Introduction
Fossil preservation is a complex mechanism that involves different processes. Taphonomy is concerned with the information content of the fossil record and the processes by which fossils are incorporated into the fossil record (Martin, Reference Martin1999). The taphonomy studies are applied to paleobiological and paleoecological questions (Behrensmeyer et al., Reference Behrensmeyer, Kidwell and Gastaldo2000). In addition, the postmortem modification of shelly fossils can provide valuable insights into paleohydraulic and diagenetic conditions (Kidwell and Bosence, Reference Kidwell and Bosence1991). In general, the taphonomy of marine invertebrates has been extensively explored compared to that of continental invertebrates.
Arthropod cuticles consist of chitinous fibers embedded in a protein matrix, cross-linked by catechol, aspartate, and histidyl moieties (Schaefer et al., Reference Schaefer, Kramer, Garbow, Jacob, Stejskal, Hopkins and Speirs1987). Calcification, in the form of calcium carbonate, further strengthens the cuticle of many crustaceans and extinct trilobites. Such biomineralized skeletons dominate the marine fossil record of arthropods (Gupta et al., Reference Gupta, Michels, Briggs, Evershed and Pancost2006). Spinicaudatans (‘clam shrimps’) occur in continental facies from the Devonian to the present (Tasch, Reference Tasch1969; Thiéry, Reference Thiéry1996). The exoskeleton of these invertebrates consists of a thin outer epicuticle and an internal procuticle. The latter is multilamellar and positioned parallel to the surface; often, the exocuticle and endocuticle are divided into sublayers (Warth, Reference Warth1969; Rieder et al., Reference Rieder, Abaffy, Hauf, Lindel and Weishäupl1984; Vannier et al., Reference Vannier, Thiéry and Racheboeuf2003).
Continental invertebrates in Mesozoic facies are mainly represented by spinicaudatans, ostracods, mollusks, and insects. The spinicaudatans are considered good biostratigraphic markers due to their wide geographic distribution (Kozur and Weems, Reference Kozur and Weems2010). They are small branchiopod crustaceans and have a soft body enclosed in a chitin-phosphate bivalve carapace that is often the only element preserved in the fossil record (Tasch, Reference Tasch1969; Thiéry, Reference Thiéry1996). Observations by Stigall et al. (Reference Stigall, Babcock, Briggs and Leslie2008) suggested that the carapace of spinicaudatans is preserved primarily in calcium phosphate, reflecting the original skeletal composition. Recently, Astrop et al. (Reference Astrop, Sahni, Blackledge and Stark2015) demonstrated that the mechanical properties and chemical composition of the carapace from living clam shrimps could affect the susceptibility of carapaces to taphonomic processes. However, the mechanisms of fossilization in spinicaudatans are not completely understood, and few studies have been carried out regarding their taphonomy.
Relatively few chemical studies on the elemental composition of spinicaudatan carapaces have been performed using scanning electron microscopy (SEM) in combination with energy-dispersive X-ray spectrometry (EDS). This is the case for a number of studies of the inorganic chemical composition associated with different modes of fossil preservation (e.g., Kohn et al., Reference Kohn, Schoeninger and Barker1999; Klug et al., Reference Klug, Schulz and De Baets2009; Lin and Briggs, Reference Lin and Briggs2010).
The aim of this study is to contribute to a better understanding of the chemical preservation of fossil spinicaudatan carapaces from the Cañadón Asfalto Formation (Jurassic, Chubut, Argentina) and of the factors involved in this preservation.
Geologic setting
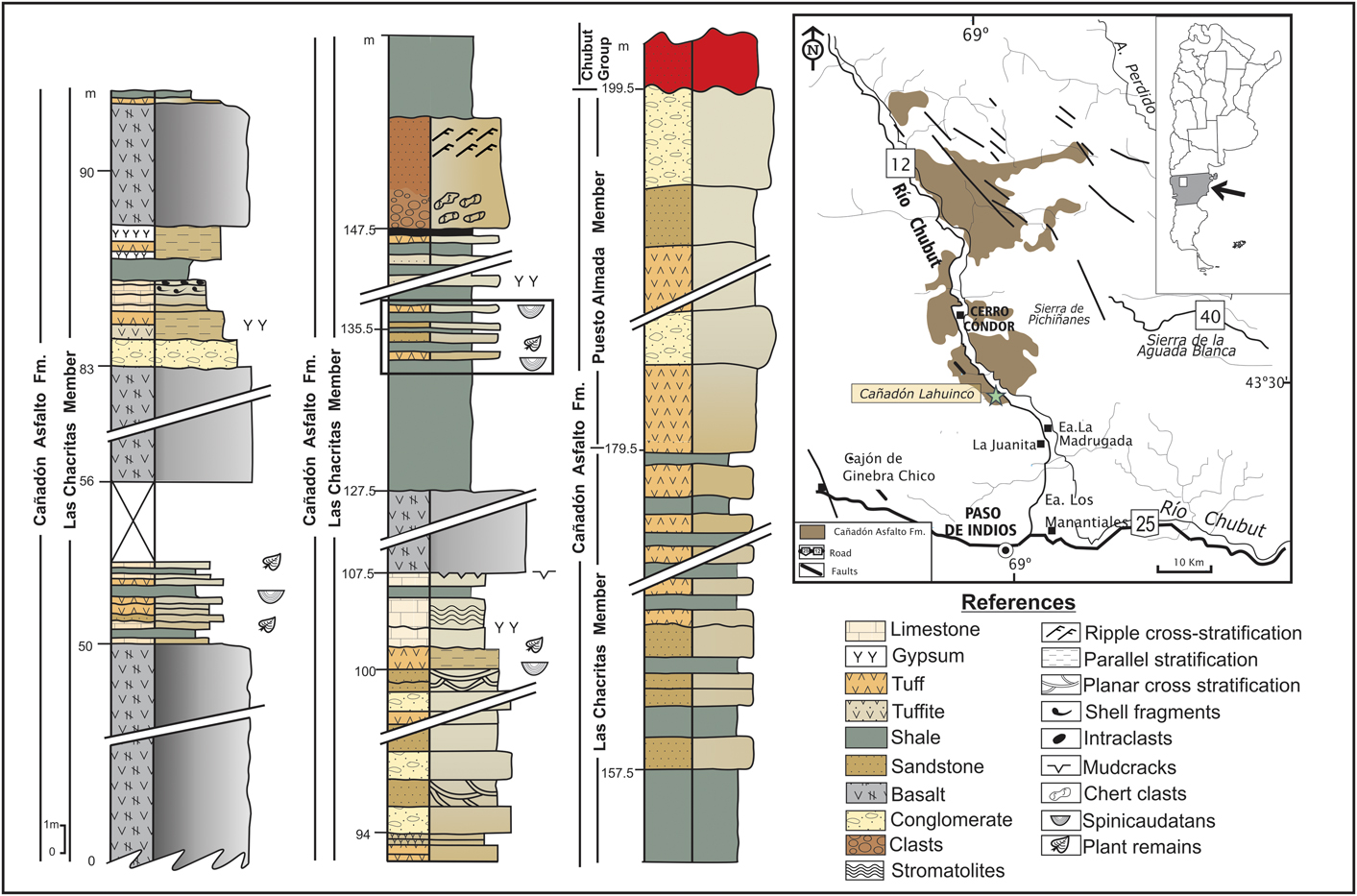
The continental Jurassic fauna in Argentina occurred in two great basins: the Cañadón Asfalto and Bahía Laura. The first basin is located in the extra-Andean region of the Chubut province, Patagonia, Argentina (Cabaleri et al., Reference Cabaleri, Volkheimer, Armella, Gallego, Silva Nieto, Cagnoni, Ramos and Panarello2010) (Fig. 1), and the latter is located in central Santa Cruz province. The Cañadón Asfalto Formation is the main unit of the Cañadón Asfalto basin (Stipanicic et al., Reference Stipanicic, Rodrigo, Baulíes and Martínez1968). The lacustrine systems of the Cañadón Asfalto Formation exhibit an important volcano-sedimentary association and represent one of the most important Jurassic continental fossil records of South America. The unit is characterized mainly by limestones, pyroclastic deposits, and basalts (Stipanicic et al., Reference Stipanicic, Rodrigo, Baulíes and Martínez1968; Turner, Reference Turner1983; Cabaleri et al., Reference Cabaleri, Volkheimer, Armella, Gallego, Silva Nieto, Cagnoni, Ramos and Panarello2010, Reference Cabaleri, Benavente, Monferran, Narváez, Volkheimer, Gallego and Do Campo2013).
