Introduction
Alligatorweed [Alternanthera philoxeroides (Mart.) Griseb.] is a noxious invasive weed belonging to Amaranthaceae that originated in the Parana River region in South America, but is now extensively distributed in almost all continents (Liu et al. Reference Liu, Horvath, Li and Liu2019). This stoloniferous and amphibious perennial invasive weed greatly threatens biodiversity, ecosystems, and agricultural production in the invaded areas (Fan et al. Reference Fan, Yu, Liu, Yu, Han and Wang2015; You et al. Reference You, Han, Fang and Du2016). Under dry terrestrial conditions, this invasive weed can form very dense stands and displace native communities (Wang et al. Reference Wang, Li and Wang2005a), and these dense stands can block cropland irrigation and drainage systems. In aquatic conditions, it can cover the entire water surface, which may decrease oxygen exchange, prevent water flow, and even increase flood damage (Wang et al. Reference Wang, Li and Wang2005a). Clonal integration and phenotypic plasticity were proposed as important traits that can facilitate the successful adaptation of A. philoxeroides to a wide range of environmental conditions (Dong et al. Reference Dong, Alpert, Zhang and Yu2015). For example, in aquatic habitats, leaves are large, with long internodes and wide cavities, whereas in terrestrial habitats, leaves are small, with short internodes and thin cavities (Gao et al. Reference Gao, Geng, Yang, Hu and Yang2015; You et al. Reference You, Han, Fang and Du2016). Moreover, under drier terrestrial conditions, adventitious roots of A. philoxeroides will grow long, thick, and fleshy to form storage roots. Storage roots have been proposed to be beneficial for propagation when grown in dry habitats, because the aboveground parts will die during cold winters, but the storage roots will survive and produce adventitious roots and buds in the following year (Gao et al. Reference Gao, Geng, Yang, Hu and Yang2015; Lou et al. Reference Lou, Wang, Deng and Wei2003).
The formation and development of storage roots in plants is a complex biological process that involves morphogenesis and dry matter accumulation (Sun et al. Reference Sun, Xiao, Duan, Guo, Qi, Liao, Zhao, Liu, Zhou and Li2015). Carbohydrates (e.g., sucrose, starch) and phytohormones (e.g., cytokinins, indole-3-acetic acid, ethylene) play crucial roles in the formation of storage roots (Ravi et al. Reference Ravi, Chakrabarti, Makeshkumar and Saravanan2014), but the changes in carbohydrate and hormone levels during A. philoxeroides storage root formation are unclear. Furthermore, a lack of genomic information of A. philoxeroides restricts our ability to characterize genes that relate to phenotypic variation and genes involved in environment acclimatization regulation.
High-throughput sequencing has been widely used to identify differentially expressed genes (Zhu et al. Reference Zhu, Jia, Yang, Xia, Zhang, Jia, Zhou, Nie, Yin, Ma and Liu2019c) and is particularly useful for those species without genomes available (e.g., non-model plant species; Lin et al. Reference Lin, Wu, Lin, Zhang, Xu, Wang, Wu, Chen, Lin, Lin, Wang, Zhao and Wu2018). In A. philoxeroides, several RNA-sequencing (RNA-seq) projects have been performed in different tissues under various treatment conditions. For instance, Li et al. (Reference Li, Xu, Wang, Pan and Lu2015) performed high-throughput RNA sequencing to screen the A. philoxeroides gene regulatory network in the root under low potassium stress. Gao et al. (Reference Gao, Geng, Yang, Hu and Yang2015) revealed the genome-wide transcriptional responses of A. philoxeroides (aerial tissues) to altered hydrological conditions. Genome-wide transcriptional studies provide mechanistic understanding of the genetic mechanisms responsible for biological processes and phenotypic plasticity in this stoloniferous and amphibious perennial species. However, physiological changes between adventitious and storage roots and corresponding gene reservoirs responsible for the formation of storage roots remain unclear.
In this study, changes in sugar and hormone content between adventitious and storage roots were characterized to reveal their roles during the formation of storage roots. Moreover, to explore key genes associated with sugar metabolism and phytohormone signal transduction, four sets of A. philoxeroides RNA-seq data were downloaded from the Sequence Read Archive (SRA) category of the National Center for Biotechnology Information (NCBI) database and used as input to carry out a de novo transcriptomic assembly. The expression patterns of candidate genes were further validated using quantitative reverse transcriptase polymerase chain reaction (qRT-PCR). Our study will provide useful information concerning changes in critical substances during the root-swelling process and a gene resource for elucidating the environmental acclimation mechanism.
Materials and Methods
Plant Materials and Growth Conditions
Alternanthera philoxeroides samples were harvested from a test field at the Yangtze University (Jingzhou, China). Vegetative parts of at least 150 plants were sampled and cut so as to produce 5-cm-long nodal segments. The 5-cm nodal segments were then grown in bowls (5.5-cm diameter by 14.5-cm height) containing sandy loam soil (sandy soil:loam soil = 1:1 [v/v]) in the university greenhouse. The cultivation temperature was set to 28/25 C with a 14/10-h day/night cycle. The basal section of each sample was 2.5-cm long, and the apical section was 2.5-cm long. After 20 and 70 d of growth, the plants grew adventitious roots (diameter < 1.5 mm) and storage roots (diameter > 1.5 mm), respectively (Figure 1A and B). Adventitious root and storage root samples were collected, frozen in liquid nitrogen, and stored at −80 C for further analysis.

Figure 1. Phenotype and sugar and hormone content of adventitious and storage roots. (A) adventitious roots; (B) storage roots after 20 and 70 d of growth in soil. Scale bar: 1.5 cm. (C) Changes in sugar and hormone content between storage and adventitious roots. The measurements were conducted after 20 and 70 d of growth in soil, respectively. Abbreviations: FW, fresh weight. ABA, abscisic acid; IAA, indoleacetic acid; tZR, tZ riboside; tZ, trans-zeatin; iP, isopentenyladenine; iPR, isopentenyladenosine; BR, brassinosteroid; JA, jasmonic acid; SA, salicylic acid. (D) Changes in soluble amino acid content between storage and adventitious roots. Data are mean ± SE (n = 8). An asterisk (*) indicates significant differences in comparison to the adventitious root at P < 0.05 (Student’s t-test).
Biomass, Sugar, and Hormone Content
Adventitious and storage roots of A. philoxeroides were harvested after 20 and 70 d, respectively, of growth in the university greenhouse, with every 60 to 70 adventitious roots or storage roots being mixed as one biological replicate. Each treatment had three replicates. The roots were separated and washed in double-distilled water, blotted dry, and weighed. Dry weights were determined after oven-drying at 80 C for 72 h until constant weight was observed. Total water content was calculated using fresh weight minus dry weight. Soluble sugar was extracted and measured according to Zhu et al. (Reference Zhu, Xu, Hu, Han, Yin, Li and Gong2015) using anthrone ethyl acetate reagent. Briefly, the samples were extracted with 4 ml of 80% (v/v) ethanol at 80 C for 40 min and were then centrifuged at 2,000 × g for 15 min. The precipitate was re-extracted with 2 ml of 80% (v/v) ethanol and recentrifuged. The supernatants were used for sugar analyses. The absorbance of the supernatant was measured at 630 nm. Each treatment included five replications.
