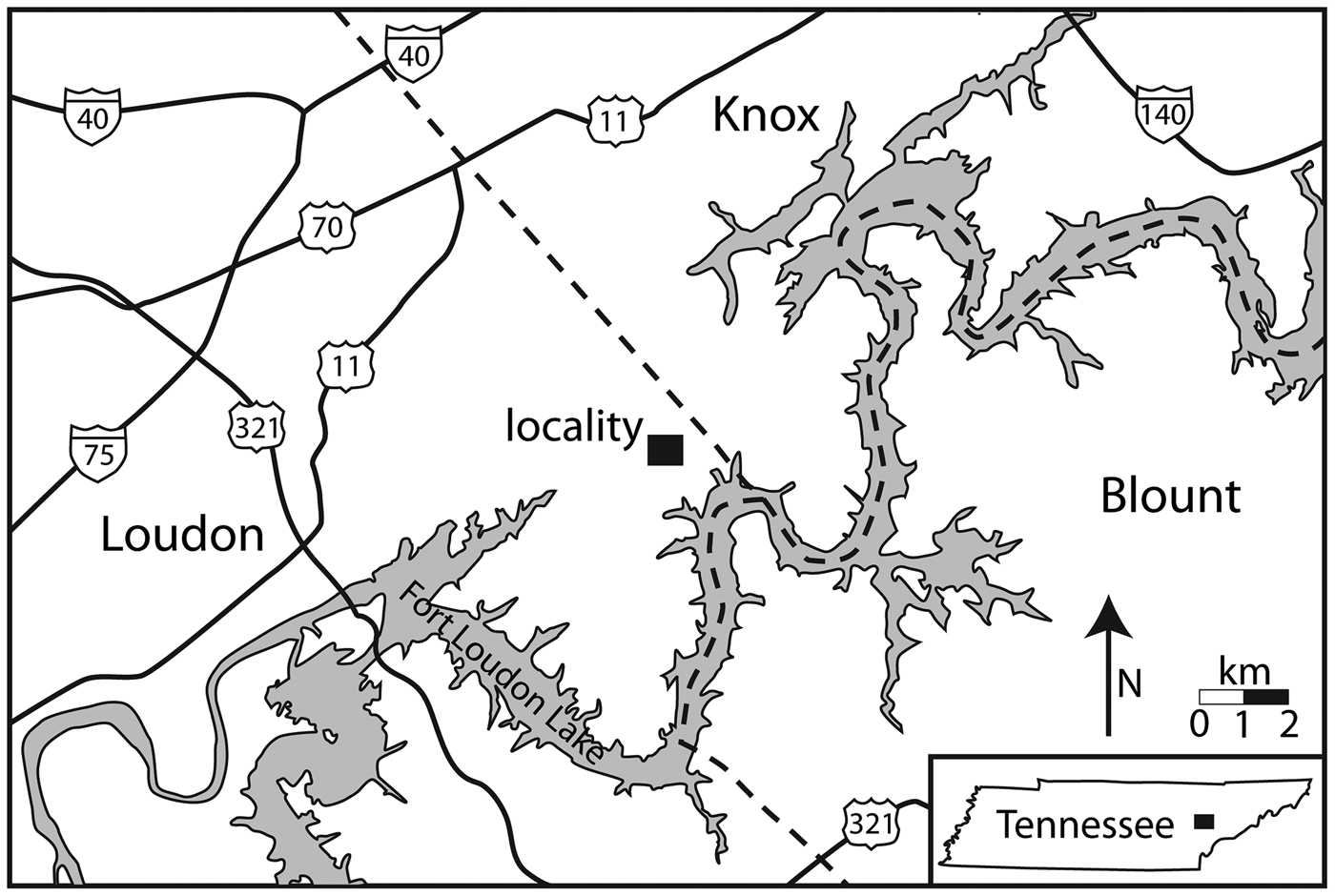
Figure 1. Geographic map and location of the collecting locality east of Lenoir City, Loudon County, Tennessee.
The sponge fauna described here was collected from the Upper Ordovician (Middle Ordovician in older literature) Lenoir Limestone, which is a gray, nodular, argillaceous limestone with minor interbedded shale. Limestones range from bioclastic wackestones to packstones and bear an open marine fauna. The fauna is dominated by large complete to fragmented sponges, pelmatozoan ossicles, trilobite sclerites, brachiopods, gastropods and bryozoans. The presence of large digitate crinoid holdfasts suggests that some areas had a soft to firm bottom.
Sponges were collected from the edge of a small sinkhole from a heavily weathered exposure of the Lenoir sponge beds. Sponges were seen both weathering free from matrix and incorporated into limestone slabs ranging from a few to tens of cm thick. The slabs ranged from wakestones to bioclastic grainstones dominated by echinoderm ossicles. Some of the beds were lightly bioturbated with grain filled and mud filled burrows.
Sponge specimens included mostly free and toppled specimens in the rock matrix. More fragile forms were often somewhat compressed and fragmented with spongocoels filled with matrix. The toppled and fragmented nature of some of the sponges suggests that they were subjected to high energy associated with storm events. No clear evidence was seen locally of in place bioherms of other organic buildups.