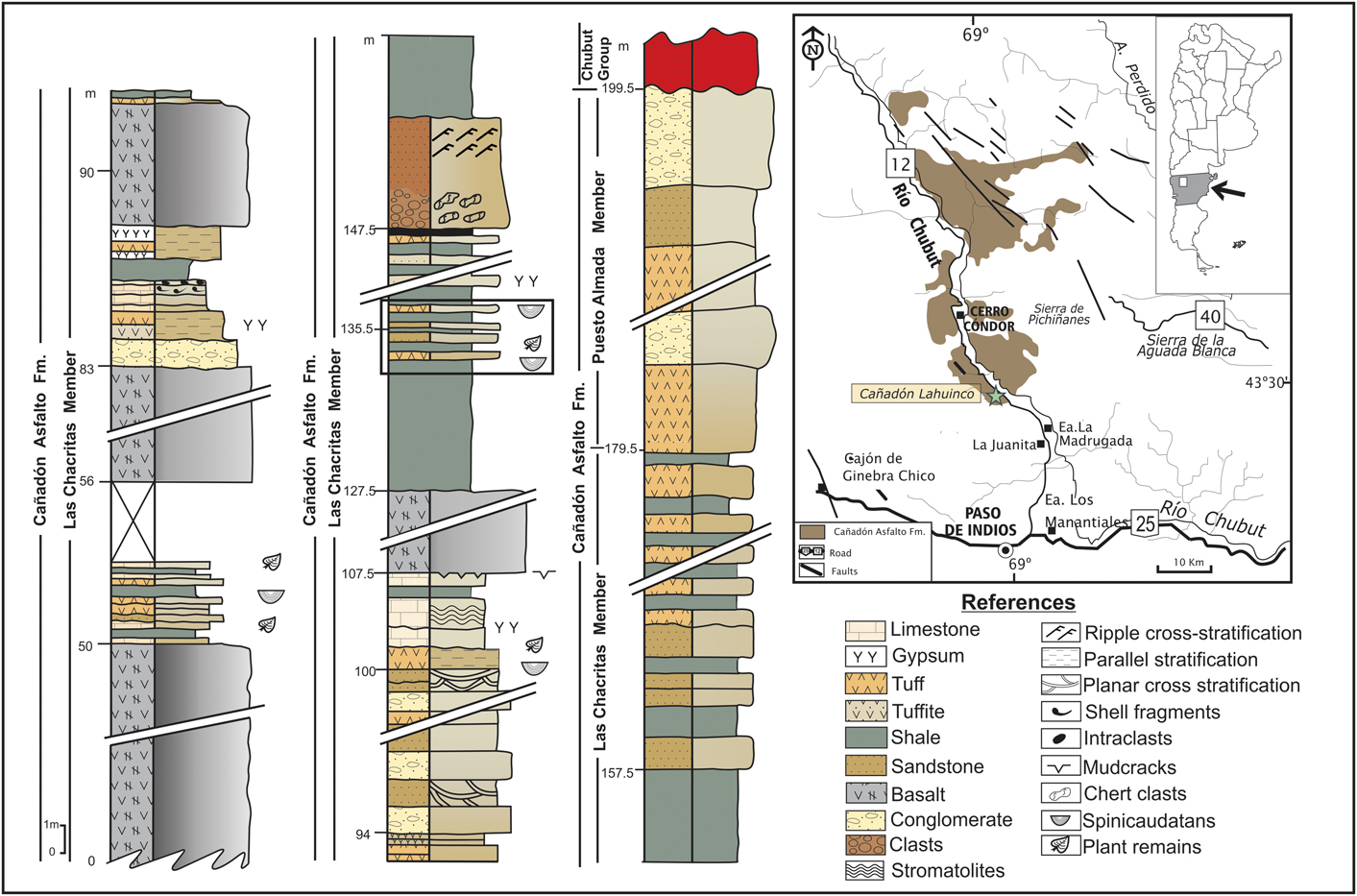
Figure 1 Lahuincó Creek locality: columnar section, both members of the Cañadón Asfalto Formation. Stratigraphic location of the spinicaudatans samples analyzed (square). Modified from Volkheimer et al. (Reference Volkheimer, Quattrocchio, Cabaleri and García2008).
The Cañadón Asfalto Formation is made up of two members, the lower Las Chacritas Member and the upper Puesto Almada Member (Silva Nieto et al., Reference Silva Nieto, Cabaleri and Salani2003; Cabaleri et al., Reference Cabaleri, Volkheimer, Armella, Gallego, Silva Nieto, Cagnoni, Ramos and Panarello2010). The former is represented by carbonate lacustrine facies, massive and stratified limestones, and stromatolitic limestones. The levels are interbedded with pyroclastic deposits (tuff and tuffites), mudstones, shales, sandstones, conglomerates, and basalt flows, dykes, and sills. The sedimentary strata are characterized by the presence of invertebrates, plant remains, and palynomorphs (Cabaleri and Armella, Reference Cabaleri and Armella1999; Volkheimer et al., Reference Volkheimer, Quattrocchio, Cabaleri and García2008; Cabaleri et al., Reference Cabaleri, Volkheimer, Armella, Gallego, Silva Nieto, Cagnoni, Ramos and Panarello2010).
The samples of spinicaudatans originated from the Las Chacritas Member near Lahuincó creek, south of Cerro Cóndor village (Fig. 1). The facies association of this member is represented mainly by carbonate sedimentation in lacustrine systems and river deposits. The sequences are characterized by limestones interbedded with shales, tuffs, tuffaceous levels, sandstones, and conglomerates (Cabaleri et al., Reference Cabaleri, Volkheimer, Armella, Gallego, Silva Nieto, Cagnoni, Ramos and Panarello2010). The spinicaudatans were obtained from the shale lithofacies (Fig. 1). The lacustrine facies of the Las Charitas Member near Lahuincó creek are represented by different environments, ranging from mudflats to palustrine and wetlands (lower and upper). A feature of the lake system is the absence of a defined coastline and shortage of deltaic deposits, indicating that surface water was not the main contribution to the basin (Cabaleri et al., Reference Cabaleri, Volkheimer, Armella, Gallego, Silva Nieto, Cagnoni, Ramos and Panarello2010). The lake sedimentation in the basin is controlled by the climate, as indicated by the expansion and contraction sequences (Gallego et al., Reference Gallego, Cabaleri, Armella, Silva Nieto, Volkheimer, Ballent, Martínez, Monferran and Páez2011; Volkheimer et al., Reference Volkheimer, Quattrocchio, Cabaleri, Narváez, Rosenfeld, Scafati and Melendi2015). During the wet season (expansion episodes), the water entering the basin flooded the littoral areas and favored the sedimentation of carbonate mud, with microbialithic and black shale levels characteristic of marginal environments. The contraction occurs in dry seasons, and these episodes caused water evaporation in marginal areas, forming evaporite deposits (Cabaleri et al., Reference Cabaleri, Armella and Silva Nieto2005). In marginal areas with exposed sediments, paleosols were formed (Volkheimer et al., Reference Volkheimer, Quattrocchio, Cabaleri and García2008). The absence of fluvial inputs in the wetlands suggests that the water supply was mainly underground, rather than superficial, due to the low slope of the basin (Cabaleri et al., Reference Cabaleri, Armella and Silva Nieto2005).
