



1. Introduction
Recent geological maps of Việt Nam show the country to comprise, in north–south orientation, five distinct continental fragments separated by major shear or suture zones (Burrett et al. Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014, fig. 1). Sedimentary rocks mapped as Cambrian span three of these terranes (Phạm, Reference Phạm2008) (Fig. 1), and their fossils and mineral content may be used to explore the original geological affinities of these fragments and thus to test various tectonic hypotheses recently proposed to explain their disposition. Here we provide new data on fossils and detrital zircons from the poorly known Sông Đà terrane and detrital zircons from both the North Việt Nam terrane immediately to the north of the Sông Đà terrane, and from the Trường Sơn terrane, immediately to its south. This study is part of a series of papers that focus on upper Cambrian sedimentary rocks of south and southeast Asia; their widespread occurrence allows for comparison among rocks whose collective depositional age span can be constrained to within a few million years (e.g. Peng et al. Reference Peng, Hughes, Heim, Sell, Zhu, Myrow and Parcha2009; Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011; Hughes, Reference Hughes2016; Myrow et al. Reference Myrow, Hughes, McKenzie, Pelgay, Thompson, Haddad and Fanning2016; Wernette et al. Reference Wernette, Hughes, Myrow and Sardsud2020 a,b, Reference Wernette, Hughes, Myrow and Aye2021). Such information is pertinent to a long-standing problem in Gondwanan geology: how was equatorial Gondwana configured prior to the opening of the Palaeotethys Ocean?
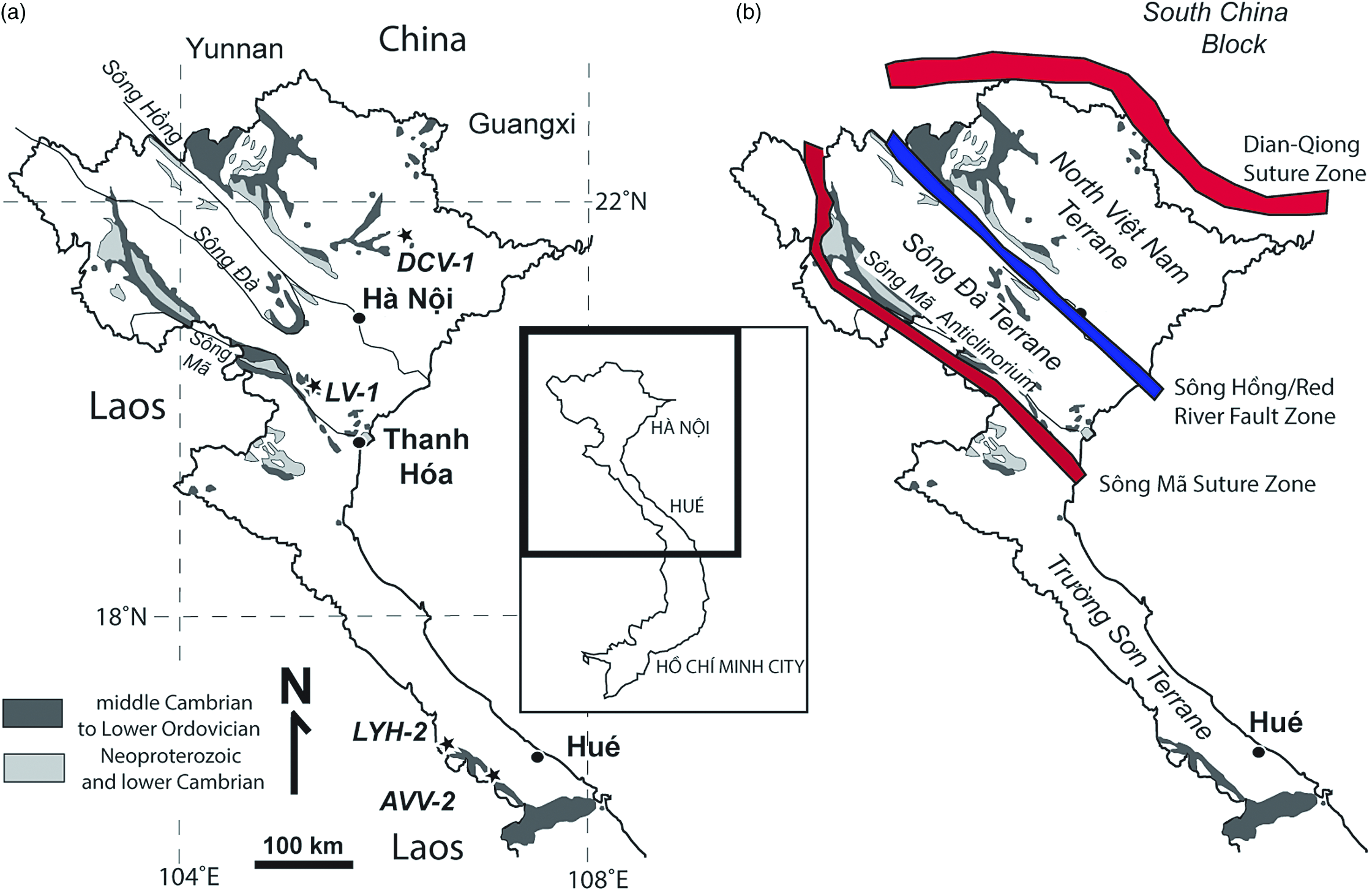
Fig. 1. Map of northern Việt Nam showing outcrops of Cambrian and stratigraphically adjacent rocks and positions of major sutures and shear zones. Inset shows the portion of Việt Nam figured along with the locations of major cities. (a) Outcrops of Cambrian and stratigraphically adjacent rocks. The courses of three major rivers, the Sông Hồng (Red River), Sông Đà and Sông Mã are also shown. Modified from Phạm (Reference Phạm2008, fig. 25). Localities from whence detrital samples were collected are indicated by stars and sample abbreviations. (b) Positions of major suture zones (red) and shear zone (blue) in northern Việt Nam, along with the names of each tectonic unit, and the position of the Sông Mã anticlinorium are identified.
Fundamental early work on Vietnamese Cambrian fossils by Mansuy (Mansuy, Reference Mansuy1915, Reference Mansuy1916), not yet comprehensively revised, has been accompanied by more recent investigations by Phạm Kim Ngân (Phạm, Reference Phạm, Peng, Babcock and Zhu2001, Reference Phạm2008) and others. Our finds of Cambrian and Tremadocian fossils were made in 2008 from Thanh Hóa province, which is also known as the west Bắc Bộ – north Trung Bộ region (Phạm, Reference Phạm2008, p. 179), and part of the ‘Sông Đà terrane’ (e.g. Burrett et al. Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014). This fault-bounded terrane lies within a geologically complex region between the South China/Yangtze and Indochina blocks. It is located south of the fast slipping Sông Hồng/Sông Chảy (Red River) fault zone (Leloup et al. Reference Leloup, Lacassin, Tapponnier, Schärer, Zhong, Lui, Zhang, Ji and Phan1995; Yin & Nie, Reference Yin, Nie, Yin and Harrison1996) (Fig. 1). Rocks north of this fault that belong to the North Việt Nam terrane (Cai & Zhang, Reference Cai and Zhang2009) have yielded the great majority of Vietnamese Cambrian fossils described to date, including all of Mansuy’s figured Cambrian material. These rocks have traditionally been considered part of the South China block (Burrett et al. Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014), although this view is not universally accepted (e.g. Cai & Zhang, Reference Cai and Zhang2009; Faure et al. Reference Faure, Lepvrier, Nguyễn, Vũ, Lin and Chen2014; Halpin et al. Reference Halpin, Trần, Lai, Meffre, Crawford and Khin2016). The Sông Đà terrane lies immediately to the south of the Sông Hồng fault. The Sông Mã suture zone defines the southern boundary of the Sông Đà terrane, and separates it from the Trường Sơn terrane (Figs 1, 2) that is here treated as part of Indochina (see Section 9 below).
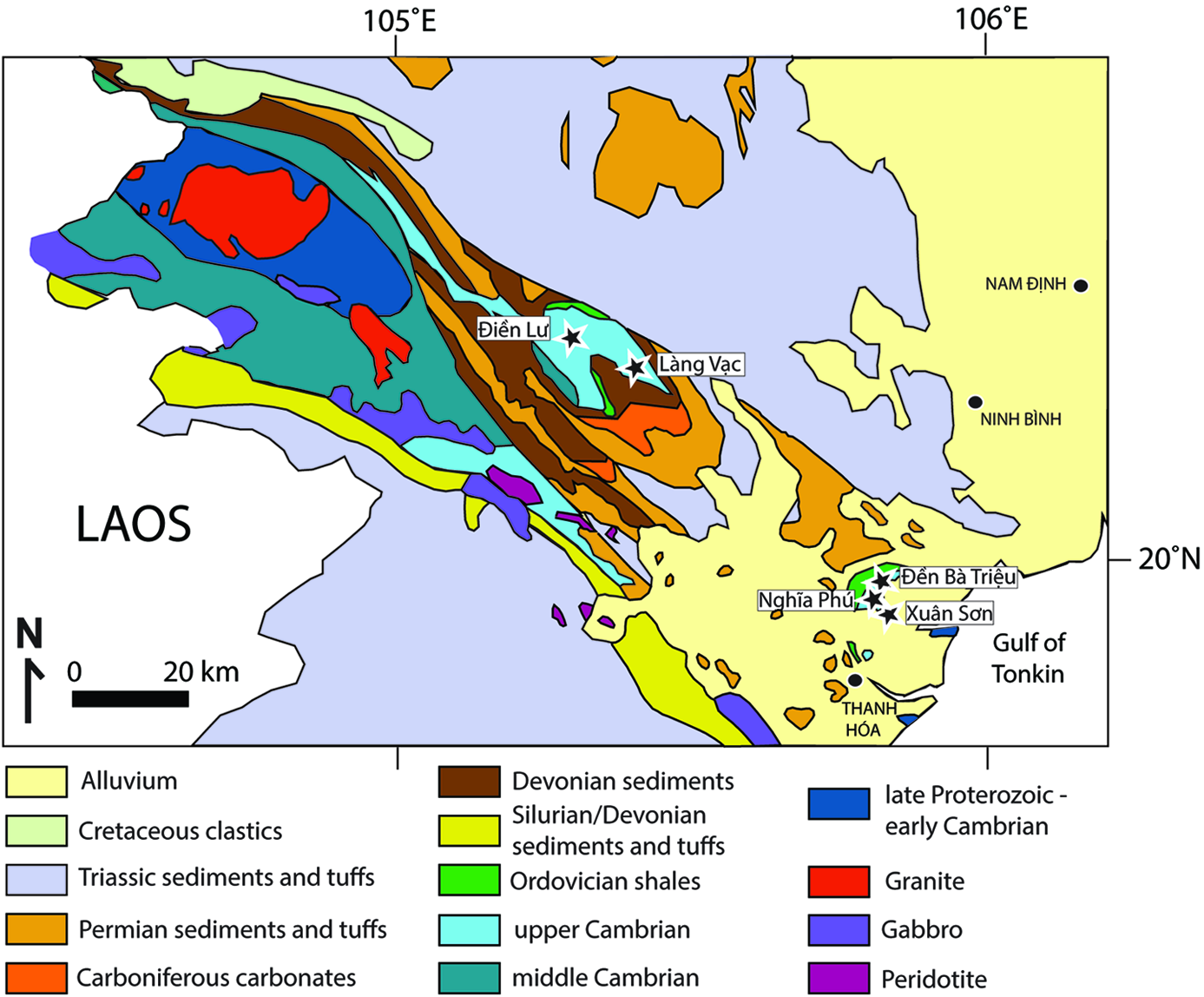
Fig. 2. Geological map of the Sông Mã anticlinorium and suture zone. The suture zone is defined by the gabbroic and peridotitic rocks, and the series of folded and faulted Palaeozoic rocks that make up the core of the Sông Mã anticlinorium. The sites from which fossils are described and discussed herein are identified. The sharp contact with Triassic sediments and tuffs north of Điền Lư marks the Sông Đà fault, considered by some as another terrane boundary. Modified from Trần (Reference Trần1973).
