


INTRODUCTION
Although identified by Antoni van Leeuwenhoek more than 3 centuries ago, Giardia has recently occupied a central stage of parasite research. The epidemiological studies conducted over the past few years indicate that a wide range of mammals, including humans and cattle, are infected by this parasite, causing a substantial burden on the global economy (Giangaspero et al. Reference Giangaspero, Paoletti, Iorio and Traversa2005). Current knowledge supports the proposal that giardiasis is a zoonotic disease and that contaminated water serves as one of the main sources of infection (Monis and Thompson, Reference Monis and Thompson2003; Smith et al. Reference Smith, Reacher, Smerdon, Adak, Nichols and Chalmers2006; Bajer, Reference Bajer2008). Various species of Giardia are recognized (Thompson, Reference Thompson, Ortega-Pierres, Caccio, Fayer, Mank, Smith and Thompson2009), and efforts have been made over the past few years to enhance the taxonomy using molecular tools (Hunter and Thompson, Reference Hunter and Thompson2005; Xiao and Fayer, Reference Xiao and Fayer2008; Thompson, Reference Thompson, Ortega-Pierres, Caccio, Fayer, Mank, Smith and Thompson2009). Based on such tools, 6 species of Giardia have been identified to date, representing 6 different assemblages, of which assemblages A and B infect humans and other mammals (Thompson, Reference Thompson, Ortega-Pierres, Caccio, Fayer, Mank, Smith and Thompson2009).
In humans, Giardia infection can be symptomatic or asymptomatic. Symptomatic giardiasis can present with fatty diarrhoea, abdominal discomfort, vomiting, malabsorption and/or weight loss (Kamda and Singer, Reference Kamda and Singer2009). In some cases, giardiasis resolves rapidly, but in other cases, it can result in chronic infection (Faubert, Reference Faubert2000). Both cell-mediated and humoral immune responses in the host against Giardia have been reported, and adaptive responses have been shown to be critical for controlling giardiasis (Faubert, Reference Faubert2000). Non-immune systems such as secretory immunoglobulins also play a role in the severity of the disease (Nayak et al. Reference Nayak, Ganguly, Walia, Wahi, Kanwar and Mahajan1987).
This parasite has a simple life cycle, with 2 morphological forms – i.e., trophozoites and cysts. Following ingestion, cysts pass through the stomach (being exposed to stomach acid), after which trophozoites are released and colonize the small intestine by longitudinal binary fission (Ghosh et al. Reference Ghosh, Frisardi, Rogers and Samuelson2001). The Giardia trophozoite (12–15 μm long) (Fig. 1, panel A), is non-invasive, and contains a ventral disc made of cytoskeletal proteins that provide support to Giardia for attachment to the enterocyte wall (Holberton, Reference Holberton1973; Ghosh et al. Reference Ghosh, Frisardi, Rogers and Samuelson2001). The resistant cysts (7–10 μm long) with thick cyst walls (Fig. 1, panel B) are responsible for the transmission of giardiasis via contaminated food or water. The cyst wall of Giardia contains insoluble filamentous materials that consist of glycoprotein, glycolipids, and amino-sugar containing oligo- and polysaccharides (Das and Gillin, Reference Das and Gillin1996; Sener et al. Reference Sener, Shen, Newburg and Jarroll2004; Ratner et al. Reference Ratner, Cui, Steffen, Moore, Robbins and Samuelson2008). Three encystation-specific cyst-wall proteins (CWP-1, -2, and -3) are expressed at the time of encystation and are concentrated within encystation-specific vesicles (ESVs) before they are targeted to the cyst wall. Besides these three CWPs, a high-cysteine non-variant cyst protein (HCNCp) is present in trophozoites and may participate in cyst production (Davids et al. Reference Davids, Reiner, Birkeland, Preheim, Cipriano, McArthur and Gillin2006).

Fig. 1. Direct interference contrast (DIC) microscopy pictures of Giardia trophozoite (A) and water-resistant cyst (B) cultured in the laboratory. The trophozoites (12–15 μm long) contain 2 nuclei (not visible in the picture) and a ventral disc (VD) made of cytoskeletal proteins that provide support to Giardia for attachment to the intestinal cell wall. The water-resistant cysts (7–10 μm long) with thick cyst walls (CW) are responsible for the transmission of giardiasis via contaminated water. VD, ventral disc; AF, anterior flagellum; PF, posterior flagellum; CW, cyst wall. Scale bar=5 μm. Reproduced with permission (Hernandez et al. (Reference Hernandez, Shpak, Duarte, Mendez, Maldonado, Roychowdhury, Rodrigues and Das2008)).
Studies conducted in recent years indicate that intestinal lipids and fatty acids influence the growth and encystation of Giardia (Farthing et al. Reference Farthing, Keusch and Carey1985; Gillin et al. Reference Gillin, Reiner, Gault, Douglas, Das, Wunderlich and Sauch1987, Reference Gillin, Reiner and Boucher1988; Lujan et al. Reference Lujan, Mowatt, Byrd and Nash1996). Most lipids are taken up by this parasite from its environment and used as required (Kaneda and Goutsu, Reference Kaneda and Goutsu1988; Mohareb et al. Reference Mohareb, Rogers, Weiner and Bruce1991). However, contrary to the earlier notion that Giardia is unable to synthesize its own lipids de novo (Jarroll et al. Reference Jarroll, Muller, Meyer and Morse1981), results from our laboratory suggest that selective phospholipids can be produced by this parasite via de novo and/or remodelling reactions (Gibson et al. Reference Gibson, Ramirez, Maier, Castillo and Das1999; Das et al. Reference Das, Castillo and Stevens2001, Reference Das, Stevens, Castillo, Villasenor, Arredondo and Reddy2002). The recently established Genome Database (www.giardiadb.org, Morrison et al. Reference Morrison, McArthur, Gillin, Aley, Adam, Olsen, Best, Cande, Chen, Mj, Davids, Dawson, Elmendorf, Hehl, Holder, Huse, Kim, Lasek-Nesselquist, Manning, Niqam, Nixon, Palm, Passamaneck, Prabhu, Reich, Reiner, Samuelson, Svard and Sogin2007), which revealed the presence of lipid synthesis and metabolic genes, further validates our observations. The focus of this article is to review progress in the field of lipid research of Giardia and to validate lipid metabolic pathways by comparison to genomic sequence information. A possible lipid biosynthesis pathway for Giardia has also been proposed.