Materials and methods
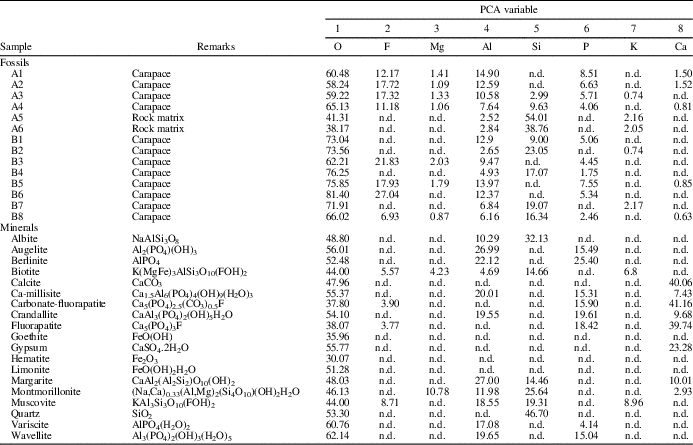
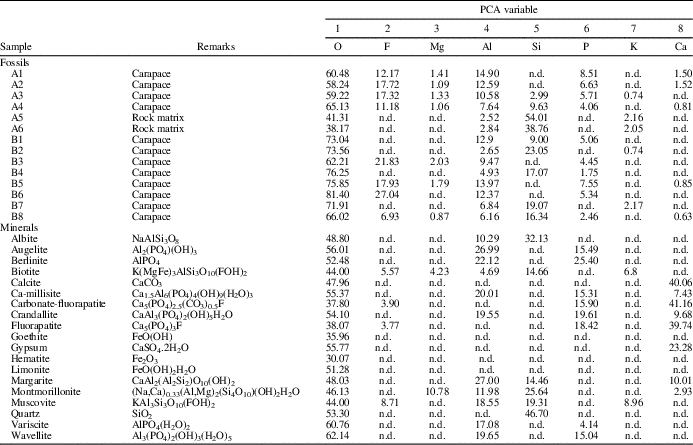
Three types of samples were analyzed: (1) fossil carapaces of Euestheria taschi (Vallati, Reference Vallati1986) (aff. Carapacestheria see Gallego, Reference Gallego2010; Monferran, Reference Monferran2015); (2) rock matrix from the same sample of carapaces; and (3) the following standards of mineral specimens (high purity, 99.5%, i.e., standard reference materials for microanalysis) for comparison with data sets 1 and 2: albite, augelite, berlinite, biotite, calcite, ca-millisite, carbonate-fluorapatite, crandallite, fluorapatite, goethite, gypsum, hematite, limonite, margarite, montmorillonite, muscovite, quartz, variscite, and wavellite (see Table 1).
Table 1 Complete elemental data (O, F, Mg, Al, Si, P, K, and Ca) for 14 samples in different zones analyzed. All measurements are expressed as w/w %. n.d.=not detected. Data set includes (1) fossil carapace (CTES-PZ –7.678); (2) rock matrix; and (3) the following mineral specimens (high purity, 99.5%; included for comparisons purposes): albite, augelite, berlinite, biotite, calcite, ca-millisite, carbonate-fluorapatite, crandallite, fluorapatite, goethite, gypsum, hematite, limonite, margarite, montmorillonite, muscovite, quartz, variscite, and wavellite.
In this study, we applied techniques such as EDS, X-ray diffraction (XRD), and subsequently multivariate analysis that allowed a chemometric approach to the chemical composition of carapace remains. This study falls within the chemometrics issue, which has been defined as “a chemical discipline that uses statistical and mathematical methods, to design or select optimum procedures and experiments, and to provide maximum chemical information by analyzing chemical data.” In short, “chemometrics concerns the extraction of relevant information from chemical data by mathematical and statistical tools” (Varmuza and Filzmoser, Reference Varmuza and Filzmoser2016, p. 16–17).
Scanning electron microscopy and energy-dispersive X-ray spectrometric analysis
Spectrochemical data were obtained using SEM in combination with EDS. The samples were carbon coated before examination. To obtain chemical compositions, energy-dispersive X-ray spectroscopy (EDS-SEM JEOL JSM-6610 LV Thermo Scientific Ultra Dry Noran System 7) was employed. Elements with atomic number greater than 5 were detected by the Sapphire Ultra-Thin Window X-ray detector with a resolution of 129.2 eV for the Mn-Kα line. An acquisition time of 200 seconds, nominal incident beam with an energy of Eo=15 keV, and a 15 mm working distance were used in all cases. The standardless analyses provided semiquantitative results due to the inconsistency in take-off angles between the measured areas and between samples (see Goldstein et al., Reference Goldstein, Newbury, Echlin, Joy, Romig, Lyman, Fiori and Lifshin1992). The PROZA phi-rho-Z matrix correction algorithm (Bastin et al., Reference Bastin, Van Loo and Heijligers1986) was used to perform matrix correction.
EDS measurements must be performed on well-polished specimens so that surface roughness does not affect the results. However, the particular features of the sample carapaces preclude polishing, thus yielding semiquantitative results (see the following section). These results have sufficient precision for multivariate statistical analyses.
Point measurements by EDS were used to monitor trends in the elemental distributions (major and minor components). The locations of measurement points were chosen at preset intervals. However, in some cases, point positions were manually modified to an adjacent area to avoid fossil or rock pore spaces. This process ensures a sufficiently flat surface area for the electron beam. At each point, the weight percentages of the following elements were obtained: O, F, Na, Mg, Al, Si, P, S, K, Ca, Ti, and Fe. The estimated reproducibility was within 10%.
X-ray diffraction analyses
X-ray diffraction (XRD) analyses were carried out using a Bruker D8 Advance X-ray diffractometer at the Center of Technology for Mineral Resources and Ceramics (CETMIC), Buenos Aires, Argentina. The spectra were acquired from a copper anode (Cu-Kα=1.54) operated at 30 kV and 10 mA. The scanning of the samples was carried out from 3° to 70° (2theta) with a step size of 0.04°, a collection time of 2.5 seconds per frame, and fixed divergence slit. The sample carapaces and material from the surrounding rock matrix were pulverized in an agate mortar and mounted on fixed sample holders for further analysis. The analysis of the diffractograms was performed using DIFFRACPLUSEVA software (Bruker AXS).
