1. Introduction
Pterosaurs from Mongolia are rare, compared with dinosaurs, and only a few pterosaur localities have been reported (Bakhurina & Unwin, Reference Bakhurina and Unwin1995; Unwin & Bakhurina, Reference Unwin, Bakhurina, Benton, Shishkin, Unwin and Kurochkin2000; Andres & Norell, Reference Andres and Norell2005). In the summer of 1998, the Mongol Highland International Dinosaur Project team, comprising researchers from China, Japan and Mongolia, discovered a nearly complete skeleton with skull and other fragments of pterosaurs in the Tsagan-Tsab Formation at Tatal, in the region of Kobdo, western Mongolia (Fig. 1). The Tsagan-Tsab Formation has generally been dated as latest Jurassic to Early Cretaceous (Shuvalov & Trusova, Reference Shuvalov and Trusova1976; Shuvalov, Reference Shuvalov and Martinson1982). Correlation with other localities in western Mongolia suggests an Early Cretaceous age for the Tsagan-Tsab Formation at Tatal (Bakhurina & Unwin, Reference Bakhurina and Unwin1995). The first pterosaur material from Tatal was described by Bakhurina (Reference Bakhurina1982), who described a new species of Dsungaripterus, D. parvus, on the basis of a distinct tibia. Additional specimens from the same locality (Bakhurina, Reference Bakhurina1983, Reference Bakhurina1984) confirmed their dsungaripterid affinities, but they are distinct from Dsungaripterus, so a new generic name Phobetor was erected (Bakhurina, Reference Bakhurina1986). Later, it was found that Phobetor is preoccupied and a replacement name will be published shortly (Bakhurina & Unwin, Reference Bakhurina and Unwin1995; Unwin et al. Reference Unwin, Bakhurina, Lockley, Manabe and Lü1997; Unwin & Bakhurina, Reference Unwin, Bakhurina, Benton, Shishkin, Unwin and Kurochkin2000; D. Unwin, pers. comm. to Lü, 2001). The nearly complete specimen collected from western Mongolia in 1998 provides a great deal of information leading to an increased understanding of the Mongolian pterosaur Phobetor.

Figure 1. The fossil locality of dsungaripterids. Δ indicates the fossil site.
2. Systematic palaeontology
Institutional abbreviations: GIN – Institute of Geology, Mongolian Academy of Sciences, Ulan Bataar, Mongolia; IVPP – Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy, Beijing, China.
PTEROSAURIA Kaup, Reference Kaup1834
PTERODACTYLOIDEA Plieninger, Reference Plieninger1901
Family dsungaripteridae Young, Reference Young1964
Amended diagnosis. Anterior parts of the upper and lower jaws toothless. Short rod-like parietal process extends posterodorsally, and the sagittal crest on the skull bears striations, unlike those seen in Tapejaridae, a sub-orbital foramen is formed by postorbital process of the jugal and the lachrymal.
Included genera. Dsungaripterus Young, Reference Young1964; Noripterus Young, Reference Young1973 (= Phobetor Bakhurina, Reference Bakhurina1986); Lonchognathosaurus Maisch, Matzke & Sun, Reference Maisch, Matzke and Sun2004; Domeykodactylus Martill et al. Reference Martill, Frey, Diaz and Bell2000; Puntanipterus Bonaparte & Sanchez, Reference Bonaparte and Sanchez1975; and possible Germanodactylus (Bennett, Reference Bennett2006).
Genus Noripterus Young, Reference Young1973 (= Phobetor Bakhurina, Reference Bakhurina1986)
Type species. Noripterus complicidens Young, Reference Young1973
Referred material. Dsungaripterus (Phobetor) parvus Bakhurina, Reference Bakhurina1986
Revised diagnosis. Skull with a developed sagittal crest, which begins above the interval between the 7th and the 8th tooth position (from anterior to posterior) of the upper jaw, extending posteriorly along the midline of the skull and terminating above at the level of the middle of the dorsal rim of the orbit. Anterior toothless parts of both jaws straight. Ratio of the length of the mandibular symphysis to that of the lower jaw approximately 0.54. Deep groove on the midline of the dorsal surface of the dentition part of the mandibular symphysis. Teeth laterally compressed with sharp tips. Thirty teeth on the upper jaw and 20 teeth on the lower jaw. Six teeth on upper jaw below the margin of the nasoantorbital opening. The alveoli are not expanded into protuberances. The dentition in the upper jaw extends about one-third further posteriorly than that of lower jaw. Ratio of tibia to femur length is approximately 1.7.
