


1. Introduction
Geographically widespread, organic-rich, dark-grey to black mudstone and shale intervals have been repeatedly deposited through geological time and are strongly associated with times of high eustatic levels (e.g. Fischer & Arthur, Reference Fischer, Arthur, Cook and Enos1977; Leggett et al. Reference Leggett, McKerrow, Cocks and Rickards1981). Dark-grey to black shelf mudstones are often limited to what are now understood to be transgressive systems tracts during eustatic rise with the rise of low-oxygen, deeper slope water across the shelf and deposition limited to hemipelagic muds (e.g. Hallam & Bradshaw, Reference Hallam and Bradshaw1979; Hallam, Reference Hallam1992; Wignall & Newton, Reference Wignall and Newton2001; Bjerrum, Bendtsen & Legarth, Reference Bjerrum, Bendtsen and Legarth2006). It is possible that development of relatively warm epeiric seas with reduced oxygen solubility led to increased burial and preservation of organic material, a condition that followed from increased insolation and warming of shelf seas with onlap and the development of low-density freshwater caps that limited exchange of oxygen and carbon dioxide following rain events on shallow seas (Landing, Reference Landing2012, Reference Landing, Derby, Fritz, Longacre, Morgan and Sternbach2013 a, b; Landing & Smith, Reference Landing and Smith2012).
It is significant to this report that one interval of early Palaeozoic time was characterized by long-term, widespread, organic-rich mudstone deposition that locally spanned cratonic to continental slope settings (Landing, Reference Landing2012). By one model (Landing, Geyer & Bartowski, Reference Landing, Geyer and Bartowski2002; Landing, 2007, Reference Landing2012), an interval of dark mudstone deposition was initiated at the end of the latest early Cambrian Hawke Bay regression. This dark mudstone resulted from a thickening and intensification of the dysoxic mid-water mass on the upper continental slope of the Laurentia continent (Fig. 1). This Hatch Hill dysoxic interval (Landing, Geyer & Bartowski, Reference Landing, Geyer and Bartowski2002) proved to be remarkably persistant and corresponded to a c. 22 Ma interval of high eustatic levels and global temperatures that persisted into earliest Ordovician time (Landing Reference Landing, Derby, Fritz, Longacre, Morgan and Sternbach2013 a, b). Reduced storminess and limited oceanic mixing were primary causes of the thickened mid-water dysoxic mass, which expanded and intensified to bring low-oxygen slope-derived water across several continents with eustatic rise (e.g. Landing, Benus & Whitney, Reference Landing, Benus and Whitney1992; Landing, Reference Landing2012). This model links eustatic rise to increased global insolation by the consideration that shallow seas covered non-vegetated, dominantly tropical Cambrian continents and that epeiric sea warming led to increased oceanic and global temperatures. The model (Fig. 1) mirrors Jenkyns’ (Reference Jenkyns1980, Reference Jenkyns1985, also Cobianchi & Picotti, Reference Cobianchi and Picotti2001) linkage of shelf black shale deposition to onlap by low-oxygen water that originated in deeper environments during Cretaceous time.
A result of high eustatic levels during early Palaeozoic time was deposition of the widespread Alum Shales which extend in outcrop from northern Norway to the island of Bornholm in the Baltic region (Martinsson, Reference Martinsson and Holland1974; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011; Nielsen & Schovsbo, Reference Nielsen and Schovsbo2007). Similarly, dark-grey to black mudstones with intervals of sandstone siltstone span the English Midlands and extend along eastern North America from eastern Massachusetts to eastern Newfoundland (Rushton, Reference Rushton and Holland1974; Allen, Jackson & Rushton, Reference Allen, Jackson and Rushton1981; Landing, Reference Landing, Nance and Thompson1996; Rushton et al. Reference Rushton, Brück, Molyneux, Williams and Woodcock2011). This black mudstone deposition is therefore prominent across the higher temperate latitude, separate, Baltic and Avalonia continents (see Landing et al. Reference Landing, Geyer, Brasier and Bowring2013, fig. 2, and Landing, Westrop & Bowring, Reference Landing, Westrop and Bowring2013, fig. 1, for global palaeogeographic reconstructions). In both regions, organic-rich mud deposition began during middle middle Cambrian time (i.e. traditional middle Acadian Epoch of Avalonia) and persisted until Tremadocian time.

Figure 1. Global hyperwarming model for onlap-offlap facies patterns in tropical and higher latitude continents; modified from Landing (Reference Landing2012, fig. 5).
A traditional interpretation is that cratonic black shale deposition reflects distal, quiescent deep-marine settings with stratified water columns that are likely accompanied by upwelling and nutrient-rich water masses, poorly aerated to anoxic shelf settings and restricted shelf basins (e.g. Hallam & Bradshaw, Reference Hallam and Bradshaw1979; Potter, Maynard & Pryor, Reference Potter, Maynard and Pryor1980; Hay & Cisne, Reference Hay, Cisne and Keith1988; Ettensohn & Brett, Reference Ettensohn and Brett2002; Farrell, Briggs & Gaines, Reference Farrell, Briggs and Gaines2011). This deep-water or stagnant basin model (Seilacher, Reference Seilacher1982), with proposed depths in excess of 100 m and with molybdenum/uranium signatures interpreted to indicate anoxia, has been applied to the Alum Shale succession in Baltica (Thickpenny, Reference Thickpenny, Stow and Piper1984; Thickpenny & Leggett, Reference Thickpenny, Leggett, Brooks and Fleet1987; Pukkonnen & Rammo, Reference Pukkonnen and Rammo1992; Gill et al. Reference Gill, Lyons, Young, Kump, Knoll and Saltzman2011).
However, a number of black-shale-dominated units, ranging in age from Devonian to Cretaceous, record surprisingly dynamic depositional settings. This black shale deposition includes: mud transport by a number of mechanisms (currents, waves, mass movements); varying water mass oxygenation with considerable variation in abundance and diversity of burrowers and skeletalized metazoans; and the presence of event beds and erosion surfaces (e.g. Leckie et al. Reference Leckie, Singh, Goodarzi and Wall1990; Sageman, Wignall & Kauffman, Reference Sageman, Wignall, Kauffman, Einsele, Ricken and Seilacher1991; Schieber, Reference Schieber1994; Wignall & Newton, Reference Wignall and Newton2001). Reliance on these physical indicators of depositional setting, rather than geochemical proxies such as molybdenum and uranium ratios, indicates that most Alum Shale successions record quite shallow-water deposition (W. Newby, unpub. M.Sc. thesis, Colorado State University, 2012; Hints et al. Reference Hints, Hade, Suesso and Voolman2014).
The long-term dark-grey to black mudstone deposition beginning during the middle middle Cambrian Period (middle Acadian Epoch) of Avalonia hosts two lithologically distinct stratigraphic intervals that are consistent with the conclusion that eastern (British) and western (Atlantic coastal North America) Avalonia were part of a unified continent (Landing, Reference Landing, Nance and Thompson1996). The lower is a shallow-water sandstone-rich interval in the middle middle Cambrian deposits in Wales (Ffestiniog Flags; e.g. Crimes, Reference Crimes1970; Allen, Jackson & Rushton, Reference Allen, Jackson and Rushton1981) and England (Mancetter Grits and Shales; e.g. Taylor & Rushton, Reference Taylor and Rushton1972). Coeval units in North American Avalonia include the Cavendish Formation in eastern Newfoundland (Landing & Westrop, Reference Landing, Westrop, Landing and Westrop1998) and synonymous ‘Agnostus Cove Formation’ in southern New Brunswick (Hayes & Howell, Reference Hayes and Howell1937; Landing, Reference Landing, Nance and Thompson1996). Higher up, a greenish mudstone and thin sandstone interval is present in the upper lower Tremadocian deposits. This interval features shallow-marine faunas and wave-dominated structures occur in England (Shineton Shales) and a coeval unnamed interval in Cape Breton Island (Landing, Reference Landing, Nance and Thompson1996; Landing & Fortey, Reference Landing and Fortey2011).
By comparison, the dominant dark-grey to black mudstones of the middle Acadian Epoch – Tremadocian of Avalonia have received little sedimentological study and synthesis. One exception has been the Maentwrog Formation in North Wales. Allen, Jackson & Rushton (Reference Allen, Jackson and Rushton1981) reported apparent turbidite beds and probable contourites in the Maentwrog which they interpreted as ‘deep-sea’ facies, which oddly appeared under the wave-dominated Ffestiniog Formation. Subsequently, hummocky cross-stratification and wave ripples were reported and the Maentwrog Formation is now interpreted as a relatively shallow-water lithesome (Bose, Reference Bose1983; Young et al. Reference Young, Martin, Dean and Rushton1994).