Individual sugars of adventitious and storage roots, including sucrose, fructose, and glucose, were extracted and measured according to Zhu et al. (Reference Zhu, Guo, Feng, Jia, Han and Gong2016) using a sugar analysis system (high-performance liquid chromatography [HPLC], Shimadzu, Kyoto, Japan). Root samples were harvested and extracted with 80% (v/v) aqueous ethanol three to four times. After the supernatant was evaporated, the residue was redissolved in water and filtered through a 0.22-μm membrane HPLC filter (Whatman, Maidstone, UK). Abscisic acid (ABA), indoleacetic acid (IAA), brassinosteroid (BR), gibberellin (GA), jasmonic acid (JA), SA (salicylic acid), and cytokinin (trans-zeatin [tZ] and isopentenyladenine and the corresponding ribosides tZR and iPR) content in adventitious and storage roots was extracted according to Yuan et al. (Reference Yuan, Zhong, Shu, Du, He, Yuan, Sun and Guo2015) and Noguchi et al. (Reference Noguchi, Fujioka, Takatsuto, Sakurai, Yoshida, Li and Chory1999) with some modifications. Samples were homogenized and extracted in 1 ml of methanol:acetic acid:water (12:3:5 [v/v/v]) for 12 h at 4 C. After centrifugation for 10 min at 8,000 × g (4 C), the supernatant was used to determine plant hormone content, and the pellet was extracted at least three times. The extraction solution was concentrated with a vacuum rotary evaporator. The residue was dissolved with 0.5 ml methanol:acetic acid:water (12:3:5 [v/v/v]) and filtered through a 0.22-μm filter prepared for HPLC (Agilent Technologies 1200 series, Santa Clara, CA, USA) analysis. The flow rate was 0.8 ml min−1, and the injection volume was 10 μl. The quantities of plant hormones in the sample were expressed on a fresh weight basis (μg g−1). There were five replicates for sugars, ABA, IAA, BR, GA, JA, SA, and cytokinin analyses.
Soluble amino acids in adventitious and storage roots were detected using HPLC (Shimadzu). A 0.5-g sample was homogenized with 6 M hydrochloric acid solution. The acid solution and 1 ml of homogenized solution were transferred to glass ampoules filled with nitrogen for 5 min, then hydrolyzed at 110 C for 18 h in flame-sealed glass ampoules. The hydrolyzed samples were transferred to 1.5-ml centrifuge tubes and centrifuged at 12,000 × g for 5 min. Finally, 0.5 ml of the supernatant was vacuum-dried and redissolved in 0.5 ml hydrochloric acid solution (0.1 M) for determination of soluble amino acids. For detection of soluble amino acids, 20 μl of leucine, 100 μl of triethylamine phenyl, and 100 μl of isothiocyanate solution were added to 200-μl samples of the soluble amino acid extractions. After incubation at room temperature for 30 min, 400 μl of normal hexane was added, the entire mixture was vortexed vigorously for 15 s, followed by centrifugation at 12,000 × g for 2 min. The lower liquid was taken for HPLC analysis. Twenty microliters of each soluble amino acid extraction was separated on a 4.6 by 250 mm Diamonsil AAA column (Dikma, Beijing, China). The samples were eluted from the column at a flow rate of 1.0 ml min−1 at 35 C. The peaks of soluble amino acids were detected with a UV detector at 254 nm. There were three biological replicates for soluble amino acid analyses.
Identification of Unigenes Associated with the Formation of Storage Roots
Illumina RNA-seq data belonging to four A. philoxeroides transcriptome sequencing BioProjects (serial numbers: PRJNA256235, PRJNA256237, PRJNA263573, and PRJNA268359) were downloaded from the SRA category of the NCBI database. The Illumina reads obtained were used as input to create the de novo assembly. Briefly, the SRA Toolkit subroutine fastq-dump was used to transform SRA format data into fastq format, then trimmed by Trimmomatic to exclude the low-quality bases and reads (Bolger et al. Reference Bolger, Lohse and Usadel2014). The remaining high-quality reads were used to perform de novo assembly using Trinity (v. 2.0.6), with annotation using TransDecoder (v. 5.3.0) (Haas et al. Reference Haas, Papanicolaou, Yassour, Grabherr, Blood, Bowden, Couger, Eccles, Li, Lieber, MacManes, Ott, Orvis, Pochet and Strozzi2013) and expression profiling using RSEM (v. 1.2.19) (Li and Dewey. Reference Li and Dewey2011). Default parameters were used for each software during the de novo assembly process. Seven public databases, including Swiss-Prot (a manually annotated and reviewed protein sequence database), Pfam (Protein family), Nr (NCBI non-redundant protein sequences), KOG/COG (Clusters of Orthologous Groups of Proteins), KEGG (Kyoto Encyclopedia of Genes and Genomes), GO (Gene Ontology), and eggNOG (a database of orthologous groups and functional annotation), were used to perform gene function annotation with the cutoff value set as 1 × 10−10 (Zhu et al. Reference Zhu, Yin, Liang, Liu, Jia, Huo, Wu, Yang and Gong2019a). Based on KEGG annotation, unigenes (the clustered contigs designated during the Trinity assembly) related to storage root-swelling processes, including sucrose metabolism and ABA, IAA, and BR signal transduction, were manually extracted. Expression levels of these genes were then collected, and the R package pheatmap was used to illustrate their expression profiles (Fang et al. Reference Fang, Jiang, He, Ma, Liu, Wang, Zhang and Yin2020).
Quantitative Reverse Transcriptase Polymerase Chain Reaction Analysis
Two sets of samples were collected and used for qRT-PCR analysis. The first set of samples included adventitious and storage roots of A. philoxeroides harvested after 20 and 70 d of growth in the university greenhouse, respectively, as described earlier. The second set of samples (adventitious roots grown in water and storage roots grown in the soil underwater) were collected from a natural pond environment (112.153°E, 30.358°N). The storage roots of the second set of samples, which were grown in soil underwater, were harvested when their diameters were similar to those of the first set of A. philoxeroides samples grown in the greenhouse. Total RNA was isolated from collected tissues using TRIzol™ reagent (Tiangen, Beijing, China), and the integrity and quality of RNA was assessed by performing 15 g L–1 agarose gel electrophoresis. The quantity and quality of RNA samples were measured with a NanoDrop™ 2000 Spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). Single-stranded cDNA for qPCR analysis was synthesized from 1 μg of total RNA using HiScriptTMReverse Transcriptase (Thermo Scientific) according to the manufacturer’s protocol, including a special step for genomic DNA digestion. Real-time qRT-PCR was performed using SYBR Green Master Mix (Vazyme, Nanjing, China) on a CFX 96 Real-Time PCR system (Bio-Rad, Hercules, CA, USA) as previously reported (Yin et al. Reference Yin, Fang, Sun, Zhang, Zhang, Lu, Wang, Ma and Zhu2018a). The relative quantity was calculated using the 2−ΔΔCt method. For each sample, there were three biological replications and two technical replications. The qRT-PCR primers used in this assay were designed using Primer Premier v. 5.0 and are listed in Supplementary Table 1. The housekeeping gene actin reported by Song and Su. (Reference Song and Su2013) was used as the internal control.
Statistical Analysis
Data were subjected to ANOVA using SPSS v. 19.0. The differences were considered significant at P < 0.05.
