


1. Introduction
In the process of word production, a speaker formulates a concept, selects the corresponding lexical item, encodes its phonology, and produces it via articulatory organs (Levelt, Roelofs & Meyer, Reference Levelt, Roelofs and Meyer1999). This process is more complicated in bilinguals whose two languages share the same conceptual representation, but use two separate lexical representations (Kroll & Stewart, Reference Kroll and Stewart1994). In addition, as indicated by accumulative evidence (Colomé, Reference Colomé2001; Costa & Caramazza, Reference Costa and Caramazza1999; Costa, Caramazza & Sebastian-Galles, Reference Costa, Caramazza and Sebastian-Galles2000; Costa, Miozzo & Caramazza, Reference Costa, Miozzo and Caramazza1999; Fink & Goldrick, Reference Fink and Goldrick2015; Giezen & Emmorey, Reference Giezen and Emmorey2016; Guo & Peng, Reference Guo and Peng2006; Hoshino & Kroll, Reference Hoshino and Kroll2008; Poarch & van Hell, Reference Poarch and van Hell2012; Starreveld, De Groot, Rossmark & van Hell, Reference Starreveld, De Groot, Rossmark and van Hell2014), when bilinguals speak one language, the representation of the other language is also activated in parallel. Thus, during the word production, bilinguals need a lexical selection mechanism to guarantee selecting the target lexical item in the target language.
Two main prevailing accounts have been put forward to address this issue: the language specific selection hypothesis and the language non-specific selection hypothesis. According to the language specific selection hypothesis, although the bilingual's two languages are both activated, only the lexical items in the target language are considered for lexical selection (Costa et al., Reference Costa, Miozzo and Caramazza1999). In contrast, the language non-specific hypothesis holds that lexical items in both languages compete for selection. Bilinguals need to inhibit the lexical item in the non-target language (e.g., Green, Reference Green1998) to guarantee the one that corresponds to the concept in the intended language finally gets selected because of its relatively higher degree of activation. Of note, it has been postulated that the lexical selection mechanism might vary as a function of the proficiency of the second language (L2). Specifically, L2 learners with relatively low proficiency level of L2 rely on a language non-specific selection mechanism, whereas highly proficient bilinguals might develop a language specific selection mechanism (Costa & Santesteban, Reference Costa and Santesteban2004; Costa, Santesteban & Ivanova, Reference Costa, Santesteban and Ivanova2006). The current study examines the lexical selection mechanism in unbalanced bilinguals in particular.
One influential language non-specific selection model is the Inhibitory Control Model (IC Model, Green, Reference Green1998). This model assumes that to guarantee greater activation of the lexical item in the target language, language control is executed during two major phases. In the first phase, the target language task schema and the non-target language task schema compete with each other. Typically, the target language task schema wins and acquires a relatively higher activation than the non-target language task schema. In the second phase, the target language task schema selectively inhibits the activation of the lexical items in the non-target language. Additionally, the IC model holds that the inhibition exerted on the non-target language lexical item is reactive. The extent of inhibition depends on the proficiency of the non-target language. The stronger language is typically activated greater than the weaker language, thus it needs more inhibition as the non-target language.
Support of the IC model has been found in studies using the language switching task. In this task, bilinguals are asked to switch between languages while naming pictures or digits. There are two types of trials: switch trials and non-switch trials. In switch trials, the naming language of the current trial differs from that in the previous trial. In contrast, in non-switch trials, two consecutive trials are named in the same language. The difference in response times (RTs) and error rates (ERs) between the switch and non-switch condition is referred to as switching costs. For example, switching costs in L1 refer to costs from switching into L1, namely the difference between L1 switch condition (L2-L1) and L1 non-switch condition (L1-L1). The IC model offers a ready explanation for switching costs (for other explanations see recent reviews, Baus, Branzi & Costa, Reference Baus, Branzi, Costa and Schwieter2015; Bobb & Wodniecka, Reference Bobb and Wodniecka2013; Declerck & Philipp, Reference Declerck and Philipp2015). In switch trials, the target language in the current trial needs to recover from its previous inhibition as a non-target language, leading to longer RTs and higher ERs. Furthermore, the IC model predicts that the switching costs between two unbalanced languages should be asymmetric, based on its assumption that the extent of inhibition relies on the proficiency level of the non-target language. In unbalanced bilinguals, the switching costs in the stronger language should be larger than those in the weaker language. In balanced bilinguals, in contrast, the switching costs should be comparable in the two directions. Indeed, an asymmetrical pattern was reported in a number of studies examining unbalanced bilinguals or trilinguals (e.g., Costa & Santesteban, Reference Costa and Santesteban2004, Experiment 1; Fink & Goldrick, Reference Fink and Goldrick2015, Experiment 2; Linck, Schwieter & Sunderman, Reference Linck, Schwieter and Sunderman2012; Meuter & Allport, Reference Meuter and Allport1999; Philipp, Gade & Koch, Reference Philipp, Gade and Koch2007, Experiment 1). In addition, a symmetrical pattern was observed in highly proficient bilinguals or trilinguals (e.g., Calabria, Hernández, Branzi & Costa, published online January 13, 2012; Costa & Santesteban, Reference Costa and Santesteban2004, Experiments 2, 3 & 4; Costa et al., Reference Costa, Santesteban and Ivanova2006, Experiments 1 & 2; Declerck, Thoma, Koch & Philipp, Reference Declerck, Thoma, Koch and Philipp2015b).
Additional support for the IC model is obtained from the n-2 repetition paradigm, which examines trilinguals’ lexical production performance (Declerck et al., Reference Declerck, Thoma, Koch and Philipp2015b; Guo, Liu, Chen & Li, Reference Guo, Liu, Chen and Li2013a; Philipp et al., Reference Philipp, Gade and Koch2007; Philipp & Koch, Reference Philipp and Koch2009). Identical to the language switching task, this task asks the participants to name pictures or digits based on cues for the naming language. There are two critical conditions: the n-2 repetition condition and the n-2 non-repetition condition. In the n-2 repetition condition (e.g., naming in the sequence of L1-L2-L1), the naming language of Trial n is the same as that of Trial n-2 (i.e., the trial ahead of Trial n-1, which precedes Trial n). In the n-2 non-repetition condition (e.g., naming in the sequence of L3-L2-L1), the naming languages of Trial n and Trial n-2 differ. The performance difference between those two conditions is defined as the n-2 repetition cost, which has been taken as a marker of persisting inhibition. The logic is that if the language of Trial n-2 (e.g., L1) is inhibited in Trial n-1 (e.g., L2), it requires efforts to recover from this inhibition in Trial n (e.g., L1), where it serves as the target language again. Indeed, n-2 repetition costs were observed in multiple studies examining trilinguals (Guo et al., Reference Guo, Liu, Chen and Li2013a; Philipp et al., Reference Philipp, Gade and Koch2007, Experiment 2; Philipp & Koch, Reference Philipp and Koch2009). For example, Declerck et al. (Reference Declerck, Thoma, Koch and Philipp2015b) tested Turkish–German–English trilinguals who were highly proficient in German and relatively less proficient in English. Participants were asked to name digits according to the geometric shapes cuing the naming languages. Results showed that n-2 repetition costs were present in all three languages. In addition, the costs in the weaker English were smaller than those in the stronger Turkish and German. In support of the IC model, these findings suggest that inhibition is implemented during word production in trilinguals who are highly proficient in their L2. Besides, the extent of inhibition exerted on a language is determined on its proficiency level, such that the less proficient L3 requires less inhibition relative to the proficient L1 and L2.