2. Sông Đà geological setting
The Sông Đà terrane is divided into two regions (Fig. 1): a more northerly portion dominated by a swath of Mesozoic rocks that includes a small inlier reportedly bearing Cambrian rocks, and a southern part, the Sông Mã anticlinorium (Findlay & Phan, 1997; Findlay, Reference Findlay1998) (Fig. 2). The border between these areas is the Sông Đà fault, which some consider to be an additional terrane boundary (Khương, Reference Khương2009; Bùi et al. Reference Bùi, Ngô, Khương, Golonka, Nguyễn, Song, Itaya and Yagi2017). All our material comes from the anticlinorium south of the fault, including from several inliers of Cambrian rock occurring laterally along the strike of the anticlinorium, which emerge from the Quaternary cover a few kilometres north of Thanh Hóa city. The geology of the region is complex, with patches of fossil-bearing lower Palaeozoic rock cropping out amidst strongly metamorphosed and sheared rocks of both sedimentary and igneous protolith. The area has been mapped in detail by officers of the Việt Nam Research Institute of Geosciences and Mineral Resources (Trần, 1973) (Fig. 2), but exposure is patchy, making it difficult to establish continuous sections, to assess local folding and faulting, and to give accurate estimates of unit thickness. Reasons for basing our map (Fig. 2) on the 1973 (Trần, 1973) map rather than the 1979 map (Trần, 1979) are given by Findlay & Phan (Reference Findlay and Phan1997, p. 16). Stratigraphic sections and maps of our five fossil-bearing sections are available in Phạm (Reference Phạm2008), whose stratigraphic nomenclature we follow herein. Several of our localities, such as that at Điền Lư, are located adjacent to strongly deformed rocks that make up the Sông Mã suture zone (Findlay & Phan, 1997; Findlay, Reference Findlay1998).
Although maps suggest that the Cambrian crops out extensively in northern Việt Nam (Figs 1, 2), much of the rock mapped as Cambrian is quite highly deformed and unlikely to yield fossils. Assignment of such rocks to the Cambrian is based mainly on stratigraphic relationships and metamorphic history, and requires further verification in many cases. For this reason, we are unsure whether the view of Findlay & Phan (Reference Findlay and Phan1997), that within the Sông Đà terrane there is a sharp contrast between non-metamorphosed and metamorphosed Cambrian rocks, is securely founded.
The lowest unit from which Cambrian fossils have been recovered is the Sông Mã Formation (Phạm, Reference Phạm, Trịnh, Dương, Nguyễn, Lê, Đặng, Nguyễn, Lương, Nguyễn, Nguyễn, Vũ, Nguyễn, Phạm, Nguyễn, Nguyễn, Trần, Nguyễn, Trịnh, Nguyễn and Nguyễn1980), which reportedly sits unconformably above the presumably Proterozoic (Trần, 1979) Nậm Cô Formation (Findlay & Phan, 1997) or its lateral equivalent, the Sa Pa Formation (Fig. 3). The Sông Mã Formation, which may be up to up to 790 m thick in places, is reported to comprise conglomerate, schist and carbonate (Phạm, Reference Phạm2008) and is also said to contain ‘metabasites’ (Findlay & Phan, 1997, p. 14) and ultramafic rocks (Phạm, Reference Phạm2008). Our collections from this formation were from an inlier near Thanh Hóa and from Điền Lư: in both localities only carbonate facies are represented (Fig. 2). The overlying Hàm Rồng Formation, from which many of our samples were collected, is reported to range from 290 to 1150 m thick and is carbonate but may also contain rare sandstone and siltstone intervals (Phạm, Reference Phạm2008). In our excursion to the region, other than extremely thin claystone horizons, the only siliciclastic rocks observed were in the overlying Đông Sơn Formation, although this unit too, is carbonate dominated (Figs 3, 4, 5). Some authors recognize the Điền Lư Formation as a lateral equivalent of the higher parts of the Hàm Rồng Formation and lowest parts of the Đông Sơn Formation (e.g. Phạm & Lương, Reference Phạm and Lương1996) (Fig. 3), but here we follow Phạm (Reference Phạm2008).

Fig. 3. Lithostratigraphic schemes for the Cambrian and Lower Ordovician of the Thanh Hóa region. Fmn. – Formation.

Fig. 4. Chart showing the stratigraphic occurrence of Cambrian and Tremadocian trilobites and brachiopods from the Thanh Hóa region, in relationship to other sections in the region. Stratigraphic placement is based on occurrence data for these trilobites known elsewhere in equatorial Gondwana (Zhou & Zhen, 2008). Only those taxa recognized to generic level or lower are included. Coloured bars represent taxa in our collections. Those with black bars give our tentative identifications from reports published previously (see text for discussion). We show the lithology for the Hàm Rồng Formation only spanning those intervals from which fossils have been collected to date. The continuity of the Cambrian sedimentary record is not well constrained in either the East Bắc Bộ or Thần Sa areas either, which we indicate by a figurative gap in the succession that does not necessarily indicate a hiatus.

Fig. 5. Stratigraphic section of the Hàm Rồng Formation at Đền Bà Triệu, spanning the horizons yielding collection 1.
3. Localities, horizons, biostratigraphy and regional context
3.a. New collections from Thanh Hóa region (Sông Đà terrane)
Outcrops of Cambrian rock around Thanh Hóa city are inliers in which exposed sections are short (Fig. 2), so we relied on biostratigraphical correlation with more continuous sections elsewhere, particularly in China (Zhou & Zhen, 2008), to order those collections whose succession could not be directly determined in the field (Fig. 4). Collections are thus listed in their presumed stratigraphic order of occurrence, from the youngest collections stratigraphically downwards.
3.a.1. Đông Sơn Formation
Làng Vạc 2 (LV-2), 20° 13.782′ N, 105° 22.225′ E; leiostegiid genus and species indet. 1 (for sketch section see Phạm, Reference Phạm2008, p. 58).
Làng Vạc 1 (LV-1), 20° 13.691′ N, 105° 22.376′ E; Troedssonia wimani, leiostegiid genus and species indet. 2, Billingsella sp. cf. B. tonkiniana. Detrital zircon sample LV-1 taken 2 m below trilobite-bearing horizon (for sketch section see Phạm, Reference Phạm2008, p. 58).
3.a.2. Hàm Rồng Formation
Nghĩa Phú (NP-1), 19° 802′ N, 105° 49.165′ E; Eosaukia buravasi, Koldinioidia sp. indet. and Plectotrophia sp. aff. P. imparicostata (for sketch section see Phạm, Reference Phạm2008, p. 56). Other taxa reported but not figured from this horizon include Calvinella walcotti (Phạm, Reference Phạm2008).
Đền Bà Triệu 3 (DBT-3), 19° 55.661′ N, 105° 49.004′ E; Saccagonum sp. indet. Some disarticulated echinoderm ossicles were also recorded at this section. Phạm Kim Ngân’s locality TH16-/8 (for sketch section see Phạm, Reference Phạm2008, p. 55). Approximately 200 m stratigraphically above DBT-2 collection.
Đền Bà Triệu 2 (DBT-2), 19° 55.620′ N, 105° 48.861′ E; Shirakiella guangnanensis, Billingsella sp. cf. B. tonkiniana and Palaeostrophia sp. cf. P. jingensis. Phạm Kim Ngân’s locality TH16-/5 (for sketch section see Phạm, Reference Phạm2008, p. 55). Approximately 100 m stratigraphically above DBT-1 collection.
Đền Bà Triệu 1 (DBT-1) (see Fig. 5), 19° 55.567′ N, 105° 48.861′ E; Prosaukia sp. indet., Shergoldia sp. cf. S. trigonalis and Palaeostrophia sp. cf. P. jingensis. Phạm Kim Ngân’s locality TH16-/1 (for sketch of entire section see Phạm, Reference Phạm2008, p. 55). Other taxa reported but not figured from this horizon include Pseudokoldinia sp., Blountia? sp. and Billingsella? sp. (Phạm, Reference Phạm2008).
3.a.3. Sông Mã Formation
Điền Lư, on north side of road to Bá Thước (DL-1), 20° 18.431′ N, 105° 16.992′ E; Proasaphiscus latifrons (for sketch section see Phạm, Reference Phạm2008, p. 58). Phạm (Reference Phạm2008) mentioned Anomocarina?, Blackwelderia sinensis, Cyclolorenzella tonkinensis, Damesella brevicaudata and Neodrepanura sp.
Xuân Sơn (XS-1), 19° 54.399′ N, 105° 50.638′ E; Kaotaia xuanensis sp. nov., Solenoparops sp. indet., eostrophiid brachiopod (for sketch section see Phạm, Reference Phạm2008, p. 56).
3.b. Other relevant trilobite reports from Thanh Hóa region (Sông Đà terrane)
In the earliest report of lower Palaeozoic fossils from the Thanh Hóa region, Mansuy (Reference Mansuy1920) reported a trilobite comparable to the dikelocephalinid Asaphopsis and considered it of Tremadocian age (Jacob, Reference Jacob1921), but formal descriptions have been sparse. Stokes (Reference Stokes2008) reported this fauna to be from Indochina, but it is part of the Sông Đà terrane. Asaphopsis villebruni, Pseudokainella sp., Bienvillia (Desmetia) sp. and Leiobienvillia sp. were also mentioned but not described or figured by Phạm (Reference Phạm2008) in his treatise on the Cambrian Geology of Việt Nam. They were reported as occurring in the upper parts of the Đền Bà Triệu section, stratigraphically above our collection 3. Asaphopsis jacobi, Asaphopsis immanis and Annamitella asiatica were also reported from the Hàm Rông – Đông Sơn section (Phạm, Reference Phạm2008).
Phạm (Reference Phạm2008) illustrated some additional taxa from Cambrian rocks in the Thanh Hóa area. The determinations of these are discussed below.
3.b.1. Hàm Rồng Formation
Bá Thước: cranidium and pygidium assigned to Dikelocephalus sp. (see Phạm, Reference Phạm2008, p. 194, pl. 12, fig. 1). The figured cranidium has bacculae and is clearly dikelocephalinid, not dikelocephalid. It is strikingly similar to Monkaspis. The co-occurring pygidium is very large, and while it could be dikelocephalinid, it lacks marginal spines that characterize this group. If it belongs to another taxon, a candidate may lie within Paracoosia.
Điền Lư: cranidium of Blackwelderia sinensis (see Phạm, Reference Phạm2008, p. 191, pl. 4, fig. 3). As the palpebral lobes of this specimen are missing it could belong to either Blackwelderia or Parablackwelderia (e.g. Peng et al. Reference Peng, Babcock and Lin2004 a, pls 28–30).
3.b.2. Sông Mã Formation
Điền Lư: cranidium of Metanomocare grandiformis n. sp. (see Phạm, Reference Phạm2008, p. 191, pl. 4, fig. 4). The figured counterpart cranidium resembles that figured by Lermontova (Reference Lermontova, Vologdin, Lermontova, Yavorsky and Janischevsky1940, pl. 47, fig. 5) as Metanomocare petaloides, but is also comparable to Paracoosia and Afghanocare (see Peng et al. Reference Peng, Babcock and Lin2004 a, pls 47, 48).
Tuần Giáo, Điện Biên: cranidium and pygidium of ‘Yohoaspis phadinensis’ Phạm (Phạm, Reference Phạm2008, p. 194, pl. 10, figs 4, 5). Two strongly deformed specimens, a cranidium and a pygidium, were used by Phạm Kim Ngân to erect this species, but no systematic description was provided and we consider this species a nomen nudum. The proportions of the cranidium encourage comparison with those ptychopariids bearing relatively long glabellae and short frontal areas, such as Nangaoia (e.g. Yuan et al. Reference Yuan, Zhao, Li and Huang2002, pl. 42).
An echinoderm plate referred to Pleurocystites? was figured from the Hàm Rồng Formation at Làng Vạc (Phạm, Reference Phạm2008, p. 192, pl. 5, fig. 7).
4. Sedimentology
The measured section at the site of Đền Bà Triệu collection 1 (Fig. 5) is dominantly fine grainstone with a few slightly coarser grainstone beds and scattered shale beds up to 36 cm thick. The fine grainstone is mostly thinly bedded with a few blocky weathering beds up to 75 cm thick, some with parallel lamination. The internal sedimentary structures are in many cases difficult to discern, but the most common is parallel lamination. A few beds between 11 and 14 m in the section contain angle-of-repose cross-bedding up to 18 cm thick. Additional sedimentary structures include intraclast-rich beds, ball-and-pillow structures (at 12.7 m) and a few beds with quasi-planar lamination and hummocky cross-stratification in the upper 6 m of the section.
The quasi-planar lamination and hummocky cross-stratification are a record of storm deposition in relatively shallow water under the influence of either complex oscillatory flow or combined flows with current and wave components (Southard, Reference Southard1991; Arnott, Reference Arnott1993; Myrow & Southard, Reference Myrow and Southard1996; Dumas et al. Reference Dumas, Arnott and Southard2005). The abundant parallel lamination records slightly higher velocity flows in upper plane bed conditions. The intraclasts are also consistent with high-energy storm deposition, as they represent rip-ups of consolidated to early cemented surficial sediment. The relative scarcity of shale beds suggests that the depositional environment was close to shore, i.e., in the lower shoreface to transition zone into the proximal offshore region.