In this report, an upper Cambrian succession dominated by the black mudstones traditionally interpreted as a ‘stagnant’ or ‘anoxic’ basin facies is described because it comprises a presently unique sequence on the Avalonia continent. The short succession probably corresponds to Matthew's (Reference Matthew1903) ‘MacAdam Shore’ section and lies just at the waterline on the north shore of East Bay in Bras d'Or Lake in Cape Breton Island (Fig. 2, section MaS). The section is notable among all of the Avalonian black siliciclastic mudstone sequences that we have investigated, in that it features a number of closely spaced, fossiliferous limestones. The brachiopod and trilobite assemblages from these limestones demonstrate not only condensation of the section, but also record high-energy depositional conditions. In addition, a limestone conglomerate in the middle of the sequence records an important forced regression that led to subaerial conditions and the formation of a calcrete in middle Furongian sediments.
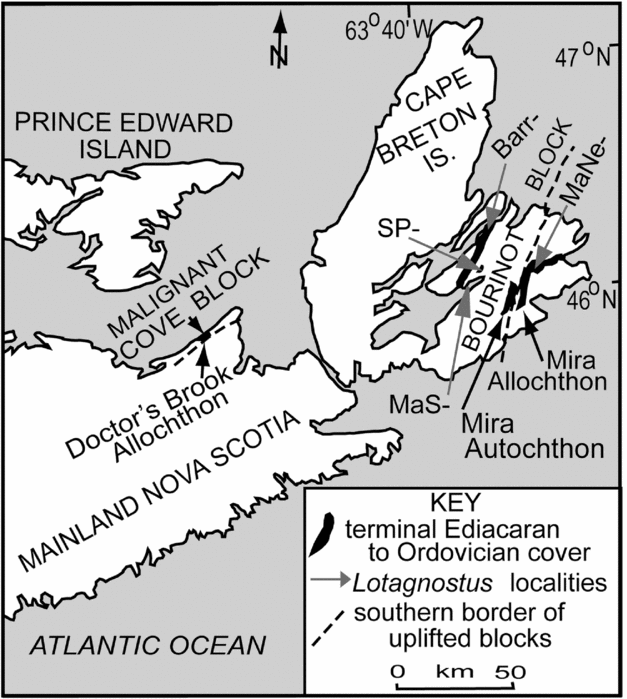
Figure 2. Generalized geologic map of Cape Breton Island area with outcrop belts of terminal Ediacaran–Ordovician cover sequences (black). Lotagnostus–bearing assemblages (grey arrows) from black mudstone-dominated sections at localities noted by Hutchinson (Reference Hutchinson1952) illustrate regional cover of Chesley Drive Group: Barr-,10 m section in Leiches Creek with large, sparsely fossiliferous calcareous nodules, section 0.5 km south of Rte 223 (Boisdale Road) – Leiches Creek intersection (Hutchinson, Reference Hutchinson1952, GSC locality 18596); MaNe-, short, black mudstone-dominated sections up- and downstream from MacKeigen Road Bridge, sections roughly on axis of McLean Brook syncline (Hutchinson, Reference Hutchinson1952, GSC localities 18600–18602; Westrop, Adrain & Landing, Reference Westrop, Adrain and Landing2011); MaS-, isolated section on north shore of East Bay, 0.5 km west of mouth of MacDonald Brook; Sp-, 2.5 m black mudstone section with fossiliferous nodules, headwaters of Spruce Brook and above Devonian granite, c. 900 m north of Eskasoni Road (Hutchinson, Reference Hutchinson1952, GCS locality 18597).
2. Geological setting
2.a. Cape Breton Island and Avalonia
Southern and southeastern Cape Breton Island, the easternmost part of Nova Scotia, lies in the Avalon Zone, a terrane in the NE Appalachians, SE Ireland, southern Britain and parts of western Europe that formed with collision of the late Ediacaran – Ordovician Avalonian continent with Laurentia and Baltica. This terrane during the Caledonian–Acadian orogen was later fragmented by strike-slip faulting during the Carboniferous Alleghenian–Hercynian orogeny by the West Gondwana – Laurussia collision (Williams & Hatcher, Reference Williams and Hatcher1982; Landing, Reference Landing, Nance and Thompson1996, Reference Landing2004, Reference Landing2005; Geyer et al. Reference Geyer, Elicki, Fatka, Żylińska and McCann2008; Linnemann et al. Reference Linnemann, Romer, Pin, Alexandrovski, Buła, Geisler, Kachlik, Kremińska, Mazur, Motuza, Murphy, Nance, Pisarevsky, Schultz, Ulrich, Wiszniewska, Żaba, Zeh and McCann2008). Avalonian Cape Breton Island has a more highly metamorphosed subduction/arc ‘basement’ unconformably overlain by a chlorite-grade, terminal Ediacaran–Ordovician cover sequence (Fig. 1). Rather than recording a ‘Pan-African orogeny’, the replacement of the subduction/arc sequence by terminal Proterozoic rift-volcanic and volcaniclastic rocks took place as the result of a ridge–trench collision (Keppie et al. Reference Keppie, Nance, Murphy and Dostal2003). The Avalonian cover sequence includes the rift succession and overlying, commonly fossiliferous marine sandstone- and mudstone-dominated units deposited in transtensional basins (see Avalonian stratigraphic sections in Landing, Reference Landing, Nance and Thompson1996; Landing, Westrop & Bowring, Reference Landing, Westrop and Bowring2013; Landing et al. Reference Landing, Geyer, Brasier and Bowring2013).
The cover sequence stratigraphy allows Cape Breton Island to be related to Avalonian palaeogeography. The MacAdam Shore (MaS) section lies on the NE margin of Avalonia in the Bourinot belt, which formed an uplifted block on the Avalonian marginal platform during earliest Cambrian time (Landing, Reference Landing, Nance and Thompson1996; Fig. 1). The basement of the Bourinot belt includes middle Proterozoic marble and schist (George River Group; Hutchinson, Reference Hutchinson1952) and Ediacaran leucogranites (553 +/− 25 Ma; Keppie & Smith, Reference Keppie and Smith1978) and is non-conformably overlain by middle Cambrian, near-shore sandstones (Hutchinson, Reference Hutchinson1952). Upper Cambrian black mudstones of the same age as the MaS succession extend from the Bourinot block further southeast into the Mira allochthon (Fig. 2), which is also referable to the Avalonian marginal platform because it has a thick Ediacaran(?) – lowest Cambrian succession with lower rift facies, a middle wave-dominated siliciclastic shelf unit and an upper tidalite sandstone (Landing, Reference Landing1991, Reference Landing, Nance and Thompson1996, Reference Landing2004). The age of the Bourinot belt basement likely reflects deep erosion of uplifted blocks prior to Cambrian deposition (e.g. Dallmeyer et al. Reference Dallmeyer, Doig, Nance and Murphy1990), and does not provide evidence for distinct Cambrian terranes in the Bourinot and Mira areas as concluded by Barr & Raeside (Reference Barr and Raeside1989) and Barr & White (Reference Barr, White, Nance and Thompson1996).
2.b. Chesley drive group
The siliciclastic mudstone-dominated Chesley Drive Group (Landing, Reference Landing, Nance and Thompson1996) provides evidence that Cape Breton Island was part of a unified Avalonian terrane during early Palaeozoic time. It is the youngest lower Palaeozoic cover sequence unit on the Avalonian basement in Cape Breton Island and forms a lithologically uniform blanket that extends from the Mira allochthon to the Bourinot area (Fig. 2). The geographic extent of these black mudstones provides further confirmation that these two regions were simply adjacent parts of Avalonia (Landing, Reference Landing, Nance and Thompson1996, Reference Landing2004). The distribution of the Chesley Drive Group in all of the terminal Ediacaran–Ordovician outliers of Cape Breton Island is shown by an illustration of the Chesley Drive localities from which Hutchinson Reference Hutchinson(1952; see also Westrop, Adrain & Landing, Reference Westrop, Adrain and Landing2011; this report) reported the agnostoid arthropod Lotagnostus Whitehouse, Reference Whitehouse1936.