More recently, the upsurge of event-related potential (ERP) research has provided valuable insights on the time course of language control during bilingual word production, due to the high temporal resolution of the technique. A key ERP component, the N2, has been particularly informative. The N2 is a negative-going waveform peaking around 200–350 ms after the onset of the stimulus over the frontal-central scalp (for a review, see Folstein & Van Petten, Reference Folstein and Van Petten2008). It has been associated with cognitive control of interference (Nieuwenhuis, Yeung & Cohen, Reference Nieuwenhuis, Yeung and Cohen2004), such as response inhibition (Jackson, Jackson & Roberts, Reference Jackson, Jackson and Roberts1999; Schmitt, Münte & Kutas, Reference Schmitt, Münte and Kutas2000; Thorpe, Fize & Marlot, Reference Thorpe, Fize and Marlot1996) and response conflict monitoring (e.g., Nieuwenhuis, Yeung, Van Den Wildenberg & Ridderinkhof, Reference Nieuwenhuis, Yeung, Van Den Wildenberg and Ridderinkhof2003). Previous ERP studies on bilingual word production have mainly examined the N2 component as an index of language control. However, the findings available are inconsistent across studies. Jackson, Swainson, Cunnington and Jackson (Reference Jackson, Swainson, Cunnington and Jackson2001) conducted the first ERP study to investigate bilingual lexical selection mechanism. They asked unbalanced bilinguals to perform a cued language switching task. ERP data showed that switch trials elicited a more negative N2 component than non-switch trials over the frontal electrodes. In addition, the frontal N2 deflection was significant when switching into L2, but not in the other direction. Jackson and colleagues interpreted these patterns in support of the IC model: when switching into the L2, the dominant L1 needs to be strongly inhibited; thus, a robust N2 effect in the L2 was observed. Interestingly, Christoffels, Firk and Schiller (Reference Christoffels, Firk and Schiller2007) reported a reversed N2 effect, i.e., the N2 amplitude was more enlarged for non-switch trials compared with switch trials. Additionally, this modulation was present when switching into L1, but not when switching into L2. One interpretation Christoffels and colleagues proposed was that the reversed N2 effect reflects more conflict processing for L1 non-switch trials. Specifically, unbalanced bilinguals were biased toward their L2 when they were required to switch between languages. Such a bias would create more conflict for L1 non-switch trials, the waveform of which exhibited a larger N2 magnitude. In contrast, Verhoef, Roelofs and Chwilla (Reference Verhoef, Roelofs and Chwilla2009) did not find reliable N2 modulation in either direction when the cue was presented ahead of the picture to the unbalanced Dutch–English bilinguals.
Noteworthy, these studies examined stimulus-locked ERPs in the cued language switching task, which could be associated with inhibition at the lemma level. Cue-locked ERPs, on the other hand, could be related to language control at the language schema level, as the target language schema competition starts upon cue presentation (see Guo, Ma & Liu, Reference Guo, Ma and Liu2013b, for related discussion). Verhoef, Roelofs and Chwilla (Reference Verhoef, Roelofs and Chwilla2010) examined cue-locked ERPs in the cued language switching task. Results showed that in the time window of 350–500 ms post-cue onset, more negative going waveforms were elicited by switch trials compared to non-switch trials in both L1 and L2 over the frontal electrodes. These later deflections were associated with engaging the target language. Nevertheless, an alternative interpretation under the framework of the IC model would be that language control occurred during language schema competition phase (see also Guo et al., Reference Guo, Ma and Liu2013b).
As shown above, the ERP evidence from the language switching task is mixed. Guo et al. (Reference Guo, Ma and Liu2013b) further examined the locus of inhibitory control during trilingual word production with the n-2 repetition task. Cue-locked ERPs showed that the N2 modulation (more negative N2 elicited by the n-2 non-repeat trials) was small or marginal over a few electrodes in the mid-line. On the other hand, stimulus-locked ERPs showed a robust N2 effect (more negative N2 elicited by the n-2 repeat trials) over the majority of sites across the scalp. One interpretation of these findings is that inhibitory control is absent during the language schema competition phase. Rather, it occurs at the lemma selection phase, where the target language schema inhibits the lexical items in the non-target language.
As shown by studies reviewed above, a great deal of evidence suggests that unbalanced bilinguals rely on the inhibitory control mechanism for lexical selection during word production (e.g., Declerck et al., Reference Declerck, Thoma, Koch and Philipp2015b; Jackson et al., Reference Jackson, Swainson, Cunnington and Jackson2001; Meuter & Allport, Reference Meuter and Allport1999; Philipp et al., Reference Philipp, Gade and Koch2007; but see Finkbeiner, Almeida, Janssen & Caramazza, Reference Finkbeiner, Almeida, Janssen and Caramazza2006).
Although a number of studies examined the lexical selection mechanism in different bilingual populations, exceptionally few have investigated the plasticity of the mechanism. As mentioned earlier, Costa and colleagues (Costa & Santesteban, Reference Costa and Santesteban2004; Costa et al., Reference Costa, Santesteban and Ivanova2006) proposed a hypothesis on the developmental trajectory of the bilingual lexical selection mechanism. Specifically, less proficient L2 learners rely more on the inhibitory control mechanism for lexical selection, whereas highly proficient bilinguals depend more on a language-specific selection mechanism. This hypothesis was introduced based on the findings of symmetrical switching costs between languages varying in proficiency (e.g., Costa & Santesteban, Reference Costa and Santesteban2004; Costa et al., Reference Costa, Santesteban and Ivanova2006). For example, Costa and Santesteban (Reference Costa and Santesteban2004, Experiment 4) tested trilinguals who were highly proficient in their L1 and L2 and less proficient in their L3. They observed symmetrical switching costs between the L1 and the L3 (but see Declerck et al., Reference Declerck, Thoma, Koch and Philipp2015b, where asymmetric n-2 repetition costs between the highly proficient L1 (and the L2) and the less proficient L3 was found.). Costa and colleagues interpreted the lack of asymmetry as evidence against inhibition. Instead, the researchers proposed that multilinguals that are highly proficient in the L2 implement a language specific selection mechanism. In a later study, Costa et al. (Reference Costa, Santesteban and Ivanova2006) further hypothesized that lexical robustness of the weaker language modulates the lexical selection mechanism. In a series of experiments, Costa et al. tested multilinguals that were highly proficient in their L1 and L2, relatively proficient in their L3 and L4, and at the initial stage of L5 learning. Results revealed symmetrical switching costs between L2 and L3 (Experiment 2) and asymmetrical switching costs between L3 and L4 (Experiment 3) and between L1 and L5 (Experiment 4). Note that in each experiment, the language pairs largely differed in proficiency. These patterns led the researchers to conclude that the lexical robustness of the weaker language needs to reach a certain threshold before the language-specific selection mechanism is engaged. Otherwise, the inhibitory control mechanism is employed. This hypothesis received support from Schwieter and Sunderman's (Reference Schwieter and Sunderman2008) study on bilinguals. In this study, unbalanced bilinguals performed a verbal fluency task in L2 and a cued language switching task. The verbal fluency (VF) task required the participants name as many items in a semantic category as possible in L2 within 60 seconds. The VF score was taken as index of the lexical robustness. Results showed that VF score correlated with the asymmetry of switching costs. Participants with lower VF scores exhibited asymmetrical switching costs. In addition, the asymmetry reduced as the VF score increased and disappeared at a certain threshold. These findings support the idea that L2 lexical robustness predicts the shift of the lexical selection mechanism in bilinguals.
Taken together, the findings reviewed above suggest that the bilingual lexical selection mechanism during word production changes as a function of the L2 proficiency. Converging evidence indicates that unbalanced bilinguals rely on the inhibitory control mechanism for lexical selection, potentially at the levels of language schema and lemma. Highly proficient bilinguals, however, may develop an alternative lexical selection mechanism, such as a language-specific mechanism. Many factors contribute to the development of the L2, such as building the L2 vocabulary, increasing the frequency of L2 use, increasing the experience of switching between two languages, etc. Notably, it has been proposed that language switching experience may play an important role in how the bilinguals’ lexical selection mechanism adapts over time (e.g., Christoffels et al., Reference Christoffels, Firk and Schiller2007; Costa & Santesteban, Reference Costa and Santesteban2004; Garbin, Costa, Sanjuan, Forn, Rodriguez-Pujadas, Ventura, Belloch, Hernandez & Avila, Reference Garbin, Costa, Sanjuan, Forn, Rodriguez-Pujadas, Ventura, Belloch, Hernandez and Avila2011). For example, Christoffels and colleagues (Reference Christoffels, Firk and Schiller2007) observed symmetrical switching costs in unbalanced bilinguals who frequently switch between languages. They speculate that extensive experience in language switching may modulate language control mechanisms and patterns of switching cost (a)symmetry within unbalanced bilinguals. Garbin et al. (Reference Garbin, Costa, Sanjuan, Forn, Rodriguez-Pujadas, Ventura, Belloch, Hernandez and Avila2011) also pointed out that due to social factors, their participants (early highly proficient Spanish–Catalan bilinguals) regularly engage in language switching according to their interlocutors and these frequent language switching experiences might bring about the changes in brain structure related to language switching.