Materials
Repository and institutional abbreviation
The specimens are housed under the prefix MMNHC in the McClung Museum of Natural History and Culture (MMNHC) at the University of Tennessee.
Systematic paleontology
Class Demospongea Sollas, Reference Sollas1875
Order Orchocladina Rauff, Reference Rauff1895
Family Anthaspidellidae Ulrich in Miller, Reference Miller1889
Genus Rugocoelia Johns, Reference Johns1994
Type species
Rugocoelia eganensis Johns, Reference Johns1994.
Rugocoelia loudonensis new species
Figure 2.1–2.7

Figure 2. (1–7) Rugocoelia loudonensis n. sp. (1) Lateral view showing the concentric, regularly spaced ridges and the basal view of the deep central cavity, holotype MMNHC 13274, scale bar 10 mm; (2) lateral view of a complete specimen, MMNHC 13275, scale bar 10 mm, (3) dorsal view of a complete specimen showing the open, broad spongocoel and thin walls, MMNHC 13276, scale bar 10 mm; (4) detail of the spicular net in longitudinal view, showing typical anthaspidellid organization, MMNHC 13277, scale bar 1 mm; (5) detail of the previous photograph, showing regular anthaspidellid skeleton with main columnar trabs separating rounded pores and thin regularly intercalated intraparietal trabs, scale bar 1 mm; (6) transverse thin section showing straight canals passing through the entire wall, MMNHC 13278, scale bar 1 mm; (7) transversal thin section showing a different sector of the wall with better-defined intraparietal spicules between canals, scale bar 1 mm. (8–11) Rhopalocoelia regularis Raymond and Okulitch. (8) Lateral view of an almost complete specimen with smooth external surface, MMNHC 13263, scale bar 10 mm; (9) longitudinal thin section of the same specimen figured in (8), showing thick walls and deep spongocoel and the typical anthaspidellid scalariform skeleton with mid wall trab divergence, scale bar 5 mm; (10) detail of (9) showing a closer view of the spicular net of vertically bending trabs and transversal radial canals, some of them expanded at mid wall or near the gastral cavity, scale bar 1 mm; (11) almost complete longitudinal section showing the expansion of sponge diameter, a part of the tangential view of the skeleton showing rounded pores, scale bar 1 mm. (12, 13) Psarodictyum sp. (12) Complete open conical to palmate specimen, with a pedunculated termination MMNHC 13265, scale bar 10 mm; (13) fragmented lamellate form showing thin walls, MMNHC 13266, scale bar 10 mm.
Type specimens
Holotype MMNHC 13274, and eight paratypes MMNHC 13275-13282 from the Lenoir Limestone East of Lenoir City, Loudon Co., Tennessee, Upper Ordovician (Sandbian).
Diagnosis
Rugocoelia with thin walls, 7 mm on average, and wide central cavity. Surface marked by regular circumferential ripple-like ridges 1–2 mm high, 1–1.5 mm across, and a spacing of 3–5 mm between ridge crests. Radial canals vertically stacked, separated by parietal distances of 0.4–1.1 mm. Canals straight, ~0.2–0.7 mm in diameter. Anthaspidellid skeleton, with two trabs between radial canals in tangential section, and commonly two or three parietal trabs in transverse section.
Occurrence
Lenoir Limestone (Upper Ordovician) East of Lenoir City, Loudon, Co., Tennessee.
Description
Lamellate, open conical or bowl-shaped sponge sometimes slightly pedunculated, with overall height reaching 70 mm. The maximum diameter occurs at the top of the sponge reaching 60 mm. Relatively thin walls, 7 mm on average, outlining a wide open central cavity that reaches almost to the base of the sponge body. Surface marked by regular circumferential ripple-like ridges 1–2 mm high, 1–1.5 mm across and a spacing of 3–5 mm between ridge crests.
Radial canals vertically stacked, separated by parietal distances of 0.4–1.1 mm. Canals straight, ~0.2–0.7 mm in diameter. Rare vertical canals occur in transverse section, 0.2 mm in diameter.
Scalariform anthaspidellid skeletal net, regularly with two trabs between radial canals in tangential section, and commonly two or three parietal trabs in transverse section. The longitudinal section also shows main columnar trabs separating pores and thin regularly intercalated intraparietal trabs. Surface of trab divergence along and parallel to gastral margin.
Amphiarborescent monoclonids dominate, but polyclonid and Y-shaped dendroclones are also common in endosomal parietal walls. Endosomal spicules 0.35–0.7 mm long.
Ectosomal dermal layer not well preserved. Dermal pores usually 0.4–0.9 mm across, generally spaced 0.4–0.9 mm apart in both vertical and parietal directions. Spiculation and pore pattern of ectosome reflects that of underlying endosome.
Etymology
Rugocoelia loudonensis: from Loudon Co., Tennessee.