The Chesley Drive Group of Cape Breton Island was divided by Hutchinson (Reference Hutchinson1952) into two formations, an upper Cambrian ‘MacNeil Formation’ and a Lower Ordovician ‘MacLeod Brook Formation’, with the latter supposedly characterized by the absence of calcareous nodules (Hutchinson, Reference Hutchinson1952, p. 29; Barr, White & MacDonald, Reference Barr, White and MacDonald1992, Barr et al. Reference Barr, White, MacDonald and Raeside1997). However, calcareous nodules occur throughout the upper Cambrian and lower Tremadocian dark-grey to black mudstones succession (i.e. Hutchinson, Reference Hutchinson1952, p. 29, who contradicted himself by noting calcareous nodules in both the upper Cambrian and lower Tremadocian black mudstones). Calcareous nodules in the Tremadocian mudstones are also recorded by Landing et al. (Reference Landing, Bowring, Fortey and Davidek1997), Landing (Reference Landing1997) and Landing & Fortey (Reference Landing and Fortey2011). Indeed, lithologically identical upper Cambrian – Tremadocian black mudstones with or without nodules also occur in eastern Newfoundland (Jenness, Reference Jenness1963) and southern New Brunswick (Landing, Taylor & Erdtmann, Reference Landing, Taylor and Erdtmann1978; Landing, Reference Landing1980) and Rhode Island (Landing, Reference Landing, Nance and Thompson1996). No lithological basis therefore exists anywhere in Avalonia for separating upper Cambrian from Tremadocian mudstones. Consequently, the ‘Chesley Drive Group’ was proposed for this interval throughout Avalonian North America and all existing local, formation-level subdivisions were abandoned (Landing, Reference Landing, Nance and Thompson1996). A completely comparable and coeval lithologic mudstone-dominated interval in British Avalon is the Maentwrog Group of Wales (e.g. Allen, Jackson & Rushton, Reference Allen, Jackson and Rushton1981).
Although exposed in heavily overgrown, tectonized short sections and spot localities, the Chesley Drive Group in Cape Breton Island yields rich Furongian–Tremadocian faunas dominated by trilobites (e.g. Hutchinson, Reference Hutchinson1952; Landing & Fortey, Reference Landing and Fortey2011; Landing et al. Reference Landing, Reyes Patrucco, Andreas and Bowser2012). The fossils are primarily from dark-grey, thin and nodular to bedded limestones. The dominant dark-grey to black siliciclastic mudstones commonly lack fossils, perhaps due in part to early dissolution of calcareous remains, although the typical absence of burrows and benthic, phosphatic or organic-walled macrofossils suggests hypoxic to anoxic bottom-water environments (e.g. Sageman, Wignall & Kauffman, Reference Sageman, Wignall, Kauffman, Einsele, Ricken and Seilacher1991; Schovsbo, Reference Schovsbo2001). Landing & Fortey (Reference Landing and Fortey2011) noted that olenid trilobites, conodonts and chitinozoans occur locally in the dark mudstones and calcareous nodules in the Chesley Drive Group and seem to reflect transient higher oxygen environments, with intervening anoxic intervals possibly responsible for olenid trilobite ‘kill horizons’. A wave-rippled, green-grey mudstone succession within the Chesley Drive Group has lingulate brachiopods, asaphid and shumardid trilobites, and the foraminiferan Bathysiphon A. Sars in G. O. Sars (Reference Sars1872) and records an interval of shallow, dysoxic environments identical to, and coeval with, the Tremadocian Shineton Shales of England.
According to Hutchinson (Reference Hutchinson1952, p. 49), faunas collected in previous studies from the MacAdam Shore include species of the olenids Peltura Milne Edwards, Reference Milne Edwards1840 and Sphaerophthalmus Angelin, Reference Angelin1854 and the agnostoid arthropod Lotagnostus Whitehouse, Reference Whitehouse1936, and have been assigned to the Peltura Superzone. Our new samples demonstrate the presence of older faunas of the Leptoplastus and, probably, Parabolina superzones (see Nielsen et al. Reference Nielsen, Weidner, Terfelt and Høyberget2014, fig. 1, for zonal nomenclature). The occurrence of Lotagnostus is significant because this genus is under consideration as an index for the base of the youngest ‘Stage 10’ Cambrian deposits (Peng et al. Reference Peng, Babcock, Zhu, Zuo and Dai2013); our collections amplify recent objections (Westrop, Adrain & Landing, Reference Westrop, Adrain and Landing2011) to the use of species of Lotagnostus as global biostratigraphic indices.
3. Localities
The MacAdam Shore (MaS) section of this report likely corresponds to Matthew's (Reference Matthew1903) ‘MacAdam Shore’ locality, an outcrop for which he did not provide precise locality information except to note that it was on the north shore of the East Bay of Bras d'Or Lake. We found only one isolated outcrop of Chesley Drive Group in this area (Fig. 2, locality MaS), and it has a fauna comparable to that reported by Matthew (Reference Matthew1901, Reference Matthew1903). This black mudstone-dominated MaS section is only several metres wide, crops out just above lake level, and dips 60°S. The Cambrian rocks are overlain at an angular unconformity by cliff-forming, yellow-weathering sandstone of the lower Carboniferous (middle–upper Visean) Windsor Group (Boehner, Reference Boehner1986; van de Poll, Gibling & Hyde, Reference van de Poll, Gibling, Hyde and Williams1995) (Fig. 3). Section MaS includes highly fossiliferous wacke- to rudstone beds that are unlike the isolated, sparsely or non-fossiliferous calcareous nodules, nodular limestones or sideritic nodules in highly pyritic, barren, black mudstone typical of other North American Avalonian sections of the Chesley Drive Group (Landing, Taylor & Erdtmann, Reference Landing, Taylor and Erdtmann1978; Landing et al. Reference Landing, Bowring, Fortey and Davidek1997; Landing, Reference Landing1980, Reference Landing, Nance and Thompson1996; Landing & Fortey, Reference Landing and Fortey2011).

Figure 3. Lithology and fossils of section MaS. Low outcrop of Chesley Drive Group, section dips 60°S at the shore of Bras d'Or Lake and is exposed by erosion of cliff of overlying, horizontally bedded Windsor Group sandstone.
The nodular to bedded limestone beds at the MaS section were thoroughly sampled (c. 10 kg samples) in the laboratory for crack-out fossils (rhynchonelliform brachiopods, trilobites and agnostoids). In addition, 6.0 kg samples were disaggregated in formic acid for conodontiform remains, pyritized sclerites and originally or secondarily phosphatized remains. However, no such remains were recovered.
4. Synopsis of trilobite and agnostoid faunas
4.a. Olenid faunas
Trilobites and agnostoid arthropods are conspicuous components of several of the shell beds (Fig. 3) and are far better preserved than in Avalonian Britain, where sclerites are typically compacted in shale (e.g. Lake, Reference Lake1913). The MaS collections will be taxonomically documented in detail elsewhere, but a brief summary is presented below.
The oldest olenid fauna, at MaS-3.9, is still under study but includes sclerites assigned to Leptoplastus cf. ovatus Angelin, Reference Angelin1854 (Fig. 4f–j) on the basis of the broad, well-rounded glabella, short genal spine and pygidium with three pairs of marginal spines. However, L. abnormis Westergård, Reference Westergård1944 shares these features and is differentiated largely on thoracic morphology. For this reason, we use open nomenclature. According to Terfelt, Ahlberg & Eriksson (Reference Terfelt, Ahlberg and Eriksson2011), both L. ovatus and L. abnormis occur in the L. ovatus Zone of the upper half of the Leptoplastus Superzone of Nielsen et al. (Reference Nielsen, Weidner, Terfelt and Høyberget2014), which is middle Jiangshanian in age. This collection is of particular importance because it provides an age bracket on the matrix of the calcrete clast-bearing conglomerate from which the collection was made (Fig. 3, see below).

Figure 4. Olenid trilobites from the Chelsey Drive Group, Cape Breton Island, Nova Scotia. Scale bars are 2 mm in length. (a–e) Sphaerophthalmus c.f. arcus Høyberget & Bruton, Reference Høyberget and Bruton2012, from collection MaS-5.8. (a) Free cheek, NBM 17877, dorsal view, ×15. (b) Cranidium, NBM 17878, dorsal view, ×12. (c–e) Cranidium, NBM 17879, dorsal, anterior and lateral view, ×16. (f–j) Leptoplastus cf. ovatus Angelin, Reference Angelin1854, from collection MaS-3.9. (f, g) Pygidium, NBM 17880, dorsal and posterior views, ×15. (h) Free cheek, NBM 17881, dorsal view, ×16. (i, j) cranidium, NBM 17882, dorsal and anterior views, ×16.
Some 2 m above the calcrete clast conglomerate, collection MaS-5.8 is dominated by a species that until recently would be have been assigned to Eoctenopyge Henningsmoen, Reference Henningsmoen1957 but which now belongs to Sphaerophthalmus Angelin, Reference Angelin1854 as revised by Høyberget & Bruton (Reference Høyberget and Bruton2012). In having a well-developed anterior arch, Sphaerophthalmus cf. arcus Høyberget & Bruton, Reference Høyberget and Bruton2012 (Fig. 4a–e) resembles the type cranidia (Høyberget & Bruton, Reference Høyberget and Bruton2012, fig. 7a–e) of this species from the Alum Shale of Norway. However, it differs in having a distinctly emarginate margin to the anterior border (preserved completely in Fig. 4c–e), more firmly incised SO (sulcuc O, or the occipital furrow) that maintains an even depth, more robust genal spine (compare Fig. 4a and Høyberget & Bruton, Reference Høyberget and Bruton2012, fig. 7G) and a seemingly larger palpebral lobe that extends forward to a point just beyond S2. Sphaerophthalmus flagellifer Angelin, Reference Angelin1854 (Høyberget & Bruton, Reference Høyberget and Bruton2012, fig. 9) also has a conspicuous arch, although less so than S. cf. arcus, and differs most clearly in the shorter, advanced genal spine (compare Fig. 4a with Høyberget & Bruton, Reference Høyberget and Bruton2012, fig. 9E, F, and Henningsmoen, Reference Henningsmoen1957, pl. 18, figs 2, 3). Although S. cf. arcus may ultimately prove to represent a new species, closely related species occur in the upper Protopeltura Superzone and MaS-5.8 could be as young as the Ctenopyge similis Zone, which yields S. arcus in the Oslo region of Norway (Høyberget & Bruton, Reference Høyberget and Bruton2012, fig. 2).