To the best of our knowledge, no study has directly examined the plasticity of the language control mechanism during unbalanced bilinguals’ word production. Such an investigation will provide significant insights on the developmental trajectory of L2 learners’ language control ability. Thus, the present study addresses the issue of whether short-term extensive language switching experience changes the lexical selection mechanism in unbalanced bilinguals. We tested two groups of unbalanced Chinese–English bilinguals: one was the experimental group and the other was the control group. The experimental group received an extensive training in a language switching task for eight consecutive days, whereas the control group received no training at all. Both groups participated in an ERP session of language switching task before (Day 2) and after (Day 11) training. The current paradigm used a relatively long cue-to-stimulus interval (CSI) of 800 ms to allow us to investigate whether language control and its plasticity occurs at the language schema competition phase or at the lemma selection phase.
2. Material and Methods
2.1 Participants
51 unbalanced Chinese–English bilinguals participated in this study. Participants signed the informed consent form before the experiment, which was approved by the Institutional Review Board of the Imaging Center for Brain Research of Beijing Normal University. All participants were right-handed with normal or corrected-to-normal vision, no color vision disorders and no cognitive dysfunction. They were randomly assigned to the experiment group (N=26) and the control group (N=25). Two participants in the control group were excluded from the analysis, because less than 60% of the trials in each condition were left after excluding incorrect trials, outliers, and trials with excessive EEG artifacts. Thus, there were 26 participants (16 female) in the experiment group, with an average age of 22.23 (SD=2.05), and 23 participants (12 female) in the control group, with an average age of 23.22 (SD=2.41). The two groups of participants were matched in age, t(47)=−1.55, p=.128.
All participants started to learn English at an average age of 10.65 years old (SD=2.04), and the two groups were matched in the age of acquisition of English, t(47)=−1.13, p=.266. They all passed the College English Test-Band 4 (CET-4), a mandatory English proficiency test for College students in Mainland China, with a mean score of 528 (SD=48.37). The two groups were also matched in CET-4 grades, t(47)<1. According to a 2 (group: experimental group or control group) × 2 (language: L1 or L2) repeated measures ANOVA performed to their self-rating scores about language proficiency on a scale of 1 to 10 (1=not proficient; 10=highly proficient), their L1 self-rating score (M=7.70, SD=1.09) was significantly higher than their L2 score (M=5.11, SD=1.30), F(1, 47)=265.04, p<.001, suggesting the participants were dominant in their L1. Additionally, the two groups were matched in their L1 and L2 self-rating scores, as indicated by the non-significant interaction between group and language, F(1, 47)=1.65, p=.205.
Furthermore, the two groups were matched in intelligence tested by the Raven's Standard Progressive Matrices test, t(47)<1, in general cognitive control ability tested by the Simon task, t(47)<1, and in working memory capacity tested by the operational-span (O-Span) task, t(47)<1. Table 1 summarizes the language background and general cognitive ability information of the two groups. Also, according to the self-reporting scores in a socio-economic status questionnaire, the two groups were comparable in parents’ education and career, and monthly family per capita income.
Table 1. Means (Standard Deviations in Parentheses) for Individual Language Proficiency and General Cognitive Measures in the Two Groups.

Note: L2 AOA is the acquisition of age of English. L1 self-rating score is the average rating score of Chinese proficiency in reading, writing, speaking, and speech comprehension. L2 self-rating score is the average rating score of English proficiency in reading, writing, speaking, and speech comprehension. O-Span performance (%) is the percentage of correct recall.
2.2 Material
The stimuli were drawn from the entire set of Snodgrass and Vanderwart (Reference Snodgrass and Vanderwart1980) pictures, including 128 black-in-white line drawings. Eight pictures served as practice trials or filler trials, while the remaining 120 were randomly assigned to three picture sets (Sets A, B, and C) for the three experimental phases (i.e., pre-test, training, and post-test). Property data from the Chinese norms (Zhang & Yang, Reference Zhang and Yang2003) and the English norms of the pictures (Snodgrass & Vanderwart, Reference Snodgrass and Vanderwart1980) across three picture sets are presented in Table 2. As shown in the table, the stimuli across three picture sets were matched in naming consistency, familiarity, image agreement, and visual complexity. In addition, the Chinese names and English names of the pictures across three picture sets were also matched in terms of frequency and number of syllables respectively.
Table 2. Means (Standard Deviations in Parentheses) for Properties of the Three Sets of Pictures.

Note: The index of naming consistency is H, which reflects the uncertainty of naming (Zhang & Yang, Reference Zhang and Yang2003).
2.3 Procedure
For the experimental group, the experiment was composed of three phrases: pre-test, training, and post-test. The pre-test phrase lasted for two days. On Day 1, participants were required to do the Simon task in order to measure their attentional control ability (e.g., Bialystok, Craik, Klein & Viswanathan, Reference Bialystok, Craik, Klein and Viswanathan2004), the O-Span task in order to measure their working memory span (e.g., Colzato, Spapé, Pannebakker & Hommel, Reference Colzato, Spapé, Pannebakker and Hommel2007; Conway & Engle, Reference Conway and Engle1996), and the Raven's Standard Progressive Matrices task in order to measure their fluid intelligence (e.g., Raven, Raven & Court, Reference Raven, Raven and Court1998; Zhang & Wang, Reference Zhang and Wang1989). The execution order of tasks was counterbalanced across participants. Participants also completed a language history questionnaire and a socio-economic status questionnaire. On Day 2, participants performed the cued picture naming language switching task. Their behavior and electrophysiology data were recorded at the same time. After pre-test, the training phrase started and lasted for eight consecutive days (i.e., from Day 3 to Day 10). It took participants about 40 min on average to do the language switching training task on each training day. On Day 11, in the post-test, participants conducted the same cued picture naming language switching task as they did in pre-test with both the behavior and electrophysiology data collected. For the control group, participants only took part in the pre-test and the post-test.
Pre-test and post-test
A cued picture naming language switching task was used in the pre-test and the post-test phases. Participants were asked to name pictures in the language indicated by the color of the cue. The task included four conditions, namely L1 switch, L1 non-switch, L2 switch, and L2 non-switch.
During the pre-test phase, Set A (or Set C) of pictures was used, whereas at the post-test phase, Set C (or Set A) was used. The order of sets to be used was counterbalanced across participants. In this way, each participant was tested with different sets of pictures in the two phases. This design aimed to avoid the repetition effect caused by the repetition of the same stimuli used in the pre- and post-test.
The procedure was the same for both phases. Before the formal test, participants were familiarized with the Chinese and English names of the pictures. In the formal test, each trial started with a fixation point (“+”) presented at the center of the screen for 500 ms, followed by a blank screen of 300 ms. After that, a blue or red frame appeared at the center of the screen for 800 ms. Then a picture was presented within the frame for 1000 ms. The participants were asked to name the picture as quickly and accurately as possible in the target language cued by the color of the frame. A microphone and a recorder were used to record the responses within 2200 ms since picture onset. After the frame and the picture disappeared, a blank screen of 1200–1600 ms randomly was presented prior to the next trial. The jittered interstimulus interval (ISI) was used to avoid systemic noise induced by regular trial presentation.