Results and Discussion
Changes in Sucrose and Hormone Content during Storage Root Formation
Previous studies showed that A. philoxeroides has limited genetic diversity, especially in China (Chen et al. Reference Chen, Xiong, Pan, Shen, Geng, Xu, Chen and Zhang2015; Wang et al. Reference Wang, Li and Wang2005a). Chen et al. (Reference Chen, Xiong, Pan, Shen, Geng, Xu, Chen and Zhang2015) identified a total of four levels of genome size and three kinds of ploidy in A. philoxeroides from Argentina, the United States, and China, but only one genome size and one ploidy level were found in samples from China. Moreover, varying genome sizes (5.72 to 8.50 pg per 2C nucleus) with low genetic differentiation in the internal transcribed spacer sequences among biotypes from different areas suggested that the genome size changes may contribute to the stronger invasive ability of A. philoxeroides (Chen et al. Reference Chen, Xiong, Pan, Shen, Geng, Xu, Chen and Zhang2015). Wang et al. (Reference Wang, Li and Wang2005a) analyzed genetic diversity of seven A. philoxeroides populations in China using multiple random amplified polymorphic DNA and inter-simple sequence repeats markers and reported extremely low genetic diversity of this invasive species in China. These studies imply that low genetic diversity did not affect the success of its expansion in China. Most studies on A. philoxeroides were focused on physiological processes (e.g., leaf/stem morphology, biomass allocation, and phytochemical constituents), whereas the underlying mechanisms leading to adventitious root enlargement remain unclear. In this study, the root traits, root sugars, soluble protein, and hormone content were determined in both adventitious and storage roots.
As can be seen in Figure 1, after 20 d of growth, plants had grown adventitious roots (diameter < 1.5 mm; Figure 1A) and after 70 d of growth, plants had grown storage roots (diameter > 1.5 mm; Figure 1B). Root-trait analysis showed that dry weight, root length, and root surface area of storage roots increased compared with adventitious roots (Table 1). Moreover, root/shoot ratio and total water content increased during the transition process (Table 1).
Table 1. Changes in root traits and total water content between storage roots and adventitious roots.a

a Data are mean ± SE (n = 8). Different letters in a column indicate a significant difference between sampling times (P < 0.05, Student’s t-test).
b Root traits (root dry weight, root dry total length, root total volume, root surface areas) were measured after 20 and 70 d of growth in soil.
c The root/shoot ratio = total weight of root/total weight of shoot.
Sugar is an energy source for growth and one of the signaling molecules in plant cells (Takahashi et al. Reference Takahashi, Sato-Nara, Kobayashi, Suzuki and Suzuki2003). Maintenance of the balance among sugar production, translocation, partition, and use is critical for the development and growth of plants (Zhu et al. Reference Zhu, Gong and Yin2019b). During cold winters, aboveground tissues of A. philoxeroides growing in dry habitats die and storage roots become the propagation source for the next growing season. Storage root formation and development are complex processes that involve dry matter (e.g., carbohydrate, storage proteins) accumulation as well as phytohormone regulation (Ravi et al. Reference Ravi, Chakrabarti, Makeshkumar and Saravanan2014). In our study, soluble sugar and individual sugars, including sucrose, fructose, and glucose, were measured, and total sugar content increased 11-fold in storage roots compared with adventitious roots (Figure 1C). Compared with adventitious roots, sucrose and fructose content was significantly increased in storage roots, whereas glucose content was significantly decreased. Sucrose was the major sugar in roots, representing more than 80% and 50% of total sugars in storage roots and adventitious roots, respectively (Figure 1C). Sucrose has been found to be the main base material for root growth and plays important roles in both the development of adventitious root and adventitious root enlargement (Ravi et al. Reference Ravi, Chakrabarti, Makeshkumar and Saravanan2014). Moreover, sucrose has been suggested to function as a specific signal in meeting the sugar requirement for the development of storage roots and other sink organs (Sun et al. Reference Sun, Xiao, Duan, Guo, Qi, Liao, Zhao, Liu, Zhou and Li2015). In sweetpotato [Ipomoea batatas (L.) Lam.], exogenous injection of sucrose was reported to enhance storage root production (Tsubone et al. Reference Tsubone, Kubota, Saitou and Kadowaki2000). However, it is still unknown how sucrose is involved in the formation of storage roots in A. philoxeroides. Further studies are needed to explore the activities of enzymes involved in carbohydrate metabolism, which is quite complex and tissue- and development-stage dependent.
Compared with adventitious roots, ABA and BR content was significantly decreased by 97% and 39% in the storage roots, respectively. ABA is one of the most important plant hormones and plays a vital role in regulating plant physiological metabolism; it has been suggested that ABA regulates the thickening of storage roots by promoting cellular division at the meristem (Tanaka et al. Reference Tanaka, Takahata and Nakatani2005). Previous studies suggested that the regulatory effect of ABA on the growth of roots varies among species. For example, in rice (Oryza sativa L.), exogenous ABA increases root biomass and modifies root morphology, including density and number of adventitious roots (Liu et al. Reference Liu, Ren, Zhu, Wu and Liang2018). By contrast, in some legume species, ABA was reported to depress the development of lateral roots (Di et al. Reference Di, Jian, Wang, Chen, Ding, Du, Lu, Li and Liu2018; Liang and Harris. Reference Liang and Harris2005). In the present study, ABA content was lower than IAA content but higher than content of other hormones detected in the roots (Figure 1C). ABA content was significantly decreased in storage roots, suggesting the participation of ABA in the regulation of the root-swelling process. BR is an endogenous polyhydroxy steroidal phytohormone that is involved in many plant processes, including root growth and development (Guo et al. Reference Guo, Zhou, Ren, Jia, Hua, Xu, Lv, Zhao and Wei2018). Vardhini et al. (Reference Vardhini, Sujatha and Rao2011) reported that foliar supplementation of BR stimulated the growth of radish (Raphanus sativus L.) roots, which was associated with increased levels of reducing sugars, soluble protein, and starch in the storage roots of radish. In A. philoxeroides, BR content significantly decreased in the storage roots compared with adventitious roots, but the precise roles of BR in the formation and/or development of storage roots are still unknown and need further investigation.
In adventitious roots of A. philoxeroides, our results suggested that IAA content was the highest among all seven kinds of tested hormones, but IAA content decreased 94% in the storage root compared with the adventitious root (Figure 1C). The plant hormone IAA is the most important natural auxin; it is involved in virtually every aspect of developmental processes and has been shown to be a major growth substance involved in the formation of adventitious roots (Ren and Gray Reference Ren and Gray2015). Adventitious roots are largely pentarch or hexarch and contain a central pith with or without central metaxylem cells. In sweetpotato, IAA level has been proposed to be critical for formation of hexarch or pentarch adventitious roots, which is a prerequisite for the initiation of storage root formation (Ravi et al. Reference Ravi, Chakrabarti, Makeshkumar and Saravanan2014). Wang et al. (Reference Wang, Zhang and Wang2005b) reported that dry-storage root yield was positively correlated with the concentration of dihydrozeatin riboside, ABA, and tZR, but not with IAA, isopentenyl-adenine, or gibberellin (GA4). In this study, IAA was determined after 20 (adventitious roots, diameter < 1.5 mm) and 70 d (storage roots, diameter > 1.5 mm) of growth, although it is difficult to characterize the precise growth stages of roots. In the future, the content of IAA as well as other hormones needs to be determined at several different stages during the formation of storage roots.