Semiquantitative determinations
The use of semiquantitative EDS analysis to study fossil materials has been demonstrated to be reliable and has provided reproducible results (e.g., Klug et al., Reference Klug, Schulz and De Baets2009; Lin and Briggs, Reference Lin and Briggs2010; Thomas et al., Reference Thomas, Chinsamy, Conrad and Kandel2012; Previtera et al., Reference Previtera, D’Angelo and Mancuso2013; Benavente et al., Reference Benavente, D’Angelo, Crespo and Mancuso2014). The absolute method (as opposed to a relative method) used here does not employ standards (e.g., Vázquez et al., Reference Vázquez, De Leyt and Riveros1988, Reference Vázquez, De Leyt and Riveros1990; Barrea and Mainardi, Reference Barrea and Mainardi1998; D’Angelo et al., Reference D’Angelo, Perino, Marchevsky and Riveros2002). The samples provide all the experimental information required since the physical-chemical processes involved are well known. The standardless, or absolute, analysis is based on the fundamental parametric method, in which all parameters are derived from the fundamental parameter database (theoretical equations) as well as precise models of the X-ray tube, detector, and geometry. As mentioned, and because of the sample features, obtaining highly polished (smooth) surfaces for a quantitative analysis is not possible. The surface roughness results in inconsistencies in takeoff angles between the measured zones and between samples (Goldstein et al., Reference Goldstein, Newbury, Echlin, Joy, Romig, Lyman, Fiori and Lifshin1992, Reference Goldstein, Newbury, Joy, Lyman, Echlin, Lifshin, Sawyer and Michael2007), yielding semiquantitative results. Thus, semiquantitative elemental concentrations were obtained from the measurements of X-ray fluorescence intensities for each element in the specimen. The characteristic X-ray fluorescence intensity emitted by each element in a sample is recorded and then compared with the corresponding intensity I(j) emitted by a standard of concentration C(j). As a first approximation, the intensity Ij may be considered as proportional to the mass concentration Cj of element j:
 $${\rm Ij\,/\,I}\left( {\rm j} \right){\rm {\equals}Cj\,/\,C}\left( {\rm j} \right)$$
$${\rm Ij\,/\,I}\left( {\rm j} \right){\rm {\equals}Cj\,/\,C}\left( {\rm j} \right)$$
Comparisons with the standards permitted eliminating physical and geometrical factors, which are very difficult to determine (Goldstein et al., Reference Goldstein, Newbury, Echlin, Joy, Romig, Lyman, Fiori and Lifshin1992, Reference Goldstein, Newbury, Joy, Lyman, Echlin, Lifshin, Sawyer and Michael2007). Matrix effects must be taken into account using correction factors (i.e., ZAF). Z and A factors represent generation, scattering, and absorption effects, whereas the F factor involves secondary fluorescence enhancement (e.g., Philibert, Reference Philibert1963). Considering that these effects may differ between the sample and the standard, the ZAF correction is necessary to accurately relate the sample composition with the measured, characteristic X-ray fluorescence intensities (Reed, Reference Reed1993). The magnitudes of the Z, A, and F correction factors are strongly dependent on the experimental conditions, mainly the X-ray energy of the incident beam, takeoff angle, and composition differences of the standards used for comparisons. All of the elements in the sample contribute to the matrix effects, which are related to elemental concentrations. The ZAF terms are calculated from suitable, long-accepted, and well-established physical models. Corrections for matrix effects are routinely carried out by applying the phi-rho-Z matrix correction algorithm (i.e., PROZA; e.g., Bastin et al., Reference Bastin, Van Loo and Heijligers1986; Bastin and Heijligers, Reference Bastin and Heijligers1990).
The results presented here are from semiquantitative determinations of major and minor components by means of EDS. The estimated statistical errors (not shown) in elemental concentrations can be as large 10%. The concentration data were recalculated to 100%.
Multivariate statistical analysis
The semiquantitative data were analyzed by PCA, which is a nonparametric, pattern-recognizing method. The main idea behind this method is to reduce the dimensionality of a data set consisting of a large number of variables while retaining as much of the variance of the original data set as possible. This goal is achieved by transforming the original group of variables to a new set of variables, that is, the principal components (PCs; Hammer et al., Reference Hammer, Harper and Ryan2001). PCs are linear combinations of the original variables, uncorrelated (orthogonal), and are ordered so that the first few components capture most of the variation present in all the original variables (e.g., Jolliffe, Reference Jolliffe2002; Lattin et al., Reference Lattin, Carroll and Green2002; Rencher, Reference Rencher2002; Anderson, Reference Anderson2003; Johnson and Wichern, Reference Johnson and Wichern2007; Izenman, Reference Izenman2008). The main utility of PCA is to detect the formation of groups, systematic separation, or the presence of outliers (Jolliffe, Reference Jolliffe2002). Detecting which variables (or groups of variables) most influence separations in the data is also possible by analyzing the loading plots, that is, coefficients of the linear combinations associated with each PC (Jolliffe, Reference Jolliffe2002).
We retained the number of components with an explained cumulative variance of approximately 80% (Kaiser, Reference Kaiser1960; see Kendall, Reference Kendall1965 for other methods). Our aim was to determine a set of data groupings to evaluate the set as a function of the EDS-derived data (elemental composition). PCA was performed using STATISTICA (StatSoft Inc., 2004) on raw data consisting of eight variables.
Repository and institutional abbreviation
The materials studied here were collected near the Lahuincó creek. This location is on the west side of the Chubut river, near provincial road No. 12 and 12 km south of the Cerro Condor village (43.30572°S, 69.8267°W). The original samples include a fossil association of conchostracans, bivalve mollusks, and ostracods. The material is deposited in the repository of ‘Dr. Rafael Herbst’ Paleontological Collections, Paleozoology section CTES-PZ (Centro de Ecología Aplicada del Litoral, Centro Científico Tecnológico Nordeste, Consejo Nacional de Investigaciones Científicas y Técnicas, Corrientes, Argentina).
Results
The EDS analysis allowed for the discernment of differences in elemental chemical compositions between carapaces of Euestheria taschi Vallati, Reference Vallati1986 (CTES-PZ –7.678) and the surrounding rock matrix. The carapace EDS spectra showed high-intensity peaks of P and Ca, indicating a predominantly calcium phosphate composition (Fig. 2.1, 2.2). By contrast, the rock-matrix-EDS spectra exhibited O and Si peaks, indicating the presence of the major components of silicate compounds. However, minor elements (i.e., Mg, K, and Al ) were also recorded in the carapace samples. The point analyses A1–A4 were taken on an anteroposterior carapace zone, and the points A5 and A6 were assigned to the rock matrix (Fig. 2.2; Table 1). The points B1–B8 correspond to the ventral carapace zone (Fig. 2.3). In all cases, the chemical compositions exhibited differences and similarities among analyzed points.