INTERACTIONS WITH INTESTINAL LIPIDS AND FATTY ACIDS
Because Giardia is continuously exposed to bile acids and dietary fats in the small intestine, it was proposed that lipids and fatty acids play important roles in regulating growth, encystation and excystation. Fatty acids from the intestine kill Giardia in vitro, whereas mucous and bile salts protect the parasite from being killed by fatty acids and other small intestinal factors (Reiner et al. Reference Reiner, Wang and Gillin1986; Das et al. Reference Das, Reiner, Zenian, Hogan, Koss, Wang and Gillin1988). Bile acids were also proposed to facilitate the transport of intestinal lipids into Giardia by forming mixed micelles (Das et al. Reference Das, Schteingart, Hofmann, Reiner, Aley and Gillin1997). The intestinal factors include aggregated and non-aggregated fats, lipases and secretory immunoglobulins (Farthing et al. Reference Farthing, Keusch and Carey1985; Reiner et al. Reference Reiner, Wang and Gillin1986). Free fatty acids generated from phospholipids and triglycerides are detrimental to the growth of Giardia (Reiner et al. Reference Reiner, Wang and Gillin1986; Das et al. Reference Das, Reiner, Zenian, Hogan, Koss, Wang and Gillin1988). Studies suggest that dodecanoic (C12:0) acid (also known as lauric acid) possesses an anti-giardial property at a reasonably low concentration (Rayan et al. Reference Rayan, Stenzel and McDonnell2005). This medium-chain fatty acid accumulates inside trophozoites and alters membrane permeability and integrity. Giardia has the machinery to neutralize the toxic effects of free fatty acids by forming complexes with membrane proteins, lipids, and carbohydrates (Das et al. Reference Das, Traynor-Kaplan, Reiner, Meng and Gillin1991; Gibson et al. Reference Gibson, Ramirez, Maier, Castillo and Das1999; Touz et al. Reference Touz, Conrad and Nash2005).
The role of bile and fatty acids in inducing the encystation of Giardia was first proposed by Frances D. Gillin. In classic experiments, Gillin and her colleagues showed that a mixture of primary bile acid (glycocholate) and fatty acid (oleic acid or myristic acid) promotes in vitro encystation (Gillin et al. Reference Gillin, Reiner, Gault, Douglas, Das, Wunderlich and Sauch1987, Reference Gillin, Reiner and Boucher1988). Subsequently, cholesterol and an excess amount of bovine bile, which Giardia obtains from the growth medium, were shown to induce encystation (Kane et al. Reference Kane, Ward, Keusch and Pereira1991; Lujan et al. Reference Lujan, Mowatt, Byrd and Nash1996). Interestingly, the homologues of sterol regulatory-element-binding proteins (SREBPs) were identified in Giardia and found to regulate the expression of cwp genes during encystation (Worgall et al. Reference Worgall, Davis-Hayman, Magana, Oelkers, Zapata, Juliano, Osborne, Nash and Deckelbaum2004). Giardia expresses 4 genes linked to cholesterol biosynthesis, which are up-regulated during its differentiation into a cyst (Hernandez and Wasserman, Reference Hernandez and Wasserman2006). Several proteins of the parasite can undergo post-translational modification by the intermediate of cholesterol (isoprenyl-group) biosynthetic pathway (Lujan et al. Reference Lujan, Mowatt, Chen and Nash1995), and it is possible that these modifications of giardial proteins are important for maintaining membrane integrity and functions.
IMPORT OF LIPIDS AND FATTY ACIDS BY GIARDIA
Phospholipids and fatty acids are important constituents of all eukaryotic membranes, including those of Giardia. Because of its limited lipid synthesis ability (Das et al. Reference Das, Stevens, Castillo, Villasenor, Arredondo and Reddy2002), lipids in Giardia are acquired from the small intestine of the host, in which the trophozoites are exposed to free and conjugated fatty acids, various sterols, phospholipids and bile acids (Stevens et al. Reference Stevens, Gibson, Adam, Maier, Allison-Ennis and Das1997). Lujan et al. (Reference Lujan, Byrd, Mowatt and Nash1994) showed that lipoprotein-like receptor molecules are present in trophozoites, which allow them to internalize serum-lipoproteins through a cytochalasin-D-sensitive pathway. Using fluorescent lipid analogues, we have shown that trophozoites are able to internalize lipids directly from the culture medium and transport them to various locations, including the plasma and nuclear membranes, the cytoplasm and the endoplasmic reticulum (ER). The cellular localization of each particular lipid analogue studied is distinct. We confirmed that the incorporation of fluorescently-labelled lipid analogues is not dependent on respective fluorophores (i.e. BODIPY or NBD), rather solely on the intrinsic properties (hydrophobicity and hydrophilicity) of lipid probes (Stevens et al. Reference Stevens, Gibson, Adam, Maier, Allison-Ennis and Das1997; Gibson et al. Reference Gibson, Ramirez, Maier, Castillo and Das1999; Das et al. Reference Das, Castillo and Stevens2001). Results indicate that ceramide and phosphatidylglycerol (PG) show preferential localization at perinuclear membranes, whereas phosphatidylcholine (PC) is incorporated into plasma and flagellar membranes. Palmitic acid (PamA) and sphingomyelin (SM) label the nuclear envelopes and the plasma membrane. Phosphatidylethanolamine (PE) is localized to the plasma membrane and in certain cytoplasmic structures adjacent to the plasma membrane.