Referred specimens. One nearly complete skeleton with a skull (GIN125/1010) (Figs 2, 3a, b; Tables 1, 2); one incomplete skull (GIN125/1010-1) (Fig. 3c) and other specimens from one quarry (Fig. 5). Specimens GIN125/1010-2, GIN125/1010-3 and GIN125/1010-4 belong to a large pterosaur, while other, smaller bones belong to a single small individual, therefore, GIN125/1010-2, GIN125/1010-3 and GIN125/1010-4 most likely belong to one large individual and are assigned to Dsungaripterus weii, and the small-sized individual is assigned to Noripterus complicidens.

Figure 2. Photograph (a) and outline drawing (b) of Noripterus (GIN125/1010). Abbreviations: cap – carpus; cv – cervical vertebrae; d – dentary; dr – dorsal ribs; f – femur; h – humerus; j – jugal; mc – manual claw; mx – maxilla; nof – nasoantorbital opening; or – orbit; pc – parietal crest; pt – pteroid; pm – premaxilla; rd – radius; sc – scapula; ti – tibia; ul – ulna; wm – wing metacarpal; wph1–wph2 – wing phalanx 1 and wing phalanx 2.

Figure 3. (a) The skull of Noripterus (GIN125/1010), (b) a close up of its dentition and (c) partial skull of Noripterus (GIN125/1010-1a). Abbreviations: j – jugal; nof – nasoantorbital opening; pm+m – premaxilla and maxilla; q – quadrate; qj – quadratojugal; sc – saggital crest; ts – tooth sockets; Scale bars = 50 mm.
Table 1. Measurements of the skull of Noripterus (GIN125/1010)

Notes: pre – preserved length; est – estimated length.
Table 2. Measurements of the postcranial skeletons

Locality and horizon. Tatal, western Mongolia. Tsagan-Tsab Formation, Late Jurassic–Early Cretaceous (Shuvalov, Reference Shuvalov and Martinson1982) (early Early Cretaceous, Bakhurina, Reference Bakhurina1982). GPS data: 48°45′15″N, 92°35′52″E; Bone bed GPS data: 48°45′30″N, 92°34′42″E.
3. Description
Skull. The skull is eroded around the braincase, but its outline is distinctive except for a small posterior part. The length of the skull is estimated to be 210 mm from the rostral tip to the distal end of the sagittal crest. The ratio of the long axis of the nasoantorbital opening to the length of the ventral margin of the skull is approximately 0.36. The ventral margin of the skull is straight in lateral view. The dorsal margin of the skull, which forms a distinct sagittal crest (midline crest) on its dorsal part, displays a wavy outline in lateral view (Figs 2, 3a, b), similar to that of other dsungaripterids, such as Dsungaripterus weii (Young, Reference Young1973) and ‘Phobetor’ (Bakhurina, Reference Bakhurina1984). The sagittal crest is preserved as thin fragmentary bone. It originates dorsal to a position between the 7th and 8th tooth of the upper jaw (counting from the tip of the upper jaw) and extends posteriorly, terminating at about the level of the posterodorsal corner of the nasoantorbital opening. A rod-like parietal process is preserved as an impression, similar to the parietal posterior process of Dsungaripterus weii (Young, Reference Young1973). The anterior margin of the sagittal crest is nearly vertical, and the anterior part of the sagittal crest is highest, gradually becoming lower posteriorly. The highest part is at the level of the tooth positions 8–15 of the upper jaw. The antorbital and nasal opening are confluent, forming a large triangular nasoantorbital opening. The posterior margin of the nasoantorbital opening is straight. The long axis of the opening is about 65 mm. Teeth are absent from the anterior parts of the upper and lower jaws for approximately one-third of the length of the jaws.
There is a distinct, nearly horizontally oriented groove on the anterolateral part of the upper jaw, which may indicate the suture between the maxilla and the premaxilla (Fig. 3a, b). The premaxilla is located dorsal to the suture and it extends posteriorly to contact with the crest, and anteriorly to the tip of the upper jaw. The suture between the nasal and the premaxilla is unclear. Although the tip of the upper jaw is missing, what is left of the premaxilla indicates that the tip was pointed. The rostral end of the premaxilla and maxilla is straight.