Remarks
The Tennessee material is, in many respects, similar to the type species Rugocoelia eganensis Johns, Reference Johns1994 from the Lower Ordovician, Egan Range, Nevada. However, the wall of our material is considerably thicker and the diagnostic ridges are more tightly spaced than the type species.
Genus Rhopalocoelia Raymond and Okulitch, Reference Raymond and Okulitch1940
Type species
Rhopalocoelia clarkii Raymond and Okulitch, Reference Raymond and Okulitch1940.
Rhopalocoelia regularis Raymond and Okulitch, Reference Raymond and Okulitch1940
Figure 2.8–2.11
Type specimen
Holotype, Museum of Comparative Zoology 9332, Sevier Formation (Upper Ordovician) east Knoxville, Tennessee.
Occurrence
Lenoir Limestone (Upper Ordovician) east of Lenoir City, Loudon, Co., Tennessee. The type species was described by Raymond and Okulitch, Reference Raymond and Okulitch1940 from Sevier Formation (Upper Ordovician), east of Knoxville, Tennessee. The species was also reported from the San Juan Formation (Dapingian - Darriwilian) Cerro La Silla, Niquivil, and Villicum sections, San Juan Province, Argentine Precordillera (Carrera, Reference Carrera and Benedetto2003).
Description
Long, cylindrical sponge, slightly expanded towards the top, with deep and wide spongocoel. Minimum diameter at the base 12 mm and a maximum diameter near the top 15 mm, with thin walls averaging 4 mm thick. Smooth, poorly preserved differentiated dermal layer composed of irregularly disposed monoclonids mainly among external pores.
Fine anthaspidellid skeleton with a ladder-like appearance of the spicular mesh with the surface of skeletal trab divergence near the gastral surface. Very fine vertical trabs, united by small monoclonid desmas (trab separation 0.2–0.3 mm), meet the external surface at high angles.
Horizontal radial canals, disposed in regular stalked series, range 0.3–0.5 mm in diameter. Some of the canals are slightly expanded at mid wall or near the gastral surface.
Materials
Two almost complete specimens MMNHC 13263, MMNHC 13264.
Remarks
The genus Rhopalocoelia Raymond and Okulitch, Reference Raymond and Okulitch1940 was erected to include tubular and cylindrical forms with an axial cavity that extends almost to the base, numerous branched radial canals, and a smooth exterior surface. Raymond and Okulitch (Reference Raymond and Okulitch1940) proposed two species in the genus Rhopalocoelia. The type species, Rhopalocoelia clarkii Raymond and Okulitch, Reference Raymond and Okulitch1940, from New York and Vermont and Rhopalocoelia regularis Raymond and Okulitch, Reference Raymond and Okulitch1940 from Sevier Formation, east of Knoxville, Tennessee.
Rhopalocoelia regularis has a smaller spicule mesh than R. clarkii, and a more regular arrangement of canals, although they are sinuous and sometimes bifurcate, unlike how they are characterized in the genus diagnosis. Raymond and Okulitch (Reference Raymond and Okulitch1940) described R. regularis as having large canals, which were sketched as being somewhat bulbous toward the gastral margin. De Freitas (Reference De Freitas1989), in his revision of some of the anthaspidellid genera, recognized large straight, not bulbous radial canals with a 0.4–0.6 mm diameter in R. regularis. We have observed in our material the same body form and canal morphology described by Raymond and Okulitch (Reference Raymond and Okulitch1940) in their description of the type material of R. regularis, with canals straight and slightly expanded towards the gastral margin. Rhopalocoelia regularis reported form the Argentine Precordillera (Carrera, Reference Carrera and Benedetto2003) shows comparable external form and structural characteristics like those found in the type species. A well-preserved dermalia occurs in some Argentinean specimens, which is not as well developed in the Tennessee material.
Genus Psarodictyum Raymond and Okulitch, Reference Raymond and Okulitch1940
Type species
Psarodictyum magnificum Raymond and Okulitch, Reference Raymond and Okulitch1940.
Psarodictyum sp.
Figure 2.12, 2.13
Occurrence
Lenoir Limestone (Upper Ordovician) East of Lenoir City, Loudon, Co., Tennessee.
Description
Discoidal, open obconical to funnel-shaped sponge, ~20 cm in maximum diameter expanding upward from short stem-like area. Gastral and dermal surfaces generally smooth except for regular weak radial grooves and pits of aligned canal openings. Sponge wall thin (5 mm), with prominent radial canals (1 mm in average), and straight trabs in characteristic anthaspidellid structure. Plane of trab divergence at mid wall. Dendroclones disposed in slender ladder-like series; most spicules I-shaped, forming canal walls.
Materials
One complete specimen MMNHC 13266 and a fragmentary specimen MMNHC 13265 from the Lenoir Limestone east of Lenoir City, Loudon Co., Tennessee, Upper Ordovician (Sandbian).
Remarks
The poor preservation of the collected material prevents a more precise identification and description. However, the main characteristics observed are consistent with the assignment to the genus Psarodictyum Raymond and Okulitch, Reference Raymond and Okulitch1940.