5. Taphonomy
The Thanh Hóa carbonate specimens have a peculiar taphonomy in that they are all preserved in dolomite, a lithology in which fossil form is usually destroyed during diagenesis. Hammering the Thanh Hóa Cambrian dolomite of the Sông Mã and Hàm Rồng formations fails to ‘crack out’ fossils, of which there are no obvious traces in thin-section. However, owing to intense monsoonal weathering, dolomite has locally rotted into loosely consolidated rhombs that occur along seams separating better consolidated bedsets. When these rotted rocks are gently pried apart, composite moulds of fossils re-emerge in some cases. As the dolomite rhombs are no longer cemented together, these fossils are so fragile that they can by smeared by touch alone. Hence, upon recovery specimens were immediately consolidated with an adhesive. We used Butvar and acetone when available, and Elmer’s/School Glue diluted with water when not. Specimens preserved in this way more closely resemble those preserved in sandstone, rather than carbonate, in that they have three-dimensional relief, but the shell is entirely lost. Because there was no void preserved where the trilobite exoskeleton or brachiopod valves dissolved, the fossils are akin to composite moulds, although it is unclear whether they combine the features of internal and external surfaces: preservation is too coarse to determine features at this scale.
This peculiar taphonomic mode has implications for the morphology preserved. Firstly, fine details of the skeletal surface texture structure, such as pustulation, terracing or other ornament, are not preserved. This limits our ability to recognize those taxa in which ornament plays an important role in taxonomy, such as among the dikelocephalid trilobites. Secondly, as the fossils must be consolidated immediately to prevent destruction, there is little opportunity to prepare the fossils to reveal unexposed parts. These factors limit the quality of the material that can be recovered and thus limited our ability to make systematic determinations based on delicate features. Lastly, most of these fossils have also undergone tectonic deformation but, because we lacked slabs with multiple deformed specimens and evidence of the principal extension direction is scarce, retrodeformation was attempted only for a single specimen (Fig. 12).
6. Systematic palaeontology
The trilobite taxonomy is by Xuejian Zhu, Nigel Hughes, Shanchi Peng and Shelly Wernette, and the new taxon should be attributed to those authors. The brachiopod taxonomy is by David Harper and Nigel Hughes. Specimens are housed in the type collection of the Bảo Tàng Địa Chất, the Department of Geology and Mineral Resource’s Geological Museum, no. 6, Phạm Ngũ Lão, Hà Nội, Việt Nam, under the specimen prefix ‘BT’. Non-type material is held in the collections of the Cincinnati Museum Center.
Family Ptychopariidae Matthew, Reference Matthew1887
Genus Kaotaia Lu in Lu et al. Reference Lu, Zhu, Qian and Wang1962
Type species. Alokistocara magnum Lu, Reference Lu1945, from the Kaotai Formation, Guizhou, South China.
Discussion. This genus has been discussed at length in Peng et al. (Reference Peng, Hughes, Heim, Sell, Zhu, Myrow and Parcha2009).
Kaotaia xuanensis sp. nov.
Figure 6c–f, h, j, m

Fig. 6. Trilobite sclerites from the Sông Mã Formation at Xuân Sơn. All specimens are coated with ammonium chloride sublimate prior to digital photography; specimens are internal moulds unless otherwise stated. All specimens cranidia except (h, j, m) which are pygidia. Scale bars: (a) = 1 mm; (b, c, e–m) = 2 mm; (d) = 4 mm. (a) Solenoparops sp. indet., latex cast of external mould, BT1/598. (b) Solenoparops sp. indet., BT2/598. (c) Kaotaia xuanensis new species, holotype, BT3/598. (d) Kaotaia xuanensis, paratype, BT4/598. (e) Kaotaia xuanensis, paratype, BT5/598. (f) Kaotaia xuanensis, paratype, latex cast of external mould, BT6/598. (g) Solenoparops sp. indet., latex cast of external mould, BT7/598. (h) Kaotaia xuanensis paratype, BT8/598. (i) Solenoparops sp. indet., BT9/598. (j) Kaotaia xuanensis, paratype, BT10/598. (k) Solenoparops sp. indet., BT11/598. (l) Solenoparops sp. indet., BT12/598. (m) Kaotaia xuanensis, paratype, BT13/598.
1980 Inouyia sp. Phạm, p. 46, pl. 1, fig. 1.
2008 Inouyia sp. Phạm, pl. 4. fig. 2.
Etymology. For Xuân Sơn, the type locality in Thanh Hóa province.
Types. Holotype: internal cranidial mould (BT3/598). Paratypes include four cranidia (BT4/598–BT6/598, BT10/598) and two pygidia (BT8/598, BT13/598). Also BT1/581 (Phạm, Reference Phạm2008, pl. 4. fig. 2). Non-figured topotypes: four cranidia (CMCIP87732–87735).
Diagnosis. Kaotaia with proportionally wide (tr.) fixigena and short (sag.) anterior border; anterior border furrow shallow.
Description. Cranidium subrectangular in large holaspids, wider than long, moderately convex, transverse anteriorly. Glabella short, subtriangular, moderately convex, tapering forward, truncate or evenly rounded anteriorly, occupying ∼0.6 of cranidial length in smaller holaspids, ∼0.55 in larger holaspids, bearing three pairs of evenly spaced, distinct but weakly incised lateral furrows; S1 with inner half more oblique than outer half; S2 and S3 transverse; occipital furrow shallow, transverse, deepened abaxially; occipital ring crescentic. Axial furrow firmly incised in smaller holaspids. Anterior border short (sag.), flat or gently convex, shortening slightly abaxially; anterior border furrow weakly incised, gently curved, preglabellar field more than twice length of anterior border (sag.). Preglabellar boss distinct in larger holaspids, absent in smaller ones; eye ridge gently arching forward then rearward abaxially, following fixigenal slope, with adaxial end opposite glabellar anterolateral margin; palpebral lobe narrow (tr.), slightly curved, slightly oblique to sagittal line, with posterior end opposite S1 and anterior end opposite S3 in large holaspids, proportionately longer in small holaspids. Anterior branch of facial suture diverging forward slightly at an angle of 0–10 degrees from sagittal axis to anterior border furrow, turning inward and forward to cross anterior border in a long curve; posterior branch divergent rearward defining triangular posterolateral border; posterior border furrow short (exsag.), shallow and defining transverse posterior border to fulcrum then curving steeply ventrally and rearward. Pygidium elliptical with two clearly defined axial rings plus one weakly incised ring and rounded terminal piece. Two pairs of pleural furrows and one pair of interpleural furrows extend almost to border.
Discussion. Assignment of this broad form to Kaotaia is based on the presence of the distinct preglabellar boss. The new species is readily distinguished from other species in the genus by its proportionally wide fixigena and its short anterior border. Among Kaotaia, the anterior border of K. xuanensis is most similar to that of K. yongshanensis Luo in Luo et al. Reference Luo, Jiang and Tang1994, but that species has inflated eye ridges and notably narrower fixigenae. The specimen figured by Phạm (Reference Phạm, Trịnh, Dương, Nguyễn, Lê, Đặng, Nguyễn, Lương, Nguyễn, Nguyễn, Vũ, Nguyễn, Phạm, Nguyễn, Nguyễn, Trần, Nguyễn, Trịnh, Nguyễn and Nguyễn1980, pl. 1, fig. 1), although convergent with Inouyia, belongs within K. xuanensis because of its subtriangular glabella. The glabella of Inouyia capax, the type species (see Zhang & Jell, Reference Zhang and Jell1987, p. 50, fig. 3.6), is quadrate. Inouyia capax also lacks the prominent anterior border furrow seen in K. xuanensis and other Kaotaia.
Sông Đà occurrence. In the dolomitic Sông Mã Formation. At Xuân Sơn, co-occurring with Solenoparops sp. indet., also from Nghĩa Trang, Hoằng Hóa (Phạm, Reference Phạm2008, p. 191, pl. 4, fig. 1) together with an eostrophiid brachiopod.
Family Solenopleuridae Angelin, Reference Angelin1854
Genus Solenoparops Zhang, 1963
Type species. Solenoparia luna Endo, Reference Endo1944 (= S. taitzuensis Resser & Endo, Reference Resser, Endo, Endo and Resser1937).
Solenoparops sp. indet.
Figure 6a, b, g, i, k, l
Material. Figured: Six cranidia: BT1/598, BT2/598, BT7/598, BT9/598, BT11/598, BT12/598. Unfigured material two cranidia (CMCIP87736–87737) and possibly a third (CMCIP87738).
Discussion. The shape of the glabella and short preglabellar field are very similar to Solenoparops granulus Zhang in Lu et al. Reference Lu, Zhang, Zhu, Qian and Xiang1965, which is a junior homonym of Solenoparops granulus (Endo, Reference Endo, Endo and Resser1937) nom. corr. herein (pro Solenoparops granulosa (Endo, Reference Endo, Endo and Resser1937), comb. nov. Zhang in Lu et al. Reference Lu, Zhang, Zhu, Qian and Xiang1965). The latter has line priority over Solenoparops granulus Zhang in Lu et al. Reference Lu, Zhang, Zhu, Qian and Xiang1965. In our material the anterior border is proportionally longer (sag.) than that of S. granulus sensu Zhang in Lu et al. Reference Lu, Zhang, Zhu, Qian and Xiang1965, and it is easily distinguished from that species. As the specimens are badly preserved and few in number, we leave this form in open nomenclature.
Sông Đà occurrence. In the dolomitic Sông Mã Formation at Xuân Sơn, co-occurring with Kaotaia xuanensis sp. nov.
Family Proasaphiscidae Zhang, 1963
Genus Proasaphiscus Resser & Endo in Kobayashi, Reference Kobayashi1935
Type species. Proasaphiscus yabei Resser & Endo in Kobayashi, Reference Kobayashi1935
Proasaphiscus latifrons (Mansuy, Reference Mansuy1916)
Figure 7a–r

Fig. 7. Proasaphiscus latifrons from the Sông Mã Formation at Điền Lư, on north side of road to Bá Thước. All specimens are internal moulds and were coated with ammonium chloride sublimate prior to digital photography. Scale bars: all 5 mm except (e, i, k) = 2.5 mm. (a–j) cranidia; (l, m, p–r) pygidia. (a) BT1/599; (b) BT2/599; (c) BT3/599; (d) BT4/599; (e) BT5/599; (f) BT6/599; (g) BT7/599; (h) BT8/599; (i) BT9/599; (j) BT10/599; (k) hypostome, BT11/599; (l) BT12/599; (m) BT13/599; (n) left free cheek, BT14/599; (o) left free cheek, BT15/599; (p) BT16/599; (q) BT17/599; (r) BT18/599.
Material. Figured: ten cranidia (BT1/599–BT10/599), one hypostome (BT11/599), two free cheeks (BT14/599, BT15/599) and five pygidia (BT12/599, BT13/599, BT16/599–BT18/599). Unfigured: seven cranidia (CMCIP87742–87748), four free cheeks (CMCIP87749–87752) and 13 pygidia, one of which is a counterpart (CMCIP87753–87765).
Discussion. This morphotype with a long and wide anterior border and preglabellar field and a tapering glabella appeared iteratively in several Cambrian groups, but we assign these specimens to Proasaphiscidae based on the relative proportions of the glabella, fixigenae and anterior border; the large eyes and the form of the pygidium exclude it from early ptychopariid homeomorphs. The anterior border is striking for its length, inflation and firmly incised border furrow. The cranidium resembles that of Huayuanaspis Peng et al. Reference Peng, Babcock and Lin2004 b but lacks the occipital spine diagnostic of that genus. The cranidium is also closely similar to that of several Manchuriella species (see Guo et al. Reference Guo, Zan and Luo1996, pl. 52, figs 11–14) and also to Kuruktagella laevigata Zhang, 1981, although that genus was considered by its author to belong in Pterocephalidae. Some cranidia from southeastern Yunnan assigned by Luo et al. (Reference Luo, Hu, Hou, Gao, Zhan and Li2009) to Proasaphiscus latifrons (Mansuy, Reference Mansuy1916) are so similar to our material that we consider them to be conspecific pending the results of ongoing revision of Mansuy’s type material and documentation of the intraspecific variation encompassed therein.
Sông Đà occurrence. In highly weathered dolomite of the Sông Mã Formation on a dip slope near Điền Lư (DL-1), on north side of road to Bá Thước. This species is also recorded in the North Việt Nam block/southeastern Yunnan (Mansuy, Reference Mansuy1916; Luo et al. Reference Luo, Hu, Hou, Gao, Zhan and Li2009).