The maxilla almost forms the whole lower margin of the upper jaw, except for the posterior part, which should be the jugal, but the suture between the maxilla and the jugal is not clear. The maxilla bears 15 teeth on its posterior portion, but there is no tooth on its anterior part. The distance between teeth is greater in the middle portion of the dentition than in the anterior and posterior portions of the upper and lower jaws, where the space between teeth is shorter, about 3.4 mm. The space between two teeth is approximately equal to the anteroposterior width of one tooth. The alveoli are not expanded, unlike those of Dsungaripterus, which are bulbous (Young, Reference Young1964, Reference Young1973). The anterior eight teeth are larger than the posterior seven teeth, and the third and the fourth teeth are the largest. The teeth are laterally compressed, and triangular in lateral outline. Unlike those of Dsungaripterus, the teeth bases of the present specimen Noripterus are narrow and much elongated. The tips of the teeth are blunt and the tips of the anterior six teeth point posteroventrally, while those of the posterior nine teeth point anteroventrally. All the teeth are located in the middle portion of the upper jaw. Both the anterior and posterior margins of the teeth are smooth. The posterior portion of the dentition bears small teeth and the spaces between the teeth are moderate, unlike the condition in Dsungaripterus weii, where two or three teeth are closely implanted (Young, Reference Young1973). There is no groove or ridge on the anterior portion of the palatal surface. The posterior part of the lower margin of the upper jaw is a very thin plate-like bone, most likely the jugal, but the suture between the jugal and the maxilla is unclear. The quadratojugal is wider than the jugal, but the suture between the quadratojugal and the quadrate is unclear. Only the lower portion of the quadrate is preserved, but it is sufficient to show that its position relative to the ventral margin of the skull is inclined about 120°.
The orbit is round (Fig. 3a), and its upper margin is well preserved. A round opening is present in the front of the anterodorsal corner of the orbit, which may represent a lachrymal opening. The bones around the lower and posterior margins of the orbit are crushed, obscuring the true shapes of the orbit and lower temporal fenestra. The dorsal margin of the orbit and the highest point of the nasoantorbital opening are at the same level.
Lower jaw. The lower jaw is relatively well preserved and bears ten teeth located mainly in the middle portion of the jaw. The dentary forms most of the lower jaw and has a ventral outline lacking a crest. There is no distinct dorsal coronoid eminence at the posterior end of the mandible either. The position of the articulation between the skull and mandible is almost under the anteroventral corner of the orbit, although this may be slightly exaggerated due to the preservation.
The tip of the lower jaw is toothless, straight and pointed. The sharply pointed teeth are of almost equal size. The mandibular symphysis bears seven teeth pairs. Posterior to the last tooth, both the upper and the lower margins of the lower jaw are nearly parallel, while the portion which bears the first five teeth is the deepest. In lateral view, the ventral margin of the lower jaw is slightly curved upwards from the position of the first tooth, while the upper margin of the lower jaw is nearly straight (Fig. 3). The mandibular symphysis is long, comprising slightly more than half the jaw length (the ratio is 0.54). A deep sagittal groove appears on the middle of the dorsal surface of the tooth-bearing part of the mandibular symphysis (Figs 3b, 4e), as in the holotype of Noripterus (Young, Reference Young1973) (Fig. 4f, g). Anterior to the dentition, a low sharp ridge appears on the middle of the dorsal surface of the mandibular symphysis (Fig. 4e). It is not clear whether this sharp ridge is present in the holotype of Noripterus or not, due to the absence of this portion.

Figure 4. (a) Right femur (GIN 125/1010-1c) of Noripterus in anterior view. (b) Right tibia and fibula of Noripterus (GIN125/1010-2). (c) Right wing metacarpal of the holotype Dsungaripterus weii. (d) New specimen of Dsungaripterus weii (GIN125/1010-3). (e) Comparison of the lower jaws of Noripterus (GIN125/1010) and the (f) photograph and (g) outline drawing of holotype (IVPP, V. 4062) in dorsal view. Abbreviations: dg – deep groove; sr – sharp ridge. Scale bar = 50 mm in (c) and (d); 20 mm in (e) and (f).
Axial skeleton (Fig. 2). Only four vertebrae are distinctively visible, probably representing the second to fifth cervicals. They are of similar size. The ventral surfaces of these four cervical vertebrae are flat. A postexapophysis is present. The neural spine is not preserved. There is a distinct lateral pneumatic foramen on the centra of the fourth and fifth cervical vertebrae. There is no evidence of cervical ribs.
Dorsal vertebrae. Five proximal ends of the stout dorsal ribs are preserved with their associated vertebrae, which is not well preserved. The neural spines of the anterior dorsal vertebrae are fused into a plate-like structure, and although the exact shape of the dorsal vertebrae is unclear, the fused neural spines show that their centra were fused into a notarium, similar to that of Dsungaripterus weii (Young, Reference Young1964, Reference Young1973).