Psarodictyum magnificum Raymond and Okulitch, Reference Raymond and Okulitch1940 from Valcour Island, New York and Mingan Island, Canada (Rigby and Desrochers, Reference Rigby and Desrochers1995) has a large, discoidal form, which is slightly different from the more conical to palmate grow form of the Tennessee specimens. Psarodictyum planum Raymond and Okulitch, Reference Raymond and Okulitch1940 is smaller, palmate, with a preserved stem area, but it has significantly coarser canals.
Sponges in the family Anthaspidellidae, Miller, Reference Miller1889 (Psarodictyum Raymond and Okulitch, Reference Raymond and Okulitch1940 and Patellispongia Bassler, Reference Bassler1927) are both broad, saucer-shaped to open conical-shaped with thin walls. Both have a distinct radiate skeletal structure on both gastral and dermal surfaces. They lack the multiple openings that characterized the gastral surface of Anthaspidella Ulrich and Everett in Miller, Reference Miller1889. However, clear distinction of these two genera is lacking, and Psarodictyum and Patellispongia could be synonyms. A comprehensive study of all species assigned to both genera is needed to clarify this possible synonymy, which is beyond the scope of this study.
Family Streptosolenidae Johns, Reference Johns1994
Remarks
The family Streptosolenidae was erected by Johns (Reference Johns1994) to circumscribe genera such as Streptosolen, Lissocoelia, and Hesperocoelia that bear a much more irregular and typically finer skeleton than those of the typical Anthaspidellidae (e.g., Anthaspidella, Archaeoscyphia or Calicocoelia). Although the radial canals may still appear vertically stacked, they are often sinuous and intertwining and frequently exit into vertical, axial apochetes. In addition, the endosomal spiculation contains a larger proportion of polyclonid dendroclones or even rhizoclones, that frequently attach to each other instead of the trabs.
Johns (Reference Johns1994) also established that the dendroclones are often not horizontal where two trabs connect. Furthermore, the trabs that they connect may not be at the same depth within the sponge wall. The result is a more complex, three-dimensional skeleton than other similar genera.
Genera included in the Streptosolenidae are separated from the Anthaspidellidae by the greater irregularity of their skeleton and the greater complexity of the canal systems that result. Apart from Streptosolen, Lissocoelia, and Hesperocoelia, other typical genera included in the Streptosolenidae are: Allosacus, Aulocopium, Hudsonospongia, and Eospongia.
Genus Allosacus Raymond and Okulitch, Reference Raymond and Okulitch1940
Type species
Allosacus proxilus Raymond and Okulitch, Reference Raymond and Okulitch1940 from the Ottosee Limestone at Dickinson's Mill North of the Mendota Valley in Virginia, Tennessee and the Lenoir Limestone, Knoxville, Tennessee.
Allosacus pedunculatus new species
Figure 3

Figure 3. (1–6) Allosacus pedunculatus n. sp. (1) Lateral, ventral, and dorsal views of the holotype MMNHC 13267, a complete discoidal, pedunculated sponge, scale bar 10 mm; (2) dorsal view of the paratype MMNHC 13268 showing the shallow depression on top and the external radial grooves on the surface, scale bar 10 mm; (3) details of the previous photograph showing a closer view of the radial deep grooves, scale bar 5 mm; (4) transversal thin section showing the irregular, mostly radial spicular net, scale bar 5 mm; (5) enlargement of (4) showing the irregular distribution of varied dendroclone morphologies in the parietal walls, cut by radial canals, scale bar 1 mm; (6) lateral view of the external fringe of the sponge, showing emergence of radial canals as open rounded pits (arrows), scale bar 1mm.
Type specimens
The complete holotype, MMNHC 13267 and a paratype MMNHC 13268 are both from the Lenoir Limestone east of Lenoir City, Loudon Co., Tennessee. Upper Ordovician (Sandbian).
Diagnosis
Discoidal pedunculate sponge, with convex upper surface, marked by large, sinuous and commonly branched radial canals that converge toward a central oscular depression. Radial subsurface canals end as circular openings in the sponge margin. Skeleton composed by densely packed, dendroclone (monoclonid and polyclonid) spicules and loose spicules irregularly distributed in the walls between canals, “C-shaped” megascleres, and irregular triclonid forms.
Occurrence
Lenoir Limestone (Upper Ordovician) east of Lenoir City, Loudon, Co., Tennessee.
Description
Discoidal to irregularly hemispherical and slightly pedunculated sponge, with convex upper surface. The greatest diameter is just above the base of the sponge. The maximum sponge diameter is 5 cm, 1.2 cm high, with an oscular diameter of 1.8 cm. The upper surface is marked by large, sinuous and commonly branched radial canals that converge toward a central oscular depression. These radial horizontal canals also occur below the main surface as small inhalant canals (0.8–1 mm in diameter). Radial subsurface canals end as circular openings in the sponge margin. The spicular mesh is mainly composed by densely packed, loose or welded dendroclone spicules. Among the loose spicules irregularly distributed in the walls between canals, “C-shaped” spicules and irregular “Y-shaped” forms rarely occur (Figs. 3, 5).