Family Shirakiellidae Kobayashi, Reference Kobayashi1935
Genus Shirakiella Kobayashi, Reference Kobayashi1935
Type species. Shirakiella elongata Kobayashi, Reference Kobayashi1935.
Shirakiella guangnanensis Luo in Luo et al. Reference Luo, Hu, Hou, Gao, Zhan and Li2009
Figure 8a–e

Fig. 8. Shirakiella guangnanensis cranidia from the Hàm Rồng Formation at Đền Bà Triệu, collection 2. All specimens are internal moulds and were coated with ammonium chloride sublimate prior to digital photography. White scale bars are 5 mm long. (a) BT1/600; (b) BT2/600; (c) BT3/600; (d) BT4/600; (e) BT5/600.
2008 Blountia? Phạm, p. 194, pl. 10, fig. 3.
Material. Five cranidia from Đền Bà Triệu collection 2 (BT1/600–BT5/600).
Discussion. It is likely that S. guangnanensis is itself a synonym of an earlier described species, and probably of the type species, S. elongata. Formal synonymy is hindered by the fact that Kobayashi’s 1935 plates do not show the frontal area clearly, which, in some specimens preserved in limestone assigned to this species by Qian (Reference Qian1994, pl. 13, figs 1, 2), show a very short cranidial anterior border and border furrow, related to retraction of the facial suture from the anterior margin of the cephalon. These structures are not evident in either S. guangnanensis or in our specimens, but this could be due to poor preservation. The specimen illustrated by Phạm (Reference Phạm2008) may be laterally compressed but closely resembles that in BT1/600. BT2/600 is a poor specimen doubtfully assigned to this species, and may have a large occipital spine. A specimen from Bá Thước referred to Quadraticephalus? by Phạm (Reference Phạm2008, p. 192, pl. 7, fig. 2) resembles S. guangnanensis in a general way but apparently has wider fixigenae and a longer frontal area.
There is a possibility that these cranidia, which are the only fossils in our collection from this locality, represent a proceratopygid. Such an assignment would accord better with their inferred stratigraphic occurrence above beds containing Shergoldia cf. S. nomas and Prosaukia sp. The general form of the cranidium does resemble taxa such as Proceratopygye (e.g. Lu & Lin, Reference Lu and Lin1989, pl. 24, fig. 12), but we prefer assignment to Shirakiella because of the far forward placement of the relatively small eyes and the lack of the preoccipital tubercle.
Phạm (Reference Phạm2008, pl. 10, fig. 3) referred a similar cranidium to Blountia? The overall shape of the cranidium and glabella resemble that of this genus, but a notable difference is the absence of an anterior border and anterior border furrow in the Vietnamese specimens. Such features are characteristic of Blountia (see Walcott, Reference Walcott1916; Pratt, Reference Pratt1992), and so this material is excluded from that genus.
Sông Đà occurrence. In the dolomitic Hàm Rồng Formation collection 2 at Đền Bà Triệu. Also at locality TH16-/5, Hoằng Hóa (Phạm, Reference Phạm2008, p. 194, pl. 10, fig. 3). This species is also recorded in the North Việt Nam block (Luo et al. Reference Luo, Hu, Hou, Gao, Zhan and Li2009).
Family Saukiidae Ulrich & Resser, Reference Ulrich and Resser1930
Genus Prosaukia Ulrich & Resser, Reference Ulrich and Resser1930
Type species. Dikelocephalus misa Hall, Reference Hall1863 from the Lone Rock Formation, Wisconsin, USA.
Prosaukia sp. indet.
Figure 9b–d, f–i

Fig. 9. Trilobite sclerites from the Hàm Rồng Formation at Đền Bà Triệu, collection 1. All specimens are coated with ammonium chloride sublimate prior to digital photography; specimens are internal moulds unless otherwise stated. White scale bars are 5 mm long except (f-h) = 2.5 mm. (a) Shergoldia cf. S. trigonalis, cranidium, BT1/601. (b) Prosaukia sp., cranidium, BT2/601. (c) Prosaukia sp., cranidium, BT3/601. (d) Prosaukia sp., cranidium, BT4/601. (e) Shergoldia cf. S. trigonalis, pygidium, BT5/601. (f) Prosaukia sp., latex of counterpart pygidium, BT6b/601. (g) Prosaukia sp., pygidium, BT7/601. (h) Prosaukia sp., pygidium, BT8/601. (i) Prosaukia sp., left free cheek, BT9/601. (j) Shergoldia cf. S. trigonalis, pygidium, BT10/601. (k) Shergoldia cf. S. trigonalis, pygidium, BT11/601. (l) Shergoldia cf. S. trigonalis, pygidium, BT12/601.
Material. Figured: Three cranidia (BT2/601–BT4/601), one free cheek (BT9/601) and three pygidia (BT6b/601–BT8/601). Unfigured: four cranidia (CMCIP87766–87770) and three pygidia (CMCIP8771–87773).
Discussion. Specimens are deformed and quite poorly preserved. The anterior border is separated from the glabella by a short but distinct preglabellar field and by the anterior border furrow which shallows adaxially. While a preglabellar field is present in various dikelocephalids, its relative shortness (sag.) and the axial shallowing of the border furrow are characteristic of the widespread genus Prosaukia (e.g. Zhang et al. Reference Zhang, Xiang, Liu and Meng1995, p. 84, pl. 37, figs 8–10) as are the broad and anteriorly tapering glabella and the anteriorly divergent facial suture branches. Hoytaspis Ludvigsen & Westrop, Reference Ludvigsen and Westrop1983, known from Laurentia and Sibumasu (Ludvigsen & Westrop, Reference Ludvigsen and Westrop1983; Shergold et al. Reference Shergold, Burrett, Akerman and Stait1988), has a similar overall form, but our material differs from this genus owing to Hoytaspis having a more parallel-sided glabella, defined S2 and strongly pustulose sculpture. The Thanh Hóa material resembles North American species of Prosaukia in the location of the eye, the midpoint of which is opposite the anterior part of L1 and not further forward as in other Gondwanan, short preglabellar field–bearing dikelocephalids such as Caznaia (see Shergold, Reference Shergold1975), Andersonella (see Shergold, Reference Shergold1991) and Hoytaspis? thanisi Shergold et al. (Reference Shergold, Burrett, Akerman and Stait1988). Some Chinese Prosaukia also have more forward positioned eyes (e.g. P. rotundolimbata Endo & Resser, Reference Endo and Resser1937 and P. resseri Endo & Resser, Reference Endo and Resser1937), although Sibumasu’s Prosaukia have eyes opposite or slightly posterior to S1 (Wernette et al. Reference Wernette, Hughes, Myrow and Sardsud2020 a) as in the Laurentian forms. The Thanh Hóa pygidia are transversely semi-elliptical in outline with three axial rings plus a terminal piece and pleurae that are apparently equally divided. Prosaukia misa, the generic type species known from the lower Sunwaptan of North America, differs in having four axial rings, a postaxial ridge and a wider and especially longer pleural region, but differences of this magnitude are common within dikelocephalid genera. Among other Gondwanan Prosaukia, only Prosaukia angulata (Mansuy, Reference Mansuy1916, p. 34, pl. 5, fig. 12c) may possess fewer than four axial rings, but this is unclear from the available published material; the fourth axial ring of Thailand’s Prosaukia tarutaoensis (Kobayashi, Reference Kobayashi1957) is very poorly defined but is part of a longer, narrower axis than that of Prosaukia sp. herein (Wernette et al. Reference Wernette, Hughes, Myrow and Sardsud2020 a). While Eosaukia has a similarly transverse, paucisegmented pygidium, with sometimes as few as two segments (Kobayashi, Reference Kobayashi1957), we are confident this pygidium is not misassociated, because no Eosaukia cranidia have been recovered at the Đền Bà Triệu locality and Eosaukia has a pair of ridges on its terminal piece, of which there is no evidence in the specimens considered here.
Prosaukia angulata (Mansuy, Reference Mansuy1915; see Lu in Lu et al. Reference Lu, Zhang, Zhu, Qian and Xiang1965) is also from Việt Nam, but from rocks located north of the Sông Mã fault. Despite their similarly transverse and paucisegmented pygidia, the material herein is unlikely to be P. angulata owing to its long and robust occipital spine, which is unknown in previously figured specimens of P. angulata, though P. angulata var. chinensis (Sun, Reference Sun1924) may possess a small occipital node (Lu et al. Reference Lu, Zhang, Zhu, Qian and Xiang1965, fig. 21). The free cheek of P. angulata possesses well-defined lateral and posterior border furrows that merge adaxially at the base of the genal spine. The furrows on the associated librigena from Thanh Hóa (Fig. 9i) are relatively shallow and more closely parallel the posterior border.
Sông Đà occurrence. In the dolomitic Hàm Rồng Formation in collection 1 from Đền Bà Triệu co-occurring with Shergoldia cf. S. trigonalis.
Eosaukia Lu, Reference Lu1954
Type species. Eosaukia latilimbata Lu, Reference Lu1954 from the Sandu shale, Guizhou, China.
Eosaukia buravasi Kobayashi, Reference Kobayashi1957
Figure 10a–l
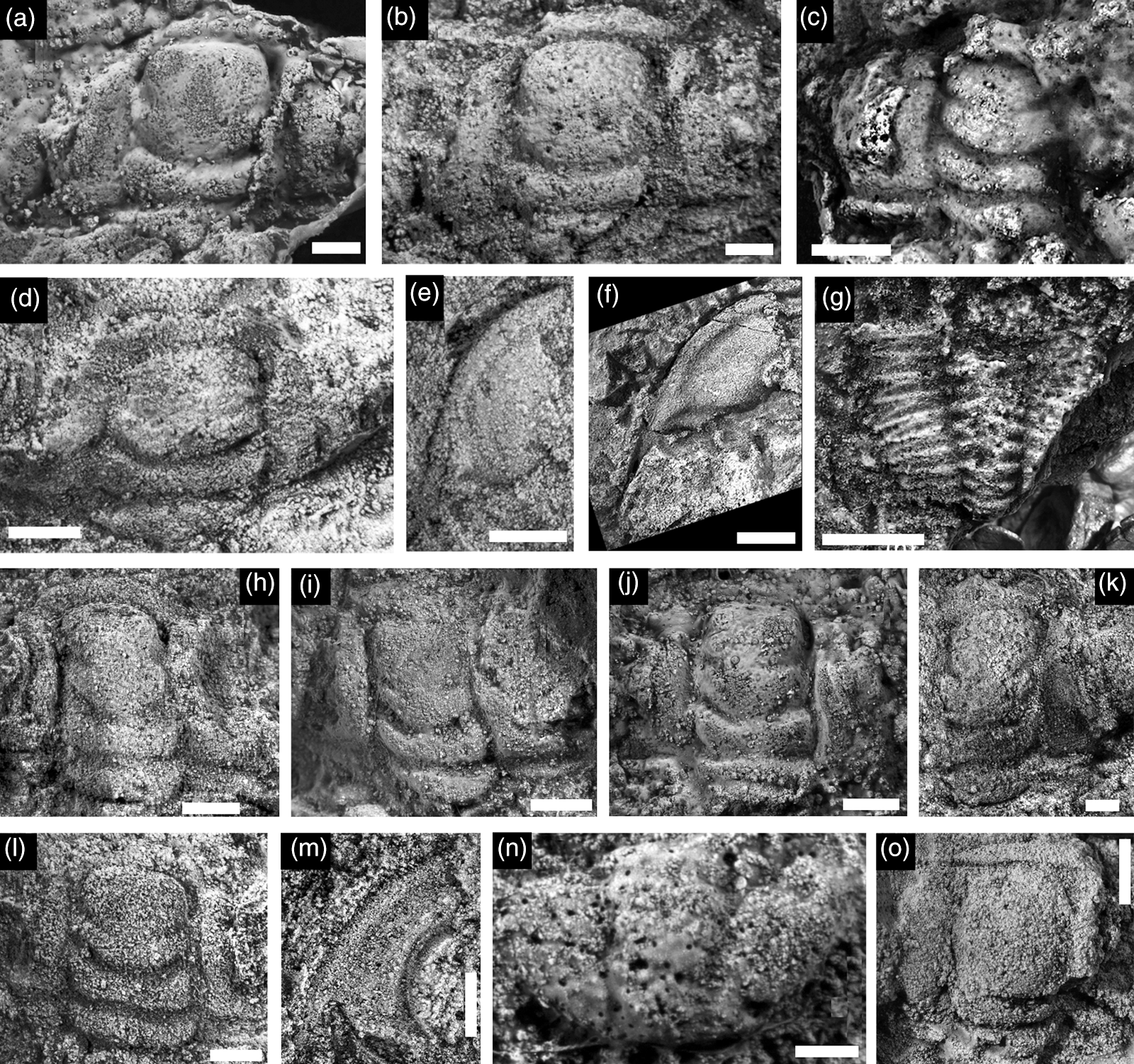
Fig. 10. Trilobite sclerites from the Hàm Rồng Formation at Nghĩa Phú and from the Đông Sơn Formation at Làng Vạc locality 2. All specimens are internal moulds unless otherwise stated and are coated with ammonium chloride sublimate prior to digital photography. Scale bars (a–c) = 2 mm, (d–o) = 4 mm. All Eosaukia buravasi cranidia from Nghĩa Phú unless otherwise stated: (a) BT1/602; (b) BT2/602; (c) BT3/602; (d) BT4/602; (e) left free cheek, BT5/602; (f) left free cheek, BT6/602; (g) thorax BT7/602; (h) BT8/602; (i) BT9/602; (j) BT10/602; (k) BT11/602; (l) BT12/602. (m) Haniwa? sp. indet., left free cheek, BT13/602. (n) Koldinioidia sp. indet., pygidium, BT14/602. (o) Leiostegiid genus and species indet. 1, cranidium, Đông Sơn Formation, Làng Vạc locality 2, BT15/602.