Scapula (Fig. 2). Both scapulae are poorly preserved. They are plate-like bones and elongate. The shaft of the scapula is curved medially. The width of the shaft is about 11 mm. Both ends are not well preserved, and their shapes are unclear.
Humerus (Fig. 2). Both humeri are present. The left one is well preserved, while the right is damaged. The proximal end is extensively expanded and consists of a large humeral head and a well-developed deltopectoral crest. A pneumatic foramen is lacking on the proximal end of the humerus. The proximal end is much more expanded than the distal one. In posterolateral view, the shaft of the humerus is slightly curved, with the middle part of the shaft narrowest. The posterolateral surface under the humeral head is slightly concave, but the surface of the deltopectoral crest is slightly convex. The margin between the deltopectoral crest and the humeral head is curved, but the margin of the humeral head is straight. The articular surface of the humeral head is expanded and convex. The distal end of the humerus is expanded, and D-shaped in distal view. The ulnar condyle of the humerus is large and there is a sharp margin above the lateral margin of the ulnar condyle, which is wing-like in shape, and may be the insertion area for M. flexor carpi ulnaris.
Ulna and radius. The proximal ends of both bones are not well preserved. Towards the distal end, both bones are almost parallel. The radius is weaker than the ulna. Both bones seem to be fused distally. The radius has an elongate opening on the distal anterior surface.
Pteroid. The pteroid is nearly complete and is a slender, rod-like bone, about one-half the length of the ulna. Its proximal end is tightly contacting the distal end of the radius, thus the morphology of the articulation is unclear. The proximal end is wider and plate-like and its anterior surface is round and convex, while the posterior surface bears a deep groove, which may be the area for inserting the wing membrane.
Carpals. Carpals are naturally articulated with the distal ends of the ulna and the radius. Distal carpals are fused into a rectangular unit.
Metacarpus. Metacarpals I–III do not reach the distal carpals, but they occupy approximately half the length of the metacarpal IV. Digit II is almost complete, and the lengths of phalanx 1 and phalanx 2 are nearly the same. The proximal end of phalanx 1 is larger than its distal end. Phalanx 2 is much more slender than phalanx 1. The claw is heavily curved and its articular end is much larger than the distal articular end of phalanx 2, which would have provided the claw with greater mobility. There is a deep groove along the lateral surface of the claw, as in other pterosaurs.
Digit IV. Only two phalanges are preserved. The first phalanx is well preserved on the left side, but only the middle part of the right bone is preserved. It is longer than the wing metacarpal. Both ends are expanded. The extensor tendon process is fused into the proximal end of the first wing phalanx, indicating that it is an adult individual (Frey & Martill, Reference Frey and Martill1998). The lower part of the proximal end forms a concave articular surface, which contains the distal end of wing metacarpal IV. The distal end of the first wing phalanx has a ‘foot-like’ extension, and its more obtuse ‘heel’ ends anteriorly. The more acute ‘toe end’ is oriented posteriorly. The second wing phalanx only lacks a small portion distally. It is more slender than the first one, with the proximal end much expanded. The ventral surface is slightly more convex than the dorsal surface. Ventrally, there is no longitudinal ridge along the shaft.
Femur. Only the right femur is preserved, and the femoral head is missing. The femoral shaft is nearly constant in width except for the slightly expanded distal and proximal ends. In lateral view, the femoral shaft is only slightly posteriorly curved.
Tibia and fibula. Both the left and right tibia and fibula are preserved. The tibia is about 1.7 times as long as the femur. The proximal end of the tibia is much wider than its distal end, which is only slightly expanded. The fibula is much weaker and smaller than the tibia, and its proximal end is widened anteroposteriorly. It strongly contacts the lateral surface of the tibia, and becomes rod-like and slender distally, where it is fused to the tibia.
Tarsal. The tarsal is located near the distal end of the right tibia, but it does not articulate with the tibia. It is irregular in shape and distinctly separated from the tibia.
4. Other specimens collected from the same area
4.a. A partial skull and postcranial elements
There are several postcranial elements and a partial skull from one quarry (Table 3). No repeated bones except one large cervical vertebra and an incomplete larger wing metacarpal were discovered in the quarry. Except for the large cervical vertebra and the wing metacarpal IV, which clearly belong to a larger individual, all other material appears to belong to one relatively small individual.