The youngest assemblage, at MaS-6.15, is dominated overwhelmingly by olenid trilobites but also includes rare agnostoid arthropods. The majority of the sclerites belong to Sphaerophthalmus cf. alatus Boeck, Reference Boeck and Keihau1838 (Fig. 5i–k), which has been revised by Høyberget & Bruton (Reference Høyberget and Bruton2012, p. 438) who considered a palpebral lobe opposite the S1 glabellar furrow to be diagnostic of S. alatus. However, cranidia from Cape Breton Island consistently have the palpebral lobes further forward and opposite the L2 lobe. Ctenopyge (s.l.) cf. tumida Westergård, Reference Westergård1922 (Fig. 5l, m), is represented by more than a dozen cranidia, all of which differ from the cranidium figured by Terfelt & Ahlberg (Reference Terfelt and Ahlberg2010, fig. 3h) from Scania, southern Sweden, in having more anteriorly positioned palpebral lobes and consequently longer (exsag.) posterolateral projections. A single, incomplete cranidium (not figured) records the presence of Parabolinites Henningsmoen, Reference Henningsmoen1957, but cannot be identified to the species level. These taxa, along with the agnostoids discussed below, indicate that MaS-6.15 belongs in the Peltura Superzone as revised by Nielsen et al. (Reference Nielsen, Weidner, Terfelt and Høyberget2014), and most likely the lowest Ctenopyge tumida Zone (see also Terfelt & Ahlberg, Reference Terfelt and Ahlberg2010). As such, MaS-5.8 and MaS-6.15 may represent successive zones from the Baltic succession.

Figure 5. Olenid trilobites and agnostoid arthropods from the Chelsey Drive Group, Cape Breton Island, Nova Scotia. Scale bars are 2 mm in length. (a–e) Lotagnostus aff. trisectus (Salter, Reference Salter1864), collection MaS-6.15. (a–c) Cephalon, NBM 17883, dorsal, lateral and anterior views, ×15. (d, e) Pygidium, NBM 17884, posterior and dorsal views, ×13. (f–h) Pseudagnostus cf. rugosus Ergaliev, Reference Ergaliev1980, collection MaS-6.15; pygidium, NBM 17885, dorsal, lateral and posterior views, ×13. (i–k) Sphaeophthalmus cf. alatus (Boeck, Reference Boeck and Keihau1838), collection MaS-6.15; cranidium, NBM 17886, lateral, dorsal, and anterior views, ×15. (l, m) Ctenopyge (s.l.) cf. tumida (Westergård, Reference Westergård1922), collection MaS-6.15. (l, m) Cranidium, NBM 17887, dorsal and anterior views, ×14.
4.b. Agnostoid faunas
Agnostoids in collection MaS-6.15 include a single pygidium (Fig. 5f–h) that we assign to Pseudagnostus cf. rugosus Ergaliev, Reference Ergaliev1980. It resembles those from Sweden identified as Pseudagnostus rugosus by Terfelt & Ahlberg (Reference Terfelt and Ahlberg2010, fig. 3f, 4) in possessing scrobiculate genae, but has a relatively broader posteroaxis. The only pygidium figured from the type area in Kazakhstan is poorly illustrated (Ergaliev, Reference Ergaliev1980, pl. 17, fig. 4; Ergaliev & Ergaliev, Reference Ergaliev and Ergaliev2008, pl. 49, fig. 12) but appears to have a narrower posteroaxis than our specimen, as does a pygidium from northwest Hunan, China (Peng, Reference Peng1992, fig. 12G).
Nearly 40 sclerites of Lotagnostus from MaS-6.15 record a strongly furrowed species (Fig. 5a–e) allied with L. trisectus (Salter, Reference Salter1864) and assigned here to L. aff. trisectus. They are far better preserved than specimens from Avalonian Britain (e.g. Rushton, Reference Rushton2009, figs 1A-1, P, 2A–D, G, K–L) which are invariably flattened and often tectonically deformed, so that detailed comparisons are difficult. Cephala from MaS-6.15 have convex glabellae that stand well above the genae with a conspicuous break in slope (Fig. 5b, c). All pygidia have strong trisection of the posteroaxis with well-defined notular furrows (Fig. 5d, e). Lotagnostus cf. trisectus of Westrop, Adrain & Landing (Reference Westrop, Adrain and Landing2011) from higher levels in the Chelsey Drive Group is differentiated by, among other features, non-scrobiculate pygidial acrolobes (Westrop, Adrain & Landing, Reference Westrop, Adrain and Landing2011, figs. 2, 3A–C) and ill-defined glabellar F2 and M3 (Westrop, Adrain & Landing, Reference Westrop, Adrain and Landing2011, fig. 3D–G), and likely represents a new species.
Lotagnostus aff. trisectus is also similar to a species from Baltica traditionally identified as Lotagnostus trisectus (e.g. Westergård, Reference Westergård1922; Rushton, Reference Rushton2009) but assigned to L. americanus (Billings, Reference Billings1860) in some recent studies (Peng & Babcock, Reference Peng and Babcock2005; Ahlberg & Terfelt, Reference Ahlberg and Terfelt2012). As noted by Westrop, Adrain & Landing (Reference Westrop, Adrain and Landing2011), there is very little published information on Lotagnostus from Baltica with just a single cephalon and pygidium repeatedly illustrated in several publications (see Rushton, Reference Rushton2009, fig. 2E–F, H–J for the most recent photographs of these specimens). However, Ahlberg & Terfelt (Reference Ahlberg and Terfelt2012, fig. 4a–f) figured an additional cephalon and pygidium. They demonstrate an L. trisectus-like morphology, with strongly tripartite pygidial posteroaxis and strongly convex glabella, which is unlike L. americanus from the type locality in Laurentia (Westrop, Adrain & Landing, Reference Westrop, Adrain and Landing2011, figs 5, 6; Rushton, Reference Rushton2009, fig. 1J–O). We remain unconvinced that such disparate morphotypes record a single, globally distributed species and, despite arguments to the contrary (Peng et al. Reference Peng, Babcock, Zhu, Zuo and Dai2013), consider reports of purported Lotagnostus americanus to be unacceptable as an index for the base of the uppermost Cambrian ‘Stage 10’ (see Westrop, Adrain & Landing, Reference Westrop, Adrain and Landing2011 for further discussion).
Ahlberg & Ahlgren (Reference Ahlberg and Ahlgren1996) and Ahlberg & Terfelt (Reference Ahlberg and Terfelt2012, fig. 2) showed that agnostoids are confined to particular segments of the Furongian succession of Baltica. Three agnostoid-bearing intervals are separated by packages of olenid zones from which they are absent. Section MaS conforms to this general pattern, albeit with the caveat that agnostoids are rare (<5% of trilobite and agnostoid sclerites present) where they do occur.
Almost all of the section lies within one of the Baltic and Avalonian intervals free of agnostoids that extends from the upper Parabolina Superzone (with shell beds of a rhynchonelliform brachiopod species traditionally identified as Orusia lenticularis Wahlenberg, Reference Wahlenberg1821; e.g. Walcott, Reference Walcott1912; Henningsmoen, Reference Henningsmoen1958; discussed in Section 5.f) through almost all of the Protopeltura Zone, with the uppermost 35 cm of the section recording the lower part of the succeeding agnostoid-bearing strata with Lotagnostus. Controls on agnostoid distribution are unclear; Ahlberg & Terfelt (Reference Ahlberg and Terfelt2012, p. 1009) speculated that diagenetic dissolution of sclerites might be partly responsible for their local absence from Alum Shale successions, although this explanation seemingly fails given the occurrence of olenids and calcitic brachiopods in at least some of their agnostoid-free intervals. However, apparent environmental controls on the occurrences of agnostoids in both Baltica and Avalonia (discussed in Section 6) add to the concerns of using the first appearance of any species of Lotagnostus as a datum for Stage 10.