Bioactive cytokinins include iP, tZ, and the corresponding ribosides iPR and tZR. tZ-type cytokinins (tZR and tZ) are the dominant cytokinins in both adventitious and storage roots. Compared with adventitious root, tZ, tZR, and iP levels were significantly lower, whereas iPR was significantly higher in the storage root (Figure 1C). Physiological studies have suggested cytokinins, together with other hormones like GA and auxin, play crucial roles in the formation of storage roots (Tanaka et al. Reference Tanaka, Kato, Nakayama, Nakatani and Takahata2008). In storage roots of sweetpotato, tZR was proposed to be the major active endogenous cytokinin, the content of which increased rapidly during the development of storage roots (Tanaka et al. Reference Tanaka, Takahata and Nakatani2005). However, in the adventitious roots of A. philoxeroides, tZ was the major cytokinin form and was significantly reduced in the storage root. Moreover, Sojikul et al. (Reference Sojikul, Saithong, Kalapanulak, Pisuttinusart, Limsirichaikul, Tanaka, Utsumi, Sakurai, Seki and Narangajavana2015) suggested that cytokinins play a role in sucrose mobilization into metabolic pathways and storage functions in cassava (Manihot esculenta Crantz). In this study, tZ content displayed opposite trends compared with sucrose content in adventitious roots (20 d) and storage roots (70 d). These results implied the participation of cytokinins in storage root development, and the content of different forms of cytokinin (e.g., iP, tZ, iPR, and tZR) in A. philoxeroides should be investigated at more time points during storage root formation. The roles of cytokinins in sucrose mobilization during storage formation need to be ascertained.
No changes of 17 kinds of soluble amino acid content were detected between storage and adventitious roots (Figure 1D). However, the soluble protein content increased in the storage root compared with the adventitious root. Storage proteins have been identified in tubers of several species, including potato (Solanum tuberosum L.) and yam (Dioscorea alata L.). Synthesis and mobilization of storage proteins were not studied here, and it would be reasonable to speculate that the inconsistency between soluble amino acid content and soluble protein content may be partly associated with storage protein metabolism. Soluble proteins have been reported to accumulate in the perennial weeds dandelion (Taraxacum officinale F.H. Wigg) and chicory (Cichorium intybus L.) and are associated with nitrogen storage or stress responses, including overwintering (Amini and Ehsanpour Reference Amini and Ehsanpour2005; Cyr and Derek Reference Cyr and Derek1990). Thus, it can be assumed that the increased soluble protein content in the storage root of A. philoxeroides may contribute to plants responding to abiotic stresses (e.g., cold and/or freezing stress) and increased soluble protein content may undergo profound changes in plants under land and water conditions.
Our results suggest that plant hormones and sucrose participate in the swelling process of storage roots. Previous studies indicate that ABA regulates root growth and the thickening of storage roots by itself or through interaction with cytokinins, ethylene, and auxin (Ravi et al. Reference Ravi, Chakrabarti, Makeshkumar and Saravanan2014; Rowe et al. Reference Rowe, Topping, Liu and Lindsey2016). Moreover, the internal balance of phytohormones such as ABA and cytokinins may also be crucial to storage root development (Ravi et al. Reference Ravi, Chakrabarti, Makeshkumar and Saravanan2014). In the future, by further revealing the function of these phytohormones and many other hormones in storage root formation regulation, we should be able to elucidate how these hormones regulate root growth and bring new understanding to the control of storage root development in A. philoxeroides.
De Novo Assembly and Functional Annotation of the Alternanthera philoxeroides Transcriptome
Although A. philoxeroides displays significant phenotypic variation when grown in different environments, very little genetic diversity within and among populations was found, especially in China, where it grows as a uniformly hexaploid population, suggesting transcriptome reprogramming plays essential roles in phenotypic adaptation of this weed (Chen et al. Reference Chen, Xiong, Pan, Shen, Geng, Xu, Chen and Zhang2015; Wang et al. Reference Wang, Li and Wang2005a; Xu et al. Reference Xu, Zhang, Fu and Lu2003). Several RNA-seq projects performed in A. philoxeroides studied different tissues under variable treatments and provided a large number of reads for de novo assembly (Gao et al. Reference Gao, Geng, Yang, Hu and Yang2015; Li et al. Reference Li, Xu, Wang, Pan and Lu2015; Liu et al. Reference Liu, Horvath, Li and Liu2019). As different RNA-seq projects were designed to handle specific problems, an individual project may not be enough to explore transcripts comprehensively, especially for A. philoxeroides, a species that lacks a completely sequenced genome. Therefore, an alternative strategy is to combine different RNA-seq data together to perform de novo assembly and obtain a more comprehensive transcript reservoir (Iyer et al. Reference Iyer, Niknafs, Malik, Singhal, Sahu, Hosono, Barrette, Prensner, Evans, Zhao, Poliakov, Cao, Dhanasekaran, Wu and Robinson2015). In this study, 62.5 Gb of data from four projects were collected and used as input to perform the de novo assembly (Table 2).
Table 2. RNA-seq data sets used for Alternanthera philoxeroides de novo assembly.

In total, 421,372 unigenes (the clustered contigs designated during the Trinity assembly) were assembled. Total gene length reached 230,842,460 bp, and average length and N50 length were 548 bp and 621 bp, respectively. Among them, 43,430 (10.31%) unigenes were longer than 1,000 bp (Figure 2A). BUSCO (Benchmarking Universal Single-Copy Orthologs) was used to assess the completeness of unigenes and the ratio of “Complete BUSCOs” reached 77.08%, suggesting that unigenes were relatively intact and suitable for mining candidate genes involved in various metabolic pathways (Simão et al. Reference Simão, Waterhouse, Ioannidis, Kriventseva and Zdobnov2015). Then these unigenes were functionally annotated using seven public databases (Yin et al. Reference Yin, Liu, Ma, Wu, Li, Zhu and Han2018b). In total, 119,222 unigenes were successfully annotated by at least one of the databases and specifically 30,832 (25.9%), 46,394 (38.9%), 62,768 (52.6%), 66,366 (55.7%), 66,525 (55.8%), 66,689 (55.9%), 96,797 (81.2%), and 106,758 (89.5%) unigenes were annotated by the COG, KEGG, GO, KOG, Swiss-Prot, Pfam, eggNOG, and Nr databases, respectively (Figure 2B). A low ratio of unigenes (28%) was functionally annotated, which is consistent with the fact that large numbers of transcripts in the transcriptome belong to non–protein coding RNAs (ncRNAs), which constitute a substantial portion of transcribed sequences in animals and plants; in Arabidopsis, at least 50% of transcripts are ncRNAs, and with the development of the next-generation sequencing technologies and transcriptomic analysis approaches, more and more ncRNAs have been detected in plants and animals (Liu et al. Reference Liu, Jung, Xu, Wang, Deng, Bernad, Arenashuertero and Chua2012; Severing et al. Reference Severing, Faino, Jamge, Busscher, Kuijer-Zhang, Bellinazzo and Pajoro2018; Yuan et al. Reference Yuan, Zhang, Dong, Sun, Lim, Liu and Lu2016).