There were eight blocks in total. Each block consisted of one filler trial and 40 critical trials, with ten trials for each condition. All trials were pseudorandomly presented. To make sure each picture was present in all four conditions, the 40 pictures were randomly assigned to 4 subsets, and then assigned to different conditions in different blocks based on the Latin square design. The whole test lasted for approximately 25 to 30 minutes. The color-language mapping was counterbalanced among participants. Before the formal test, the participants were given 8 practice trials to familiarize them with the procedure.
Training
We also used the cued picture naming language switching task for the training phrase with pictures in Set B. Prior to training, participants were familiarized with the Chinese and English names of the pictures.
In training phase, each trial started with a fixation point (“+”) presented at the center of the screen for 300 ms. It was followed by a blank screen of 200 ms. Then an orange or green frame appeared at the center of the screen for 500 ms (or 300 ms, or 100 ms, or 0 ms). The rest of the procedure was identical to that of the pre- and post-test phrases, except that the inter-trial blank screen was 1200 ms. The ISI was fixed. The colors of the frame in training were different from those in the pre- and post-test phases. This design aimed to eliminate the repetition effect caused by the same color cues.
As described above, the current training regime adopted reducing cue-to-stimulus intervals (CSIs). Some previous studies on cognitive training (Jaeggi, Buschkuehl, Jonides & Shah, Reference Jaeggi, Buschkuehl, Jonides and Shah2011; Owen, Hampshire, Grahn, Stenton, Dajani, Burns, Howard & Ballard, Reference Owen, Hampshire, Grahn, Stenton, Dajani, Burns, Howard and Ballard2010) suggest that gradually increasing the difficulty of the training task could maximize the training effect. As indicated by previous language switching literature, lengthened CSI provides longer preparation time and helps overcome the residual inhibition, leading to smaller switching costs (e.g., Costa & Santesteban, Reference Costa and Santesteban2004; Fink & Goldrick, Reference Fink and Goldrick2015; Ma, Li & Guo, Reference Ma, Li and Guo2016; Verhoef et al., Reference Verhoef, Roelofs and Chwilla2009; see similar findings using sequence-based language switching paradigm, Declerck, Philipp & Koch, Reference Declerck, Philipp and Koch2013). In other words, one way of increasing the difficulty of the language switching task is to shorten the CSI. Therefore, the current design used decreasing CSI from 500 ms, 300 ms, 100 ms to 0 ms during the training phase, with each CSI employed for two consecutive days, to increase task difficulty gradually. To maintain the novelty of the task and minimize the potential effect caused by specific cue-language mapping, on the two days using the same CSI, the cue-language mapping was reversed. A previous study found that reversing the cue-task mapping would introduce proactive interference from the formerly established cue-task associations and demand more to reorient and to remember the currently relevant cue-task mapping (Gade & Koch, Reference Gade and Koch2007). Thus, we inferred that language schema competition might benefit more from the present training regime, which was confirmed by the results and discussed in details in the discussion section.
On each day, there were four training sessions. Each session consisted of four blocks. Each block was composed of 1 filler trial and 40 critical trials distributed evenly across four conditions. The trials were presented in a pseudorandom order. Each session lasted approximately 10 minutes, resulting in 40-minute training per day. The cue-language mappings were counterbalanced across participants. Eight practice trials were provided before the formal training to familiarize the participants with the training procedure.
2.4 EEG data recording and analysis
Participants’ electroencephalograms (EEGs) were recorded from a 32-channel Quik Cap (NeuroScan Inc.) for the language switching task during the pre-test and the post-test. Horizontal electro-oculograms (EOGs) were recorded by two electrodes placed at the external canthus of each eye, while vertical EOGs were recorded by two electrodes attached around 1 cm above and below the left eye. EEGs obtained in all electrodes were referenced to the right mastoid during recording. Electrode impedances were kept below 5 kilo ohms (kΩ). The EEGs were sampled at a rate of 500 Hz, and filtered with a 0.05-100 Hz band-pass.
Trials corresponding to language error and stimuli-locked phase corresponding to naming error were removed firstly off-line. Trials affected by ocular artifacts were corrected by the arithmetic built in the Neuroscan analysis software. All electrodes were re-referenced to the average of both mastoids. Then EEGs were filtered at a low-pass of 30Hz (24 dB). After that, EEGs were segmented to 700-ms epochs time-locked to the cue with a baseline of 100 ms pre-cue presentation. Likewise, 700-ms epochs time-locked to the stimulus were segmented, with a baseline of 100 ms before stimulus onset. Epochs with voltages exceeding ±100 μV were rejected to eliminate data contaminated by eye blinks, eye movement and muscle movement. To guarantee the ratio of signal to noise, for both the cue-locked epochs and the stimulus-locked epochs, there must be at least 48 (60%) epochs left after the pre-processing in each condition for each participant. Cue-locked and stimulus-locked epochs in each condition were then grand-averaged respectively. Two participants’ data in the control group were rejected due to inadequate number (less than 48, 60%) of trials in each condition.
Following previous language switching studies (e.g., Christoffels et al., Reference Christoffels, Firk and Schiller2007; Jackson et al., Reference Jackson, Swainson, Cunnington and Jackson2001; Verhoef et al., Reference Verhoef, Roelofs and Chwilla2009), the present study focuses on the N2 component, examining the potential effect of language switching training on N2 deflections between switch and non-switch conditions in the two languages. Based on visual inspection of the cue-locked grand average waveforms for both groups in both pre- and post-test, for each condition, N2 peak latency in the 200–350 ms time window and N2 mean amplitude in the time window 30 ms before and 30 ms after the peak latency were extracted from F3, FZ, F4, FC3, FCZ, FC4, C3, CZ and C4 electrode sites. Besides, based on previous studies (Christoffels et al., Reference Christoffels, Firk and Schiller2007; Jackson et al., Reference Jackson, Swainson, Cunnington and Jackson2001; Verhoef et al., Reference Verhoef, Roelofs and Chwilla2009) and visual inspection of the stimulus-locked grand average waveforms for both groups in pre- and post-test, N2 mean amplitudes were extracted from two time windows (to detect potential effects more subtly, see also Christoffels et al., Reference Christoffels, Firk and Schiller2007; Guo et al., Reference Guo, Ma and Liu2013b), 180–250 ms and 250–350 ms post stimulus-onset, for each condition in the same nine electrode sites as aforesaid. A 2 (group: experimental group or control group) × 2 (test session: pre-test or post-test) × 2 (language: L1 or L2) × 2 (trial type: switch trials or non-switch trials) × 9 (electrodes: F3, FZ, F4, FC3, FCZ, FC4, C3, CZ, and C4) repeated measures ANOVA was conducted on each dependent variable. The Greenhouse-Geisser correction for non-spherity was applied when appropriate. Unadjusted degrees of freedom and adjusted p values were reported. As the present study focuses on the potential modulation of language switching training on the language control in unbalanced bilinguals, only the significant main effects of group, test session, language, trial type, and interactions involved one or more of these four factors were reported.
3. Results
3.1 Behavioral results
Only correct responses were entered into the analysis, excluding incorrect responses (5.63%). RTs beyond the range of 200–2000 ms (0.86%) and those 2.5 standard deviation slower or faster than each individual's mean RT (1.95%) were further excluded. Figures 1 and 2 show the mean RTs and ERs in different conditions of the two groups in pre-test and post-test (for behavioral results over training sessions, see Figure S1 and Figure S2 in supplementary material, Supplementary Materials).

Figure 1. Mean RTs of switch and non-switch naming in L1 and L2 during pre-test and post-test for the two groups. Error bars show one standard error.

Figure 2. Mean ERs of switch and non-switch naming in L1 and L2 during pre-test and post-test for the two groups. Error bars show one standard error.