4.c. Biostratigraphical evidence for stratigraphical condensation
The trilobite and agnostoid faunas show that some Avalonian successions are at least as condensed as those of Baltica (e.g. Terfelt, Ahlberg & Eriksson, Reference Terfelt, Ahlberg and Eriksson2011, fig. 1), and emphasize that such condensed sections are inadequate to provide global boundary stratotype section and points (GSSPs) for a terminal Cambrian stage. There are lingering uncertainties over the age of the lowest Orusia lenticularis (Fig. 6; discussed in Section 5.b) shell beds that influence our assessment of the extent of condensation in the lower segment of the section below the calcrete-clast conglomerate. If the lowest, closely spaced Orusia-rich horizons at 3.0 m and 3.7 m record the Parabolina spinulosa Zone as they do in Baltica and British Avalonia (e.g. Martinsson, Reference Martinsson and Holland1974; Rushton, Reference Rushton and Holland1974; Rushton et al. Reference Rushton, Brück, Molyneux, Williams and Woodcock2011), the lower–middle Leptoplastus Superzone (L. paucisegmentatus – L. crassicornis zones of Nielsen et al. Reference Nielsen, Weidner, Terfelt and Høyberget2014, fig. 1) is represented by barely 20 cm of strata. This is even thinner than the strongly condensed coeval interval in the Andrarum region of southern Sweden (60–70 cm; see Ahlberg et al. Reference Ahlberg, Månsson, Clarkson and Taylor2006, fig. 3; Terfelt, Ahlberg & Eriksson, Reference Terfelt, Ahlberg and Eriksson2011, fig. 1) and perhaps reflects the period of subaerial exposure recorded by the calcrete clast-bearing conglomerate (discussed in Section 5.e). With firm constraints on MaS-3.9 (L. ovatus Zone) and MaS-6.15 (C. tumida Zone), the upper segment of the section is also somewhat more condensed than at Andrarum. At the MaS locality, the equivalents of the uppermost Leptoplastus and Protopeltura superzones (L. stenotus – C. spectabilis zones; Nielsen et al. Reference Nielsen, Weidner, Terfelt and Høyberget2014) are 2.25 m in thickness, compared to 3.3 m at Andrarum (Terfelt, Ahlberg & Eriksson, Reference Terfelt, Ahlberg and Eriksson2011, fig. 1).

Figure 6. Orusia lenticularis coquina (from MaS-5.0) above middle Leptoplastus Superzone fauna (at MaS-3.9), NBMG 17869.
5. Deposition and correlation of MaS section
5.a. Black mudstones
The dominant dark-grey to black mudstones have a relatively small amount of fine-grained pyritohedral pyrite. Due to regional chlorite grade metamorphism, organic carbon is absent and total carbon (probably graphite) comprises 1–2% of the mudstone (5 samples tested). Despite a thorough field search and inspection of polished slabs, the mudstones show no evidence of burrowing, with the exception of a single large specimen of Chondrites Sternberg, Reference Sternberg1833 (Fig. 3), a burrow characteristic of low-oxygen facies (Ekdale & Mason, Reference Ekdale and Mason1988). No evidence of metazoan body fossils, whether originally calcareous or phosphatic (lingulate) sclerites, secondarily pyritized or phosphatized sclerites or moldic impressions of calcareous trilobites or brachiopods, were found in the black mudstones.
The scarcity of trace fossils and absence of any type of body fossils suggest that pre-compaction dissolution during diagenesis in the upper dysoxic environment does not explain the lack of fossils in the black mudstones of the MaS section (e.g. Schovsbo, Reference Schovsbo2001). Rather, persistent very-low-oxygen concentrations or high-frequency anoxic events likely accompanied deposition of the black mudstones. However, wave-rippled silt and mud pellet laminae, planar silt laminae and discontinuous, inclined silt laminae are common through the black mudstones and demonstrate a non-stagnant, wave- and current-controlled depositional environment of the black mudstone. This interpretation implies that fossiliferous limestone beds within the succession likely record periodic increases in oxygenation.
5.b. Rhynchonelliform brachiopod limestones: record of biotic change associated with a ‘biomere’ extinction?
Monospecific associations of an early orthid brachiopod comparable to traditional interpretations of Orusia lenticularis (Wahlenberg, Reference Wahlenberg1821) form medium-grey-coloured pack- to wackestone beds and nodular limestones at 3.0 and 3.7 m (Figs 3, 6, 7a). The genus also occurs as well-sorted brachiopod rudstone lenses with rare olenid fragments at 5.0 m (Fig. 6). The sorting and winnowing and convex side-up orientation associated with these brachiopod valve limestones records intervals of dynamic wave activity. In addition, the good preservation of the valves indicates limited transportation and essentially in situ specimens. The occurrence of rhynchonelliform brachiopods in sufficient abundance to form shell beds is unexpected in shallow-water upper Cambrian successions, and points towards unusual ecological conditions.

Figure 7. Lithologies from the MaS- section. (a) Fining-upward, monospecific Orusia lenticularis rudstone at horizon MaS-3.0, NBMG 17865. (b–e) Sections through the MaS-3.9 conglomeratic limestone with dyke of pyritic siliciclastic mudstone (psm); (b, d) large, dark grey, fine-grained, diagenetic calcareous nodules at top of bed (cns) and lenses of phosphatic sand and granules (ph) and pyrite sand (ps) that yielded Leptoplastus cf. ovatus sclerites, respectively NBMG 17866 and NBMG 17867; (b–d) clasts of non-fossiliferous, light to medium grey, laminar caliche hardpan caliche (cal) composed of microsparitic carbonate laminae with quartz sand (small white grains), scattered darker-grey glaebules (gl) and subhorizontal, caliche calcite spar (scr); (b, d, e) pebbles include rounded white clasts dominated by syntaxial calcite rims (sc) and sparry (bladed) calcite (bs) that envelope fragments of black siliciclastic mudstone and caliche; (d) pyritic siliciclastic mudstone (psm) dyke fills intraclast space; (e) enlarged view of clast with bladed (sparry) calcite coating (bc) in Fig. 3d; (f) dark-grey, sparse trilobite-agnostoid wackestone (lower part) with middle dark packstone layer broken into pull-apart clasts (pcl), upper half of sample an olenid-rare agnostoid fragment and lime mudstone granule grainstone, sample MaS-6.15, NBMG 17868.
The high abundance of Orusia lenticularis parallels the early Sunwaptan (Jiangshanian) succession in Laurentian North America, where the orthids Eoorthis Walcott, Reference Walcott1908 and Billingsella Hall & Clarke, Reference Hall and Clarke1892 appear abruptly and form rudstone beds in shallow subtidal settings in the wake of the trilobite extinctions at the base and top of the Irvingella major Zone (e.g. Berg, Reference Berg1954; Bell & Ellinwood, Reference Bell and Ellinwood1962; Grant, Reference Grant1965; Freeman & Stitt, Reference Freeman and Stitt1996; Westrop, Reference Westrop1996; S.R. Westrop, unpub. data). Abundant orthids extend into the lower Sunwaptan as far as the lower Saratogia Zone in Oklahoma (Freeman & Stitt, Reference Freeman and Stitt1996); similar brachiopod shell beds are associated with the end-Sunwaptan extinction in Texas (Winston & Nicholls, Reference Winston and Nicholls1967) and Alberta (S. R. Westrop, unpub. data, 2015).
Orusia lenticularis was described originally from Baltica (Sweden) and is reported widely, although the absence of a modern systematic revision may limit the utility of such records. Throughout Baltica, the species occur in both zones of the Parabolina Superzone (Nielsen et al. Reference Nielsen, Weidner, Terfelt and Høyberget2014), and may form shell beds in the younger P. spinulosa Zone (e.g. Terfelt, Reference Terfelt2003). Rushton & Weidner (Reference Rushton and Weidner2010) have shown that the highest occurrence of Irvingella, a cranidium that they assign to I. major Ulrich and Resser, in Walcott, is in the underlying P. brevispina Zone. As in Laurentia, orthid shell beds therefore appear immediately above Irvingella. Records of Orusia from Avalonia are also predominantly from the P. spinulosa Zone, although Rushton (in Rushton et al. Reference Rushton, Brück, Molyneux, Williams and Woodcock2011, p. 31) indicates that the genus occurs in higher strata of Stage 10 in the Malvern area of the English Midlands and in the Ogof-ddu section in North Wales (Allen, Jackson & Rushton, Reference Allen, Jackson and Rushton1981, p. 314). Similarly, Matthew's (Reference Matthew1901, Reference Matthew1903) record of Orusia with Peltura scarabaeoides in southern New Brunswick and Cape Breton Island is assignable to Stage 10. In our MaS section, Orusia also ranges into higher Jiangshanian strata than in Baltica, with the youngest shell lenses at MaS-5.0 (Fig. 3) bracketed by an upper Leptoplastus Superzone fauna at MaS-3.9 and an upper Protopeltura Superzone fauna at MaS-5.8 m. The lowest Orusia shell beds at MaS-3.0 and MaS-3.7 (Fig. 3) are of uncertain age as there is no biostratigraphic bracket below them. However, based on all other first appearances of Orusia recorded elsewhere in Avalonia and in Baltica, they are likely no older than the Parabolina Superzone and probably correlate with its P. spinulosa Zone.