Figure 2. Annotation and function enrichment of unigenes. (A) Length range of annotated unigenes. (B) Number of all annotated unigenes and number of unigenes with annotation in the specific database. (C) Distribution of the BLAST hits of Alternanthera philoxeroides unigenes in Nr database. The numbers in the labels represent the number and percentage of unigenes in different species (n, %). (D) GO (Gene Ontology) classification of unigenes. Percentage and number of genes are shown in three main categories: cellular component (blue), molecular function (red), and biological process (green). (E) Distribution of genes in the transcriptome by COG (Clusters of Orthologous Groups of Proteins) functional classification. A total of 30,832 unigenes were grouped into 26 COG classifications.
Nr Annotation
As shown in Figure 2C, the organism similarity of A. philoxeroides unigenes to other plant species was analyzed through comparison of the assembled unigenes with nonredundant proteins in the NCBI Nr database. These A. philoxeroides unigenes showed the closest matches with beet (Beta vulgaris L.) (25,582, 24%), followed by quinoa (Chenopodium quinoa Willd.) (21,529, 20%), and spinach (Spinacia oleracea L.) (10,036, 9.4%), all three of which are in the same family as A. philoxeroides, the Amaranthaceae. No other matches from the Nr database were greater than 1% similarity, for example, barley (Hordeum vulgare L.) (919, 0.86%), wine grape (Vitis vinifera L.) (826, 0.77%), cucumber (Cucumis sativus L.) (684, 0.64%), coyote tobacco (Nicotiana attenuata Torr. ex S. Watson) (681, 0.64%), Basidiobolus meristosporus (646, 0.61%), Dorcoceras hygrometricum CBS (632, 0.59%), and Arachis ipaensis (571, 0.53%), suggesting A. philoxeroides unigenes are more homologous to transcripts of Chenopodiaceae plants.
GO Annotation
GO is a widely used function classification system for genes that can help predict the physiological role of unigenes (Xia et al. Reference Xia, Xu, Zhai, Li, Luo, He and Huang2011). Thus, Blast2GO was used to annotate and categorize unigenes into GO functions (Zhou et al. Reference Zhou, Zhu, Zhao, Fang, Wang, Yin, Chu and Ma2018). As shown in Figure 2D, a total of 62,768 unigenes were assigned into 54 GO functional categories. Among these 54 GO categories, biological process, cellular component, and molecular function were most highly representative, accounting for 16 (29.6%), 15 (27.8%), and 23 (42.6%), respectively. The most representativeGO terms in these three subclasses were cell (GO:0005623), catalytic activity (GO:0003824), and metabolic process (GO:0008152) with 29,926, 31,972, and 34,373 genes, respectively.
COG Annotation