Separate 2 (group: experimental group or control group) × 2 (test session: pre-test or post-test) × 2 (language: L1 or L2) × 2 (trial type: switch trial or non-switch trial) repeated-measures ANOVAs were conducted on RTs and ERs, respectively. RTs results showed a significant main effect of trial type, F(1, 47)=74.10, p<.001, ηp 2 =.612, suggesting that naming in switch trials (1014 ms) was significantly slower than naming in non-switch trials (979 ms), resulting in a switching cost of 35 ms. None of the main effect of group, F(1, 47)<1, test session, F(1, 47)=2.60, p=.114, ηp 2 =.052, language, F(1, 47)=2.09, p=.154, ηp 2 =.043, the interactions between group and test session, F(1, 47)=2.51, p=.120, ηp 2 =.051, between group and language, F(1, 47)=2.88, p=.096, ηp 2 =.058, between group and trial type, F(1, 47)=2.28, p=.138, ηp 2 =.046, between test session and language, F(1, 47)<1, between group, test session and language, F(1, 47)=1.81, p=.185, ηp 2 =.037, between test session and trial type, F(1, 47)<1, between group, test session and trial type, F(1, 47)<1, between language and trial type, F(1, 47)=2.93, p=.093, ηp 2 =.059, between group, language and trial type, F(1, 47)<1, between test session, language and trial type, F(1, 47)=1.51, p=.225, ηp 2 =.031, or between group, test session, language and trial type, F(1, 47)<1, reached significant level.
According to the analyses for the ERs, the main effect of test session was significant, F(1, 47)=10.40, p=.002, ηp 2 =.181, revealing lower ERs in post-test (4.50%) than in pre-test (6.07%). The main effect of language reached significance, F(1, 47)=17.65, p<.001, ηp 2 =.273, with more inaccurate naming in English (6.42%) than in Chinese (4.15%). In addition, the main effect of trial type was significant, F(1, 47)=16.74, p<.001, ηp 2 =.263, suggesting that naming in switch trials (5.76%) was less accurate than naming in non-switch trials (4.81%). The results also revealed a significant interaction effect between test session and language, F(1, 47)=9.49, p=.003, ηp 2 =.168. Further simple effect test found that for naming in English, performance in post-test (5.08%) was significantly better than that in pre-test (7.76%), p=.001. For naming in Chinese, however, there was no significant difference between performance in post-test (3.93%) and that in pre-test (4.37%), p=.322. The interaction between test session and trial type was also significant, F(1, 47)=5.98, p=.018, ηp 2 =.113. Further simple effect test showed that in pre-test, naming in switch trials (6.80%) was significantly less accurate than naming in non-switch trials (5.33%), p<.001, whereas in post-test, no significant difference was observed (4.71% and 4.29%), p=.212.
3.2 ERP results
Cue-locked ERP results
Figures 3 and 4 present the cue-locked ERPs for different conditions in pre-test and post-test for the experimental group and the control group, respectively. All conditions in pre- and post-test elicited an N100 component, a P100 component, and an N2 component, the main component of interest of the present study. Based on visual inspection, the critical difference between the post-test and the pre-test seems to occur on the latency and magnitude of the N2 component, a pattern which was confirmed by the results of ANOVAs.

Figure 3. The grand average waveforms for switch (dash line) and non-switch trials (solid line) in L1 (blue) and L2 (red) in cue phase for the experimental group in pre-test (left panel) and post-test (right panel) at three representative electrode sites.

Figure 4. The grand average waveforms for switch (dash line) and non-switch trials (solid line) in L1 (blue) and L2 (red) in cue phase for the control group in pre-test (left panel) and post-test (right panel) at three representative electrode sites.
N2 peak latency
The results of ANOVA conducted on N2 peak latencies showed that the main effect of test session was significant, F(1, 47)=13.01, p=.001, ηp 2 =.217, indicating that N2 peak latency in the post-test (272 ms) was shorter than that in the pre-test (285 ms). More importantly, the interaction between test session and group was significant, F(1, 47)=8.51, p=.005, ηp 2 =.153. Further simple effect test revealed that for the experimental group, N2 peak latency in the post-test (262 ms) was earlier than that in the pre-test (285 ms), p<.001. In contrast, for the control group, the difference between the N2 peak latency in the post-test (282 ms) and pre-test (284 ms) was not significant, p=.638 (see Figure 5).
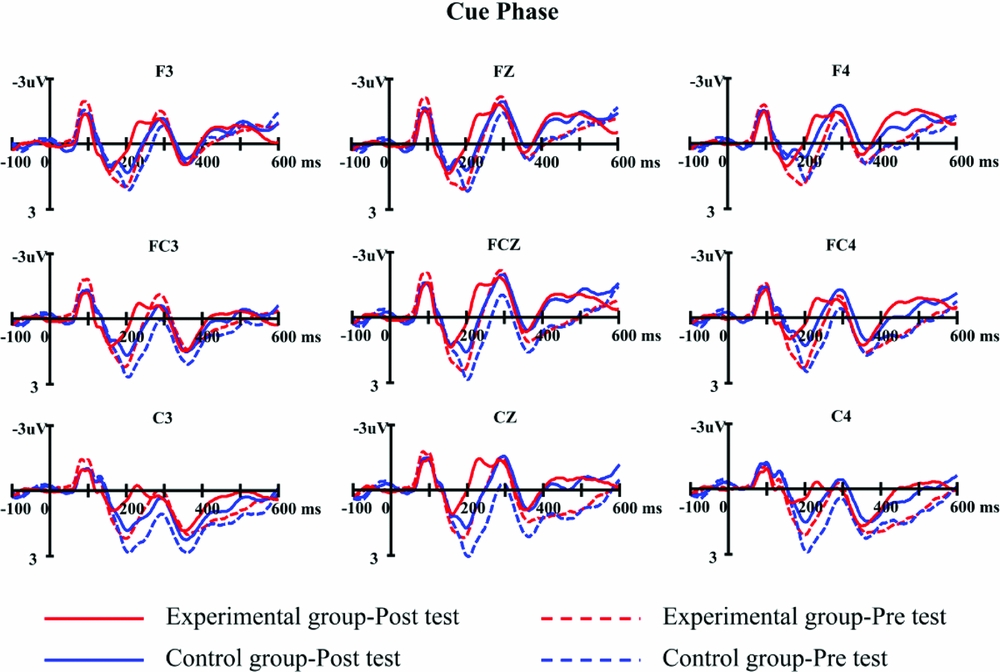
Figure 5. The grand average waveforms collapsed across languages and trial types in cue phase for the experimental group (red) and control group (blue) in pre-test (dash line) and post-test (solid line) at nine electrode sites.
N2 mean amplitude
The results of the ANOVA on mean amplitude of N2 revealed a significant main effect of test session, F(1, 47)=11.34, p=.002,ηp 2 =.194, suggesting that the mean amplitude of the N2 in the post-test (-1.67μV) was more negative than that in the pre-test (-0.96μV). The main effect of trial type was marginally significant, F(1, 47)=4.02, p=.051,ηp 2 =.079, suggesting that N2 elicited by switch trials (−1.42μV) tended to be more negative than that by non-switch trials (−1.20μV). There was also a significant interaction between language and group, F(1, 47)=8.13, p=.006,ηp 2 =.148. Further simple effect test revealed that for the experimental group, the N2 induced by Chinese naming (−1.79μV) was larger than that induced by English naming (−1.44μV), p=.048. In contrast, for the control group, the difference between the N2 induced by English naming (−1.20μV) and Chinese naming (−0.83μV) was marginally significant, p=.051. Additionally, the interaction of test session and electrodes was significant, F(8, 376)=3.42, p=.007,ηp 2 =.068. Further analyses showed that except for F3 and FZ, the N2 component in the post-test was more negative than that in the pre-test over the remaining 7 electrode sites, ps<.05.
Stimulus-locked ERP results
Figures 6 and 7 present the stimulus-locked ERPs across different conditions in the pre-test and the post-test for the experimental group and the control group respectively. All conditions elicited an N100 component, a P100 component, followed by an N2 complex lasting from about 180 ms to 350 ms post stimulus onset. Based on visual inspection, the difference between the post-test and the pre-test seems to occur on the amplitude of N2, which was confirmed by the results of ANOVAs.