Table 3. Measurements of GIN 125/1010-1

4.a.1. Partial skull
A partial skull (GIN125/1010-1a), which is missing its posterior part and the tip of the anterior part, is preserved (Fig. 3c). Visible sutures between the maxilla/jugal and maxilla/premaxilla are missing. There are nine tooth sockets preserved in the maxilla. The shape of the tooth sockets is elongate, indicating that the tooth base was narrow and elongate, unlike that of Dsungaripterus weii, in which the tooth base is broad and sub-oval (Young, Reference Young1964, Reference Young1973). The tooth socket is wider anteroposteriorly than lateromedially. The nasoantorbital opening is nearly triangular. The anterior margin of the opening is arched. The broken surface of the sagittal crest shows that the crest is trabecular, and filled with small cavities, enclosed by bony struts. About 11 teeth are sparsely distributed among the all bones. The teeth are laterally compressed with a slight medial curvature. All the tooth crowns are smooth. The tips of the teeth are moderately sharp. Considering that no teeth are preserved in situ in the partial skull (GIN125/1010-1a), these teeth are assigned to this specimen.
4.a.2. Postcranials (GIN125/1010-1b) (Fig. 5; Table 3)
Except for the relatively small size, the characters of the bones are similar to those of specimen GIN125/1010. Descriptions of elements which are not present in the specimen GIN125/1010 are added, based on specimen GIN125/1010-1b.

Figure 5. (a) Postcranial skeleton of different individual of Noripterus (GIN125/1010-1b). (b) Close-up of teeth scattered among bones.
Both specimens GIN125/1010 and GIN125/1010-1b are assigned to Noripterus, based on the morphologies of the bones. Several teeth are scattered among the bones. They are identical to the teeth of the specimen (Fig. 5b) GIN125/1010. The wing metacarpal is straight. Both wing metacarpals display a distinct roughened area present on the ventral margin, which is 14 mm away from the distal end of the wing metacarpal. A similar roughened area is also present on the wing metacarpal of Dsungaripterus weii (Young, Reference Young1964). The proximal end of the wing metacarpal is expanded dorsoventrally. A distinct crest is present on the ventral margin of the proximal end. The extensor process is fused into the proximal end of the first wing phalanx, indicating that this animal is at least sub-adult. The radius is straight, with expanded proximal and distal ends. The articular surface of the distal end is clear. The ulna is missing its middle portion, but its distal and proximal ends are preserved. Its distal end is much wider than that of the radius, the ratio of the diameter of the radius to that of the ulna being approximately 0.95. Wing phalanx 3 is straight with suboval cross-section of the shaft. Wing phalanx 4 also displays a straight shaft, although it is broken into four small parts. A partial right tibia and fibula are preserved, but their proximal ends are missing. The distal end of the fibula is fused into the tibia.
4.a.3. Right femur (GIN 125/1010-1c) (Fig. 4a)
This femur is almost complete and is 120 mm long. The upper part of the shaft is slightly curved laterally and posteromedially. The lower one-third of the shaft is straight. There is a distinct neck between the femoral head and the shaft, and the caput is steeply directed. The great trochanter is well developed. The femoral head is not ball-shaped, and it is wider lateromedially than anteroposteriorly. In dorsal view, the middle portion of the femoral head is widened. It seems that there is a large opening on the anterior surface of the proximal end, which may extend into the femoral head. There is a shallow groove on the anterior surface of the distal end, dividing it into two condyles, and the medial one is larger than the lateral one.
4.b. Right tibia and fibula (GIN125/1010-2) (Fig. 4b)
The tibia, as preserved, is 109 mm long, missing only its distal end. It is heavily crushed. The proximal end of the tibia is expanded. The articular surface for the distal end of the femur is sub-oval. There is a weak groove along the posterior surface of the tibia, which extends to its distal end. The proximal end of the fibula is thin and plate-like and expanded anteroposteriorly, but it becomes rod-like, extending distally, then fuses to the tibia, as in Noripterus complicidens (Young, Reference Young1973). The width of the proximal end of the fibula is the same as that of the tibia. The articular end of the tibia is slightly concave, but in anteroposterior view, the margin of the proximal end is horizontal, unlike that of the holotype of Phobetor (Bakhurina, Reference Bakhurina1982, Reference Bakhurina1984), in which the articular surface of the proximal end of the tibia is oblique.