In Laurentia, the appearance of rhynchonelliform brachiopods a short distance above the base of the Sunwaptan Stage is part of the biotic changes associated with trilobite extinction at the end of the ‘Pterocephalid Biomere’. It is tempting to suggest that the apparently coeval entry of Orusia in successions of Avalonia and Baltica is also a record of similar, albeit more cryptic, biotic change in quite different palaeogeographic settings than tropical Laurentia. Compared to Laurentia, Paibian – early Jiangshanian (equivalent to the Steptoean Stage of Laurentia) trilobite faunas of Avalonia and Baltica are of much lower diversity and often dominated by one olenid genus, which is Olenus Dalman, Reference Dalman1827 (e.g. Terfelt et al. Reference Terfelt, Ahlberg and Eriksson2011, fig. 1).
The disappearance of Olenus along with such non-olenid genera as Proceratopyge Wallerius, Reference Wallerius1895 may well record the same extinction interval as in Laurentia, followed by the immigration of brachiopods. That is, the end-Steptoean extinction event (or the end of the ‘Pterocephaliid Biomere’; Westrop & Cuggy, Reference Westrop and Cuggy1999) is best expressed in Laurentia but can also be identified in other continents and terranes (see also Westrop & Adrain, Reference Westrop and Adrain2013 for evidence that this extinction can be recognized in the Karatau–Naryn Terrane of Kazakhstan).
Orusia-rich horizons also offer information on environmental conditions and sea-level changes. In Avalonian Britain, they appear with marine onlap and the onset of black mudstone deposition just above the shallow-water sandstones of the Ffestiniogg Formation of the Welsh Basin (Allen, Jackson & Rushton, Reference Allen, Jackson and Rushton1981). Similarly, Orusia is the first fossil to appear with onlap across a karstic limestone with caves in the Alum Shale at Kinnekulle, Sweden (Lehnert et al. Reference Lehnert, Calner, Ahlberg and Harper2012). These occurrences are in agreement with the shallow-marine, more oxygenated water column habitat given to the species in Swedish Baltica (Dworatzek, Reference Dworatzek1987), where the species occurs in Alum Shale facies that Ahlberg & Terfelt (Reference Ahlberg and Terfelt2012) interpreted as regressive.
The biofacies represented by the monospecific Orusia lenticularis assemblages at 3.0 m and 3.7 m records high-energy, wave-dominated and, presumably, better-oxygenated facies than intervals that lack orthids and yield only olenid trilobites (e.g. Ahlberg et al. Reference Ahlberg, Szaniawski, Clarkson and Bengtson2005, Reference Ahlberg, Månsson, Clarkson and Taylor2006). Orusia lenticularis lenses re-appear with continued submergence following forced regression, subaerial exposure and initial transgression indicated by the calcrete-clast conglomerate at 3.9 m (discussed in the following section). It is, however, uncertain whether occurrences of the Orusia biofacies represent short- or long-term intervals of higher bottom-water oxygenation and ambient energy.
5.c. Calcrete-bearing conglomerate: clast types
The MaS-3.9 bed is an intraclast conglomerate with a surprisingly complex history of development that included sea-level fall, an interval of subaerial exposure and the onset of the succeeding rise in sea level. This conglomerate also yields olenid trilobites from its matrix, which help bracket the timing of sea-level changes.
5.c.1. Carbonate nodule and siliciclastic mudstone clast
Multiple slabs cut through the MaS-3.9 bed show that about half of the clasts consist of pebble- to cobble-sized, discoid fragments of dark-grey, non-fossiliferous microspar (Fig. 7b, d, e; ‘cnc’) with depleted carbon isotope values (δ13C = −17‰) consistent with methanogenic carbonate nodules (e.g. Hicks et al. Reference Hicks, Compton, McCracken and Vecsei1996). These dark-grey nodules are common in the Chesley Drive Group in Maritime Canada and eastern Newfoundland (e.g. Landing, Reference Landing1980, fig. 2; Landing & Fortey, Reference Landing and Fortey2011). A small number of clasts are composed of black siliciclastic mudstone of the type that dominates the MaS section (Fig. 7b, clast labeled ‘mc’).
5.c.2. Calcrete clasts
Associated fragments of a lighter-grey, laminated limestone are composed of a dense mosaic of blocky calcite microspar crystals that range up to 1.0 mm in diameter (Fig. 7b, ‘cal’) and include abundant quartz sand. In addition, rounded, darker-grey clasts with rounded glaebules (Fig. 7b, ‘gl’ at arrow) and calcite-filled, subhorizontal fractures occur in the light-coloured clasts (Fig. 7b, c; ‘src’ at arrow). A number of limestone clasts dominated by blocky calcite spar have a large proportion of dark-grey grains composed of siliciclastic mudstone (Fig. 7c, small ‘cal’ clasts).
The laminated clasts best suggest a subaerial environment of formation because they are comparable to laminar caliche hardpan or the laminar calcrete of Goudie (Reference Goudie, Goudie and Pye1983). Laminar calcrete is composed of microsparitic carbonate with small glaebules and abundant quartz sand (e.g. Esteban & Klappa, Reference Esteban, Klappa, Scholle, Bebout and Moore1983, fig. 75). The isotopic composition of these clasts (δ13C = −7‰) is consistent with the depletion in 13C compared to marine limestones, which average ± 0‰ (e.g. Schlesinger, Marion & Fontey, Reference Schlesinger, Marion and Fontey1989; Ainóczi et al. Reference Ainóczi, Horváth, Demény and Mindszenty2006). Further evidence of a subaerial environment during development of the laminar calcrete clasts is shown by the subhorizontal, roughly parallel white fractures filled with calcite (Fig. 7b, c, ‘src’). These calcite-filled fractures are comparable to the hair-like cracks in calcretes (e.g. Allen, Reference Allen and Wright1986, fig. 14d).
The smaller ‘dirty’ pebbles with abundant black siliciclastic grains scattered in a matrix of microspar are also regarded as a caliche (Fig. 7c, ‘cal’). These smaller pebbles show the disordered fabric of Brewer's (Reference Brewer1964) agglomeratic fabric (also Allen, Reference Allen and Wright1986, fig. 14c).
5.c.3. Nodular phosphate and reworked pyrite sand
The finest-grained, reworked fragments in MaS-3.9 occur between the pebbles and cobbles. These include rounded, black, sand- to granule-sized fragments of nodular phosphate (Fig. 7b, ‘ph’) reworked from the black Chesley Drive Group mudstone. In addition, rounded grains of fine- to coarse-grained pyrite sand also occur in the matrix between the pebbles (Fig. 7b, ‘py’). The clastic pyrite sand, also reworked from the black mudstones, would have been unstable in oxic environments; its occurrence suggests strongly dysoxic bottom waters during accumulation of the MaS-3.9 conglomerate. Other occurrences of clastic pyrite during Precambrian (Simpson & Bowles, Reference Simpson and Bowles1977), Middle Ordovician (Landing, Benus & Whitney, Reference Landing, Benus and Whitney1992), Silurian (Love, Reference Love1971) and Devonian (Baird & Brett, Reference Baird and Brett1986) time are all related to basins with anoxic to strongly dysoxic bottom waters. However, the association of well-preserved (non-abraded and non-transported) sclerites of olenid trilobites with the pyritic sand would seem to preclude entirely anoxic bottom water (e.g. Fig. 1).
5.d. Calcrete-bearing conglomerate: cements
Two types of pre-burial cements coat the clasts in the conglomerate. They are more conspicuous than the microcrystalline calcite that lines the hair-like cracks in the calcrete clasts and which comprises the agglomeratic fabric.
5.d.1. Syntaxial calcite cement
The older cement is an almost-opaque white calcite cement that forms thick rinds around a number of the calcrete pebbles (Fig. 7b, d, ‘sc’). This rind is composed of unicrystalline calcite, as shown by its rhomboidehedral cleavage (Fig. 7b, upper ‘sc’). Syntaxial overgrowths are common by a process of ‘secondary enlargement’ on unicrystalline grains such as echinoderm ossicles (e.g. Bathurst, Reference Bathurst1975). However, polycrystalline grains, as for the caliche clasts in MaS-3.9, can also have syntaxial rims comprising one crystal of calcite (e.g. Evamy & Shearman, Reference Evamy and Shearman1969).
The environment of formation of the thick syntaxial calcite rims is problematical. Relatively coarse-grained (i.e. sand-sized or coarser) calcite crystal cements are rare in any diagenetic environment. Coarse-grained spar crystals are less commonly associated with pedogenesis than with early diagenesis in shallow subaerial environments with a supersaturation of calcium and bicarbonate. These environments can include shoreline muds of high-salinity lakes (Hay & Kyser, Reference Hay and Kyser2001).