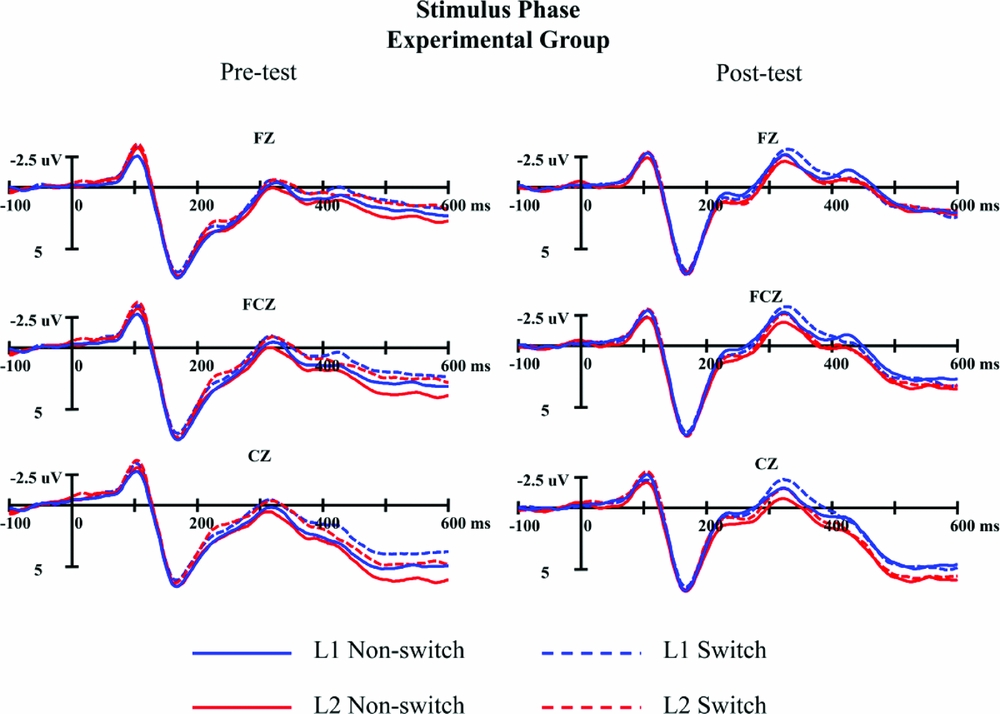
Figure 6. The grand average waveforms for switch (dash line) and non-switch trials (solid line) in L1 (blue) and L2 (red) in stimulus phase for the experimental group in pre-test (left panel) and post-test (right panel) at three representative electrode sites.
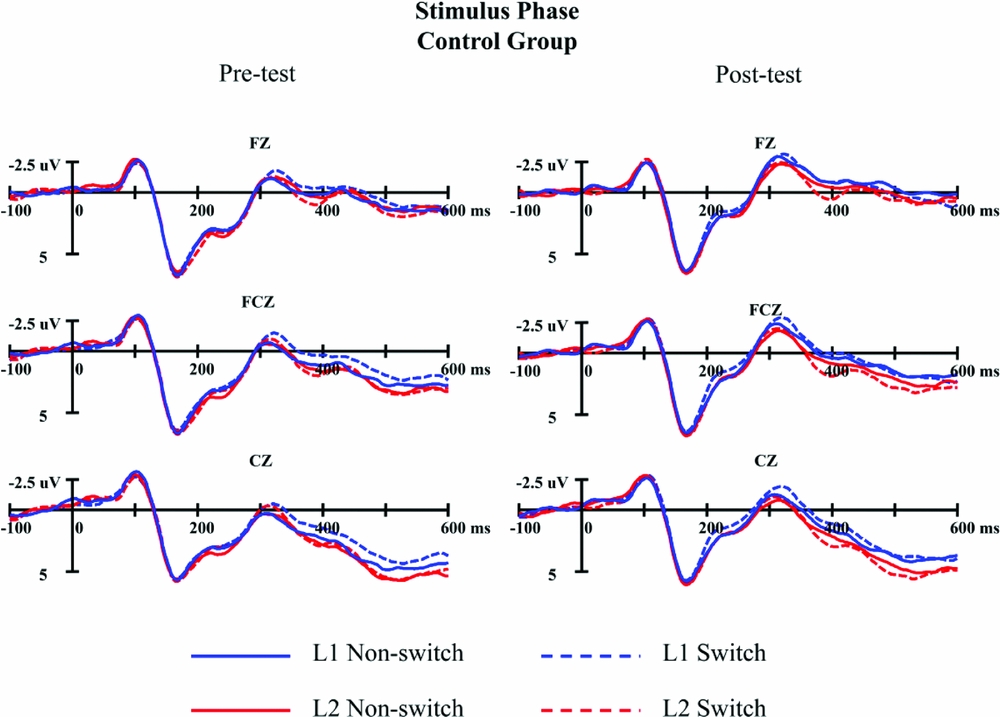
Figure 7. The grand average waveforms for switch (dash line) and non-switch trials (solid line) in L1 (blue) and L2 (red) in stimulus phase for the control group in pre-test (left panel) and post-test (right panel) at three representative electrode sites.
N2 mean amplitude in time window 180–250 ms
The results of ANOVA on N2 mean amplitude in 180–250 ms after stimulus onset showed a significant main effect of test session, F(1, 47)=23.29, p<.001,ηp 2 =.331, indicating that the N2 was more negative-going in the post-test (3.11μV) than that in the pre-test (4.38μV). The main effect of language was significant, F(1, 47)=4.87, p=.032,ηp 2 =.094, with a more negative N2 elicited by Chinese naming (3.63μV) than that elicited by English naming (3.86μV). The results also found a significant main effect of trial type, F(1, 47)=4.72, p=.035, ηp 2 = .091, revealing that switch trials (3.62μV) induced a larger N2 than non-switch trials (3.87μV). The interaction effect between test session and electrodes was significant, F(8, 376)=6.81, p<.001, ηp 2 =.127. Further simple effect test revealed that the N2 in the post-test was significantly larger than that in the pre-test over all electrode sites, F3(MD=−1.16, p<.001), FZ(MD=−1.45, p<.001), F4(MD=−1.63, p<.001), FC3(MD=−1.00, p<.001), FCZ(MD=−1.32, p<.001), FC4(MD=−1.61, p<.001), C3(MD=−.81, p=.003), CZ(MD=−1.04, p<.001), C4(MD=−1.38, p<.001). The results also showed a significant interaction between trial type and electrodes, F(8, 376)=6.38, p<.001, ηp 2 =.119. Further simple effect test revealed that the N2 effect (a larger N2 in switch trials compared to non-switch trials) was found in FC3, FCZ, C3 and CZ electrode sites, ps<.022, while no such robust effects showed in the other five electrode sites, ps > .075.
N2 mean amplitude in time window 250–350 ms
The ANOVA results on the N2 mean amplitude in 250–350 ms after stimulus onset were consistent with those in 180–250 ms. The main effect of test session was significant, F(1, 47)=45.20, p<.001,ηp 2 =.490, with a more negative N2 in post-test (0.29μV) than that in pre-test (1.77μV). The main effect of language was also significant, F(1, 47)=8.69, p=.005,ηp 2 =.156, suggesting that N2 elicited by naming in Chinese (0.86μV) was larger than naming in English (1.21μV). The main effect of trial type also reached significance, F(1, 47)=10.51, p=.002,ηp 2 =.183. Switch trials (0.87μV) induced a more negative N2 than non-switch trials (1.19μV). The results also showed a significant interaction between test session and electrodes, F(8, 376)=7.29, p<.001, ηp 2 =.134. Like the results in 180–250 ms, over all electrode sites, the N2 in the post-test was significantly larger than in the pre-test, F3(MD=−1.39, p<.001), FZ(MD=−1.72, p<.001), F4(MD=−2.06, p<.001), FC3(MD=−1.13, p<.001), FCZ(MD=−1.59, p<.001), FC4(MD=−1.95, p<.001), C3(MD=−.81, p=.001), CZ(MD=−1.29, p<.001), C4(MD=−1.42, p<.001). The interaction effect of trial type by electrodes was also significant, F(8, 376)=8.34, p<.001, ηp 2 =.151. Further analyses revealed a significant N2 effect in electrodes FC3, FCZ, FC4, C3, CZ and C4, ps<.025, while there was only a marginally significant effect in F3, p=.056, and no such effect in other sites, ps > .118.