1957 ‘Eosaukia’ buravasi Kobayashi, p. 376, pl. 5, figs 1–6, 10, 14–20, ?7–9, ?13.
1988 ‘Eosaukia’ buravasi Kobayashi; Shergold et al., p. 310, fig. 4O–X.
?2007 Ptychaspis? sp. aff. P. cacus (Walcott, Reference Walcott1905); Shergold et al., p. 65, fig. 38.
2008 Calvinella walcotti Mansuy; Phạm, p. 194, pl. 10, fig. 1.
Material. Figured: Nine cranidia (BT1/602–BT4/602, BT8/602–BT12/602), two free cheeks (BT5/602, BT6/602) and one partially articulated trunk (BT7/602). Unfigured: seven cranidia (CMCIP87781–87787) and two free cheeks (CMCIP87788–87789), plus one cranidium from Phạm (Reference Phạm2008, pl. 10, fig. 1).
Discussion. There are two morphological end-members contained within this collection, demonstrating variety in the width of fixed cheeks, width of the glabella, the extent to which SO undulates medially, the incision of S2 and the presence and size of an occipital node. Some specimens (e.g. Fig. 10c) exhibit intermediate character states, suggesting that this variation is intraspecific. It is not strongly size-related, but it may be at least partially controlled by deformation as variants Figure 10a and 10c are orthogonal to each other on the same slab and show different forms.
The variation within this collection is a subset of the variation within Eosaukia buravasi collections found in the Tarutao Group of Thailand. Eosaukia buravasi resembles Eosaukia bella (Walcott, Reference Walcott1906) in many respects, but E. bella possesses a more strongly medially shortened (sag.) anterior border and more curved palpebral lobes. Eosaukia micropora (Qian, Reference Qian1985) is also similar, but there is no evidence in the Việt Nam material of E. micropora’s exceptionally robust occipital spine (Lee & Choi, Reference Lee and Choi2011). The longer, narrower cranidia resemble Mictosaukia in all but the anterior divergence of the abaxial anterior border. Recovery of pygidia in the Sông Đà Formation may help in further refining taxonomic affinity.
Sông Đà occurrence. This species occurs in some abundance in the dolomitic Hàm Rồng Formation in a quarry at Nghĩa Phú along with Koldinioidia sp. indet. Also in same formation at Trinh Hà, Hoằng Hóa (Phạm, Reference Phạm2008, pl. 10, fig. 1).
Family Shumardiidae Lake, Reference Lake1907
Genus Koldinioidia Kobayashi, Reference Kobayashi1931
Type species. Koldinioidia typicalis Kobayashi, Reference Kobayashi1931 from the Fengshan Formation, Liaoning, NE China.
Koldinioidia sp. indet.
Figure 10n
Material. Figured: A single pygidium (BT14/602).
Discussion. The outline of the pygidium is reminiscent of that of Koldinioidia orientalis (Mansuy, Reference Mansuy1916) (see Zhu & Peng, Reference Zhu and Peng2006). The axis tapers rearward more slowly and is consequently proportionally wider than that of K. orientalis, which distinguishes this specimen from that species. As this is only a single, poorly preserved pygidium we do not assign it to any species.
Sông Đà occurrence. In the Hàm Rồng Formation at Nghĩa Phú quarry along with Eosaukia buravasi.
Family Remopleurididae Hawle & Corda, Reference Hawle and Corda1847
Genus Haniwa Kobayashi, Reference Kobayashi1933
Type species. Haniwa sosanensis Kobayashi, Reference Kobayashi1933 from the Tsinania Zone of the Chosan (= Sosan) area, Korea. Fengshan Formation, Liaoning, NE China.
Haniwa? sp. indet.
Figure 10m
Material. Figured: a single librigena (BT13/602).
Discussion. This librigena has a very large, strongly arched eye, advanced genal spine and narrow genal field, conditions common among species of Haniwa. However, without being able to determine whether the librigena is yoked and without an associated Haniwa cranidium, we leave the generic assignment as tentative. The placement of the genal spine is more similar to that of Haniwa quadrata Kobayashi, Reference Kobayashi1933 than to the more advanced spine of Haniwa sosanensis Kobayashi, Reference Kobayashi1933 (see Sohn & Choi, Reference Sohn and Choi2007 and Park & Choi, Reference Park and Choi2011).
Sông Đà occurrence. In the Hàm Rồng Formation at Nghĩa Phú quarry along with Eosaukia buravasi.
Family Leiostegiidae Bradley, Reference Bradley1925
Two cranidia from different but possibly stratigraphically equivalent sites at Làng Vạc show the distinctive structure of the anterior of the glabella and cranidial anterior border that are characteristic of this family.
Leiostegiid genus and species indet. 1
Figure 10o
Material. Figured: a single fragmentary cranidium (BT15/602).
Discussion. This cranidium strongly resembles Pseudocalymene szechuanensis (Lu in Lu et al. Reference Lu, Zhu, Qian and Wang1962; also see Liu in Zhou et al. Reference Zhou, Lui, Meng and Sun1977, pl. 56, fig. 15), and might be conspecific with it. However, Pseudocalymene Pillet, Reference Pillet1973 is distinguished from Chosenia Kobayashi, Reference Kobayashi1934 by the absence of spines on the pygidium. Given that we have no pygidium, at present we prefer to leave this specimen in open nomenclature.
Sông Đà occurrence. Locality Làng Vạc collection 2 in Hàm Rồng Formation float material collected just west of Làng Vạc village.
Leiostegiid genus and species indet. 2
Figure 11a
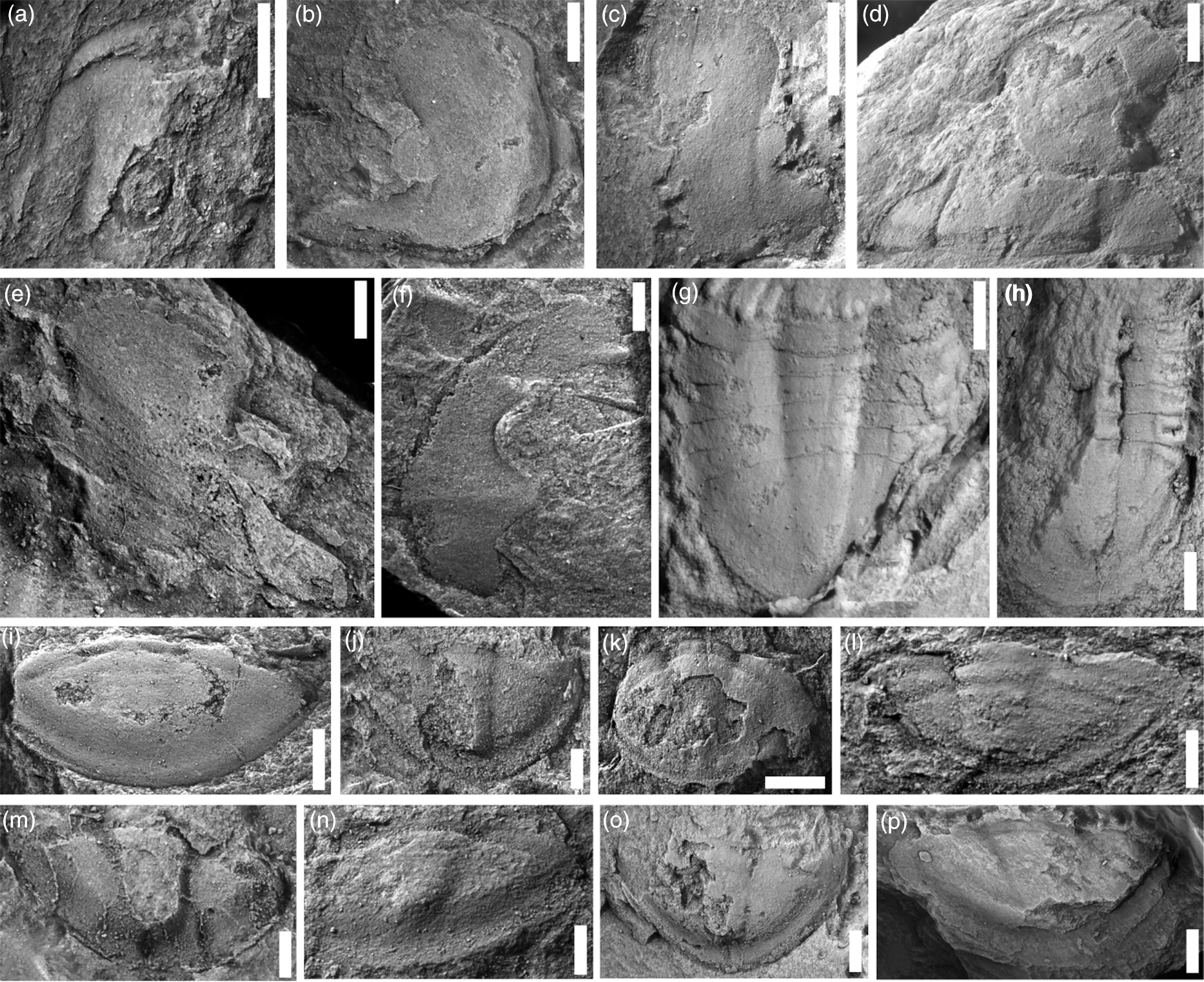
Fig. 11. Trilobite sclerites from the Đông Sơn Formation at Làng Vạc locality 1. All specimens are coated with ammonium chloride sublimate prior to digital photography; specimens are internal moulds unless otherwise stated. Scale bars: (a, b, d, g, i, k, o, p) = 4 mm; (c) = 8 mm; (e, f, j, l–n) = 2 mm; (h) = 5 mm. (a–e) cranidia; (i–p) pygidia. (a) Leiostegiid genus and species indet. 2, BT1/603. (b–p) Troedssonia wimani: (b) BT2/603; (c) BT3/603; (d) BT4/603; (e) BT5/603; (f) left free cheek, BT6/603; (g) latex cast of counterpart of trunk, BT7/603; (h) trunk, BT8/603; (i) BT9/603; (j) BT10/603; (k) BT11/603; (l) BT12/603; (m) BT13/603; (n) BT14/603; (o) BT15/603; (p) BT16/603.
Material. Figured: a single fragmentary cranidium (BT1/603).
Discussion. This cranidium is differentiated from leiostegiid genus and species indet. 1 by the arched anterior border and anteriorly shallowing axial furrow. As these differences might result from deformation, our treating them as separate taxa is tentative. The single fragmentary cranidium does not permit specific assignment.
Sông Đà occurrence. In Đông Sơn Formation green shales collected in situ (LV-1) near Làng Vạc village, co-occurring with Troedssonia wimani.
Family Tsinaniidae Kobayashi, Reference Kobayashi1935
Genus Shergoldia Zhang & Jell, Reference Zhang and Jell1987
Type species. Tsinania nomas Shergold, Reference Shergold1975, Chatsworth Limestone, Queensland, Australia.
Shergoldia cf. Shergoldia trigonalis (Kobayashi, Reference Kobayashi1933)
Figure 9a, e, j–l
2008 Tsinania sp. Phạm, p. 194, pl. 10, fig. 2.
Material. Figured: One cranidium (BT1/601) and four pygidia (BT5/601, BT10/601–BT12-601). Unfigured: seven pygidia (CMCIP87774–87780) along with one pygidium from Phạm (Reference Phạm2008).