4.c. Right wing metacarpal (GIN125/1010-3) (Fig. 4d)
One right wing metacarpal IV is preserved; it is 211 mm long and 13 mm wide anteroposteriorly, and 7.5 mm wide dorsoventrally. It is the largest among the Tatal pterosaur metacarpals, compared with the shortest one, which is 128 mm long. The proximal end of the wing metacarpal in GIN125/1010-3 is missing, but the most proximal part preserved is wider than the distal. The shaft is nearly straight. Although the bone is crushed, the outline of the proximal part is clear. Both the dorsal and ventral surfaces are round and slightly convex. Near the distal end, the ventral surface is also flat, while the dorsal surface is convex. The cross-section of the shaft is quadrilateral. The thickness of the anterior and posterior walls of the bone is the same, about 1.2 mm thick. The ventral wall of the bone is thicker than the dorsal wall. The thickness of the ventral wall is 2.86 mm, while that of the dorsal wall is 1.9 mm. Distally, there is a deep groove formed by the condyles on the articular part, which is articulated with the first phalanx of the wing finger IV. The dorsal condyle is larger than the ventral condyle and it extends much further posteriorly than the dorsal condyle. There is a distinct and roughened area on the dorsal margin near the distal end (Fig. 4d). The distal margin of this area is 30 mm from the distal end of the bone. The long axis of this roughened area is 12.3 mm; it may represent the attachment site for the wing membrane, and is very similar to the condition seen in Dsungaripterus weii (Young, Reference Young1964). The wing metacarpal is identical to that of Dsungaripterus weii (Fig. 4c) in both size and shape, therefore it is assigned to this taxon.
4.d. Fourth cervical vertebra (GIN125/1010-4)
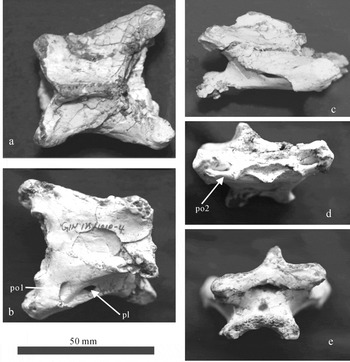
This cervical vertebra is almost complete, but exhibits some slight dorsoventral crushing (Fig. 6; Table 4). The proximal end is much wider than its posterior end and the ventral surface is flat (Fig. 6a, b). It compares well to the fourth cervical of Dsungaripterus weii (Young, Reference Young1973), therefore it is regarded as a fourth cervical vertebra. The prezygapophysis is stout and well developed, with the facet of the prezygapophysis oval and facing dorsomedially. The prezygapophysis extends far anteriorly to the anterior margin of the articular end. The anterior articular end of the centrum is concave and triangular (Fig. 6d). The dorsal margin of the anterior articular end is nearly straight. A distinct preexapophyseal articulation is located laterally to the anterior articular surface, which faces ventromedially. A pneumatic opening is present between the bases of the prezygapophysis and the preexapophyseal articulation (Fig. 6d, po2). The neural spine is low. The pleurocoel is large and is located slightly to the middle of the centrum (Fig. 6b, pl). Anteroventral to the pleurocoel, a pneumatic opening is also present near the base of the prezygapophysis (Fig. 6b, po1). The postzygapophysis is also developed. The facet of the postzygapophysis faces ventrolaterally. An epipophysis is present on the dorsal surface of the postzygapophysis. There is no hypapophysis on the ventral surface of the anterior end of the centrum. The ventral surface of the anterior end is concave, thus the posterior articular end is crescent-shaped (Fig. 6a). The postexapophysis is present on the lateral face of the articular end, but it is less developed than the preexapophysis. The size and morphology of this cervical vertebra are identical to those of Dsungaripterus weii, thus this cervical vertebra is assigned to Dsungaripterus weii.

Figure 6. The fourth cervical vertebra of Dsungaripterus weii (GIN125/1010-4) in (a) dorsal, (b) ventral, (c) lateral, (d) anterior and (e) posterior views. Abbreviations: pl – pleurocoel; po1 – pneumatic opening 1; po2 – pneumatic opening 2.
Table 4. Measurements of the fourth cervical vertebra of Dsungaripterus weii (GIN 125/1010-4)

5. Discussion
The nearly complete specimen collected from western Mongolia in 1998, and described here, is referable to the pterosaur taxon Phobetor, but a detailed comparison of its lower jaw with that of the holotype of Noripterus shows that they are almost identical, and thus the pterosaur Phobetor is considered a junior synonym of Noripterus.