The COG database is a tool to classify products of orthologous genes based on the orthology concept. All assembled unigenes of A. philoxeroides were analyzed against the COG database. A total of 30,832 unigenes were annotated to 26 protein families, accounting for 7.32% of the total unigenes (Figure 2E). In the 26 COG categories, the cluster for “Translation, ribosomal structure and biogenesis” (4,805 unigenes, 15.58%) was the largest group, followed by “Posttranslational modification, protein turnover, chaperones” (3,342 unigenes, 10.84%) and “General function prediction only” (3,214 unigenes, 10.42%). And the fewest gene sequences (17) were found in the “RNA processing and modification” cluster, while no sequences were annotated into the “Nuclear structure” category. GO and COG classifications suggest that the assembled unigenes of A. philoxeroides have diverse molecular functions and participate in multiple metabolic pathways.
KEGG Pathway Mapping
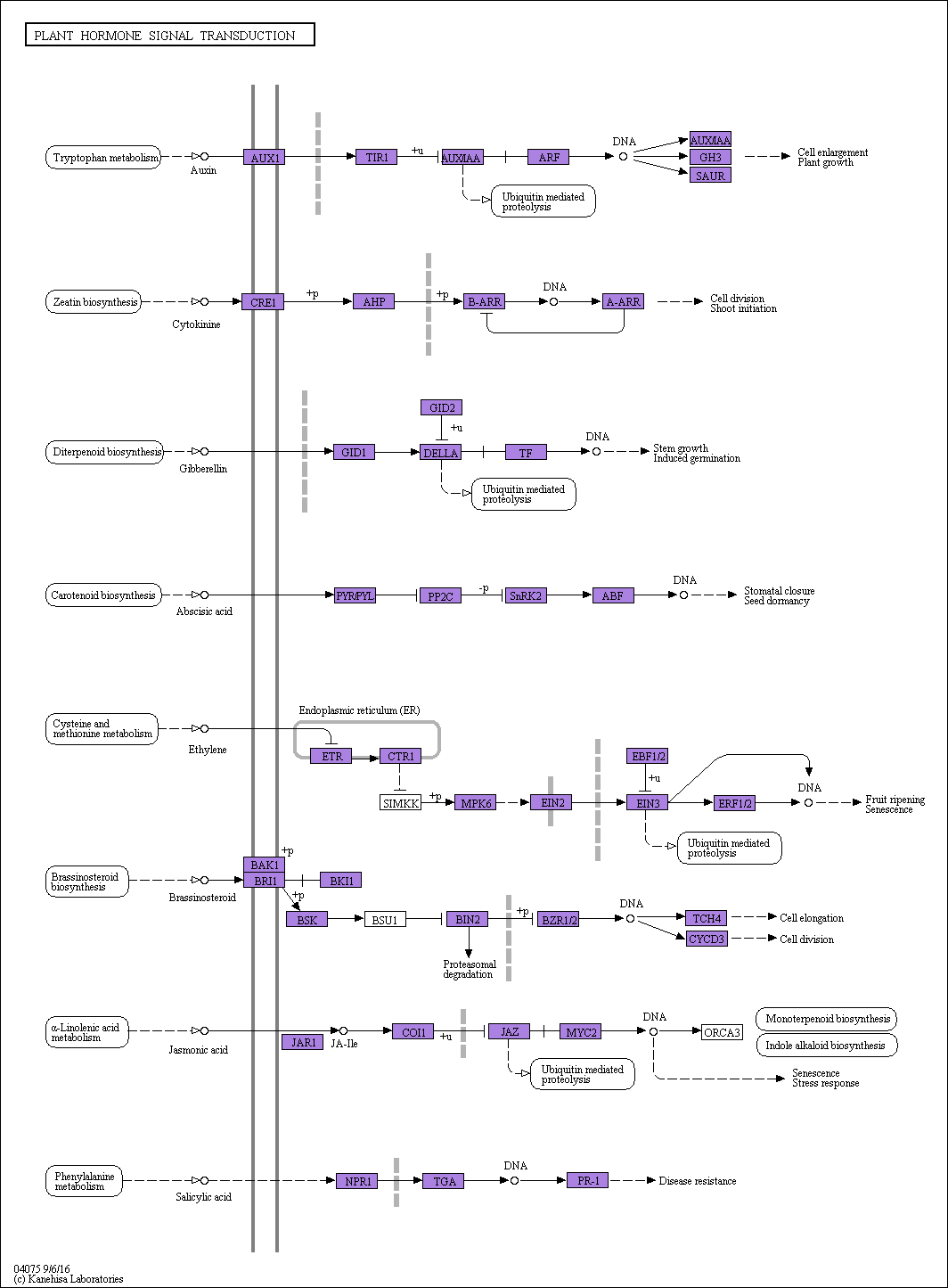
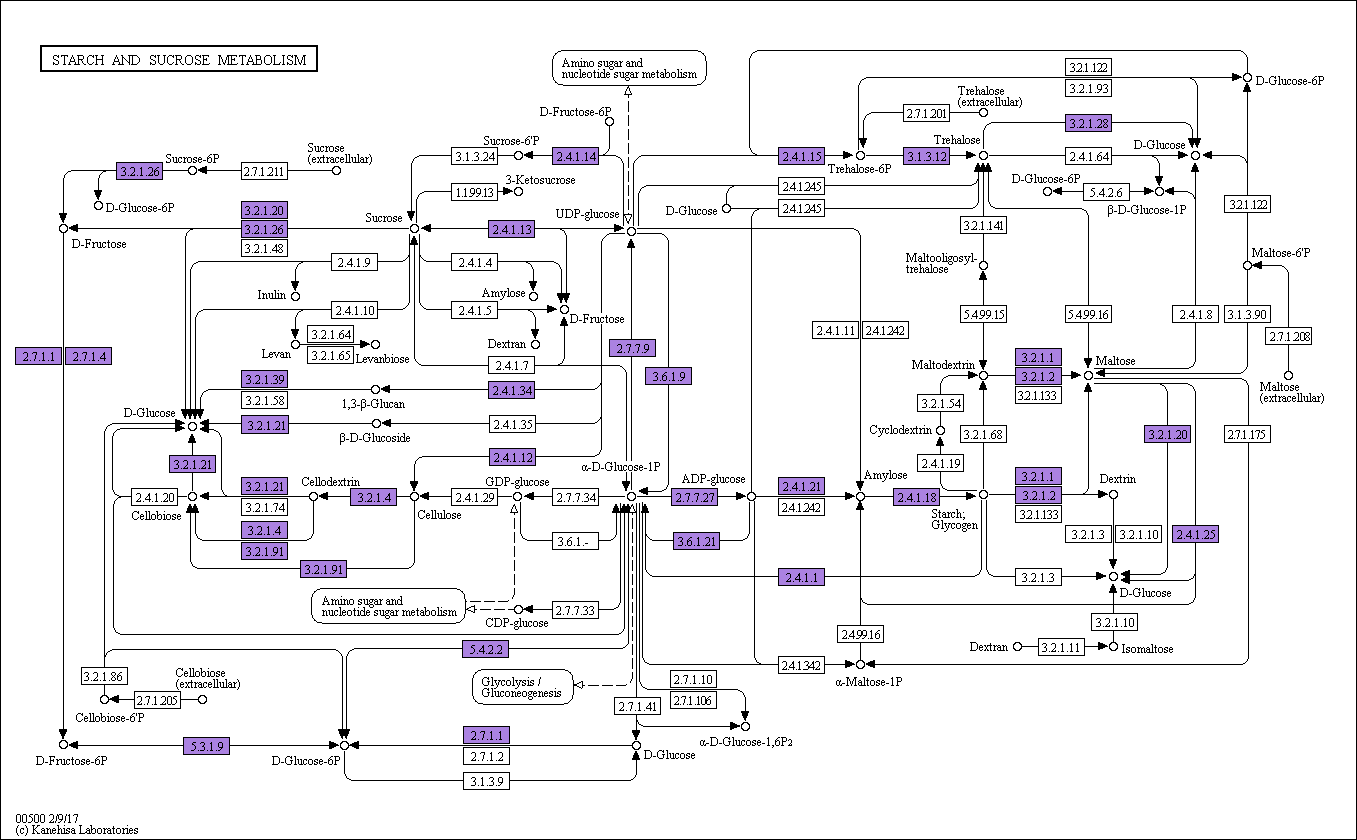
To further analyze the products of unigenes during metabolic processes and determine their biological functions in cellular processes, the assembled sequences were mapped to the reference canonical pathways in the KEGG database using BLAST. In total, 46,394 unigenes were annotated and assigned to 130 different pathways. As a result, “Ribosome” (4,116 unigenes, accounting for 8.87%), “Carbon metabolism” (2,244 unigenes, 4.84%), and “Biosynthesis of amino acids” (1,561 unigenes, 3.36%) were the most highly represented pathways (Supplementary Table 2). Moreover, 834 (1.80%) and 761 (1.64%) unigenes were assigned to the “Starch and sucrose metabolism” (ko00500; Supplementary Figure 1) and “Plant hormone signal transduction” (ko04075; Supplementary Figure 2) pathways, respectively (Supplementary Table 2).
Sucrose and Phytohormone Metabolism
Previous studies revealed that the formation and development of storage roots are complex physiological processes involving morphogenesis and dry matter (e.g., carbohydrate and storage protein) accumulation (Ravi et al. Reference Ravi, Chakrabarti, Makeshkumar and Saravanan2014). Similarly, in A. philoxeroides, the soluble sugar, hormone, and soluble protein content varied significantly between adventitious and storage roots. Plant hormone and sugar biosynthetic pathways are complex processes that involve various genes specifically expressed in different organs, but genetic resources for A. philoxeroides are sparse, which impedes the mining of molecular mechanisms behind A. philoxeroides colonization and invasiveness. To reveal expression patterns of sugar- and plant hormone–related genes, especially ABA, BR, IAA, and sucrose metabolic genes in A. philoxeroides, we selected all unigenes related to “Starch and sucrose metabolism” (ko00500) and “Plant hormone signal transduction” (ko04075) KEGG pathways and analyzed their functional annotations and expression patterns (Supplementary Table 2). To better demonstrate their expression patterns and exclude the barely expressed ones, we define four kinds of expression patterns: not expressed (FPKM value < 0.1, fragments per kilobase of exon model per million mapped reads), low-level (0.1 ≤ FPKM value < 1), medium-level (1 ≤ FPKM value < 10), and high-level (FPKM value ≥ 10) expression, which were used as criteria to describe the expression patterns of related genes identified in this research (Supplementary Table 2).