As cytoskeletal components (i.e. actin filaments and microtubules) are involved in transporting lipid molecules in a range of eukaryotes, we investigated whether the giardial cytoskeleton also participates in the uptake and recycling of fluorescent lipid molecules from plasma- to endo-membranes. In Giardia, microtubules constitute numerous structures in trophozoites, including the ventral disc, basal bodies, flagella, paraflagellar rods and median body (Crossley et al. Reference Crossley, Marshall, Clark and Holberton1986; Elmendorf et al. Reference Elmendorf, Dawson and Mccaffery2003). The basal bodies are the major microtubule organizing centre and functional equivalent of the centrosome of higher eukaryotes (Nohynkova et al. Reference Nohynkova, Draber, Reischig and Kulda2000; Correa et al. Reference Correa, Morgado-Diaz and Benchimol2004; Davids et al. Reference Davids, Williams, Lauwaet, Palanca and Gillin2008). A large set of kinesin homologues are present (Iwabe et al. Reference Iwabe and Miyata2002; Richardson et al. Reference Richardson, Simmons and Reddy2006) but, thus far, no putative homologue of myosin has been identified (Elmendorf et al. Reference Elmendorf, Dawson and Mccaffery2003). Giardia contains a single copy of the actin gene (Morrison et al. Reference Morrison, McArthur, Gillin, Aley, Adam, Olsen, Best, Cande, Chen, Mj, Davids, Dawson, Elmendorf, Hehl, Holder, Huse, Kim, Lasek-Nesselquist, Manning, Niqam, Nixon, Palm, Passamaneck, Prabhu, Reich, Reiner, Samuelson, Svard and Sogin2007), and its protein sequence reveals an ∼58% nucleotide identity to other eukaryotic actin sequences (Elmendorf et al. Reference Elmendorf, Dawson and Mccaffery2003).
We have observed that the uptake and inter-organelle transport of fluorescently labelled SM, PC, and PG are interrupted by anti-actin and anti-microtubule agents. Cytochalasin-D, an actin-depolymerizing drug, induces the formation of several tubular/vesicular structures and blocks the intracellular trafficking of ceramide and SM (Hernandez et al. Reference Hernandez, Castillo, Roychowdhury, Hehl, Aley and Das2007a; Castillo et al. Reference Castillo, Hernandez, Roychowdhury, Das, Ortega-Pierres, Caccio, Fayer, Mank, Smith and Thompson2009). Furthermore, vinorelbine (a microtubule depolymerizing agent) is effective in significantly lowering the intracellular incorporation of fluorescently labelled ceramide and SM. These observations indicate that both ceramide and SM are taken up by cells through cytoskeletal-dependent processes which require intact actin and microtubule structures. On the contrary, the uptake of PC is not dependent on cytoskeleton, because cytochalasin-D and other microtubule-depolymerizing drugs neither alter nor reduce the localization pattern of PC. Like PC, PamA intake is also not affected by anti-cytoskeleton agents (Castillo et al. Reference Castillo, Hernandez, Roychowdhury, Das, Ortega-Pierres, Caccio, Fayer, Mank, Smith and Thompson2009). We have also observed that anti-microtubule depolymerizing agents (e.g., cholchicine, albendazole and nocodazole) blocked the release of PG from the ER/perinuclear regions, suggesting that an intact microtubule structure could be essential not only for uptake and transport, but also for the recycling of PG from the ER to the cytoplasm and plasma membranes (Castillo et al. Reference Castillo, Hernandez, Roychowdhury, Das, Ortega-Pierres, Caccio, Fayer, Mank, Smith and Thompson2009). The results for cytoskeletal-based lipid transport and trafficking experiments (Castillo et al. Reference Castillo, Hernandez, Roychowdhury, Das, Ortega-Pierres, Caccio, Fayer, Mank, Smith and Thompson2009) have been summarized in a model (Fig. 2), which suggests that fluorescently labelled ceramide, SM and PG are mainly taken up by actin-dependent endocytic mechanisms (Hernandez et al. Reference Hernandez, Castillo, Roychowdhury, Hehl, Aley and Das2007a; Castillo et al. Reference Castillo, Hernandez, Roychowdhury, Das, Ortega-Pierres, Caccio, Fayer, Mank, Smith and Thompson2009). It can be postulated that soon after endocytic vesicles are released from the plasma membranes encapsulating lipid molecules, they reach the ER/perinuclear membranes on microtubule rails. Lipids, such as ceramide, SM, PC and PamA, are possibly taken up by the cells through non-endocytic pathways but, at this stage, it is not clear whether PC localized on the outer cell membrane originates from the ER/perinuclear membranes or from the inner plasma membrane (Fig. 2). Giardial lipid and fatty-acid transport proteins may also participate in translocating lipid molecules that may travel along the microtubules to reach perinuclear membranes. The presence of a fatty-acid binding protein (∼8 kDa) has been reported in Giardia (Hassan et al. Reference Hassan, Maache, De La Guardia, Cordova, Garcia, Galiana, Castroviejo, Martins and Osuna2005). The transport of ER/perinuclear PG via exocytic vesicles may be regulated by microtubule filaments and not by actin cytoskeleton (Fig. 2) but more in-depth experiments must be carried out to fully elucidate the lipid transport and trafficking in this organism.