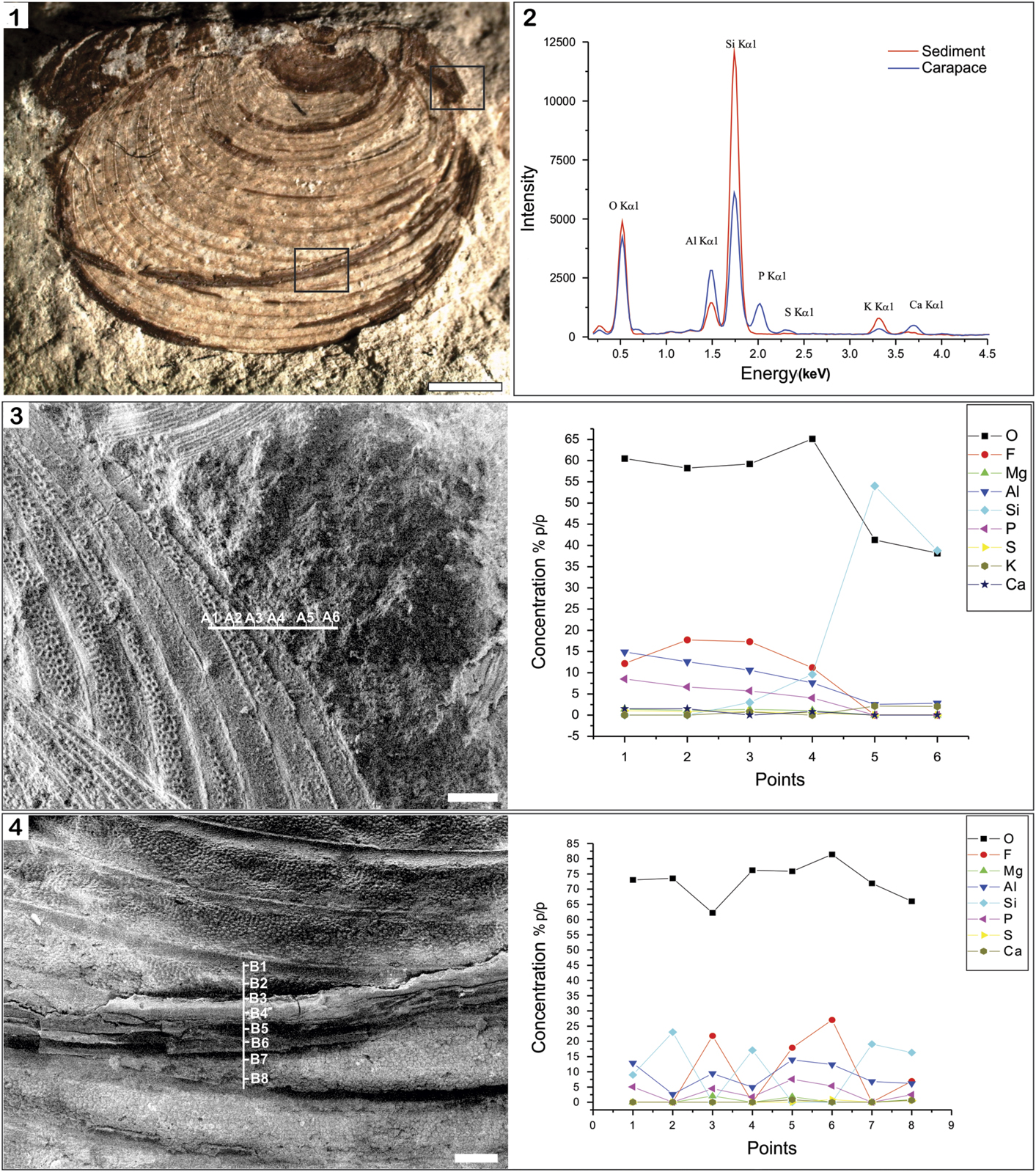
Figure 2 Energy-dispersive X-ray spectrometry (EDS) analysis of an E. taschi carapace (CTES-PZ –7.678) and rock matrix. (1) General view of the carapace; (2) EDS comparison of general EDS spectra of carapace and sediment; (3) points analyzed in the anterodorsal zone—white horizontal line indicates the approximate transect position used for monitoring trends in elemental distributions by EDS; (4) points analyzed in ventral zone—white vertical line indicates the approximate transect position used for elemental analysis by EDS. (1) Scale bar=1 mm; (3, 4) scale bar=100 μm.
Differences in mineral phases were confirmed by XRD analysis, showing that rock matrix surrounding the carapaces predominantly had quartz-cristobalite, K-feldspar, and montmorillonite (Table 2). Furthermore, the diffractogram of carapace samples showed the presence of minerals that were associated directly with the surrounding matrix material (quartz-cristobalite and montmorillonite), the presence of mineral phases such as gypsum and alunite, and other unidentified peaks that can be attributed to phosphate minerals. Unfortunately, the complete elucidation of the diffraction pattern associated with the carapaces was not possible because of: (1) the small amount of material obtained from the carapaces, and (2) the impossibility of obtaining this material without contamination from the surrounding rock matrix. Such limitations were partly overcome by a multivariate evaluation PCA [2], see supplemental data, Appendix 2) of the elemental composition data obtained from the carapace, rock matrix, and mineral specimens (see the following section). These results were compared to the XRD data.
Table 2 Results of XRD analysis.

PCA of fossil samples
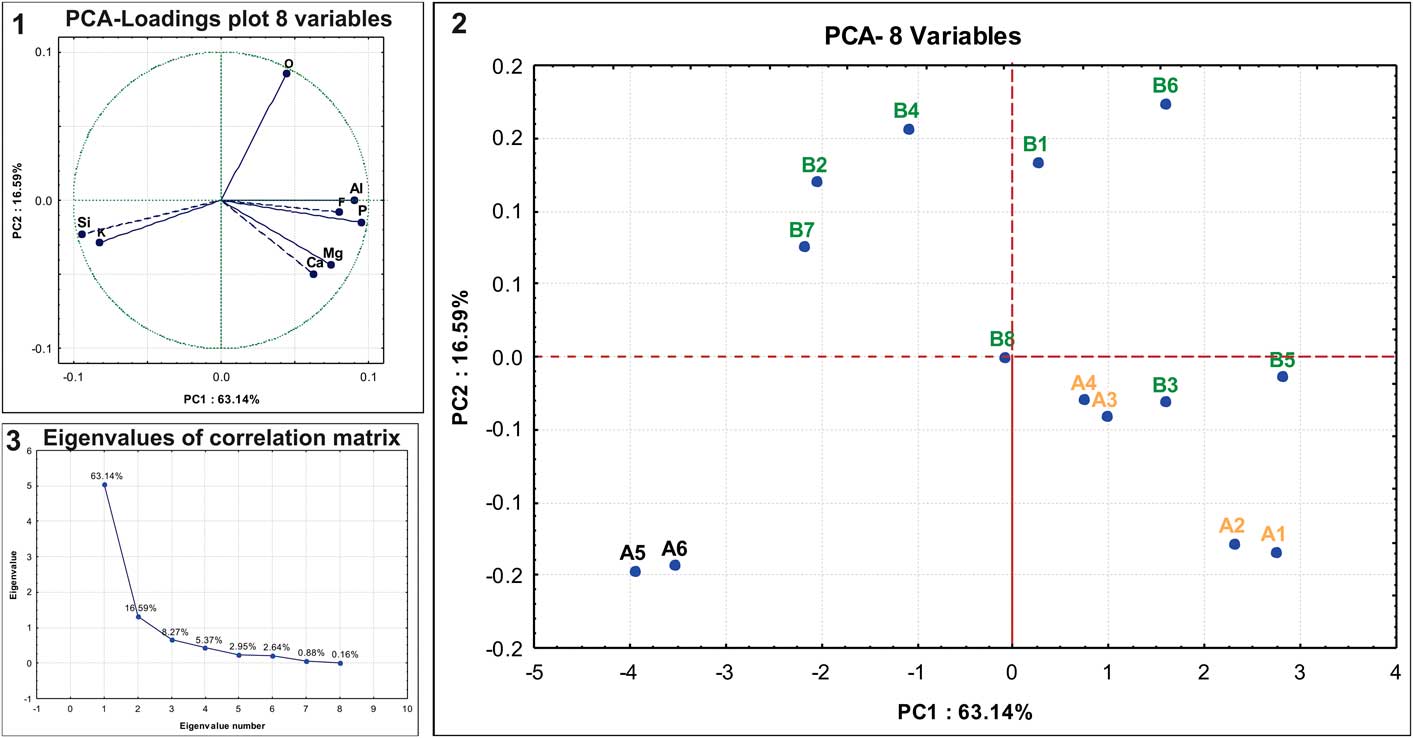
The EDS technique provided 14 data points (Table 1), which were evaluated using PCA (Tables 1–3; supplemental data, Appendix 1). Cumulatively, two components account for 79.73% of the variance (Fig. 3). The first PC (63.14% of variance) exhibits predominantly positive loadings on Al, F, and P, with the exception of Si and K, which emphasizes the relative abundance of silicate minerals (Fig. 3.1). The second PC (16.59% of variance) shows positive values for O and negative values for Ca and Mg. The points measured on rock matrix (A5–A6) exhibited the most negative scores in PC1 (Fig. 3.2), indicating a low content of P, Ca, and F and the importance of compounds containing Si and K, as shown in Table 1.