However, calcretes with sparry, displasive calcite also prove to be common in dry regions at horizons in shallow subterranean environments that lack significant biological activity and in which fluctuating groundwater levels regularly wet soft sediment (Watts, Reference Watts1978; Nash & Smith, Reference Nash and Smith2003; Bainóczi et al. Reference Bainóczi, Horváth, Demény and Mindszenty2006; Sedov et al. Reference Sedov, Solleiro-Rebolledo, Fedick, Pi pulg, Vallejo-Gómez, Lourdes Flores-Delgadillo and Kapur2008). The environment in which the white syntaxial cement grew was one with limited suspended sediment which otherwise would have coloured the cement.
High-energy benthic environments after cementation and prior to deposition of the MaS-3.9 conglomerate are indicated by the well-rounded form of the clasts rimmed by syntaxial calcite. Probable wave-associated abrasion wore away all pyramidal faces of the syntaxial cement and even locally abraded the syntaxial cement to expose the calcrete nucleus of some clasts (Fig. 7d, middle ‘sc; on image).
5.d.2. Bladed calcite cement
Radiaxial calcite spar crystals developed on a wide variety of clasts in the MaS-3.9 conglomerate. Well-preserved, medium- to dark-grey-coloured calcite scalenohedrons occur on siliciclastic mudstone pebbles, exhumed calcareous nodules and on calcrete clasts (Fig. 7b, d; ‘bc’ for bladed calcite). The fragile scalenohedron calcite crystal rinds on the clasts would not have survived transport, and are interpreted to have grown after deposition of the clasts to occlude inter-clast spaces.
Radiating calcite cement is consistent with cementation under subaerial exposure, although it also appears in meteroric or submarine environments (James, Reference James1972). In the case of the bladed sparry cement from the MaS-3.9 conglomerate, the medium to dark-grey colour of the scalenohedrons suggests growth in turbid water produced by wave and current action on the dark-grey to black siliciclastic muds of the Chesley Drive Group.
5.e. Calcrete-bearing conglomerate: deposition
The thin MaS-3.9 bed reveals a complex history that includes a relative sea-level fall that led to subaerial exposure and production of a calcrete. In terms of geological time, calcretes form relatively rapidly (e.g. Sedov et al. Reference Sedov, Solleiro-Rebolledo, Fedick, Pi pulg, Vallejo-Gómez, Lourdes Flores-Delgadillo and Kapur2008). However, the association in bed MaS-3.9 of clasts of a light-grey, laminated calcrete and a darker-grey calcrete with an agglomeratic fabric and grains of dark siliciclastic mudstone all suggest that the bed may represent multiple episodes of calcrete formation. Other than the calcrete, evidence of subaerial exposure, such as bleaching of the underlying silicicilastic mudstone or presence of bleached (oxidized) mudstone clasts, is not present in the section.
Fragmentation of this caliche took place with at least two intervals of relative sea-level rise. The first rise led to the break-up of the caliche and was followed by the development of syntaxial calcite rims on the caliche clasts. At least one additional sea-level rise (perhaps preceded by a fall) led to rounding and severe abrasion of the calcrete clasts already coated with syntaxial cement. At this same time, it is possible that further fragmentation of the calcrete(s) took place, along with the erosion and deposition of calcareous nodules and black mudstone clasts and the accumulation of pyrite sand, small phosphate fragments and olenid trilobite debris between the larger clasts. The local presence of trough cross-bedding in the finer-grained parts of MaS-3.9 records high wave/current energies (Fig. 3). Wave ripples on the top of MaS-3.9 (Fig. 3) record the last episode of high ambient energy associated with deposition of the conglomerate bed. As the trilobites introduced during the final transgressive phase of the conglomerate's history belong to the Leptoplastis ovatus Zone, the preceeding sea-level fall and emergence probably occurred in the lower part of the Leptoplastus Superzone.
Growth of bladed, scalenohedral calcite cement took place after the final deposition of the clasts. This fragile cement grew in the turbid water that filled the open spaces between the clasts. Dark-grey mud was incorporated into this radiaxial cement, and is responsible for its dark colour. The erosion, rounding and deposition of pyrite sand indicates the presence of dysoxic water which allowed preservation of the remané pyrite as it was rounded, but apparently the bottom water had enough oxygen for olenid trilobites.
The last depositional event was the opening of small fissures that cut through the bed and allowed the injection of dark-grey, pyritiferous siliciclastic mudstone into the fissures (Fig. 7d, ‘psm’). All of the fissure fills revealed by slabbing have trilobite debris reworked from slightly higher in the MaS section. They are not upward injections of mud from under the bed (i.e. Neptunian dykes), but are fissure fills packed with soft siliciclastic mud that accumulated on top of the conglomerate bed. Fracturing of the bed either by subsidence and compaction of the underlying mud or gentle downslope creep of the bed are two possible methods by which the fissures could have originated.
As noted above, the probable lower Leptoplastus Superzone interval between the Orusia shell beds and the conglomerate is strongly condensed. There is a break in the ‘customary’ deposition of Chesley Drive Group black mud, and the evidence of subaerial exposure means that it marks a type 1 depositional sequence boundary (van Wagoner et al. Reference van Wagoner, Posamentier, Mitchum, Vail, Sarg, Louit, Hardenbol, Wilgus, Posamentier, Ross and Kendall1988) in an Alum Shale-type facies.
5.f. Upper part of MaS section
Above the calcrete clast-bearing conglomerate, the upper 2.6 m of the MaS section consists of an alternation of dark grey to black siliciclastic mudstone and light grey weathering, nodular to bedded fossil rudstone (Figs 3, 7f). The lowest of these is represented by lenses of Orusia lenticularis valves (MaS-5.0; Figs 6, 7a) with a probably wave-winnowed fabric. Higher shell beds composed of olenid trilobites at MaS-5.8 and -6.15 show an internal alternation of spar-rich and spar-poor rudstone layers (Fig. 7f) that likely record variations in winnowing and sediment supply in condensed beds (e.g. Dattilo et al. Reference Dattilo, Brett, Tsujita and Fairhurst2008). The overall succession above the caliche conglomerate therefore shows upward changes in biofacies although, as discussed in the following section, the presence of condensed shell beds may preclude a simple Waltherian interpretation. In any event, the various shell beds demonstrate periodic oxygenation of the sea floor.
Fracturing of the darker wackestone layers (Fig. 7f), whether by tension with downslope movement (sedimentary boudinage) or differential compaction of underlying sediment, is shown by repeated sections through the olenid limestones that show that the fractured wackestone layers consist of large (10+ cm), plate-like layers that are far larger than the granule-sized intraclasts in the olenid limestones. This fracturing and differential ‘sinking’ of the trilobite wackestone layers further suggests sedimentary condensation as the wackestones were relatively consolidated, either by compaction or the early growth of cement before fracture.
6. Black mudstone depositional history and upper Cambrian sea-level changes
The presence of relatively high-energy depositional intervals recorded by the fossil rudstones of section MaS seems initially inconsistent with the deeper-water setting traditionally assigned to low-oxygen environments represented by pyritic black mudstones with Chondrites as the only known trace fossil. However, recent syntheses indicate that hypoxic mudstones were deposited in relatively shallow water during intervals of extremely high sea levels and high global temperatures, with very low oxygen conditions in the epeiric seas promoted by high temperatures, freshwater caps and local water column turbidity during early–middle Palaeozoic time. Indeed, the presence of black mudstones on unconformity surfaces developed during subaerial exposure is best interpreted as indicating hypoxic epeiric sea deposition that even extended into nearshore environments (e.g. Landing, Reference Landing2012, Reference Landing, Derby, Fritz, Longacre, Morgan and Sternbach2013 a, b; Landing & Smith, Reference Landing and Smith2012). At the same time, the shell beds also demonstrate that these environments periodically became sufficiently oxygenated to support low-diversity rhynchonelliform brachiopod and trilobite faunas.
In the confines of a single stratigraphic section, our ability to interpret sea-level history is limited. Nonetheless, the calcrete-bearing conglomerate within section MaS provides evidence for at least two previously unrecognized cycles of major sea-level change punctuated by subaerial emergence. The lower of these, terminated by emergence that led to calcrete formation, may correspond in part to the middle Jiangshanian, Orusia-dominated interval of Baltica, thought to be regressive by Ahlberg & Terfelt (Reference Ahlberg and Terfelt2012, p. 1009).