Discussion. Among the tsinaniids, which are a strongly effaced group, Shergoldia possesses a notable and wide pygidial border (see Shergold, Reference Shergold1975, pl. 50, figs 3–9). Our material resembles Shergoldia trigonalis (Kobayashi, Reference Kobayashi1933) in having this and a relatively long frontal area, plus possible small bacculae. Deformation of the Thanh Hóa material makes it difficult to assess whether the glabella was originally parallel sided or tapering. Our material lacks the slightly depressed, shelf-like anterior border seen in S. cf. S. nomas from Tarutao Island, Thailand (Shergold et al. Reference Shergold, Burrett, Akerman and Stait1988) and in S. laevigata (Zhu et al. Reference Zhu, Hughes and Peng2007). Rather, our material apparently had a long frontal area with a weakly inflated anterior border as in Taipaikia (see Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011). That form does not have the pointed anterior margin seen in our specimen and consistently present in Shergoldia. Our material is thus most comparable to Shergoldia trigonalis, which also has a similar number of axial rings.
Sông Đà occurrence. In the Hàm Rồng Formation dolomite from collection 1 at Đền Bà Triệu, co-occurring with Prosaukia sp. indet. Also see Phạm (Reference Phạm2008, pl. 10, fig. 2).
Family Nileidae Angelin, Reference Angelin1854
Genus Troedssonia Poletaeva in Khalfin, Reference Khalfin1960
Type species. Symphysurus? wimani Troedsson, Reference Troedsson1937, Torsuqtagh Formation, Xinjiang, China.
Troedssonia wimani (Troedsson, Reference Troedsson1937)
1937 Symphysurus? wimani Troedsson, p. 44, pl. 4, figs 3–6, (?) 7–8.
1965 Symphysurus (Troedssonia) wimani; Lu et al., p. 532, pl. 111, figs 1–4.
1980 Symphysurus (Troedssonia) wimani; Lu & Lin, p. 128, pl. 3, figs 3, 4.
1984 Symphysurus (Troedssonia) wimani; Lu & Lin, pp. 116–17, pl. 15, figs 4–13.
1984 Troedssonia wimani; Apollonov & Chugaeva, pp. 10, 11, pl. 16, figs 4, 8, 12, pl. 18, figs 12, 14–19, pl. 21, figs 8, 9.
1984 Troedssonia wimani; Peng, p. 367, pl. 10, fig. 8b, pl. 13, figs 3, 4, pl. 14, fig. 9.
1990 Troedssonia wimani; Lu & Zhou, p. 40, pl. 15, figs 1–4.
Sông Đà material. Figured: Four cranidia (BT2/603–BT5/603), eight pygidia (BT9/603–BT16/603), one free cheek (BT6/603) and two pygidia with several thoracic segments (BT7/603, BT8/603). Unfigured: eight cranidia (CMCIP87790–87797), two free cheeks (CMCIP87789–87799), one trunk CMCIP87800) and 18 pygidia CMCIP87801–87818).
Discussion. The relatively large size and effaced form of this taxon resembles both asaphid trilobites such as Niobella and such nileid trilobites as symphysuriids. We acknowledge that, given the deformed state of our material, confident assignment to either of these two groups is challenging. These specimens from Làng Vạc are here assigned to a nileid genus on the basis of the articulating pits at the base of the glabella, the very short anterior border, the evenly curved anteriormost dorsal suture with modest expansion of the glabella at its anterolateral corner and the presence of a median suture in a trilobite with an apparently impendent hypostomal condition. Nileidae almost exclusively have anterior thoracic articulation at or close to the axis, which then becomes further removed posteriorly, whereas in asaphids the fulcrum is located more abaxially. Determining the original form of the anterior border, which is pointed sagittally in asaphids, is complicated by tectonic deformation. However, retrodeformation of the best specimen available (Fig. 12), while fully not effective in removing the effects of shear, provides no evidence of a sagittal inflection. The extended doublure of the associated free cheek, which is apparently conspecific, might suggest an impendent hypostomal attachment as is characteristic of nileids.
Having made the determination as a nileid, species assignment is straightforward as within that group this form bears all the characteristics of T. wimani.
Sông Đà occurrence. In Đông Sơn Formation green shales collected in situ in collection 1 near Làng Vạc village, co-occurring with leiostegiid genus and species indet. 2.
Phylum BRACHIOPODA
Subphylum RHYNCHONELLIFORMEA Williams et al. Reference Williams, Carlson, Brunton, Holmer and Popov1996
Class STROPHOMENATA Williams et al. Reference Williams, Carlson, Brunton, Holmer and Popov1996
Order BILLINGSELLIDA Schuchert, Reference Schuchert1893
Suborder BILLINGSELLIDINA Schuchert, Reference Schuchert1893
Superfamily Billingselloidea Schuchert, Reference Schuchert1893
Family Billingsellidae Schuchert, Reference Schuchert1893
The billingsellids are a common and distinctive group of strophomenate brachiopods that have many features that mimic orthide brachiopods where they were traditionally placed, prior to 2000. Nevertheless, the presence of a pseudodeltidium and a chilidium together with transverse flat-lying socket plates and, in particular, a secondary shell layer consisting of crossbladed laminae, indicated a reassignment of billingsellids to the Strophomenata (Williams et al. Reference Williams, Carlson, Brunton, Holmer and Popov1996). The family ranges from the middle Cambrian to the Lower Ordovician (Floian) and currently includes six genera. Two, possibly three, taxa of billingsellids are reported here, two assigned, with varying levels of confidence, to Billingsella and one tentatively related to Saccogonum.
Genus Billingsella Hall & Clarke, Reference Hall and Clarke1892
Type species. Orthis pepina Hall, Reference Hall1863, p. 134, pl. 6, figs 23–27.
Billingsella is normally ventribiconvex with variable cardinal extremities, a robust pseudodeltidium and convex chilidium together with a minute apical foramen (see also Harper & McKenzie in Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011). The dental plates are widely divergent and the ventral muscle scars are commonly impressed on a long, tongue-like callus; a subperipheral rim is occasionally developed. The other members of the family conform to this Billingsella model but have minor differences, mainly in overall shape and features of the ventral interior. Billingsella ranges from the middle Cambrian to the Lower Ordovician (Tremadocian) and has a cosmopolitan distribution. The majority of its 20 reported species are known from the Furongian. At least one, possibly two, species in this study are assigned to Billingsella.
Billingsella cf. tonkiniana Mansuy, Reference Mansuy1915
Figure 13a–g, j–l

Fig. 12. Retrodeformation of Troedssonia wimani cranidium BT5/603 using cleavage as principal extension direction: (a) original; (b) retrodeformation, showing expansion of anterior glabellar margins. Scale bar = 10 mm.
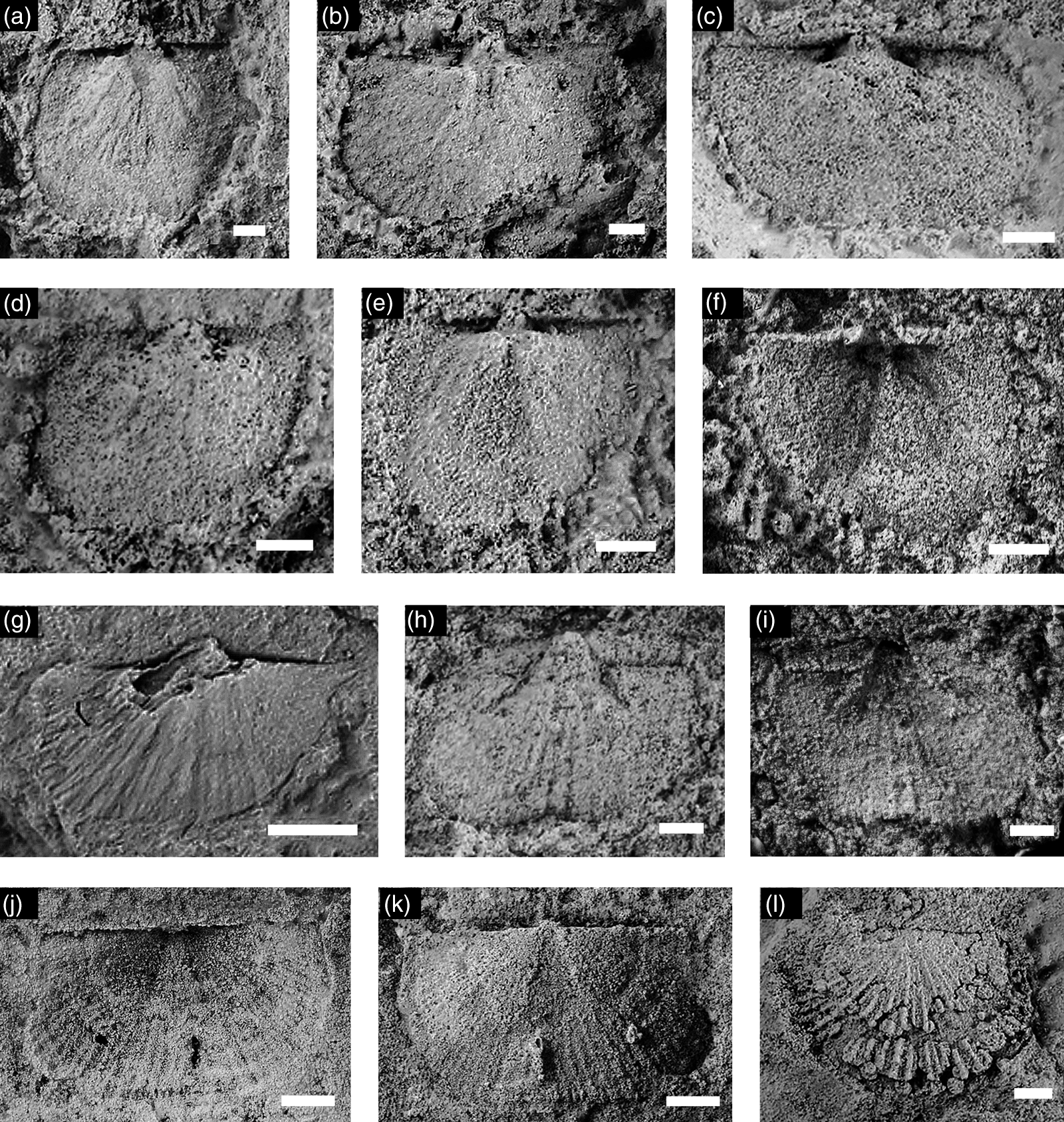
Fig. 13. Billingsellid brachiopods from the Hàm Rồng Formation at Đền Bà Triệu localities 2 and 3, and the Đông Sơn Formation at Làng Vạc locality 1. Scale bars are 2.5 mm in all figures. Locality is Đền Bà Triệu collection 2 unless otherwise stated. (a–g, j–l) Billingsella sp. cf. B. tonkiniana. (a–e) Internal moulds of dorsal valves displaying intraspecific variability: (a) BT1/604; (b) BT2/604; (c) BT3/604; (d) BT4/604; (e) BT5/604. (f) BT5/604, latex cast of dorsal valve interior. (g) BT6/604, partially exfoliated ventral exterior from Làng Vạc locality 1. (h, i) BT7/604, billingsellid gen. et. sp. indet. from Hàm Rồng Formation at Đền Bà Triệu collection 3: (h) internal mould of ventral valve; (i) latex cast of ventral exterior. (j, k) BT8/604: (j) dorsal valve interior; (k) latex cast of dorsal interior. (l) BT9/604, exterior of ventral valve.
cf. 1915 Billingsella tonkiniana Mansuy, p. 7, pl. I, fig. 2a–q.
cf. 1916 Billingsella tonkiniana Mansuy; Mansuy, p. 13, pl. I, fig. 12a–g.
cf. 2011 Billingsella cf. tonkiniana Mansuy; Harper & McKenzie in Hughes et al., p. 368, fig. 13a–k.
Material. Six dorsal internal moulds (BT1/604–BT5/604, BT8/604) and one ventral external mould (BT9/604), from Đền Bà Triệu (DBT-2), one external ventral mould from Làng Vạc (LV-1) (BT6/604).
Description. Exterior (modified from Harper & McKenzie in Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011). Biconvex valves, transverse to subquadrate outline, maximum width commonly at or just anterior to hingeline; cardinal extremities rectangular or slightly obtuse. Anterior commissure slightly sulcate. Dorsal valve convex, transverse; faint sulcus arising at or near umbo, deepening and widening anteriorly; flanked by two strong ribs. Dorsal interarea relatively short, flat; chilidium relatively small, convex occupying the apical part of the notothyrium. Ornament of strong costae and costellae, with sharp profiles and flat interspaces; arising by both branching and intercalation; concentric growth lines variably developed, accentuated at and near anterior commissure; up to ten ribs developed per 5 mm at about 5 mm from dorsal umbo, with commonly five in the sulcus.