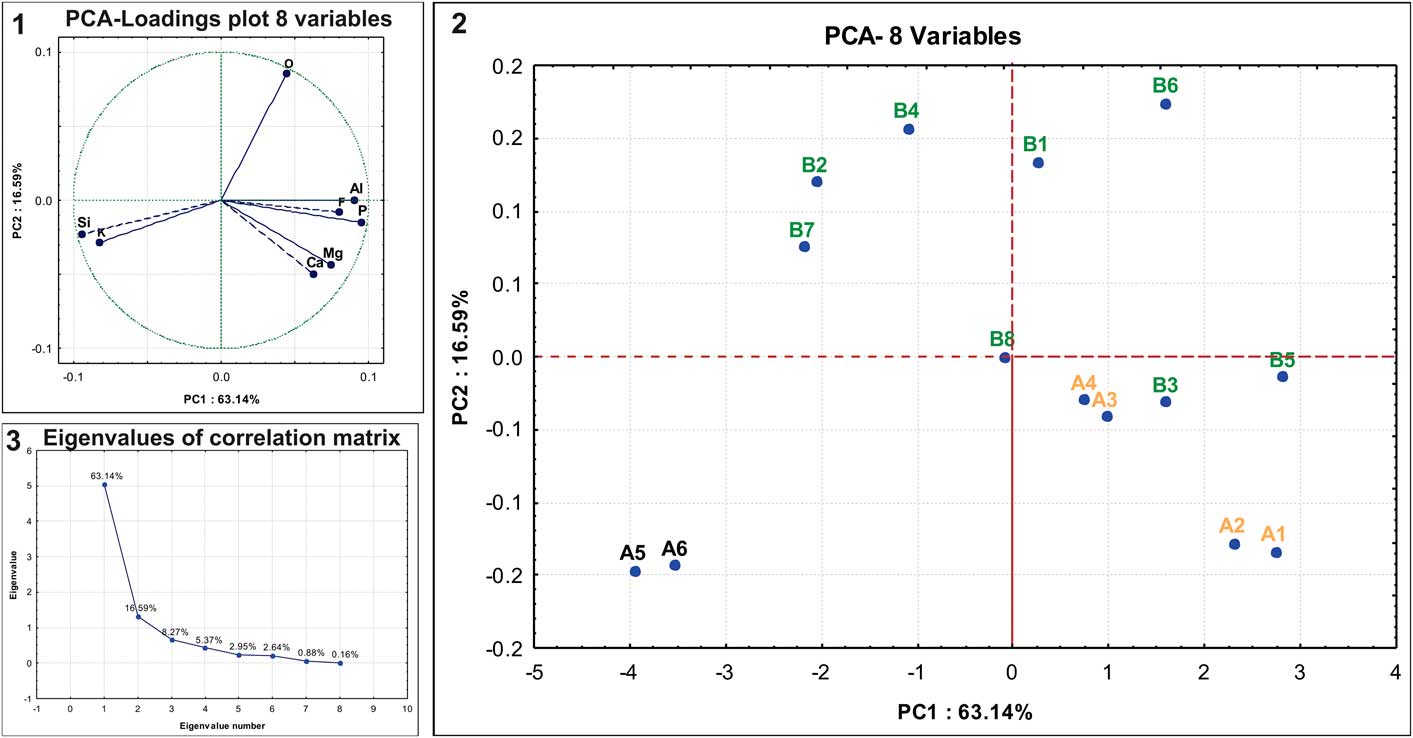
Figure 3 Principal component analysis (PCA) of energy-dispersive X-ray spectrometry (EDS) data obtained from spinicaudatan carapace remains and rock matrix. (1) Plot of PC1 vs. PC2 component loadings; (2) plot of PC1 vs. PC2 component scores; (3) scree plot.
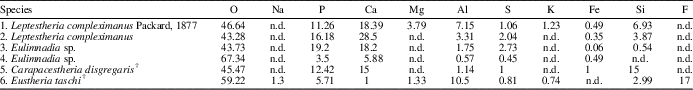
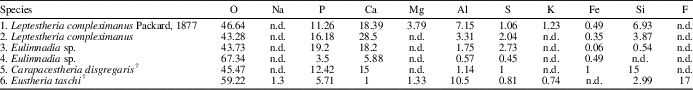
Table 3 Mean values of elemental composition (w/w %) of six individual spinicaudatan carapaces; 1–5 are according to Stigall et al. (Reference Stigall, Babcock, Briggs and Leslie2008), and the species E. taschi was analyzed in this work. n.d.=not detected.
The plot of scores (Fig. 3.2) shows the grouping of data as a function of the chemical compositions from the points analyzed on the spinicaudatan carapace remains. The rock matrix analyzed (A5–A6) exhibited the most negative scores in PC1, indicating the low content of P, Ca, and F and the importance of compounds containing Si and K, as shown in Table 1. The points measured on the carapace remains (A1–A4, B1, B3, B5, B6, and B8) exhibit the most positive scores (PC1), reflecting their high contents of P, F (except B8), Al, and Mg (except for B1 and B6). However, the other points measured (B2, B4, and B7) show a composition mixture between carapace and rock matrix, and some of these points (B2, B4, and B7) have a composition more similar to the rock matrix than to the carapace. These different chemical compositions of the carapace remains could represent the thickness of layers preserved or different conditions of layer preservation.
A second principal component analysis (PCA (2), see supplemental data, Appendix 2) was performed using data points measured on both the carapace remains and selected mineral specimens (high purity, 99.5%, standard reference materials), which were included for multivariate comparison of elemental compositions: albite, augelite, berlinite, biotite, calcite, ca-millisite, carbonate-fluorapatite, crandallite, fluorapatite, goethite, gypsum, hematite, limonite, margarite, montmorillonite, muscovite, quartz, variscite, and wavellite (Table 1).
Results showed that two principal components accounted for 49% of the cumulative variance (see Tables 1–3; supplemental data, Appendix 2). The plots of the component loadings and component scores are shown in supplemental data (Appendix 2). Considering the first three principal components, ~40% of the variance remained unexplained. Therefore, it is important to emphasize that PCA (2) of fossil and mineral samples was intended only as a preliminary analysis, which will be fully developed in future studies. PCA (2) model is a preliminary analysis that indicated a considerable correlation between EDS-derived data measured from carapace and those corresponding to phosphate species (margarite, variscite, ca-millisite, wavellite, augelite, and berlinite). The latter were in agreement with the results obtained by XRD analysis, which unequivocally indicated the presence of phosphate species.
Discussion
Data interpretation
The EDS analysis is a tool for the study of the modes of chemical preservation in spinicaudatan carapaces. Stigall et al. (Reference Stigall, Babcock, Briggs and Leslie2008) analyzed modern spinicaudatan carapaces and revealed a calcium-phosphate composition with minor amounts of Al, Si, S, and Fe (Table 3). Astrop et al. (Reference Astrop, Sahni, Blackledge and Stark2015) analyzed the elemental composition of five living spinicaudatan specimens and showed the presence of C, O, P, and Ca (Na was recorded in only one specimen). These results indicated that the spinicaudatan carapaces are composed of the chitinous component (C, O) of the cuticle and the mineralization of a calcium-phosphate complex (Ca, P). Trace amounts of Zn, S, and other elements also occurred in very low quantities (Astrop et al., Reference Astrop, Sahni, Blackledge and Stark2015). Carapacestheria disgregaris Tasch, Reference Tasch1987 from the Lower Jurassic of Antarctica was also analyzed by EDS, showing the original skeletal mineralogy (Stigall et al., Reference Stigall, Babcock, Briggs and Leslie2008). In our study, the elemental composition (major components) of E. taschi carapace exhibits chemical elements such as F, P, Ca, Al, and Mg. Table 3 shows the similarities and differences in chemical compositions between living and fossil spinicaudatan carapaces. Therefore, our results confirm that the carapace of this species preserved part of the original chemical composition or their trace elements. However, the phosphatic mineralization frequently found in carapaces is present, suggesting some diagenetic recrystallization. This is the case for our studied specimens.