Etymology
Pedunculatus from Latin, having a peduncle or stalk.
Remarks
Allosacus proxilus Raymond and Okulitch, Reference Raymond and Okulitch1940 was erected to include discoidal to mound-shape sponges with a central shallow oscular depression on top and radial canals marked as grooves in the upper surface converging towards the osculum. These characteristics are consistent with the material described in this contribution. However, in the type species, as well as in the Allosacus sp. described from Argentina (Carrera, Reference Carrera1996), the basal surface is flat to concave and completely used for attachment. This feature differs from the pedunculated morphology found in the material from the Lenoir Limestone. Furthermore, the discoidal margin in the type species and the specimen from Argentina (Carrera, Reference Carrera1996) bends downward forming the discoidal and completely attached base. This margin, in A. pedunculatus n. sp., is straight and relatively thick with marked openings for the radial canals emerging in the margin surface.
Genus Zittelella Ulrich and Everett, Reference Ulrich and Everett1890
Type species
Zittelella typicalis Ulrich and Everett, Reference Ulrich and Everett1890.
Remarks
Zittelella was not included by Johns (Reference Johns1994) in the family Anthaspidellidae nor in the family Streptosolenidae, although in the latter family he included the related genus Hudsonospongia. Finks and Rigby (Reference Finks, Rigby and Kaesler2004) retained Hudsonospongia in the Streptosolenidae, but included Zittelella within the Anthaspidellidae.
Zittelella shows similar canal structure irregularity as that seen in Hudsonospongia, but bears a more complex spiculation, at least as it was described in detail for Zittelella varians (Billings, Reference Billings1861) by Rigby and Desrochers (Reference Rigby and Desrochers1995) from Mingan Island, Canada. The material described here, from Tennessee, is similar in this respect. Furthermore, like Hudsonospongia, Zittelella shows a scalarifom, ladder-like spicule structure, but it is more irregular and scarcely marked, and the main choanosomal structure and canal walls are formed from irregularly distributed dendroclones (mainly Y and X shape). Unfortunately, the description and illustration of the spicular structure of the type species Zittelella typicalis Ulrich and Everett, Reference Ulrich and Everett1890 are not informative enough for a definitive comparison. However, Z. typicalis shows more irregular dichotomous canals and the spiculation in vertical section as illustrated by Ulrich and Everett, (Reference Ulrich and Everett1890, fig 3a) shows clearly more irregularity than the regular ladder-like structure of anthaspidellids.
Zittelella varians (Billings, Reference Billings1861)
Figure 4

Figure 4. (1–9) Zittelella varians (Billings, Reference Billings1861). (1) Lateral view of a complete specimen, MMNHC 13269, scale bar 10 mm; (2) longitudinal thin section of a complete specimen, MMNHC 13270, showing vertically stacked radial canals, scale bar 10 mm; (3) transversal thin section showing straight radial canals, scale bar 10 mm; (4) detail of (2) showing the large radial canals and the irregular skeletal net, scale bar 5 mm; (5) detail of (2) showing parietal walls with irregularly distributed dendroclones, scale bar 1 mm; (6) enlargement of (4) showing a closer view of the parietal walls, including a visible vertical parietal with a faintly marked ladder-like distribution of dendroclones, scale bar 1 mm; (7) lower part of the sponge in (2) showing the radial canals at the base and the vertical canals from the base of the sponge, scale bar 2 mm; (8) vertical canals shown in the previous photograph, separated by very thin walls with a tiny alignment of “I-shaped” monocrepid dendroclones and possibly monaxons, scale bar 2 mm; (9) transversal view, specimen MMNHC 13271, showing the base of the central cavity and the appearance of the vertical canals, scale bar 2 mm.
Type specimens
Holotype GSC 999f, h Clear Water Point, (La Grande Point) east side of Havre-Sainte Pierre, Canada (Billings, Reference Billings1861).
Occurrence
Lenoir Limestone (Upper Ordovician) East of Lenoir City, Loudon, Co., Tennessee and Mingan Island Quebec, Canada (Raymond and Okulitch, Reference Raymond and Okulitch1940; Rigby and Desrochers, Reference Rigby and Desrochers1995).
Description
Small to intermediate-size sponges ranging from conical, to expanded obconical top-shaped. Small specimens commonly pedunculated, 3 cm high and 2.9 cm in maximum diameter near the top of the sponge body. The largest specimen is open obconical, 4.3 cm high and 5.2 cm in maximum diameter near the top.
Upper surface with a shallow central depression including a moderately shallow spongocoel or oscular depression into which a cluster of vertical exhalant canals extends. External surface generally smooth, marked by shallow grooves of the radial canals on the flat top and by conspicuous pores over the rest of the sponge.