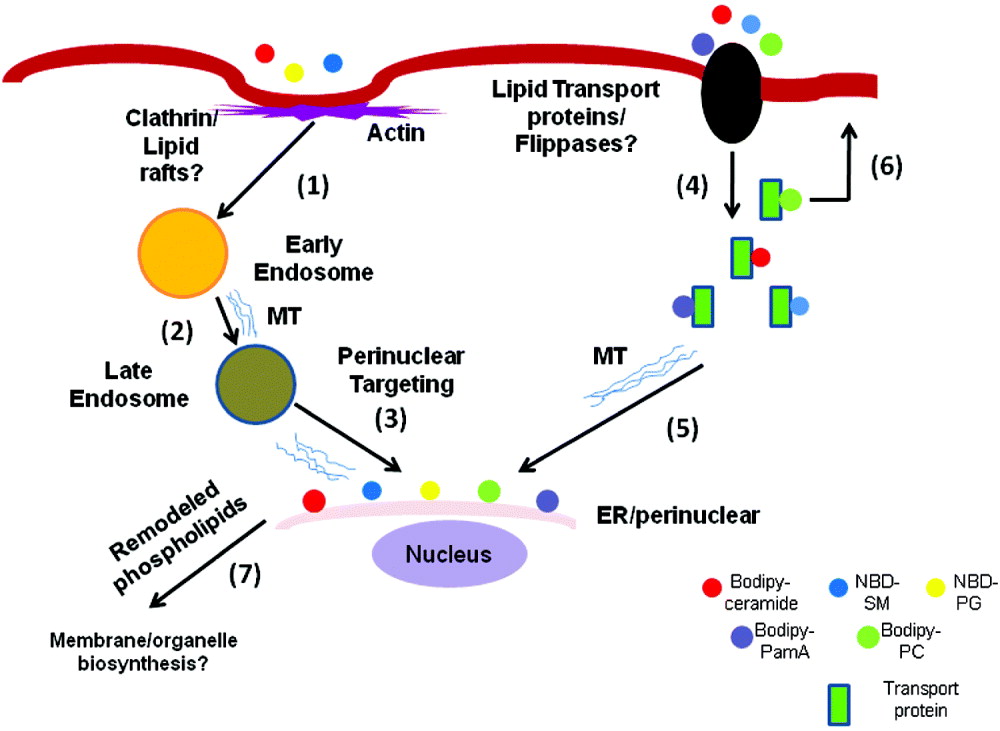
Fig. 2. Lipid import and trafficking by Giardia. The figure shows that BODIPY-ceramide, NBD-SM, and NBD-PG could be imported by actin-dependent endocytic pathways and targeted to ER/perinuclear membranes (steps 1–3) (Hernandez et al. Reference Hernandez, Shpak, Duarte, Mendez, Maldonado, Roychowdhury, Rodrigues and Das2008; Castillo et al. Reference Castillo, Hernandez, Roychowdhury, Das, Ortega-Pierres, Caccio, Fayer, Mank, Smith and Thompson2009). Membrane lipids and fatty acids like ceramide, SM, PC, and PalmA can also be taken up by a flippase-dependent, non-vesicular mechanism and migrate intracellularly. Lipid-binding proteins and microtubule filaments may participate in this process (steps 4–5). Membrane phospholipids like PC, which are mostly localized in the plasma membrane (Das et al. Reference Das, Castillo and Stevens2001), can be flipped back to the plasma membrane (step 6), although the mechanism of this outward movement is not known. It is possible that the internalized lipids are remodelled at the ER/perinuclear regions (step 7) and utilized by the parasite for the synthesis of membranes and organelles. SM, sphingomyelin; PG, phosphatidylglycerol; PalmA, palmitic acid; PC, phosphatidylcholine; MT, microtubule.
SYNTHESES OF NEW LIPIDS AND FATTY ACIDS
The synthesis and metabolism of phospholipids and fatty acids in Giardia was first investigated by Edward Jarroll and his colleagues almost 3 decades ago (Jarroll et al. Reference Jarroll, Muller, Meyer and Morse1981). Using radioactive acetate, glucose, glycerol, threonine, cholesterol and glycerol-3-phosphate, his group monitored the incorporation, utilization and subsequent conversion of these lipids into downstream metabolic products. Interestingly, it was reported that none of these radioactive precursors were converted into other lipids, and it was postulated that Giardia has little or no ability to synthesize lipids de novo. It was thus suggested that Giardia obtains most of its phospholipids and fatty acids from bovine serum and bile supplemented to the growth medium or present in dietary lipids, which are abundant in the human small intestine (Farthing et al. Reference Farthing, Keusch and Carey1985; Gillin et al. Reference Gillin, Gault, Hofmann, Gurantz and Sauch1986). This proposal was further supported by Kaneda and Goutsu (Reference Kaneda and Goutsu1988) and Mohareb et al. (Reference Mohareb, Rogers, Weiner and Bruce1991), who showed that the lipid composition in Giardia is similar to that of the growth medium. Thin-layer chromatographic analyses revealed that 4 phospholipids – i.e. PC, PE, SM, and PG are present in both encysting and non-encysting cells and remain unaltered throughout the process of encystation (Ellis et al. Reference Ellis, Wyder, Jarroll and Kaneshiro1996).
Recently, we carried out detailed analyses of phospholipids in Giardia with the help of electrospray ionization quadrupole time-of-flight mass spectrometry (ESI-qTOF-MS) (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009). The results indicated that PCs and PGs are the major phospholipids in this parasite. Analyses in negative-ion mode revealed that at least 17 different species of PGs are present with various combinations of odd- and even-numbered, carbon-containing fatty acids. Quantitative analyses further elucidated that 2 PG species containing C18:1/C16:0 and C18:1/C16:0 were most abundant, followed by C16:0/C16:0 and/or C18:0/C14:0. Although we detected more species of PCs (19 in positive-ion mode and 6 in negative-ion mode), only 1 of them (C18:1/C18:1) was abundant. In addition to the PGs and PCs, 6 species of PEs, 3 species of SMs and 2 species of PIs were also detected (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009). Interestingly, except for lyso-PCs and PCs, no other phospholipids are present in bile and serum, suggesting that many of these phospholipids (specifically PG and PE) in Giardia could be synthesized de novo via CDP-DAG and/or fatty acid and head-group remodelling pathways (Das et al. Reference Das, Castillo and Stevens2001). This proposal can be further supported by the finding that radio-isotope labelled fatty acids are directly incorporated into membrane phospholipids (Blair and Weller, Reference Blair and Weller1987; Stevens et al. Reference Stevens, Gibson, Adam, Maier, Allison-Ennis and Das1997; Gibson et al. Reference Gibson, Ramirez, Maier, Castillo and Das1999; Vargas-Villarreal et al. Reference Vargas-Villarreal, Escobedo-Guajardo, Mata-Cardenas, Palacios-Corona, Cortes-Gutierrez, Morales-Vallarta, Sampayo-Reyes and Said-Fernandez2007), indicating that Giardia has the cellular machineries to synthesize new phospholipids. Radio-isotope labelled bases (i.e. choline, inositol, ethanolamine, serine and glycerol) are also incorporated into respective phospholipids of trophozoites when added to the culture medium (Subramanian et al. Reference Subramanian, Navarro, Carrasco, Marti and Das2000; Das et al. unpublished observations). A schematic diagram of the synthesis of new phospholipids by fatty acid and headgroup exchange reactions (Das et al. Reference Das, Castillo and Stevens2001, Reference Das, Stevens, Castillo, Villasenor, Arredondo and Reddy2002) is shown in Fig. 3. In the future, it will also be interesting to investigate whether some of the phospholipids in Giardia, particularly PGs, are synthesized via the CDP-DAG de novo pathway.