The distal end of the sagittal crest of Noripterus is rod-like, representing a special posterior process of the parietal, as is the case in Dsungaripterus (Young, Reference Young1973) and Phobetor (Bakhurina & Unwin, Reference Bakhurina and Unwin1995). The dorsal margin of the sagittal crest is wave-like in lateral view, similar to the condition seen in Dsungaripterus weii, and this sagittal crest originates above the interval between the 4th and 5th tooth positions of the upper jaw (Young, Reference Young1964, Reference Young1973). The suture between the maxilla and the jugal, like most other skull connections in pterosaurs, is not clear (Padian, Reference Padian1983).
The radius is weaker than the ulna in GIN125/1010. Both bones seem to be fused at their distal end, unlike those in Dsungaripterus weii (Young, Reference Young1973). The proximal end is wider and plate-like. Its anterior surface is round and convex, while the posterior surface has a deep groove, which may be the area for inserting the wing membrane, as is the case in the wing phalanx of Rhamphorhynchus (Wellnhofer, Reference Wellnhofer1991). Both ends of the first phalanx of digit IV are expanded. There is a well-developed olecranon-like process at the proximal end as in Dsungaripterus weii (Young, Reference Young1964). Ventrally, there is no longitudinal ridge along the shaft as is seen in Dsungaripterus weii (Young, Reference Young1964). In lateral view, the femoral shaft is only slightly curved posteriorly, similar to the holotype of Noripterus complicidens (Young, Reference Young1973), but unlike the condition in Dsungaripterus weii, where the femur is strongly posteriorly curved. The tarsal is irregular in shape and unfused to the tibia, unlike its condition in Dsungaripterus, where it is fused (Young, Reference Young1973); however, this may indicate that it is sub-adult.
The sagittal crest occurs only in three families of Pterodactyloidea: Ctenochasmatidae (Buisonjé, Reference Buisonjé1981), Dsungaripteridae (Young, Reference Young1964; Wellnhofer, Reference Wellnhofer1991) and Azhdarchids (Martill & Naish, Reference Martill and Naish2006). All of these have a premaxillary sagittal crest which extends posteriorly, as does that of Tapejara, Tupuxuara and Thalassodromeus (Wellnhofer, Reference Wellnhofer1987; Wellnhofer & Kellner Reference Wellnhofer and Kellner1991; Kellner & Campos, Reference Kellner and Campos2002; Martill & Naish, Reference Martill and Naish2006). Noripterus (GIN125/1010) ( = Phobetor) is different from Ctenochasma in that Noripterus (GIN125/1010) does not have premaxillary teeth, and the tips of the upper and the lower jaws are straight. The relatively short teeth are laterally compressed, have a sub-circular cross-section and are only present on the maxilla, while in Ctenochasma, there are slender and numerous teeth in the premaxilla and the maxilla, and the teeth have a circular cross-section (Buisonjé, Reference Buisonjé1981). Noripterus (GIN125/1010) has fewer teeth than Ctenochasma, 15 teeth in the upper jaw and 11 teeth in the lower jaw on each side in Noripterus (GIN125/1010), while Ctenochasma has more than 100 long, delicate teeth.
Noripterus (GIN125/1010) differs from Dsungaripterus in that the tips of both upper and lower jaws in Dsungaripterus are curved dorsally, the teeth are blunt (knob-like) and usually the bases of the posterior three or four are very close, fusing into one, separating them from other teeth, while in Noripterus (GIN125/1010), the tips of both upper and lower jaws are straight, and the teeth are compressed lateromedially. The tips of the teeth are sharp. The upper jaw and lower jaw tooth numbers of Dsungaripterus and Noripterus (GIN125/1010) are almost identical (14 upper jaw teeth and 11 lower jaw teeth in Dsungaripterus (Young, Reference Young1973), while there are 15 teeth in the upper jaw and 11 teeth in the lower jaw in Noripterus (GIN125/1010)).
Noripterus (GIN125/1010) differs from Germanodactylus in that Germanodactylus has premaxillary teeth while Noripterus (GIN125/1010) does not. The sagittal crest appears above the orbit and extends posteriorly to form a parietal process in Noripterus (GIN125/1010). The orbit is larger and located below two-thirds maximum height of the nasoantorbital opening.
Noripterus (GIN125/1010) differs from Huanhepterus in that Noripterus has relatively shorter cervical vertebrae, while Huanhepterus has long cervical vertebrae; Huanhepterus has premaxillary teeth, and the tip of the lower jaw also has teeth, while Noripterus (GIN125/1010) does not have premaxillary teeth, and the tip of the lower jaw is toothless.