The relative thicknesses of olenid zones in the Alum Shale of southern Sweden (Ahlberg & Terfelt, Reference Ahlberg and Terfelt2012, fig. 1) provide indirect evidence for sea-level changes in Baltica that are broadly coeval with those described here for Avalonia. In southern Sweden, the Leptoplastus and basal Protopeltura superzones comprise a very thin interval, only 1.4 m thick (Ahlberg & Terfelt, Reference Ahlberg and Terfelt2012, fig. 1; numerical data from Terfelt, Ahlberg & Eriksson, Reference Terfelt, Ahlberg and Eriksson2011, fig. 1). As discussed above, the relative sea-level fall associated with the caliche clast conglomerate is likely pre-L. ovatus Zone in age. Immediately below the L. ovatus Zone in Sweden, the L. crassicornis and L. raphidophorus zones are only 10 cm each in thickness (Terfelt, Ahlberg & Eriksson, Reference Terfelt, Ahlberg and Eriksson2011, fig. 1). This stratal thinning is consistent with a sharp reduction in the rate of creation of accommodation space during late regression. Above the L. ovatus Zone, the combined thicknesses of the L. stenotus and L. neglectus zones is only 20 cm (Terfelt, Ahlberg & Eriksson, Reference Terfelt, Ahlberg and Eriksson2011, fig. 1). This interval is one of sea-level rise in Avalon; in Baltica, the L. stenotus – L. neglectus zones might record condensation that may occur in transgressive systems tracts (e.g. Kidwell, Reference Kidwell, Einsele, Ricken and Seilacher1991). The evidence is circumstantial, but we argue that the same Leptoplastus Superzone sea-level fall and rise are expressed in both Avalonia and Baltica, with the inference that they are eustatic in nature. The Leptoplastus Superzone of Baltica would likely merit further study.
The conglomerate is a condensed unit that records marine transgressions that fragmented and reworked subaerially formed calcrete and filled the inter-clast spaces with a pyritiferous mud with clastic pyrite sand, phosphate granules and trilobite sclerites. It is tempting to suggest that the overlying succession, which features replacement of rhynchonelliform brachiopod shell beds with olenid trilobite-rich shell beds, records continued upward deepening. However, in stratally thin, condensed successions that represent a considerable length of time, such as the Chelsey Drive Formation, a simple Waltherian interpretation may be inappropriate. Condensed shell beds such as those at MaS-5.8 and MaS-6.15 commonly form in transgressive intervals (e.g. Zecchin, Reference Zecchin2007, p. 24, 25). They may therefore be evidence of lower-rank cycles or ‘high-frequency sequences’ (e.g. Zecchin & Catuneanu, Reference Zecchin and Catuneanu2013) in section MaS. Under such an interpretation, the shell beds could be considered small-scale analogues of transgressive systems tracts, with the intervening mudstones recording analogues of highstand systems tracts. Longer-term sea-level change would therefore be expressed by stacking patterns of cycles. Data from sections in other localities are needed to properly evaluate these ideas, but it is conceivable that the sea-level history of condensed Cambrian black shale successions is more complex than previously suspected. Indeed, high-energy, transgressive intervals may offer a means of mixing the water column and elevating the oxygen levels demanded by the periodic appearances of skeletalized invertebrates.
The repeated occurrence of higher-energy depositional events associated with probably better-oxygenated Orusia and olenid associations through the short MaS section do not require significant changes in relative depth. The only relatively dramatic changes in depositional environments that may have occurred through the MaS section was a regression and subsequent transgression, significant in the production of subaerial environments, and a caliche followed by submergence that produced bladed calcite cement and featured the re-appearance of olenid trilobites. However, even the sea-level fall required for calcrete development at the MaS section may have been quantitatively minor, as evidence shows that the black organic-rich mudstones were deposited in shallow water in Baltica and Avalonia (discussed above). This interpretation of shallow-marine deposition of Alum Shale facies with subaerial exposure agrees with interpretations by Artyushkov, Lindström & Popov (Reference Artyushkov, Lindström and Popov2000) and Artyushkov, Tesakov & Chekhovich (Reference Atryushkov, Tesakov and Chekhovich2008), who have concluded that Cambrian–Ordovician eustatic changes were no more than 20 m in magnitude based, in particular, on evidence from Baltica.
7. Conclusions
Organic-rich, dark-grey mudstone was prominent in the middle middle Cambrian (traditional middle Acadian of Avalonia) – Tremadocian depositional history of two middle-latitude palaeocontinents. During this time interval, these mudstones covered much of Baltica. Similarly, the inner platform of Avalonia, with the exception of intervals of upper middle Cambrian (upper Acadian) shallow-water sandstones and upper lower Tremadocian wave-deposited silt- and sandstones, was also blanketed by coeval ‘Alum-Shale’-like mudstones. The traditional paradigm of stagnant, highly dysoxic to anoxic, deep-water environments to explain Alum Shale sedimentation across Baltica has ceded to an understanding that dynamic depositional processes with varying wave and current activity, changes in water mass oxygenation, variations in diversity and abundance of benthic organisms, and event beds and erosion surfaces are present in these supposedly ‘stagnant basin’ facies.
Similarly, a carbonate-bed-rich, highly condensed section in the upper Cambrian deposits of Avalonian Cape Breton Island demonstrates dramatic changes in benthic energy levels and oxygenation. Strongly dysoxic to (perhaps) anoxic black mudstones alternating with well-sorted and winnowed, often condensed, brachiopod- and olenid trilobite-rich limestones record intermittantly well-oxygenated conditions. Perhaps the most surprising development is the presence of a caliche-clast conglomerate in the Leptoplastus Superzone, that demonstrates subaerial exposure and a forced regression in this black mudstone succession followed by transgression. Patterns of stratal thinning expressed by variation in olenid zone thickness in the Leptoplastus Superzone of Baltica (Terfelt, Ahlberg & Eriksson, Reference Terfelt, Ahlberg and Eriksson2011; Ahlberg & Terfelt, Reference Ahlberg and Terfelt2012) possibly record the same sea-level changes.
Olenid trilobites and orthid brachiopods allow close biofacies and biostratigraphic comparison between the Cape Breton and other Avalonian successions with Baltica, demonstrating that barriers to faunal exchange were minimal, particularly during late Cambrian time. An Acado-Baltic Faunal Province can be recognized in the Furongian–Tremadocian deposits, although Avalonia and Baltica have significantly different macrofaunas in the earlier part of the Cambrian (e.g. Landing, Westrop & Bowring, Reference Landing, Westrop and Bowring2013). Orthid shell beds occur at correlative levels in the Baltic, Avalonian and Laurentian successions. In Laurentia, they occur in the aftermath of well-documented trilobite extinctions at the base of the Sunwaptan stage, and are likely associated with similar, but more cryptic, turnover and biotic change in Avalonia and Baltica.
Faunal similarity between Baltica and Avalonia extends to similar Lotagnostus species in dysoxic, olenid-bearing facies that resemble the compacted L. trisectus topotypes (Rushton, Reference Rushton2009) from Avalonian Britain. However, the Swedish and the Cape Breton species are morphologically unlike L. americanus from well-oxygenated lithofacies in Laurentia (see Westrop, Adrain & Landing, Reference Westrop, Adrain and Landing2011 for further discussion) and cannot be used to define an ‘L. americanus Zone’ at the base of Cambrian Stage 10 in Avalonia and Baltica. An alternative method, which defines the base of a Cambrian Stage 10 at the lowest occurrence of the euconodont Eoconodontus notchpeakensis (Miller, Reference Miller1969), just below the onset of the HERB carbon isotope excursion, has been demonstrated to work globally from shallow- to deep-marine facies on almost all Cambrian continents (Landing, Westrop & Adrain, Reference Landing, Westrop and Adrain2011). Indeed, recent reports now establish the HERB excursion and the lowest occurrence of E. notchpeakensis and Cordylodus andersi Viira & Sergeyeva (in Kaljo et al. Reference Kaljo, Borovko, Heinsalu, Khasanovich, Mens, Popov, Sergeyeva, Sobolevskaya and Viira1986) lie in the transition from the thin Parabolina lobata Zone into the Peltura paradoxa Zone in Sweden (Bagnoli & Stouge, Reference Bagnoli and Stouge2014; Terfelt, Eriksson & Schmitz, Reference Terfelt, Eriksson and Schmitz2014). It is therefore possible to recognize a base of informal Cambrian Stage 10 based on the lowest occurrence of E. notchpeakensis/C. andersi and the onset of the HERB carbon isotope excursion in Baltica without the taxonomic difficulties of attempting to force a set of morphologically distinct, mid-Furongian Lotagnostus species into a single, purportedly pandemic ‘L. americanus’.
Acknowledgements
EL acknowledges support from the New York State Museum and from earlier National Science Foundation grants. Randy Miller (New Brunswick Museum) arranged loans of trilobites and agnostoids and supplied type numbers. J. Smith, previously of the New York State Museum, generously arranged for geochemical analysis of carbon content of the MaS samples. Two anonymous reviewers provided useful suggestions. The order of the authors’ names is alphabetical and does not indicate seniority.