4. Discussion
The present study examined the impact of short-term language switching training on the lexical selection mechanism of word production in unbalanced bilinguals. Specifically, the present study trained a group of unbalanced Chinese–English bilinguals in a cued language switching task intensively for 8 consecutive days, and compared their performance with behavioral and EEG measures in a pre-test and a post-test of the same task. Further comparisons were made between the experimental group and a matched control group, which received no training.
4.1 The loci of language control
Our behavioral results revealed robust switching costs in both the experimental and the control group. That is, compared with naming in non-switch trials, naming in switch trials was slower and less accurate, which was consistent with previous findings of language switching studies (Christoffels et al., Reference Christoffels, Firk and Schiller2007; Costa & Santesteban, Reference Costa and Santesteban2004; Costa et al., Reference Costa, Santesteban and Ivanova2006; Declerck, Koch & Philipp, Reference Declerck, Koch and Philipp2012; Fink & Goldrick, Reference Fink and Goldrick2015; Gollan & Ferreira, Reference Gollan and Ferreira2009; Jackson et al., Reference Jackson, Swainson, Cunnington and Jackson2001; Linck et al., Reference Linck, Schwieter and Sunderman2012; Ma et al., Reference Ma, Li and Guo2016; Martin, Strijkers, Santesteban, Escera, Hartsuiker & Costa, published online November 5, 2013; Meuter & Allport, Reference Meuter and Allport1999; Schwieter & Sunderman, Reference Schwieter and Sunderman2008; Verhoef et al., Reference Verhoef, Roelofs and Chwilla2009, Reference Verhoef, Roelofs and Chwilla2010). The switching costs could be readily explained under the framework of IC model (Green, Reference Green1998; for other explanations, see recent reviews Baus et al., Reference Baus, Branzi, Costa and Schwieter2015; Bobb & Wodniecka, Reference Bobb and Wodniecka2013; Declerck & Philipp, Reference Declerck and Philipp2015). In switch trials, the target language in the current trial was inhibited in the former trial as the non-target language. To reactivate the target language in the current trial, previous inhibition exerted on it needs to be overcome, resulting in longer RTs and higher ERs.
Interestingly, we did not find a significant difference in the magnitude of the switching costs between the L1 and the L2. That is, there is no evidence for asymmetrical switching costs in unbalanced bilinguals, which is inconsistent with the previous studies (Costa & Santesteban, Reference Costa and Santesteban2004; Costa et al., Reference Costa, Santesteban and Ivanova2006, Experiment 1; Jackson et al., Reference Jackson, Swainson, Cunnington and Jackson2001; Meuter & Allport, Reference Meuter and Allport1999). This finding might arise from the relatively long cue-to-stimulus intervals (CSI) in the current study. To investigate the potential changes caused by bilingual language switching training during the language schema competition phase and/or the lemma selection phase, a relatively large CSI of 800 ms was used. Previous studies found that when bilinguals were provided with more time to prepare for the target language with a longer CSI, the asymmetry of switching costs could be attenuated or eliminated (e.g., Verhoef et al., Reference Verhoef, Roelofs and Chwilla2009, Reference Verhoef, Roelofs and Chwilla2010; see also Guo et al., Reference Guo, Ma and Liu2013b for related discussions). Alternatively, since the response-to-stimulus interval (RSI) was relatively long (at least 2000 ms) and variable in the pre- and post-test, there would be another possibility that such long passive decay might eliminate the asymmetry across languages (see Declerck et al., Reference Declerck, Koch and Philipp2012, and Declerck, Koch & Philipp, Reference Declerck, Koch and Philipp2015a, for related discussions).
Critically, ERP results of the present study showed that a more negative N2 component was elicited by switch trials relative to non-switch trials in ERPs time-locked to the stimulus for the experimental group and the control group. This is compatible with the N2 effect reported in previous language switching studies (e.g., Jackson et al., Reference Jackson, Swainson, Cunnington and Jackson2001; Guo et al., Reference Guo, Ma and Liu2013b). As the N2 effect has been taken as an indicator for response inhibition (Jackson et al., Reference Jackson, Jackson and Roberts1999; Schmitt et al., Reference Schmitt, Münte and Kutas2000; Thorpe et al., Reference Thorpe, Fize and Marlot1996) or conflict detection (Nieuwenhuis et al., Reference Nieuwenhuis, Yeung, Van Den Wildenberg and Ridderinkhof2003), our finding indicates that the lexical selection mechanism exerts language control during bilingual word production. However, it should be noted that our results are not in line with some previous findings (Verhoef et al., Reference Verhoef, Roelofs and Chwilla2009, Reference Verhoef, Roelofs and Chwilla2010). The evidence regarding the N2 effect in the cue- and/or stimulus-locked ERPs has been mixed. The N2 effect has been reported for both languages in the cue phase (Verhoef et al., Reference Verhoef, Roelofs and Chwilla2010), but not in the stimulus phase (Verhoef et al., Reference Verhoef, Roelofs and Chwilla2009). Conversely, using the n-2 repetition task, Guo et al. (Reference Guo, Ma and Liu2013b) only observed an N2 effect in the stimulus phase, with n-2 repetition trials inducing a more negative N2 than n-2 non-repetition trials, suggesting that inhibition of the non-target language occurs at the lemma level. We suspect that the incongruence across studies probably results from experimental factors such as the difference in the tasks and stimuli. In the present study, we observed an N2 effect in the stimulus phase, again indicating that inhibitory control occurs in lemma selection phase (Green, Reference Green1998). It will remain for ongoing research to determine whether and how potential factors contribute to the discrepant patterns available.
The present study did not show asymmetrical switching costs of two languages on the ERP component, consistent with previous studies using relatively long preparation time to select the target language (Guo et al., Reference Guo, Ma and Liu2013b; Verhoef et al., Reference Verhoef, Roelofs and Chwilla2010). This is compatible with the symmetrical switching costs in our behavioral data, which again could be attributed to the relatively long preparation time or long passive decay provided for the unbalanced bilinguals.
Additionally, the stimulus-locked ERP results showed that naming in Chinese induced a larger N2 component than naming in English. Evidence from behavior studies has accumulated that naming in the dominant language was comparable or even slower than naming in the non-dominant language under mixing-language context (Christoffels et al., Reference Christoffels, Firk and Schiller2007; Costa & Santesteban, Reference Costa and Santesteban2004; Costa et al., Reference Costa, Santesteban and Ivanova2006; Declerck et al., Reference Declerck, Philipp and Koch2013; Declerck et al., Reference Declerck, Thoma, Koch and Philipp2015b; Gollan & Ferreira, Reference Gollan and Ferreira2009; Gollan, Kleinman & Wierenga, Reference Gollan, Kleinman and Wierenga2014; Verhoef et al., Reference Verhoef, Roelofs and Chwilla2009). This phenomenon, often referred to as global slowing, has been considered as another indicator of inhibition exerted on the dominant language during bilingual language switching in word production (for a review, see Bobb & Wodniecka, Reference Bobb and Wodniecka2013). To our best knowledge, our finding is the first piece of evidence for global slowing in the N2 ERP component. Parallel to the ERP result, our RT data show that the main effect of language was not significant, reflecting the absence of the standard language dominance effect. Taken together, our findings indicate that bilinguals recruit strong inhibition on the L1 in the lemma selection phase during word production.
4.2 The effect of training on language control
First of all, our behavioral results revealed no significant effect of training in RTs. One possible reason is that the RTs reflect aggregated results of various processes after stimulus presentation (e.g., Jackson et al., Reference Jackson, Swainson, Cunnington and Jackson2001), whereas training effect might be exhibited mainly in the processing of the cue, namely during the language task schema competition phase. This speculation is confirmed by our ERP results.