In longitudinal section, convergent horizontal canals arranged in vertically stacked regular rows, meet the spongocoel normally. The arching upward inhalant canals are ~1 mm in diameter and are vertically separated by a 2 mm thick parietal wall. Numerous vertical exhalant canals, 2 mm in diameter, extend from near the base of sponge to the base of oscular depression or spongocoel. These canals are separated by thin, one spicule thick walls, composed of monoclonid desmas and possibly monaxons.
The spongocoel is moderately deep, and occupies almost half of the sponge body (21 mm deep, measured from a specimen of 43 mm high). The maximum diameter is 25 mm located at the top of the sponge. Maximum wall thickness 15 mm, also near the top.
In transverse section, the oscular depression occupies half of the sponge diameter and is irregularly surrounded by circular openings of the axial vertical canals, 1.7–2 mm in diameter. The axial canals are separated by irregular walls that are 1–2 mm thick, but occasionally only a spicule thick.
In general, the spicular skeleton is irregular and complex, dominated by I-shaped dendroclones in slightly marked ladder-like series. This basic skeleton is superposed by a complex irregular skeleton that forms thick horizontal canal walls. The skeleton around the canals is composed of irregularly disposed desmas (I-, Y-, X-, and H-shaped dendroclones; Fig. 4.5, 4.6), and most of these spicules bear curved and irregular terminations. C-shaped dendroclones also occur sporadically (Fig. 4.5, 4.6). Spicule length variable, 0.2–1.2 mm.
Materials
Five complete specimens MMNHC 13269–13273.
Remarks
The main distinctive features of Zittelella varians (Billings Reference Billings1861) include: the overall stalked obconical to conico-cylindrical form, centrally located and moderately deep spongocoel or oscular depression into which a cluster of vertical exhalant canals extends subprismatically into circular openings, and convergent horizontal canals that are vertically stacked regular rows. Size of the spiculation and canal diameter are similar to those described for specimens from Mingan Island (Raymond and Okulitch, Reference Raymond and Okulitch1940; Rigby and Desrochers, Reference Rigby and Desrochers1995).
The type species Zitelella typicalis Ulrich and Everett, Reference Ulrich and Everett1890 is very similar in size, form, and the distribution of canals. The primary difference relates to the shape of the spongocoel, which is very shallow in Z. typicalis, forming a simple central depression, into which the vertical canals open almost directly near the top of the sponge.
Other related species of the genus, such as, Z. pannosa Raymond and Okulitch, Reference Raymond and Okulitch1940 and Z. grossa Rigby and Desrochers, Reference Rigby and Desrochers1995, show coarser spiculation and canals approximately twice the size of those seen in the Tennessee material.
Paleobiogeographic significance of the Lenoir sponges
The first attempt to analyze the global distribution of Ordovician sponges, particularly those along the Laurentian margins, was performed by Carrera and Rigby (Reference Carrera and Rigby1999). Additional studies focusing on Ordovician sponge diversification patterns also incorporated paleobiogeographic information (Carrera and Rigby, Reference Carrera, Rigby, Webby, Droser, Paris and Percival2004; Carrera and Botting, Reference Carrera and Botting2008; Muir et al., Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013). Updated information published subsequently confirms the overall paleobiogeographic pattern for sponge distribution in the Laurentian paleocontinent. Most of the more recent Ordovician sponge taxonomic studies are concentrated in areas outside of North America such as China, Korea, Morocco, and Wales with new data on protomonaxonid and orchoclad demosponges and hexactinellids (Muir et al., Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013; Botting and Muir, Reference Botting and Muir2018 and references therein).
Lower Ordovician sponge faunas have received more attention primarily because of increased study of reef systems mainly in China and Korea (Rigby et al., Reference Rigby, Nitecki, Zhu, Liu, Jiang, Cooper, Droser and Finney1995; Wang et al., Reference Wang, Deng, Wang and Li2012; Choh et al., Reference Choh, Hong, Sun, Kwon, Park, Woo, Kwon, Lee and Lee2013; Adachi et al., Reference Adachi, Kotani, Ezaki and Liu2015), confirming a global Lower Ordovician interval of reef recovery following the demise of archeocyaths. Lower Ordovician sponge faunas in North America, associated with reef structures, have been recorded in classical studies by Church (Reference Church1974), Rigby and Toomey (Reference Rigby and Toomey1978), Toomey and Nitecki (Reference Toomey and Nitecki1979), Pratt and James (Reference Pratt and James1982), and Pratt (Reference Pratt, Geldsetzer, James and Tebbutt1989). Lower Ordovician bioherms usually built by microbes, sponges, and calathids occur in a broad belt along the southern and western margins of the North American continent, constituting the Laurentian sponge-algal facies (Alberstadt and Repetsky, Reference Alberstadt and Repetski1989). Sponges associated with reef structures are recorded world-wide and are restricted to tropical or subtropical areas; with main occurrences in China and Korea (Rigby et al., Reference Rigby, Nitecki, Zhu, Liu, Jiang, Cooper, Droser and Finney1995; Wang et al., Reference Wang, Deng, Wang and Li2012; Choh et al., Reference Choh, Hong, Sun, Kwon, Park, Woo, Kwon, Lee and Lee2013; Adachi et al., Reference Adachi, Kotani, Ezaki and Liu2015; Li et al., Reference Li, Li, Wang and Kiessling2015 and references therein), Siberia (Webby, Reference Webby and Bruton1984), Baltica (Kröger et al., Reference Kröger, Hints and Lehnert2017), and the Precordillera basin of Western Argentina (Cañas and Carrera, Reference Cañas and Carrera1993; Carrera et al., Reference Carrera, Astini and Gomez2017).