Sugar Metabolism–related Candidate Genes
To explore genes involved in sugar metabolism in A. philoxeroides, 85 genes involved in sucrose synthesis and hydrolysis were identified, including 16, 17, 28, and 24 for sucrose-phosphate synthase (SPS), sucrose synthase (SUS), α-glucosidase, and β-fructofuranosidase invertase, respectively (Supplementary Table 2). Generally, SPS is the key enzyme for sucrose synthesis, and invertases are primary enzymes responsible for sucrose degradation (Zhu et al. Reference Zhu, Guo, Feng, Jia, Han and Gong2016). Sucrose utilization is initiated by sucrose decomposition, which is catalyzed by two different enzymes: SUS (EC 2.4.1.13) and β-fructofuranosidase (also called β-d-fructofuranoside or invertase; EC 3.2.1.26) (Sturm. Reference Sturm1995). SUS is responsible for both sucrose synthesis and decomposition, with the decomposition function being its primarily role (Zhu et al. Reference Zhu, Guo, Feng, Jia, Han and Gong2016). SUS preserves the energy of the glycosidic bond and catalyzes sucrose decomposition into UDP-glucose and fructose, while sucrose cleavage by β-fructofuranosidase is irreversible and generates glucose and fructose. α-Glucosidases (EC 3.2.1.20) have been purified from several plant species and are divided into three types (type I to III). Type I α-glucosidase hydrolyzes heterogeneous substrates (e.g., aryl glucosides and sucrose) more efficiently than maltose (Frandsen and Svensson Reference Frandsen and Svensson1998). However, the isoforms, subcellular locations, and functions of these enzymes are still unknown in A. philoxeroides. To understand the functions of the different isoenzymes of SUS, α-glucosidase, and β-fructofuranosidase, it is important to study when and where these genes are expressed. Therefore, a heat map was drawn based on expression levels (Figure 3). Among 85 assembled unigenes related to sugar metabolism, 43 were excluded due to no or low expression in most tissues or conditions (FPKM mean value < 1), and the remaining 42 genes are shown in Figure 3. According to Figure 3, most of the SPS genes assembled in A. philoxeroides showed medium expression levels based on expression profiling analysis (Supplementary Table 2). Most SUS genes (6/10) showed higher expression levels (FPKM > 10), and the expression levels of these genes varied between different treatments and sampling times (Figure 3; Supplementary Table 2). For example, in pooled shoot samples, the expression levels of the SUS genes TR110357/c1_g2, TR110357/c1_g3, and TR110357/c1_g1 increased in the Pond treatment compared with the Up treatment (Gao et al. Reference Gao, Geng, Yang, Hu and Yang2015). However, in the stem node sample, expression levels varied during sampling times, with their expression levels being lower in the Pond treatment compared with the Up treatment (). The expression levels of several α-glucosidase genes (including TR32014/c1_g2, TR99442/c0_g1, TR231159/c0_g2, and TR23922/c0_g1) differed between the Up and Pond treatments. In the stem node sample, expression levels were generally higher in the Up treatment than in the Pond treatment. However, in pooled shoot samples, the expression levels of two genes, TR32014/c1_g2, TR99442/c0_g1, were lower in the Pond treatment compared with the Up treatment. The expression levels of several β-fructofuranosidase genes (including TR112461/c3_g1, TR112247/c1_g1; TR115034/c0_g2, and TR115034/c0_g1) were generally decreased in the Up treatment compared with the Pond treatment. These results suggest a possible role of sugar metabolism genes under different conditions, which need to be verified experimentally in future studies.

Figure 3. Sucrose synthesis and hydrolysis pathway and the expression levels of related genes. Each row represents a gene of Alternanthera philoxeroides, and each column represents a treatment. Color scale indicates the expression level of cognate genes in log2(FPKM + 1)-transformed value.
Abscisic Acid–related Candidate Genes Assembled
The ABA signal transduction network is complex and associated with many molecular components (Liang and Harris. Reference Liang and Harris2005). Key members in the ABA signal transduction pathway include the PYR/PYL (PYRABACTIN RESISTANCE1/PYR1-LIKE), PP2Cs (type 2C protein phosphatases), and SnRK2s (sucrose non-fermenting 1-related subfamily 2), which have been identified and studied in several species including Arabidopsis thaliana and tomato (Solanum lycopersicum L.) (González-Guzmán et al. Reference González-Guzmán, Rodríguez, Lorenzo-Orts, Pons, Sarrión-Perdigones, Fernández, Peirats-Llobet, Forment, Moreno-Alvero, Cutler, Albert, Granell and Rodríguez2014; Park et al. Reference Park, Fung, Nishimura, Jensen, Fujii, Zhao, Lumba, Santiago, Rodrigues, Chow, Alfred, Bonetta, Finkelstein, Provart and Desveaux2009). However, the ABA signaling pathway is largely unknown in A. philoxeroides. A comprehensive identification of putative ABA signaling pathway genes was carried out to provide a scientific basis for further functional studies. In total, 177 unigenes related to ABA signal transduction (ko04075) were assembled in this study, many of which showed low expression levels or were not expressed in most conditions in A. philoxeroides (evaluated based on expression profiling analysis; Figure 4; Supplementary Table 2). On the other hand, several genes involved in ABA signal transduction with higher expression levels (FPKM > 10) were up- or downregulated in response to different treatments. For example, the expression levels of genes assigned as PYR/PYL (TR108320/c1_g1, TR52226/c1_g1, and TR300871/c1_g4), PP2C (TR947381/c0_g1), and SnRK2 (TR259377/c0_g1) were generally low in the Up treatment, whereas their expression levels were upregulated in the Pond treatment. These results suggest that the differential expression of these hormone and sugar-related genes in various organs may be one of the reasons for the differential accumulation of sugar and ABA.

Figure 4. Abscisic acid signal pathway and the expression levels of related genes. Unigenes with low expression levels were excluded, and 70 unigenes are shown in the figure. Each row represents a gene of Alternanthera philoxeroides, and each column represents a treatment. Color scale indicates the expression level of cognate genes in log2(FPKM + 1)-transformed value.
Brassinosteroid-related Candidate Genes Assembled
In this study, the BR content in adventitious roots was significantly higher than in storage roots, indicating BR participation in storage root formation (Table 1). In plants, BRs have been found to be perceived by BRI1 (BRASSINOSTEROID INSENSITIVE1) at the cell surface (Clouse Reference Clouse2012). BAK1 (BRI1-ASSOCIATED RECEPTOR KINASE1) interacts with BRI1, and this association is promoted by BR. BSKs (BR-SIGNALING KINASES) are direct substrates of BRI1 and positive regulators of BR signaling (Clouse Reference Clouse2012). Phosphorylated BSK1 then interacts with BSU1 (BRI SUPPRESSOR1) phosphatase, promoting its interaction with the negative regulator BIN2 (BRASSINOSTEROID INSENSITIVE2). Inactivation of BIN2 leads to activation of transcription factors BZR1 (BRASSINAZOLE-RESISTANT1) and BZR2, also known as BES1 (BRI1-EMS SUPPRESSOR1), near the terminal end of BR signaling (Clouse Reference Clouse2012). In this study, according to KEGG annotation, most genes (15) assembled as BSK, followed by BRI1 (10). Only one to two genes assembled as BKI1, BIN2, and TCH4 (Figure 5).

Figure 5. Brassinosteroid signal pathway and the expression levels of related genes. Unigenes with low expression levels were excluded, and 58 unigenes are shown in the figure. Each row represents a gene of Alternanthera philoxeroides, and each column represents a treatment. Color scale indicates the expression level of cognate genes in log2(FPKM + 1)-transformed value.
Most genes annotated as BZR1/2, CYCD3, and BSK showed higher expression levels under different tissues and environments (FPKM > 10). Moreover, in the BR signal transduction pathway, many genes with higher expression levels were up- or downregulated in different treatments. For example, TCH4 had a higher expression level under the Pond condition compared with the Up condition, and its expression level varied during sampling time under the same growth conditions (Figure 5; Supplementary Table 2). Therefore, analysis of these genes involved in plant hormone signal transduction provides deeper understanding of the BR signal transduction pathway.