Dorsal interior. Notothyrial platform high, well developed with simple, bladelike cardinal process flanked by pair of suboval depressions, marking the site of the diductor scars; platform extended anteriorly as broad ridge that fades within the posterior third of the valve length. Notothyrial platform flanked by flat-lying socket ridges that extend laterally, subparallel to hingeline, thickening slightly distally. Posterior pair of adductor scars, oval, faintly impressed lateral to median ridge.
Discussion. This material has apparent similarities with B. tonkiniana Mansuy, Reference Mansuy1915, a species described briefly and illustrated from the North Việt Nam block. A similar form was described and illustrated from the Quartzite Formation, Wachi La section, Black Mountains, Bhutan (Harper & McKenzie in Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011). Harper & McKenzie (in Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011) provided a detailed discussion of Billingsella, noted morphological variation in the genus and commented on similar species; this is not repeated here. Mansuy’s species, B. tonkiniana was compared by Mansuy with the Laurentian species, B. coloradoensis (Shumard, Reference Shumard1861), hinting at biogeographical links with that province. Both the Bhutan and Vietnamese species are tectonically deformed, to varying degrees, making precise morphological comparisons difficult, but both are clearly similar. To avoid the proliferation of specific names, the Bhutanese material was compared with B. tonkiniana. This material is similarly compared with Mansuy’s species. Pending revision of Mansuy’s species, the Bhutan and Hàm Rồng taxa may be included in that species, or their distinctive characteristics could form the basis for new species (see also Harper & McKenzie in Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011).
Zhan et al. (Reference Zhan, Jin, Rong, Zhu and Han2010) noted variation in the cardinal angles, crenulations on the inner shell surface, the length of the interarea, the dorsal sulcus, the notothyrial platform and the dental plates in their descriptions of the two Billingsella species reported from SW Guangxi Province, Billingsella guangxiensis Zeng, Reference Zeng1977 and a new species they erected, B. costata. Of the two, the Hàm Rồng material is most similar to B. guangxiensis, based on the limited material available; the similarity of the latter to B. tonkiniana will be tested following pending revision of the types of that species.
Sông Đà occurrence. Hàm Rồng Formation, at Đền Bà Triệu in collection 2, and Đông Sơn Formation at Làng Vạc collection 1.
Genus Saccogonum Havlíček, Reference Havlíček1971
Type species. Saccogonum saccatum Havlíček, Reference Havlíček1971 from the Fezouata Infériere (Furongian), the Anti-Atlas Mountains of Morocco.
Saccogonum sp.
Figure 14a–f

Fig. 14. Other rhynchonelliform brachiopods from the Hàm Rồng Formation. All specimens from Đền Bà Triệu. Scale bars are 2.5 mm in all figures. (a–f) Saccogonum sp. indet. from collection 3: (a) shell cluster of moulds of dorsal (right) and ventral valves (left), BT1/605; (b) latex cast of internal of dorsal valve, BT1/605; (c) latex cast of internal of ventral valve, BT1/605; (d) larger part of same block with external moulds of ventral valves, BT1/605; (e, f) latex casts of external moulds of ventral valves, BT1/605. (g–j) Huenellid, gen. et. sp. indet. dorsal valves from collection 2: (g, h) BT2/605, (h) latex cast; (i) external mould of dorsal valve interior, BT3/605; (j) internal mould of ventral valve interior, BT3/605.
Material. Shell cluster with five ventral and two dorsal internal moulds together with some incomplete external moulds (BT1/605).
Discussion. A cluster of dorsal and ventral internal moulds from this horizon show many billingsellid characters, notably the flat-lying, transverse socket ridges, the well-developed ventral interarea and ventral muscle adductor scars impressed on a long, tongue-like callus. The material differs from Billingsella in a number of key characters; the shells are markedly dorsibiconvex, and the disposition of the ventral muscle scars with a pentagonal shape and its cardinalia are different. The material has strong similarities with Saccogonum, described and illustrated initially from the basal Fezouata Infériere Formation possibly of Furongian age, from the Anti-Atlas Mountains of Morocco (Havlíček, Reference Havlíček1971). The genus was revised, two new species (S. arenosum and S. salebrosum) were described in detail from the High Atlas and Meseta and the type species reassessed by Mergl et al. (Reference Mergl, Geyer and El Attari1998), providing more information on the ventral and dorsal interiors of this genus. In addition, a Furongian age for the genus was confirmed.
The Moroccan and Vietnamese species, particularly in the ventral valves, show some similarities with some other penecontemporary brachiopod groups. Mergl et al. (Reference Mergl, Geyer and El Attari1998) noted similarities with Protambonites and speculated that Saccogonum together with Billingsella may have given rise to the Clitambonitoidea in the Mediterranean Region. Alternatively, Saccogonum was the progenitor of the Polytoechoidea (see Topper et al. Reference Topper, Harper and Brock2013).
Sông Đà occurrence. Hàm Rồng Formation in collection 3 at Đền Bà Triệu.
Billingsellid gen. et sp. indet.
Figure 13h, i
Discussion. A single ventral valve from collection 2 at Đền Bà Triệu (BT7/604), with a moderately high interarea, a faint sulcus together with well-developed, widely divergent dental plates, is difficult to assign. Some of the features are those of a billingsellid and may belong to Billingsella but further and better-preserved material is required to confirm or otherwise this placement.
Order PENTAMERIDA Schuchert & Cooper, Reference Schuchert and Cooper1931
Suborder SYNTROPHIIDINA Ulrich & Cooper, Reference Ulrich and Cooper1936
Superfamily PORAMBONITOIDEA Davidson, Reference Davidson1853
Family Tetralobulidae Ulrich & Cooper, Reference Ulrich and Cooper1936
Subfamily Syntrophopsinae Ulrich & Cooper, Reference Ulrich and Cooper1936
Genus Palaeostrophia Ulrich & Cooper, Reference Ulrich and Cooper1936
Type species. Syntrophia orthia Walcott, Reference Walcott1905; lower Chaomidian Formation (formerly called Chaumitien Limestone, upper Cambrian); Jinan, Shandong Province, North China.
Palaeostrophia cf. jingensis Zeng, Reference Zeng1977
Figure 15a–h
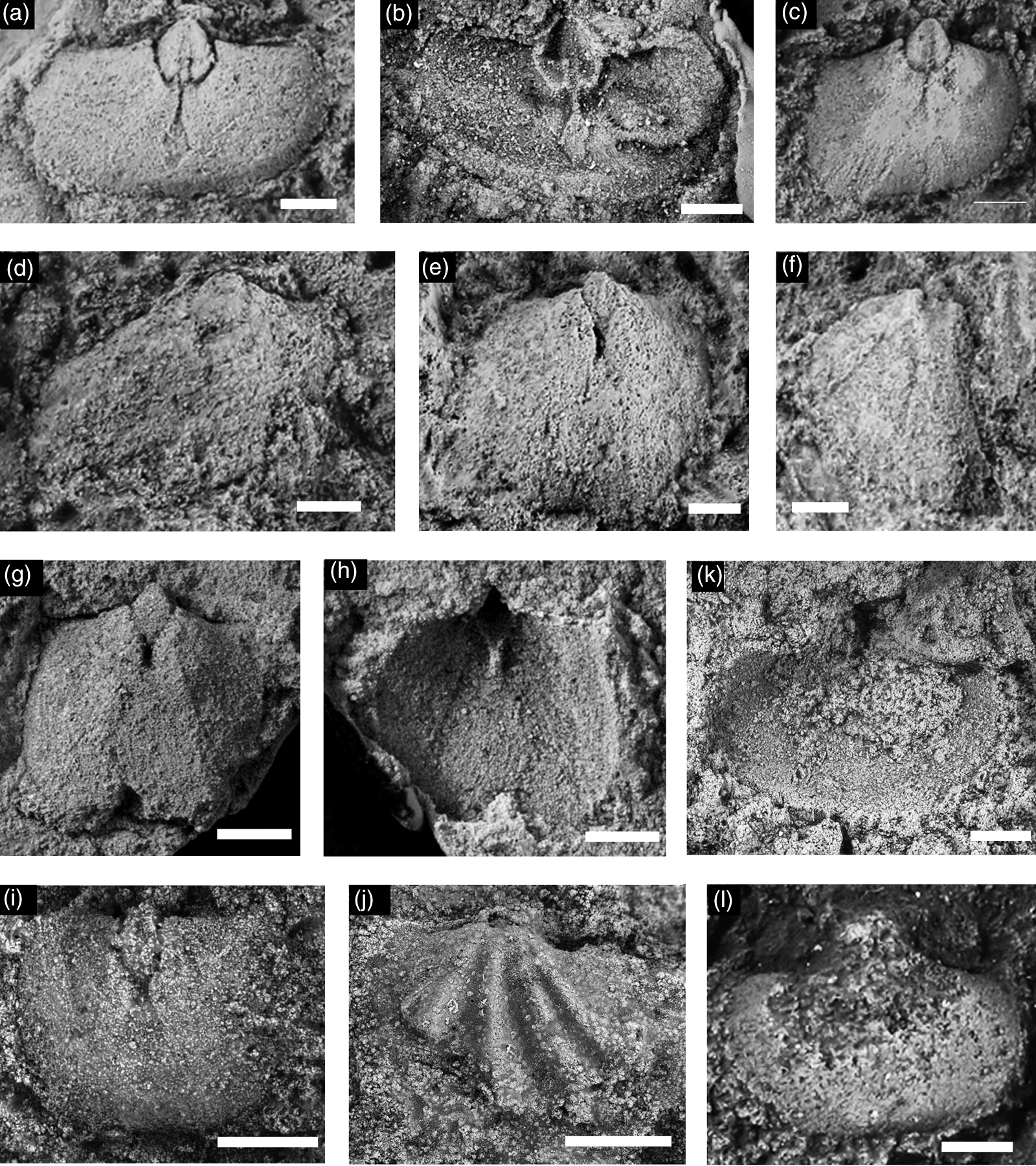
Fig. 15. Other rhynchonelliform brachiopods from the Hàm Rồng and Sông Mã formations. Scale bars are 2.5 mm in all figures. All specimens from Đền Bà Triệu collection 2 apart from (c, d, f) from Đền Bà Triệu collection 1; all interior views. (a–h) Palaeostrophia sp. cf. P. jingensis: (a–d) ventral valves: (a) BT1/606; (b) BT1/606, latex cast; (c) BT2/606; (d) BT3/606; (e–h) dorsal valves: (e) BT4/606; (f) BT5/606; (g, h) BT6/606, (h) latex cast. (i) Syntrophopsinin gen. et. sp. indet., ventral valve, BT7/606. (j) Plectotrophia aff. imparicostata, dorsal valve, Nghĩa Phú, BT8/606. (k, l) Eostrophiid gen. et sp. indet., latex cast and internal mould of ventral valve, Sông Mã Formation at Xuân Sơn, BT9/606.
?1915 Syntrophia orthia Mansuy, p. 9, pl. 1, fig. 4.
?1916 Syntrophia orthia Mansuy, p. 16, pl. 1, fig. 20.
cf. 1977 Palaeostrophia jingensis Zeng, p. 49, pl. 18, figs 4, 5.
cf. 2010 Palaeostrophia jingensis Zeng; Zhan et al., p. 119, figs 15D–O, 16–20.
Material. Three dorsal (BT4/606–BT6/606) and three ventral (BT1/606–BT3/606) internal moulds.
Description. These medium-sized, biconvex valves are subcircular to elongately suboval, rectimarginate to plicate. The ventral interior is characterized by small teeth, a large delthyrial cavity and a sessile spondylium. The dorsal interior has a deep notothyrial cavity, small sockets, lacking a cardinal process.
Remarks. This smooth syntrophopsin in is represented by a selection of variable dorsal and ventral internal moulds. Zhan et al. (Reference Zhan, Jin, Rong, Zhu and Han2010) noted variation in the size and shape of the ventral muscle scar and shapes of the shells themselves. Similar patterns are obvious in the material from Thanh Hóa. Mansuy (Reference Mansuy1915, Reference Mansuy1916) noted and illustrated specimens he considered synonymous with Syntrophia orthia Walcott from the North Việt Nam terrane. That material is in need of re-examination; the figure and description are inadequate for detailed comparative purposes but show similarities to the material from both Jingxi and Thanh Hóa.
Sông Đà occurrence. Hàm Rồng Formation, Đền Bà Triệu at all three collecting horizons.