Most of the diverse Middle to Upper Ordovician sponge faunas are dominated by demosponges belonging to the families Anthaspidellidae and Streptosolenidae. Middle and Upper Ordovician (Darriwilian to Sandbian) faunas from eastern North America have been described from Tennessee, Virginia, Illinois, Quebec, Newfoundland, and in the Canadian Arctic (Raymond and Okulitch, Reference Raymond and Okulitch1940; Rigby, Reference Rigby1967a, Reference Rigby1977, Reference Rigby1995; Klappa and James, Reference Klappa and James1980; Rigby and Desrochers, Reference Rigby and Desrochers1995).
Middle Ordovician (Darriwilian) sponges have been reported from western North America in Utah, Nevada, and California (Bassler, Reference Bassler1941; Greife and Langenheim, Reference Greife and Langenheim1963; Rigby, Reference Rigby1967b; Rigby and Chidsey, Reference Rigby and Chidsey1976; Johns, Reference Johns1994).
Carrera and Rigby (Reference Carrera and Rigby1999) suggested an east-west differential distribution of Middle–Upper (Darriwilian to Sandbian) Ordovician demosponges along the Laurentian margins. These authors presented a compiled data set with fifteen demosponge genera described from Middle Ordovician rocks of eastern North America. Ten of these genera were not recorded in western North America and five were considered endemics. Eastern Laurentian faunas are dominated by several species of the anthaspidellid genera Hudsonospongia, Psarodictyum, Rhopalocoelia, and Climacospongia; the streptosolenids Zittelella, Eospongia, and Allosacus, and the megamorinid Saccospongia (see Tables 1, 2 for an updated sponge distribution).
Table 1. Middle and Upper Ordovician (Floian to Sandbian) generic sponge records in North America.

Table 2. Upper Ordovician (Katian–Hirnantian) generic sponge records in North America.

Carrera and Rigby (Reference Carrera and Rigby1999) recognized that the main components of the western Laurentian sponge association show an expansion of the streptosolenids, including several species of the endemic genera Streptosolen, Hesperocoelia, and Verpaspongia. Anthaspidellids are represented by some endemic forms, such as Colinispongia and Egania, as well as the occurrence of several species of the genera Archaeoscyphia, Patellispongia, and Nevadocoelia. This dataset along with an updated compiled dataset allowed Muir et al. (Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013) to reach a similar conclusion. These authors also noticed that the Late Ordovician sponge faunas show an increasing differentiation along both margins, with the occurrence of hypercalcified sponges (Sphintozoans), which are absent from the Atlantic side but are abundant on the Pacific side, with many genera shared with New South Wales (see Carrera and Rigby, Reference Carrera and Rigby1999, 2004; Muir et al., Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013, and references therein).
The new records, described in this contribution, include genera previously recorded in the eastern North American margin, such as Allosacus, Psarodictyum, Rhopalocoelia, and Zitelella reported from Mingan Island (Canada) and New York (Raymond and Okulitch, Reference Raymond and Okulitch1940; Rigby and Desrochers, Reference Rigby and Desrochers1995). Rugocoelia Johns, Reference Johns1994, a typical genus from the western margin, is reported for the first time in Tennessee, although, a new species is erected maintaining a degree of differentiation at least at the species level.
These findings confirm the differential sponge faunal distribution along both Laurentian margins, because none of these forms has been reported from the western margin (present day orientation), even though some of these genera are recorded in nearby regions outside Laurentia, such as the Argentine Precordillera and Baltica (Carrera and Rigby, Reference Carrera and Rigby1999). In addition, these genera were reported from a new different locality with almost the same sponge association as those found along Eastern North America, expanding the area, and suggesting a homogeneous distribution along the margin.
Carrera and Rigby (Reference Carrera and Rigby1999) explained this differential distribution as related to possible physical barriers that prevented the free distribution of sponges along the Laurentian continental margin. These include factors relating to the sedimentary nature of both margins as primary physical barriers and associated differential climatic constraints.
Physical barriers may relate to the development of the Mississippi embayment and southern Oklahoma aulacogens (see Marshak and Paulsen, Reference Marshak and Paulsen1996; Thomas, Reference Thomas2006 and references therein) that may have exerted a barrier effect in faunal distributions leading to the differentiation of the two sponge associations. The same restriction was inferred for the broad distribution of the Lower–Middle Ordovician sponge-algal facies along both Laurentian margins as reported by Alberstadt and Repetski (Reference Alberstadt and Repetski1989). The presence of the deep-water facies in these areas (Ouachita facies, see Alberstadt and Repetski, Reference Alberstadt and Repetski1989) considerably narrowed the sponge-algal facies against the restricted facies developed in the epicontinental seas (Fig. 5).