Indoleacetic Acid–related Candidate Genes Assembled
In recent years, extensive knowledge of IAA perception and response has emerged in auxin biology, particularly in Arabidopsis thaliana. AUX1 represents an auxin-influx gene, which was originally cloned in A. thaliana. Since then, four, five, and four members have been identified in A. thaliana (Arabidopsis Genome Initiative 2000), rice (Yu et al. Reference Yu, Hu, Wang, Wong, Li, Liu, Deng and Dai2002), and barrelclover (Medicago truncatula Gaertn.) (Schnabel and Frugoli Reference Schnabel and Frugoli2004), respectively. In A. philoxeroides, 276 genes were assembled in the IAA signal pathway (Figure 5; Supplementary Table 2). Further genome-wide identification and functional analysis of IAA transporters will help us to better understand auxin transport and distribution in A. philoxeroides. Auxin signaling is mediated by several families of well-characterized genes, including the TRANSPORT INHIBITOR RESPONSE/AUXIN SIGNALING F-BOX (TIR1/AFB), the AUXIN RESPONSE FACTOR (ARF) transcription factors, and AUXIN/INDOLE-3-ACETIC ACID (Aux/IAA) transcriptional repressors. TIR1, as a receptor for auxin, is a member of an F-box protein group that contains five possible AFB proteins (AFB1 to AFB5) crucial for substrate recognition in most phases of the auxin response. In this study, 10 AFB genes were assembled in the IAA signal pathway (ko04075), and most of them showed medium or relatively low expression levels in different tissues and treatments (evaluated based on expression profiling analysis) (Figure 6; Supplementary Table 2). Auxin directly promotes rapid degradation of the Aux/IAA family of transcriptional repressors by SCFTIR1/AFB E3 ubiquitin ligase and thus allows ARF-dependent transcription (Chandler Reference Chandler2016). Aux/IAA, Small Auxin-Up RNAs (SAURs), and Gretchen hagen3 (GH3s) genes are early auxin response genes, among which Aux/IAA genes are the most studied. Aux/IAA proteins can interact with each other and act as transcriptional repressors through interaction with ARFs (Chandler Reference Chandler2016; Hagen and Guilfoyle Reference Hagen and Guilfoyle2002). In this study, the greatest number of Aux/IAA genes were assembled in the IAA signal pathway (ko04075), and most of them showed relatively higher expression levels in different tissues and treatments, especially TR93586/c2_g2, TR78426/c2_g1, TR97949/c1_g1, and TR739366/c5_g1 (Figure 6; Supplementary Table 2). The expression levels of SAURs and GH3s were generally low, except for TR45987/c4_g1. These genes assemblies will expand our understanding concerning auxin signaling in A. philoxeroides and will provide candidate genes that can guide future research.

Figure 6. Auxin (indoleacetic acid) signal pathway and the expression levels of related genes. Unigenes with fairly lower expression levels were excluded, and 78 unigenes are shown in the figure. Each row represents a gene of Alternanthera philoxeroides, and each column represents a treatment. Color scale indicates the expression level of cognate genes in log2(FPKM + 1)-transformed value.
Confirmation of Candidate Genes and Their Expression Analyses by qRT-PCR
Plant enzymes are usually encoded by multiple genes that are functionally redundant and/or divergent from each other. For example, SPS, a key enzyme that regulates sucrose biosynthesis, is encoded by different gene families that display varied expression patterns and tissue- and developmental stage–specific functions in different plants. The tissue- and developmental stage–specific expression patterns of the SPS gene family have been reported in several species such as Arabidopsis thaliana, rice, and wheat (Triticum aestivum L.), and differences in expression patterns seem to have functional significance (Castleden et al. Reference Castleden, Aoki, Gillespie, MacRae, Quick, Buchner, Foyer, Furbank and Lunn2004; Fung et al. Reference Fung, Langenkämper, Gardner and MacRae2003; Okamura et al. Reference Okamura, Aoki, Hirose, Yonekura, Ohto and Ohsugi2011; Sun et al. Reference Sun, Zhang, Larue and Huber2011). In the present study, 10 SPS genes were assembled. The expression levels and patterns of two SPS genes in adventitious and storage roots grown in both cultured and natural conditions were examined. Results show that TR49460|c0_g1 was more abundant in the root of A. philoxeroides, while TR277542|c0_g1 appeared to be expressed at relatively low levels compared with TR49460|c0_g1 (Figure 7; Supplementary Table 2). Moreover, the expression level of TR49460|c0_g1 was significantly higher in the storage root compared with the adventitious root, which correlates with increased sucrose content in the storage root. By contrast, the expression level of TR277542|c0_g1 was significantly lower in the storage root. Similarly, in the phytohormone signaling pathways, the expression patterns of examined isogenes, including PYL8, BZR1/2, BSK, and PP2C, varied during the formation of storage roots. However, the functional significance and products of these isogenes are not yet clear; it is difficult to determine the controlling genes and key forms of enzymes related to sugar and hormone metabolism based on limited gene-expression information. Further functional studies should be undertaken to understand the molecular mechanisms of these genes in regulating the formation of storage roots.

Figure 7. qRT-PCR analysis of the relative expression of sucrose metabolism– and hormone signal transduction–related gene expression in adventitious and storage roots of Alternanthera philoxeroides. Different letters indicate significant differences in comparison to the adventitious root at P < 0.05 (Student’s t-test).
In summary, storage roots play important roles in overwintering and propagation of A. philoxeroides, which largely facilitates the invasion and spread of A. philoxeroides in upland habitats. To explore the changes of metabolites during the formation of storage roots, we identified changes in sugar and phytohormone levels and mined genes associated with those processes. In this study, the sugar and plant hormone content, especially ABA, IAA, cytokinin, and BR, was altered during storage root formation, suggesting their roles in the storage root-swelling process. Through de novo assembly, functional annotation, and expression-pattern profiling, candidate genes associated with sucrose metabolism and phytohormone signal transduction were identified. qRT-PCR analysis revealed that these genes were expressed differentially during the formation of storage roots. However, we still poorly understand the functional significance and products of these isogenes, which largely constrains our ability to uncover the controlling genes and key forms of enzymes associated with sugar and hormone metabolism. These results provide a mechanistic understanding of the physiological and genetic mechanisms responsible for biological processes and phenotypic plasticity in this stoloniferous and amphibious perennial species and present candidate metabolites and a useful gene resource for further studies in elucidating the environmental acclimation mechanism and invasion management of A. philoxeroides. Further studies should be undertaken to understand (1) the expression patterns for each member of these gene families and (2) the possible functions of these genes during storage formation of A. philoxeroides.
Acknowledgments
This work was supported by the Key Laboratory Project of Qinghai Science & Technology Department (2020-ZJ-Y02), the Project of Qinghai Sciences & Technology Department (2016-ZJ-Y01), the Open Project of State Key Laboratory of Plateau Ecology and Agriculture of Qinghai University (2016-ZZ-06), the Open Project of Qinghai Key Laboratory of Vegetable Genetics and Physiology (no. 2018-ZJ-Y18), the National Natural Science Foundation of China (no. 31701911), and the Open Project Program of Engineering Research Center of Ecology and Agricultural Use of Wetland, Ministry of Education (no. KF201909). We would like to thank the Shanghai Chenshan Botanical Garden for sharing its RNA-seq data (PRJNA256235) and Beijing Biomarker Technologies Co., Ltd for assisting in data analysis. We thank Yongli Qiao for beneficial comments on the initial project design and data analysis. No conflicts of interest have been declared.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/wsc.2020.37