Fig. 3. Generation of new lipids by fatty acid and headgroup remodelling reactions. (A) Fatty acid remodelling by deacylation/reacylation reaction (the Lands cycle), in which phospholipase A2 and fatty acyl CoA transferase enzymes are involved. (B) Indicates the generation of new lipids by headgroup or base-exchange reactions (Das et al. Reference Das, Castillo and Stevens2001, Reference Das, Stevens, Castillo, Villasenor, Arredondo and Reddy2002).
Several studies suggest that sphingolipid (SL) metabolic pathways are critical for the encystation process, and that inhibition of their syntheses blocks the production of cysts in culture (Hernandez et al. Reference Hernandez, Shpak, Duarte, Mendez, Maldonado, Roychowdhury, Rodrigues and Das2008; Sonda et al. Reference Sonda, Stefanic and Hehl2008; Stefanic et al. Reference Stefanic, Spycher, Morf, Fabrias, Casas, Schraner, Wild, Hehl and Sonda2010). Only 5 SL metabolic genes have been annotated in the Giardia Genomic Database (www.giardiadb.org), and they are all transcribed differentially between trophozoites and encysting cells. These genes are: (i) giardial serine-palmitoyltransferase-1 and -2 subunit genes (gspt-1 and gspt-2), (ii) glucosylceramide synthase or glucosylceramide transferase 1 (gglct-1), and (iii) 2 acid sphingomyelinase genes (gasmase-1 and -2). The enzymatic activities of serine-palmitoyltransferases (gSPTs) and glucosylceramide transferase1 (gGlcT1) were measured and found to be up-regulated during encystation. It was observed that gSPTs (synthesize 3-ketosphinganine – the first rate-limiting step of SL biosynthesis) regulate ceramide endocytosis, which is important because Giardia is unable to synthesize ceramide de novo (Hernandez et al. Reference Hernandez, Shpak, Duarte, Mendez, Maldonado, Roychowdhury, Rodrigues and Das2008). On the other hand, gGlcT1 (catalyses the synthesis of glucosylceramide or GlcCer) is involved in encystation and cyst production by modulating the synthesis of CWPs and ESVs. Inhibition of the synthesis of GlcCer interferes with trophozoite replication and cyst formation (Hernandez et al. Reference Hernandez, Shpak, Duarte, Mendez, Maldonado, Roychowdhury, Rodrigues and Das2008; Sonda et al. Reference Sonda, Stefanic and Hehl2008). Recently, it has been demonstrated that the inhibition of GlcCer production causes cellular abnormalities, including the formation of enlarged lysosomes, clathrin localization and cell-cycle progression before blocking the overall cyst production (Stefanic et al. Reference Stefanic, Spycher, Morf, Fabrias, Casas, Schraner, Wild, Hehl and Sonda2010). Although the function of giardial SMase has yet to be elucidated, it is possible that this enzyme is involved in scavenging ceramide from SM present in the growth medium or in the milieu of the small intestine. These results indicate that ceramide and other SLs play important roles in giardial biology and differentiation.
A comprehensive analysis of fatty acids by Ellis et al. (Reference Ellis, Wyder, Jarroll and Kaneshiro1996) revealed that major fatty acids in Giardia were C16:0 followed by C18:0, C18:1, and C18:2. Small amounts of C14:0, C15:0, C17:0, C18:3, C19:0, C20:0, C22:0, C24:0, C26:0, and C28:0 were also detected. Interestingly, no dramatic differences were observed between the fatty acid content of non-encysting and encysting Giardia. The authors also determined the fatty acid compositions of low-bile (1% bile-containing growth medium) and high-bile (10% bile-containing encystation medium) and noticed that major fatty acids (i.e., C16:0, C18:0 and C18:1) were present in both media, although some quantitative differences were recorded (Ellis et al. Reference Ellis, Wyder, Jarroll and Kaneshiro1996). A detailed analysis of fatty acids by gas chromatography-mass spectrometry (GC-MS) showed that C16:0, C18:0, and C18:1 are indeed the major fatty acids in Giardia and that they remained essentially unaltered during the transition from vegetative forms to encysting (0–48 h) and mature cysts (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009). Traces of shorter-chain fatty acids – i.e. C10:0, C12:0, C14:0, and C15:0 – were also detected. It is interesting that C12:0 and C14:0 were found to be present in the adult bovine serum and bovine bile, the major sources of lipids in giardial growth and encystation medium (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009).
Our results (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009) and those of Ellis et al. (Reference Ellis, Wyder, Jarroll and Kaneshiro1996) further supported the proposal that very long-chain fatty acids (i.e., C20:0, C20:1, C21:0, C22:0, C23:0, C24:0, and C24:1) that are present in Giardia are not taken up from bile and serum and thus could be generated by the action of fatty acid elongase activity, because a similarity (BLAST) search of the giardial genome predicted the presence of fatty-acid elongase 1 gene (gelo) (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009). Free and esterified cholesterols were found to be the major neutral lipids in non-encysting and encysting stages. In addition, the presence of cholesterylesters and small quantities of ergosterol and glycerides were reported (Ellis et al. Reference Ellis, Wyder, Jarroll and Kaneshiro1996). However, the GC-MS analyses (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009) suggest that cholesterol is the only sterol present in trophozoites, encysting cells and cysts, and that it is obtained directly from the growth medium.