More specifically, ERP results in the present study show that after intensive short-term language switching training, the peak latency of N2 elicited by cue processing in the post-test was significantly shorter than that in the pre-test in the experimental group. On the other hand, no such difference was observed in the control group. Previous research has associated shorter latency of the N2 component with stronger attention control in healthy adults relative to older adults (Beck, Swanson & Dustman, Reference Beck, Swanson and Dustman1980; Schroeder, Lipton, Ritter, Giesser & Vaughan, Reference Schroeder, Lipton, Ritter, Giesser and Vaughan1995), children and younger adults (Johnstone, Pleffer, Barry, Clarke & Smith, Reference Johnstone, Pleffer, Barry, Clarke and Smith2005), and patients with multiple system atrophy (Kamitani & Kuroiwa, Reference Kamitani and Kuroiwa2009). In addition, Falkenstein, Hoormann and Hohnsbein (Reference Falkenstein, Hoormann and Hohnsbein1999) reported that stronger inhibitory ability reflected by lower false alarm rates correlated with shorter N2 latency in a Go/Nogo task. Thus, the shortened N2 peak latency observed in the present study suggests that the language control in language task schema competition phase was accelerated after language switching training.
One possible reason is that short-term intensive training with the cued language switching task improved the unbalanced bilinguals’ efficiency of establishing the language task schema based on the cue. According to the IC model (Green, Reference Green1998), the task schema comprises the mental networks constructed or adapted to achieve a certain task. The current study used decreasing CSIs over the training procedure to increase the difficulty of the training and maximize the effect of the training. In other words, the time to prepare for the target language reduced gradually, posing a higher demand for more effective language control. In addition, the cue-language mapping was reversed on the two days with the same CSI. This manipulation was to keep the novelty of the task and to minimize the potential effect of fixed cue-language mapping. The unbalanced bilinguals are likely to detect the cue rapidly to establish the target language schema more efficiently, and to recruit more cognitive resource to maintain the goal.
Interestingly, the training effect seemed to be modulated by individual difference, which was indicated by the dual peak N2 observed only in the post-test cue-locked ERPs for the experimental group (Figure 5). Close examination revealed that after training, the peak latency of N2 was shortened to a larger extent for some participants than others in the experimental group. Thus, when averaged together, the grand average waveforms manifested a dual peak N2. Unfortunately, no significant correlation was found between the training effect of N2 peak latency and the L2 AOA, L2 proficiency, intelligence, attentional control ability or working memory capacity measured in the present study. We speculated that variance in the individual difference measures due to relatively small numbers of participants might not be powerful enough for us to determine the impact of individual difference on the discrepancy in N2 latency effect. More future studies are thus needed to examine the role of individual differences in the plasticity of bilingual language control.
Notably, the training effect of the present study was present in the peak latency rather than the mean amplitude of the N2 component induced by cue processing, which is inconsistent with the previous findings of the language switching costs indexed by the N2 amplitude (Christoffels et al., Reference Christoffels, Firk and Schiller2007; Guo et al., Reference Guo, Ma and Liu2013b; Jackson et al., Reference Jackson, Swainson, Cunnington and Jackson2001; Verhoef et al., Reference Verhoef, Roelofs and Chwilla2009, Reference Verhoef, Roelofs and Chwilla2010). One possibility is that the present study is a dynamic training study, while the previous studies on language switching were static, where the strength of the language control was reflected by the amplitude of N2. However, language switching training is likely to improve the language control efficiency, just as we found in the present study that the N2 peak latency was shortened after training. In one recent study, Sullivan, Janus, Moreno, Astheimer and Bialystok (Reference Sullivan, Janus, Moreno, Astheimer and Bialystok2014) found that after 6-months L2 training, the training group manifested shortened peak latency of the N2 and P3 components in a Go/Nogo task, while the control group didn't show such changes. These findings together with the present findings thus shed light on the necessity for future research to focus on the potential effect of the N2 peak latency that could reflect the cognitive control ability.
Furthermore, it is important to note that we used a passive control group which did not receive any training to compare with the experimental group, a design adopted by many previous studies on cognitive training (e.g., Chein & Morrison, Reference Chein and Morrison2010; Jaeggi, Buschkuehl, Jonides & Perrig, Reference Jaeggi, Buschkuehl, Jonides and Perrig2008; Rudebeck, Bor, Ormond, O'Reilly & Lee, published online November 28, 2012). Although this design is likely to ensure that the control group was deprived of experiment-related language switching experience and to rule out the improvement resulting from task repetition, it should be acknowledged that it cannot exclude the potential role of other factors, such as cognitive investment, motivation, and expectations for improvement, on the effect of training (for a review, see Morrison & Chein, Reference Morrison and Chein2011; Redick, Shipstead, Harrison, Hicks, Fried, Hambrick, Kane & Engle, Reference Redick, Shipstead, Harrison, Hicks, Fried, Hambrick, Kane and Engle2013). To overcome this limitation, a more reasonable design is to adopt an appropriate active control group (Boot, Simons, Stothart & Stutts, Reference Boot, Simons, Stothart and Stutts2013; Green, Strobach & Schubert, Reference Green, Strobach and Schubert2014; Morrison & Chein, Reference Morrison and Chein2011). For the present study, one possible improvement is to include an active control group that takes a single language picture naming training for 8 days with the intended language in two consecutive days different (e.g., Day1, L1; Day 2, L2). This training regime may guarantee that the experimental group and the control group have the same amount of training of word production in both languages and exposure duration to the experimental environment, but different amount of language switching experience.
To summarize, the current study shows that after training, the N2 peak latency in the cue-locked ERPs was reduced in the experimental group, suggesting improved efficiency of establishing the target language schema. In other words, the current patterns indicate that language switching training modulates unbalanced bilinguals’ lexical selection mechanism. It's worth noting that different language switching training regimes may lead to varying training effects. To better understand whether and how specific aspects of the language switching experience modulate the bilingual lexical selection mechanism during word production, it remains to be investigated what other training regimes could be used, what the training effects are, and how to maximize the training effects.
4.3 The effect of task repetition
Comparison between the pre-test and post-test revealed that the ERs were lower in the post-test in both the experimental and control groups. In addition, the N2 amplitude was larger during the cue phase and the stimulus phase in the post-test in both groups. Notably, to minimize the effect of task repetition, different pictures were used in the pre- and the post-test. Nonetheless, the effect of task repetition was present in ERs and the N2 amplitude. The effect of task repetition in behavioral data has been well documented in the task training literature (e.g., Karbach & Kray, Reference Karbach and Kray2009; Zinke, Einert, Pfennig & Kliegel, published online March 26, 2012). Furthermore, the enlarged N2 component has been associated with increased efforts to allocate cognitive resources for executing control (Band, Ridderinkhof & van der Molen, Reference Band, Ridderinkhof and van der Molen2003; Chang, Chen, Li & Li, published online June 16, 2014; Jodo & Kayama, Reference Jodo and Kayama1992; Larson & Clayson, Reference Larson and Clayson2011; Larson, Clayson & Baldwin, Reference Larson, Clayson and Baldwin2012). Thus, the decreased ERs and the enhanced N2 amplitude in the post-test indicate that the repetition of the task leads all participants to strategically enhance their employment of cognitive sources to exert control during both phases and in turn results in improved behavioral performance.
5. Conclusions
The present study trained unbalanced Chinese–English bilinguals with the cued language switching task. Stimulus-locked ERP data showed that switch trials elicited a more negative-going N2 component than non-switch trials in both the pre-test and the post-test across the experimental and control groups, indicating that unbalanced bilinguals exert inhibitory control during the lemma selection phase. More importantly, the N2 peak latency in the cue-locked ERPs was shortened after training in the experimental group only. As cue-locked ERPs are indicative of the task schema competition phase, the observed pattern suggests that language switching training improves the efficiency of establishing the target language schema, indicating the plasticity of unbalanced bilinguals’ lexical selection mechanism during word production.
Supplementary Material
To view supplementary material for this article, please visit https://doi.org/10.1017/S1366728917000037