Figure 5. (1) Middle Ordovician paleogeographic map showing position of Laurentia and nearby paleocontinents and terrains, AP: Argentine Precordillera (from Scotese, Reference Scotese2016). (2) Darriwilian to Sandbian paleogeographic map of Laurentia, showing main sponge localities and distribution of main lithofacies belts (after Alberstadt and Repetski, Reference Alberstadt and Repetski1989) and inferred position of the equator. (M) Mingan Island, Quebec, Canada; (V) Valcour Island, New York; (D) Trenton limestone, Dixon, Illinois; (O) Ottosee Limestone, Virginia; (T) Lenoir limestones, Tennessee; (U) Utah; (N) Nevada; (C) California.
The climatic constraint could relate to the more equatorial position of the Great Basin compared to the subtropical location of the Appalachian region (Witzke, Reference Witzke, McKerrow and Scotese1990; Hodych et al., Reference Hodych, Cox and Kosler2004; Pruss et al., Reference Pruss, Finnegan, Fischer and Knoll2010; Scotese, Reference Scotese2016, and references therein). Muir et al. (Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013) emphasized the importance of this latitudinal position of Laurentia, and recognized that the western North American faunas have not been recorded elsewhere within the Middle Ordovician, and all of its characteristic faunal elements appear to have been endemic to the region. The paleocontinental reconstructions employed by Muir et al. (Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013) locate western Laurentia within a tropical band, facing northwards towards the proto-Pacific Ocean, while the eastern Laurentia margin was positioned in a subtropical region facing southward towards the proto-Atlantic, Iapetus Ocean (Fig. 5).
By the Late Ordovician (Katian–Hirnantian), geographic sponge differentiation was more accentuated (Carrera and Rigby, Reference Carrera and Rigby1999, Reference Carrera, Rigby, Webby, Droser, Paris and Percival2004; Muir et al., Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013) with the occurrence of hypercalcified sponges in Western Laurentia (the Pacific association) in a clear subequatorial distribution and a remarkably different sponge association along the eastern margin (the Atlantic association). In this Atlantic association, new suborders (Sphaerocladina, Tricranocladina, Rhizomorina, Megamorina) and families (Hindiidae, Chiastoclonellidae, Astylospongiidae, Haplistiidae) radiated and later became common elements of Silurian sponge faunas. The tectonic uplift of the Appalachians during the Upper Ordovician likely influenced the differentiation of the Atlantic sponge assemblages. The influence of the siliciclastic input is notable on this margin since the Middle Ordovician (see Miller and Mao, Reference Miller and Mao1995). The worldwide peak in sponge diversity recorded during the late Ordovician is concentrated in areas of active tectonism and orogeny (Carrera and Rigby, Reference Carrera, Rigby, Webby, Droser, Paris and Percival2004), and this signal is very strong in the Appalachian Basin.
There is a strong direct influence of environmental conditions and paleoecology on the sponge associations in more siliciclastic facies, such as those of the eastern margin of Laurentia. This may be one of the main differential constraints for sponge distribution, other than the latitudinal distinction.
Evidence based on sponge faunas is not conclusive during the Middle Ordovician—both margins share the same two main families of the orchoclad demosponges (Anthaspidellidae and Streptosolenidae) that are only differentiated at genus level. There are no clear paleoenvironmental conditions, at the scale of this analysis, that could satisfactorily explain this sponge differentiation. However, as it was pointed out above, and in the absence of other verifiable explanations, latitudinal climatic gradients are likely a factor in generation of the “north-south” Ordovician sponge distribution (Muir et al., Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013), which would also provide a way to keep the two sponge faunas distinct.
We are aware that other biological constraints of sponges (e.g., intrinsic ecological factors of the sponge species) could also be considered in this differential distribution (see for example Carrera and Rigby, Reference Carrera, Rigby, Webby, Droser, Paris and Percival2004; Carrera and Botting, Reference Carrera and Botting2008; Muir et al., Reference Muir, Botting, Carrera, Beresi, Harper and Servais2013). However, at the scale of the present analysis, intrinsic biological characteristics, other than larval distribution, appeared to be subordinate to large-scale, climatic or physical constraints.
Acknowledgments
This paper is a contribution to the International Geoscience Programme (IGCP) Project 653—“The onset of the Great Ordovician Biodiversification Event.” This study was supported by CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas): PUE 2016 (CICTERRA – CONICET). Special acknowledgement to M. Medina and the technical team of the LABGEO (CICTERRA-FCEFyN-UNC) for thin sections preparation. G. Dinkins, McClung Museum of Natural History and Culture (MMNHC) at the University of Tennessee provided assistance with the specimens. We also thank the reviewers of the manuscript, Drs. J. Botting and E. Świerczewska-Gładysz for their helpful comments.