LIPID SYNTHESIS AND METABOLIC GENES
Giardia is polyploid, and its genome is very much like the eukaryotic genome that includes linear chromosomes flanked by telomere sequences (TAGGG). The 5 chromosomes, ranging in size from 1·6 to 3·8 Mb, are constituted of 1·34×108 bp, which predicts up to 8–12 copies of each chromosome compared with the haploid genome (Yu et al. Reference Yu, Birky and Adam2002). The trophozoite stage of the parasite has 2 morphologically indistinguishable nuclei that both replicate at approximately the same time and that are transcriptionally active. Each nucleus contains approximately the same copy numbers of ribosomal RNA genes on a single chromosome (chromosome 1), which indicates that this chromosome is present in both nuclei and contains the same complement of DNA (Adam, Reference Adam2001). To understand the biology of the organism as well as to identify new drug targets, the Giardia Genome Project was initiated (www.GiardiaDB.org) in 1998 by Mitchell Sogin and his colleagues at the Marine Biological Laboratory, Woods Hole, MA, with the support from the National Institutes of Health, USA (McArthur et al. Reference McArthur, Morrison, Nixon, Passamaneck, Kim, Hinkle, Crocker, Holder, Farr, Reich, Olsen, Aley, Adam, Gillin and Sogin2000; Morrison et al. Reference Morrison, McArthur, Gillin, Aley, Adam, Olsen, Best, Cande, Chen, Mj, Davids, Dawson, Elmendorf, Hehl, Holder, Huse, Kim, Lasek-Nesselquist, Manning, Niqam, Nixon, Palm, Passamaneck, Prabhu, Reich, Reiner, Samuelson, Svard and Sogin2007). This genome project (Morrison et al. Reference Morrison, McArthur, Gillin, Aley, Adam, Olsen, Best, Cande, Chen, Mj, Davids, Dawson, Elmendorf, Hehl, Holder, Huse, Kim, Lasek-Nesselquist, Manning, Niqam, Nixon, Palm, Passamaneck, Prabhu, Reich, Reiner, Samuelson, Svard and Sogin2007) has assisted in identifying several putative homologues of lipid synthesis and metabolic genes in assemblages A (isolate WB), B (isolate GS) and C (isolate P 15) of Giardia. Table 1 demonstrates the classes of phospholipid syntheses/metabolic genes that were annotated in GenBank (http://www.ncbi.nlm.nih.gov/genbank/). These classes represent the putative genes encoding phosphatidylinositol synthase (PIS), phosphatidylglycerolphosphate synthase (PGPS), phosphatidylserine synthase (PSS) and decarboxylase (PSD). The presence of these genes in the database, together with our earlier lipidomic study that PG, PE, and PI are not obtained from the growth medium (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009), supports the hypothesis that Giardia has the ability to synthesize selective phospholipids de novo. We have observed that Giardia has a strong PSD activity and that it converts [14C]-PS to [14C]-PE instantly (Das et al. unpublished observations). It is likely that Giardia utilizes the product of the gpss gene to synthesize PS from PC and PE, respectively. In mammalian cells, 2 pss genes are present – pss-1 and pss-2. While pss-1 facilitates the formation of PS from PC, pss-2 converts PE to PS (Kent, Reference Kent1995; Dowhan, Reference Dowhan1997). Because Giardia is considered an early diverging eukaryote (Sogin et al. Reference Sogin, Gunderson, Elwood, Alonso and Peattie1989), its pss gene may encode an enzyme that functions both as gPSS1 and gPSS2. Similarly, PG may be synthesized from CDP-DAG in a reaction catalysed by PGPS, encoded by the gpgps gene expressed throughout its life cycle (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009).
Table 1. Lipids and fatty acid metabolic genes annotated in GS, WB and P15 isolates in Giardia

The genome database (Morrison et al. Reference Morrison, McArthur, Gillin, Aley, Adam, Olsen, Best, Cande, Chen, Mj, Davids, Dawson, Elmendorf, Hehl, Holder, Huse, Kim, Lasek-Nesselquist, Manning, Niqam, Nixon, Palm, Passamaneck, Prabhu, Reich, Reiner, Samuelson, Svard and Sogin2007) also suggests the presence of several classes of phospholipid-transport (PLT) ATPases or flippases (FLIPs) that allow the parasite to internalize phospholipids (particularly amino-phospholipids, which include PC and PE) from the environment in the small intestine. As the database suggests, there are several flippase genes in the WB, GS and P15 isolates of Giardia (Table 1). In an unpublished observation, we found that all of these flippase genes in the WB isolate are active and expressed differentially in trophozoites and encysting stages of the parasite's life cycle (K. Y. Aguilera and S. Das, unpublished observations). Although, presently, the reason for the existence of so many flippases is not known, it can be presumed that Giardia has evolved an efficient mechanism to internalize amino-phospholipids, particularly PC, from the intestinal environment.
Several phosphatidylinositol kinase (PIK) – and phosphatidylinositol phosphatase (PIP)-related lipid signalling genes were also annotated in the genome database for Giardia (Table 1), and many of them were shown to be involved in regulating the growth and encystation. An example is the giardial target of rapamycin (TOR), which is an analogue of the FAKB-rapamycin associated protein (FRAP)/TOR of eukaryotes expressed in dividing parasites and is not inhibited by rapamycin (Morrison et al. Reference Morrison, McArthur, Gillin, Aley, Adam, Olsen, Best, Cande, Chen, Mj, Davids, Dawson, Elmendorf, Hehl, Holder, Huse, Kim, Lasek-Nesselquist, Manning, Niqam, Nixon, Palm, Passamaneck, Prabhu, Reich, Reiner, Samuelson, Svard and Sogin2007). The bioinformatic analyses of 3 giardial PIKs genes (gpiks) [2 gpi3ks (gpi3k-1 and gpi3k-2), and 1 gpi4k] were also carried out (Cox et al. Reference Cox, Van Der Giezen, Tarr, Crompton and Tovar2006; Hernandez et al. Reference Hernandez, Zamora, Ray, Chapoy, Chavez, Valvarde, Williams, Aley and Das2007b). The analyses revealed that giardial PI3Ks, unlike higher eukaryotes, contain only catalytic (p110) but not regulatory subunits (p85) (Hernandez et al. Reference Hernandez, Zamora, Ray, Chapoy, Chavez, Valvarde, Williams, Aley and Das2007b). Transcriptional analyses demonstrated that gpiks are expressed in Giardia and are differentially regulated during encystation. In addition, 2 PI3 K inhibitors, wortmannin and LY 294002, have been shown to inhibit the replication of trophozoites in culture, supporting the notion that the activities of PIKs could be linked to the growth and encystation of Giardia (Cox et al. Reference Cox, Van Der Giezen, Tarr, Crompton and Tovar2006; Hernandez et al. Reference Hernandez, Zamora, Ray, Chapoy, Chavez, Valvarde, Williams, Aley and Das2007b). Thus, signal-transducing phospholipid molecules are synthesized in Giardia and participate in cell growth and differentiation.