On the other hand, there are differences in chemical composition and concentration between E. taschi and C. disgregaris carapaces. The Ca and P contents are much greater in C. disgregaris carapaces, while other chemical elements are not found or are at low concentrations (e.g., Al, K, F, and Mg; Table 3). These chemical differences are attributed to the variation of the mineralization process for each carapace fossil. The secondary phosphatic mineralization in C. disgregaris carapace remains would have been higher.
The SEM-EDS spectra and PCA analysis of the E. taschi carapace revealed variable chemical compositions according to the measured points (Fig. 3). In specific zones, the carapace remains showed high contents of Ca and P. However, in some other parts of the carapace, the concentrations of Ca and P decreased, while Si contents increased. This finding suggests a different diagenetic pattern for external and internal carapace layers, as in the patterns obtained by Stigall et al. (Reference Stigall, Babcock, Briggs and Leslie2008) in C. disgregaris, where a single valve showed different layers or regions preserved as either calcium phosphate or silica. These authors explained that the silica layers were present where calcium phosphate layers had been exfoliated. Therefore, considering these data, the continuum of variation in the chemical compositions in the carapace remains of E. taschi indicated that different modes of the layers had been preserved, with internal layers preserved as silica and external layers preserved as calcium phosphate (Fig. 4.1). These differences could be due to the different chemical compositions of the layers; however, there are no detailed studies on the composition of layers in living spinicaudatans.

Figure 4 Remains of Euestheria taschi carapace (CTES-PZ –7.678). (1) Diagram of the carapace preservation without obliteration and, on the right, carapace with modifications; (2) some parts of carapace with external layers preservation (in brown) and original ornamentation (reticular in positive relief). ‘Impression mode’ (reticular negative relief) corresponding to internal layers and chemical elements from rock matrix is present (white arrow); (3) in detail. Scale bars=50 μm.
Carapace taphonomy
In many instances, the fragmentation of spinicaudatan carapaces has been interpreted to be a result of biostratinomic modifications caused by hydraulic processes (transport). However, Astrop et al. (Reference Astrop, Sahni, Blackledge and Stark2015) demonstrated that the carapace is resistant to destructive physical taphonomic effects such as abrasion and fragmentation by transport and has high flexibility and resistance in high-energy environments.
Spinicaudatans readily enter the fossil record and seem much more resistant to taphonomic processes than are the soft parts of branchiopod crustaceans. This is evident by the thousands of instances of carapace preservation (Astrop and Hegna, Reference Astrop and Hegna2015). In general, physical breakage by transport in shells or carapaces tends to occur along preexisting lines of weakness such as growth lines or ornamentation such as ribbing (Brett, Reference Brett1990).
The studied specimens of E. taschi lack a breakage pattern, exhibiting isolated (not continuous) patches of carapace in the same specimen (Figs. 2, 4). Therefore, the fragmentation and exfoliation of layers observed on the fossil carapace probably occurred after burial. Stigall et al. (Reference Stigall, Babcock, Briggs and Leslie2008) indicated that the SiO2 replacement of the carapace occurred early in diagenesis, before calcium phosphate recrystallization. Accordingly, mineralic changes lead to the loss of the original properties of the carapaces (e.g., flexibility), making them more susceptible to fragmentation.
Paleoenvironmental influences
The influence of environmental factors (e.g., volcanic activity, sedimentary conditions) on the preservation of fossil invertebrates is poorly understood (e.g., Gupta and Briggs, Reference Gupta and Briggs2011). Clam shrimps have been found in a variety of paleoenvironments, including estuaries, deltas, lakes, and ponds, where neutral to alkaline water conditions and frequent volcanic ash fall prevailed (e.g., Li and Matsuoka, Reference Li and Matsuoka2012; Monferran et al., Reference Monferran, Cabaleri, Gallego, Armella and Cagnoni2016). Volcanic ash contributed to the alkalinity of water through hydration and intrastratal solution of silicic volcanic glass, which is highly active (Hay, Reference Hay1966; Jones et al., Reference Jones, VanDenburgh, Truesdell and Rettig1969; Sheppard and Gude, Reference Sheppard and Gude1969). Paleolakes influenced by volcanic activity make available a high abundance of sulfur compounds (Allison et al., Reference Allison, Maeda, Tuzino and Maeda2008), which oxidize into sulfates in aquatic environments. Furthermore, reactive iron minerals are frequently abundant in sedimentary volcanic deposits (Pan et al., Reference Pan, Sha and Fürsich2014). In addition, alkaline media are indicated by both the preservation of phosphatic minerals in ‘conchostracan’ carapaces (D’Angelo et al., Reference D’Angelo, Monferran, Volkheimer and Cabaleri2013) and cuticle loss in cuticle-free coalified layers of gymnosperm leaves (e.g., Zodrow et al., Reference Zodrow, D’Angelo, Mastalerz and Keefe2009; D’Angelo et al., Reference D’Angelo, Escudero, Volkheimer and Zodrow2011).
In addition, the presence of Ca and aluminum phosphates in lake environments is related to pH conditions and phosphate availability. Wavellite can be formed in alkaline to acidic environments with low phosphate concentrations, while montgomeryite prevails with high phosphate availability (Nriagu, Reference Nriagu1976). Variscite is unstable relative to the other aluminum hydroxyphosphates and is unlikely to be a major constituent in phosphorite beds. Variscite can be a stable phase in natural environments with significant Ca concentrations and where the values for the activities of participating ions are controlled by other reactions within the system (see Nriagu, Reference Nriagu1976). Phosphatic mineralization may occur in alkaline media, even in the case of low P contents (Gierlowski-Kordesch and Kelts, Reference Gierlowski-Kordesch and Kelts2000).
Other studies have suggested the importance of biological factors in the preservation of carapaces involving equilibria between the decay and mineralization of carapace remains. Some factors (e.g., microbial activity) produce decomposition and early mineralization in cuticle layers (Allison, Reference Allison1988; Briggs and Kear, Reference Briggs and Kear1994; Briggs, Reference Briggs2003).
Algae and bacteria have been observed on the carapace surfaces of living spinicaudatans. This suggests that algae and bacteria could have played a role in the preservation of fossil carapaces. Stigall et al. (Reference Stigall, Babcock, Briggs and Leslie2008) indicated that microbially mediated silica molded onto the inner surfaces of carapace layers and typically preserved a higher fidelity of detail than calcium phosphate layers. These authors proposed that silica precipitation is promoted by the chemical gradient set up by microbial degradation of the spinicaudatan carcass. However, further studies could deepen this idea regarding the role of microbial activity in spinicaudatan carapace preservation.
From the Jurassic of Patagonia, spinicaudatans were recorded in lacustrine facies (limestones, mudstones, and shales) associated with mud cracks, stromatolites, and organic matter (Cabaleri et al., Reference Cabaleri, Benavente, Monferran, Narváez, Volkheimer, Gallego and Do Campo2013; Monferran et al., Reference Monferran, Cabaleri, Gallego, Armella and Cagnoni2016). The Lahuincó creek locality is interpreted as an alkaline-water wetland with pyroclastic material levels represented by tuff and tuffaceous rocks (Cabaleri et al., Reference Cabaleri, Volkheimer, Armella, Gallego, Silva Nieto, Cagnoni, Ramos and Panarello2010). The deposits are characterized by clay minerals as well as analcime, kaolinite, smectite, quartz, K-feldspar, plagioclase, and calcite (Cabaleri et al., Reference Cabaleri, Benavente, Monferran, Narváez, Volkheimer, Gallego and Do Campo2013).