At present, six genera of dsungaripterid pterosaurs have been reported: Dsungaripterus (Young, Reference Young1964), Noripterus (Young, Reference Young1973; = Phobetor Bakhurina, Reference Bakhurina1986); Lonchognathosaurus (Maisch, Matzke & Sun, Reference Maisch, Matzke and Sun2004); Domeykodactylus (Martill et al. Reference Martill, Frey, Diaz and Bell2000), ‘Santanadactylus’ spixi (Bennett, Reference Bennett1994) and Puntanipterus (Bonaparte & Sanchez, Reference Bonaparte and Sanchez1975). The new material of Noripterus differs from Dsungaripterus (Young, Reference Young1973) in the relatively small size and the straight toothless jaw tips and more maxillary teeth (15 teeth) in Noripterus, while Dsungaripterus is larger, and has toothless jaw tips which are curved dorsally, and fewer (12) maxillary teeth. This difference is also seen in the holotypes of Noripterus and Dsungaripterus (Young, Reference Young1973).
Although there are some similarities between Noripterus (GIN125/1010) and Lonchognathosaurus (Maisch, Matzke & Sun, Reference Maisch, Matzke and Sun2004), such as the straight toothless jaw tips and the less expanded alveoli, their distinctness is clear. Fifteen maxillary teeth are present in Noripterus (GIN125/1010), while the number of the maxillary teeth of Lonchognathosaurus is only eight. Although the tooth row starts far anterior to the sagittal crests in both forms, the tooth row ends anterior to the nasoantorbital opening in Lonchognathosaurus (Maisch, Matzke & Sun, Reference Maisch, Matzke and Sun2004). However, the tooth row extends far posterior to the nasoantorbital opening in Noripterus (GIN125/1010). Therefore, Noripterus (GIN125/1010) is distinct from Lonchognathosaurus.
Domeykodactylus (Martill et al. Reference Martill, Frey, Diaz and Bell2000) is only preserved as a partial lower jaw. Noripterus (GIN125/1010) differs from Domeykodactylus (Martill et al. Reference Martill, Frey, Diaz and Bell2000) in that the lower jaw of Noripterus (GIN125/1010) bears ten, widely spaced teeth while that of Domeykodactylus bears at least 16, closely spaced teeth. The dental alveolar borders are much more expanded in Domeykodactylus than in Noripterus (GIN125/1010). These characters allow Domeykodactylus and Noripterus (GIN125/1010) to be easily distinguished.
It is difficult to compare Noripterus (GIN125/1010) with Puntanipterus, which is only preserved as a single tibia and corresponding fibula (Bonaparte & Sanchez, Reference Bonaparte and Sanchez1975). Although the proximal end of the tibia is oblique in Puntanipterus, while it is horizontal in Noripterus (GIN125/1010-2), the morphology of the tibia–fibula of Puntanipterus and Noripterus (GIN125/1010-2) (Fig. 4b) is similar.
6. Palaeoecological implications
Although material of Dsungaripterus is less abundant than that of Noripterus in the Tatal region, the preserved parts such as the cervical vertebra and a large metacarpal IV, which are almost identical to those of Dsungaripterus weii, indicate that it can reach the same body size as Dsungaripterus from Xinjiang with a wing span up to 5 m (Young, Reference Young1973). However, the body size of Noripterus is much smaller, and its wing span is less than one-third that of Dsungaripterus. As Young (Reference Young1973) mentioned, these two forms (Dsungaripterus and Noripterus) may have coexisted for a long time. The smaller one, Noripterus, either must have had an advantage over the large-sized Dsungaripterus (Young, Reference Young1973), or they may have had different living strategies. The tooth morphologies and the dentitions are different between these two forms; the teeth are short and stout with distinctly expanded alveoli and the posteriormost two or three closely connected in Dsungaripterus: this morphology not only allows piscivory, but also deals with hard-shelled animals (Young, Reference Young1973). The teeth of Noripterus are relatively weak, slender and flat mediolaterally, which probably worked well for fish eating, but not for dealing with hard-shelled animals. Dsungaripterus may have obtained its food from shallow parts of lakes, where shelled invertebrates may have been abundant, while Noripterus may have gone further towards the lake centre in search of fish.
Acknowledgements
We are indebted to Dr Eric Buffetaut (Paris, France), who read the first draft of this paper and made constructive suggestions. We would like to acknowledge the members of the Mongol Highland International Dinosaur Project. This project is supported by Chunichi Shinbun Co. Ltd., Kyoto Kagaku Co. Ltd., Chukyo TV Broadcasting Co. Ltd., and Tokai Bank Ltd. This research is partly supported by the National Key Basic Research and Development Program (Grant 2006CB701405).