As shown in Table 1, only 5 SL metabolic genes have been annotated in the Giardia genomic database, including the genes that encode serine-palmitoyltransferases 1 and 2 (gspt-1 and -2) – glucosylceramide transferase or GlcT-1 (gglct-1), and 2 separate acid sphingomyelinase enzymes (gasmases). All 5 genes are reported to be expressed differentially between the 2 different stages of the life cycle of Giardia, suggesting that SL pathways could be involved in modulating the growth and differentiation of this waterborne pathogen (Hernandez et al. Reference Hernandez, Shpak, Duarte, Mendez, Maldonado, Roychowdhury, Rodrigues and Das2008).
With regard to fatty acids (FAs), genomic information for Giardia infers the presence of 9 fatty-acid transport, synthesis and metabolic genes (Table 1). Three 1-acyl-sn-glycerol-3-phosphate acyltransferases (AGPATs) have been annotated, suggesting that Giardia might use the products of these genes to import fatty acids from its surrounding environment. Additional FA genes annotated are putative lysophosphatidic acid acyltransferase (glaat), elongase 1 (gelo), several long-chain fatty-acid (LCFA)-CoA ligases – LCFA-CoA ligase (glcfal), LCFA-CoA ligase 4 (glcfal4), and 3 different forms of LCFA-CoA ligase 5 (glcfal5) – and acetyl-CoA/pyruvate carboxylase (gacpc). The presence of these FA genes further indicates that a very basic but essential FA metabolism is present in Giardia, which is linked to transferring fatty acids across the membranes, forming reactive fatty-acid species (fatty acyl-CoA), acylating lysophosphatidic acid (LPA) to form phosphatidic acid (PA) and elongating and ligating fatty-acid chains (Table 1). Giardia contains 2 isoforms of secreted and cytoplasmic phospholipase B enzymes (gplb) which are responsible for the simultaneous removal of Sn1 and Sn2 fatty acids from a phospholipid (Morgan et al. Reference Morgan, Insall, Haynes and Cockcroft2004).
PROPOSED PATHWAY AND FUTURE PERSPECTIVES
Based on biochemical, cell biology, and genomic information, we have inferred a comprehensive pathway describing the synthesis and metabolism of phospholipids, neutral lipids FAs and SLs in Giardia (Fig. 4). The model reveals that a vibrant and metabolically active trophozoite synthesizes putative gPLTs (or gFLIPs) that allow the parasite to import PC and lyso-PC from the external environment by facilitated diffusion and to convert these molecules into various downstream lipids. For example, PC can be converted to PS by the enzyme encoded by gpss. Table 1 also indicates that both gpsd and gpss are present in Giardia, and that the parasite has the ability to synthesize PS from PE and PE from PS by base-exchange reactions. PG is synthesized de novo, as proposed earlier (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009), and PC may serve as a major precursor. It is likely that a novel PC-to-PG remodelling enzyme may exist and that the parasite uses this enzyme to synthesize PG directly from PC. Nevertheless, such an enzyme has yet to be identified and characterized. The gene gpgps (encoding PGPS) was identified and shown to express in non-encysting and encysting cells (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009). However, it is not known whether this gene participates in the synthesis of new PG via the CDP-DAG (de novo) pathway.

Fig. 4. Proposed lipid metabolic pathways in Giardia. The model proposes that PC and lyso-PC, which are abundant in the growth medium, can be taken up by Giardia with the help of gPLT or gFLIP (step 1). Diacylglycerol (DAG) and FA are internalized by specific transporter(s) from the growth medium (steps 2 and 3). Internalized PC can be converted to PS with the help of gPSS1-like enzyme encoded by putative gpss gene (step 4). Giardia expresses psd gene (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009), and its possible encoded product (gPSD) may facilitate PE synthesis from PS (step 5). The putative gpss can also encode gPSS2-like enzyme for the synthesis of PS from PE (step 6). Because PG is the major phospholipid in Giardia and is not present in the growth medium (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009), it is likely that Giardia has the ability to synthesize PG not only by CDP-DAG pathway (step 16) but also by the headgroup remodelling reaction shown in step 9. Similarly, PI is synthesized from PC by base or headgroup exchange reactions from PG (step 10). However, the presence of these 2 pathways – i.e. PC→PG and PG→PI – is yet to be elucidated in Giardia or other eukaryotic cells. PI can be converted to various phosphinositides facilitated by gpiks (step 11) as mentioned before (Cox et al. Reference Cox, Van Der Giezen, Tarr, Crompton and Tovar2006; Hernandez et al. Reference Hernandez, Zamora, Ray, Chapoy, Chavez, Valvarde, Williams, Aley and Das2007b). Giardial plb and lpl gene may be responsible for synthesizing glycerophosphorylcholine from PC and lyso-PC, respectively (steps 7 and 8). Diacylglycerol (DAG) obtained from the growth medium can be converted to CDP-DAG (step 13) and serves as a precursor for TAG (step 14), PG (step 15), and PI (step 16). Exogenous FAs can produce unsaturated FAs (MUFA and PUFA) and elongated FAs as depicted in steps 18 and 19. Exogenous PalmA can be used as a precursor to synthesize 3-ketosphinganine with the help of gspts (steps 20 and 21). Similarly, both ceramide and SM can be acquired from the growth medium (step 22) by endocytic and non-endocytic pathways. Exogenously obtained SM can be hydrolysed by gsmases to produce ceramide (step 23), and ceramide can be used to synthesize GlcCer (step 24). PC, phosphatidylcholine; lyso-PC, lyso-phosphatidylcholine; PS, phosphatidylserine; PE, phosphatidylethanolamine; PIP, phosphatidylinositol phosphate; PIP2, phosphatidylinositol bisphosphate; PIP3, phosphatidylinositol triphosphate; DAG, diacylglycerol; TAG, triacylglycerol; CTP, cytidine triphosphate; FA, fatty acid; MUFA, monounsaturated fatty acid; PUFA, polyunsaturated fatty acid; SM, sphingomyelin; GlcCer, glucosylceramide. For better understanding, phospholipid pathways are shown in red, neutral lipids (DAG and TAG) in black, FAs in green, and SLs in pink.