The XRD analysis was not useful for the determination of the phosphate mineral species present in the E. taschi carapace. However, PCA (2) (see supplemental data, Appendix 2), including several mineral samples (standard reference materials) and carapace samples, suggests the likely presence of some phosphate species such as augelite, berlinite, Ca-millisite, crandallite, variscite, and wavellite. In addition, results of PCA (2) indicate that carbonate-fluorapatite and fluorapatite would not be present in the carapace. Furthermore, the PCA model (supplemental data, Appendix 2) shows that carapace samples are in close affinity to variscite and wavellite species. The latter suggests that relatively low P and high Ca concentrations were available during the mineralization of carapace remains.
Overall, our experimental evidence, that is, the information derived from the XRD analysis, statistical evaluation of EDS data by PCA (2), and sedimentological and ecological inferences, suggests that E. taschi carapace mineralization occurred in alkaline media with low P and high Ca concentrations and was influenced by periodic ash fall events.
Taxonomic implications
The classification of fossil spinicaudatans relies on carapace features, such as morphology, the presence of macroornamentation (nodes and ribs), and the kinds of microornamentation from growth bands (Chen and Shen, Reference Chen and Shen1985). The latter feature represents one of the most important characteristics in the systematic classification of fossil spinicaudatans. In many cases, the preservation does not allow for the identification of the ornamentation, or the original features could be distorted. Frequently, the carapace’s ornamentations are preserved as the reflection (impression-type fossil) of the original morphology. Therefore, the identification of preserved species is important for proper descriptions in fossil spinicaudatans. Stigall et al. (Reference Stigall, Babcock, Briggs and Leslie2008) indicated that silica layers preserved fine details of the carapace microstructure while the recrystallized phosphate layers did not. Gallego et al. (Reference Gallego, Monferran, Astrop and Zacarías2013) published details on general types of carapaces for Martinsestheria codoensis Cardoso, Reference Cardoso1962, showing lamellae of different thicknesses that make up the carapaces, each with a different kind or presentation of ornamentation or morphological structure (e.g., Gallego et al., Reference Gallego, Monferran, Astrop and Zacarías2013, p. 53, fig. 4).
In our study, the carapace exhibits parts with reticular ornamentation in positive relief that present high phosphorus and calcium concentrations (highlighted picture, Fig. 4.2, 4.3). The same carapace also presents reticular ornamentation in negative relief, corresponding to the internal layer of the carapace, related to the mixture of silicate minerals with phosphorus or calcium (white arrows, Fig. 4.2, 4.3). Therefore, the same fossil carapace could exhibit two modes of ornamentation according to the preservation of different layers, and the carapace exhibits different planar views of the ornamentation. These observations also support the idea that the complete carapace was present during the burial and that some layers of the carapace were exfoliated during later stages of diagenesis or postdiagenesis.
Types of spinicaudatans preserved at the Cañadón Asfalto Formation
Spinicaudatans are abundant in several localities in the Cañadón Asfalto Formation, and their preservation is specific for each locality (Fig. 5). For example, the carapaces are preserved as black remains at Las Chacritas Creek (Fig. 5.1), brown remains at Carrizal Creek (Fig. 5.2), and white remains at Miyanao Creek (Fig. 5.3). However, sometimes only silicified spinicaudatan molds are preserved. This is the case for spinicaudatan carapaces found at Caracoles Creek (Fig. 5.4). Therefore, differences in the carapace color are mainly due to different preservation types. Tasch (Reference Tasch1982) related the preservation color of carapaces to the temperature during fossilization. According to this experimental study, a transparent color is obtained between 300°C and 400°C, a light-brown crust appears in the carapace at 450°C–600°C, a superficial white color forms from 600°C to 1,100°C, and carapaces acquire a transparent glassy characteristic at 1,200°C. The fossil conchostracans analyzed by Tasch (Reference Tasch1982) are from Blizzard Heights and Mauger Nunatak of Antarctica, where the overlying Kirkpartrick basalt flow thermally affected the sedimentary sequences of fossil conchostracans. The Cañadón Asfalto Formation presents a basalt flow between sedimentary sequences, where the temperature from magmatic activity could have affected the preservation (including the colors) of spinicaudatan carapaces. Nevertheless, several complementary studies including X-ray diffraction analysis, Fourier transform infrared analysis, the determination of rock mineral composition, and data on paleoenvironmental inferences are needed to better understand the relationships between carapace color and physical geochemical preservation conditions.

Figure 5 Remains of spinicaudatans carapace showing characteristic colors of different preservation modes; (1) black (Las Chacritas Creek); (2) brown (Karrizal Creek); (3) white (Miyanao Creek); (4) silicified carapace molds (Cañadón Caracoles locality). Scale bars=1 mm.
Conclusion
In this contribution, we present the first chemometric approach to the study of the chemical preservation modes of spinicaudatan carapaces from the Jurassic of Argentina. Carapaces of Euestheria taschi from the Lahuincó creek locality were analyzed using XRD and EDS. Semiquantitative elemental composition data obtained by EDS analysis were subsequently evaluated using a principal component analysis.
The EDS results clearly indicate that spinicaudatan carapaces were not uniformly preserved. These results show complex distribution patterns of elemental compositions in which variable amounts of major constituents (i.e., Ca, P, Al, and F) are detected in different zones of the carapaces. Although the preservation is influenced by diagenetic recrystallization, the recorded values of the elemental compositions are considered part of the original chemical composition of the carapace materials. On the other hand, some other zones of the carapace show that Ca and P contents decrease, while SiO2 (from the underlying rock matrix) contents increase. Therefore, the chemical composition variability found in E. taschi indicates that different carapace layers (with different chemical compositions) were preserved. This preservation is the result of diagenetic modifications of the carapace, while minimal (or no) biostratinomic factors may have been involved.
The ornamentation modes of carapace growth bands are correlated with the preservation modes of their layers. In this case, the original mode is the reticular ornamentation, which is visible in positive relief when the complete carapace morphology (i.e., external layers) is preserved. On the other hand, when incomplete carapace morphological characteristics (i.e., inner layers) are preserved, the reticular ornamentation is seen in negative relief.
Our paleoenvironmental evidence (i.e., sedimentological, ecological, and chemical data) suggest that mineralization of E. taschi carapaces likely occurred in alkaline media with low P and high Ca concentrations and was influenced by periodic ash fall events. The sediments at Lahuincó creek locality are interpreted as wetland paleoenvironments consisting of alkaline brackish water with pyroclastic material levels represented by tuff and tuffaceous rocks. Therefore, ash fall deposits may have played an important role in the fossil preservation at Cañadón Asfalto Formation, while the alkaline media may have favored the preservation of spinicaudatans.
Overall, our results clearly indicate that diagenetic factors played an important role in the preservation modes of the spinicaudatans carapaces.
As we show in this contribution, tracking carapace chemical signatures using chemometric tools opens new possibilities to achieve a better understanding of preservation mechanisms. Future contributions will include studies on the preservation modes of spinicaudatan remains from other Jurassic localities of Argentina.
Acknowledgments
This research was supported by grants PIP 5760 and PIP-112/201001/00034 to NGC from Argentina’s Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET). Thanks to Dr. F. Cravero (Instituto Centro de Tecnología de Recursos Minerales y Cerámica CETMIC-CONICET) for RX diffraction analysis. The Comisión Nacional de Energía Atómica (CNEA) provided logistical support at the Campamento Los Adobes during fieldwork, and G. Giordanengo produced the digital figures.
Accessibility of supplemental data
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.67sf15d.