As shown in Table 1, and also reported earlier (Morgan et al. Reference Morgan, Insall, Haynes and Cockcroft2004), Giardia has the genes that encode phospholipase B (PLB). The hydrolysis of Sn 1 and Sn 2 FAs from PC by PLB produces lyso-PC and soluble glycerophosphorylcholine. The presence of lyso-phosphatidic acid acyltransferase (LPAAT) gene (glpaat) in the genomic database suggests that this parasite also has the ability to convert lyso-PA to PA.
The pathway also proposes that most FAs can be taken up by simple and facilitated diffusion (Gibson et al. Reference Gibson, Ramirez, Maier, Castillo and Das1999). Once internalized, FAs undergo elongation and/or desaturation reactions. The presence of a giardial FA desaturase was reported earlier by Ellis et al. (Reference Ellis, Wyder, Jarroll and Kaneshiro1996); the gene (gelo) that is likely to encode elongase was annotated in the database (Table 1). Diacylglycerol (DAG) or other neutral lipids present in the medium (Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009) can also be obtained by membrane diffusion and/or via transport proteins. Intracellular DAG can form triacylglycerol (TAG) by agpat gene products, and diacylglycerol (DAG) can be activated by cytidine diphosphate (CDP) to produce CDP-DAG, which then can be used as a precursor to synthesize PI. Newly synthesized PI can be utilized to generate PIP, PIP2, and PIP3 by giardial PIKs for cellular signalling (Cox et al. Reference Cox, Van Der Giezen, Tarr, Crompton and Tovar2006; Hernandez et al. Reference Hernandez, Zamora, Ray, Chapoy, Chavez, Valvarde, Williams, Aley and Das2007b).
As mentioned, Giardia expresses gspt, gglct1, and gsmase genes, indicating a limited SL synthesis/metabolic pathway. It is possible that PalmA obtained from the growth medium is converted to Palm-CoA by acyl-CoA ligase and then is used by the parasite to synthesize 3-ketosphinganine by the action of serine-palmitoyltransferase enzymes encoded by gspts. We have proposed earlier (Hernandez et al. Reference Hernandez, Shpak, Duarte, Mendez, Maldonado, Roychowdhury, Rodrigues and Das2008) that 3-ketosphinganine regulates ceramide uptake in Giardia by controlling its endocytic machinery, and this is important because ceramide is not synthesized by this parasite de novo. The newly acquired ceramide is then used by Giardia as precursors to synthesize GlcCer by glucosylceramide synthase (encoded by gglct1) which may serve as a key regulator of encystation and cyst production (Hernandez et al. Reference Hernandez, Shpak, Duarte, Mendez, Maldonado, Roychowdhury, Rodrigues and Das2008). Taken together, it has been proposed that ceramide uptake and GlcCer synthesis is important for the encystation of Giardia (Hernandez et al. Reference Hernandez, Shpak, Duarte, Mendez, Maldonado, Roychowdhury, Rodrigues and Das2008; Sonda et al. Reference Sonda, Stefanic and Hehl2008; Stefanic et al. Reference Stefanic, Spycher, Morf, Fabrias, Casas, Schraner, Wild, Hehl and Sonda2010).
We foresee that the current review not only contributes to our understanding of the lipid pathways in Giardia but also should assist researchers in identifying unique targets for developing effective therapies in the future. One of these targets might be the enzymes of PG biosynthesis, because PG appears to be the major phospholipid in Giardia (Gibson et al. Reference Gibson, Ramirez, Maier, Castillo and Das1999; Yichoy et al. Reference Yichoy, Nakayasu, Shpak, Aguilar, Aley, Almeida and Das2009). Lipid transport and lipid-based cell signalling could be another important area for future investigation. As mentioned above, Giardia has evolved mechanisms to import exogenous lipids and cholesterol by receptor-mediated endocytosis (Lujan et al. Reference Lujan, Byrd, Mowatt and Nash1994) and traffic via clathrin-mediated and actin/microtubule-dependent pathways (Hernandez et al. Reference Hernandez, Castillo, Roychowdhury, Hehl, Aley and Das2007a). Therefore, the identification of lipoprotein-like receptors and the study of lipid transport vesicles in Giardia should open a new research area that might lead to the discovery of unique pathways and mechanisms of lipid sorting and targeting. At present, it is not fully understood how extracellular signals regulate the growth and differentiation of Giardia. Future investigation may suggest that PI3 K-based signalling is associated with this phenomenon and drives the process of encystation and excystation. Finally, it would also be fascinating to investigate whether giardial lipids and lipid metabolic enzymes are involved in host-parasite interactions.
FINANCIAL SUPPORT
The authors were supported by the National Institutes of Health (S.D., grant no. S06GM008012). Mass spectrometric analyses of lipids were carried out (in collaboration with Professor Igor C. Almeida) in the Biomolecule Analysis Core Facility at BBRC/UTEP supported by the NIH/NCRR/RCMI (grant no. 5G12RR008124). Dr Mayte Yichoy was supported in part by the Dodson Dissertation Fellowship from UT-El Paso. Mr Trevor T. Duarte, Mr Tavis L. Mendez and Ms Kristina Y. Aguilera were supported by the RISE and MARC (NIH) programs, respectively. Mr Debarshi Roy was supported by a grant from the Howard Hughes Medical Institute.