Genus Plectotrophia Ulrich & Cooper, Reference Ulrich and Cooper1936
Type species. Plectotrophia bridgei Ulrich & Cooper, Reference Ulrich and Cooper1936, p. 627 (brief description only); illustrated by Ulrich & Cooper (Reference Ulrich and Cooper1938, p. 198, pl. 40B, figs 5–7, 9–22), Wilberns Formation (upper Cambrian); Point Peak, Llano Quadrangle, Texas.
Plectotrophia aff. imparicostata Zhan et al. Reference Zhan, Jin, Rong, Zhu and Han2010
Figure 15j
aff. 2010 Plectotrophia imparicostata Zhan et al., p. 127, figs 21A–O.
Material. A single internal dorsal valve (BT8/606).
Discussion. Only one poorly preserved and incomplete dorsal valve was discovered. This species is characterized by a convex dorsal valve, a transverse outline and a distinctive costate ornament; a strong median costa, broadening and heightening anteriorly is supplemented by at least two costae on each flank, curving laterally. Traces of strong concentric growth lines are preserved. The shell has a small notothyrial cavity; the cardinalia are indistinct. The specimen shows some similarities, in terms of shape and ornament, with P. imparicostata from the lower Guole Formation (upper Furongian), Guanxi Province, southern China, with which it is compared. Zhan et al. (Reference Zhan, Jin, Rong, Zhu and Han2010) included their species within Plectotrophia, although a coarse costation is not usual for the genus and may require appropriate recognition in due course within this family.
Sông Đà occurrence. Hàm Rồng Formation from collection 2 at Đền Bà Triệu.
Syntrophopsinin gen. et sp. indet.
Figure 15i
Material. A relatively small, subquadrate valve from collection 2 at Đền Bà Triệu (BT7/606) displays long, thin, convergent plates.
Discussion. The material is insufficient to offer a more definitive assignment and indeed it is difficult to determine if this is in fact a ventral or dorsal valve. One possibility is that it is a small dorsal valve of Palaeostrophia, although it is more likely a ventral valve, because it displays evidence of a spondylium and median septum.
Family Eostrophiidae Ulrich & Cooper, Reference Ulrich and Cooper1936
Eostrophiid gen. et sp. indet.
Figure 15k, l
Material. A single poorly preserved ventral internal mould (BT9/606).
Discussion. The valve is semi-elliptical in shape, convex with a poorly developed interarea. The internal structures are unclear but it does not appear to have a spondylium, raised or sessile. Although the evidence is limited and the shell inadequately preserved, an assignment to the Porambonitoidea is probable, with a placement within the Eostrophiidae a possibility.
Sông Đà occurrence. In the Sông Mã Formation at Xuân Sơn, co-occurring with K. xuanensis sp. nov. and Solenoparops sp. indet.
Family Huenellidae Schuchert & Cooper, Reference Schuchert and Cooper1931
Huenellid gen et sp. indet.
Figure 14g–j
Material. External and internal moulds of a huenellid are illustrated from the Hàm Rồng Formation from collection 2 at Đền Bà Triệu (BT2/605, BT3/605). Two unfigured specimens are CMPIP87823–87824.
Discussion. Mansuy (Reference Mansuy1915, pl. 1, fig. 3; 1916, pl. 1, fig. 19) illustrated a single valve, assigned to Huenella orientalis Walcott from the North Việt Nam block. While it is possible all three shells are related, Mansuy’s material requires further scrutiny before the conspecifity of these forms can be confirmed.
7. Palaeoecology, biostratigraphy and biogeography
7.a. Palaeoecology
Because fossil recovery is dependent on weathering patterns (see Section 4) it is difficult to assess whether the fossil-yielding beds were typical of the original environment. The fossils we were able to recover represent concentrations of disarticulated shells, as is common in late Cambrian carbonate platform assemblages. The diversity recovered in any collection is low, but several horizons contain a mix of trilobite and brachiopod fossils; collection 3 from Đền Bà Triệu is notable for being apparently an exclusively brachiopod shell bed, particularly rich in Saccagonum sp.
7.b. Lowest Palaeozoic biostratigraphy of the Sông Đà terrane
The isolated outcrops with short stratigraphic sections, relatively low taxic diversity and poor quality of preservation place limits on biostratigraphical interpretation. The total range of the Sông Đà Cambrian fauna spans the last two series of that system, but recovery to date is confined to three of the seven stages (Fig. 4). Whether the absence of fauna from the other four stages reflects a regional lack of sedimentary rocks of these ages, or sporadic sampling to date, is currently unclear. Of the five localities we visited, three were uppermost Cambrian or lowermost Ordovician. At Đền Bà Triệu the three sampled horizons apparently span an interval of some 300 m (Phạm, Reference Phạm2008, fig. 12) and represent c. 3 million years (Fig. 4). Based on this observation and a highly approximate calculation that takes into account the inverse relationship between accumulation rate and timespan over which it is measured (Sadler, Reference Sadler1981), the interval represented by the whole of the Hàm Rồng Formation might be expected to have been over 10 million years in duration. The maximum reported thickness for the formation of ∼1150 m would not be unusual for a stratum that covers an ∼10 myr age span, but in most places the formation is reportedly thinner, being as thin as 270 m (Phạm, Reference Phạm2008). This may imply condensed (or incomplete) sections in some regions. The present lithostratigraphic system (Figs 3, 4) equates formations with Series, and while it is notable that the fauna assigned to the Hàm Rồng Formation spans the majority of the late Cambrian, biostratigraphical correlation of the Sông Đà taxa with their occurrences in strata recovered from elsewhere suggests that rocks in the formation span an interval of almost 15 million years, and that the formation has yet to yield identifiable fossils from an ∼10 million year interval during the earlier Furongian (Fig. 4).
7.c. Distribution and biogeography
7.c.1. Within northern Việt Nam
To date there are two other regions in Việt Nam from which Cambrian fossils have been recovered, both of which belong to the North Việt Nam terrane (Saurin, Reference Saurin and Rodgers1956; Phạm & Lương, Reference Phạm and Lương1996; Phạm, Reference Phạm2008) (Fig. 1). The first is referred to as East Bắc Bộ, and contains the classic Cambrian successions of the border region with China including the ‘Chang Pung’ section. A thorough revision of Mansuy’s (Reference Mansuy1915, Reference Mansuy1916) monographs, along with the resampling and stratigraphic logging of the sections in northern Việt Nam is pending. Until it is completed, a detailed comparison between the Thanh Hóa fauna from the Sông Đà terrane, and that from the Northern Việt Nam terrane cannot be completed. But in the interim, inspection of Mansuy’s figured material shows not only a similar assemblage of platform dwelling trilobites and brachiopods, but also hints that the fauna represented in northernmost Việt Nam spans a broadly similar sampling of later Cambrian time, although likely with several more stages sampled (Fig. 4). The section at Lũng Cú at 23° 21.180′ N, 105° 18.647′ E contains an interbedded succession of claystone consistent with trilobites preserved as articulated, flattened exoskeletons, and oolitic limestone, with disarticulated specimens.
The other Cambrian fossil-bearing area, Thần Sa, is to the east of east Bắc Bộ. This region shows a distinct change in facies towards a more siliciclastic-rich, red-green claystone assemblage with siltstone and minor sandstone called the Thần Sa Formation (Phạm & Lương, Reference Phạm and Lương1996); trilobites are commonly preserved articulated in claystone and are reported to belong to genera such as Hedinaspis and Charchaqia (see Rushton et al. Reference Rushton, Williams, Nguyễn, Komatsu, Siveter, Zalasiewicz, Đinh, Nguyễn, Nguyễn and Takaka2018, fig. 6b) that, along with Lotagnostus, are typical of slope settings (Phạm, Reference Phạm2008). The reported occurrence of these genera, and a specimen referable to Koldinioidia from 21° 48.296′ N, 105° 53.931′ E, which we collected in 2010, suggests a Furongian age for this unit, close to the Jiangshanian – Stage 10 boundary (Fig. 4). Elsewhere the same section is said to yield Ptychagnostus atavus (Chernysheva in Trần et al. Reference Trần, Tạ, Phan and Lê1964). Phạm (Reference Phạm2008) reported a mixed trilobite assemblage at Banloa in northernmost Việt Nam consisting of both shelf (saukiid and tsinaniid) and slope (Charchaqia sp.) forms.
Regional differentiation among trilobite and brachiopod-bearing Cambrian rocks in Việt Nam thus represents a shelf–slope transition within the North Việt Nam block, which mimics that between the Yangtze platform and Jiangnan belt on the Southern China block (Phạm, Reference Phạm2008), and a similar deepening transect persisted into Early Devonian time (Tống et al. Reference Tống, Tạ, Janvier, Nguyễn, Nguyễn and Nguyễn2013; Williams et al. Reference Williams, Komatsu, Tanaka, Nguyễn, Zalasiewicz, Vandenbroucke, Wallis and Perrier2016). Comparison between the Cambrian geology of the Sông Đà terrane with that of the North Việt Nam block indicates similarities not only in the persistence of a carbonate shelf palaeoenvironment and attendant fauna, but broadly also in the interval of the later Cambrian represented in the record. Both areas also share a notable sub-Devonian unconformity that cuts deeply into the Ordovician (Phạm & Lương, Reference Phạm and Lương1996). The Cambrian–Ordovician record is thus consistent with physical continuity between these areas at that time.
7.c.2. Regional trilobite palaeogeography
The early Palaeozoic trilobites described from the Sông Đà terrane are typical of the equatorial Gondwana shelf fauna. All of the genera recognized herein are widely known in the region. Both the fauna and the time intervals represented are consistent with a close association between the Sông Đà terrane and the North Việt Nam block during Cambrian time, but they do not require it, because their similarities are general to a wider region. For example, the trilobite genera recorded herein are typical of shelf faunas in both the South China and North China blocks (Zhou & Zhen, Reference Zhou and Zhen2008), and also more broadly along the equatorial Gondwana margin in Sibumasu, including Baoshan (Kobayashi, Reference Kobayashi1957; Sun & Xiang, Reference Sun and Xiang1979; Luo, Reference Luo1983, Reference Luo1984; Shergold et al. Reference Shergold, Burrett, Akerman and Stait1988; Wernette et al. Reference Wernette, Hughes, Myrow and Sardsud2020 a,b), northern India (Peng et al. Reference Peng, Hughes, Heim, Sell, Zhu, Myrow and Parcha2009), Bhutan (Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011), northern and central Australia (Shergold, Reference Shergold1991; Shergold et al. Reference Shergold, Laurie and Shergold2007) and Oman (Fortey, Reference Fortey1994). This is true both for the middle Cambrian elements and for most of those clustered around the Cambrian–Ordovician boundary (Shergold, Reference Shergold1988). Hence, the fauna recovered here does not more specifically localize the position of the Sông Đà terrane within equatorial Gondwana during Cambrian time. Recognition of the shelf–slope transition in the North Việt Nam block in its current roughly W–E orientation may ultimately help constrain its original placement, given that shelf–slope transitions are critical for restoring the orientation of ancient continental margins (Fortey & Cocks, Reference Fortey, Cocks, Hall and Holloway1998).
7.c.3. Late Cambrian brachiopod palaeobiology/palaeogeography
Relatively little is known about the origins and distributions of Furongian rhynchonelliform brachiopods (Harper et al. Reference Harper, Topper, Cascales-Miñana, Servais, Zhang and Ahlberg2019), although some clear trends are emerging. Laurentian and related bioregions appear to be characterized by Billingsella, syntrophidiines and some early orthoids; Avalonia, Baltica and possibly western South America have Furongian shell beds dominated by the plectorthoid Orusia, whereas higher latitude faunas including Billingsella, Protambonites and Saccogonum are less well known. The low-diversity fauna from Hàm Rồng, Việt Nam, is closely similar to the faunas described by Mansuy from the North Việt Nam block and that from broadly coeval horizons in southwestern Guangxi Province, southern China (Zhan et al. Reference Zhan, Jin, Rong, Zhu and Han2010). The latter fauna is dominated by Billingsella, Guoleella, Palaeostrophia and Plectotrophia, and Zhan et al. (Reference Zhan, Jin, Rong, Zhu and Han2010) considered that the fauna has a strong connection with those from Laurentia. The combination of Billingsella, Palaeostrophia and Plectotrophia indicates a link between southern China and this part of Việt Nam, but in the absence of a more global analysis of the distributions of Furongian brachiopods, these new data provide a testable hypothesis rather than firm evidence.
As noted previously (Harper in Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011), the Billingsella Association (Bassett et al. Reference Bassett, Popov, Holmer, Crame and Owen2002), a low-diversity assemblage rich in the eponymous genus itself or closely related forms such as Cymbithyris, dominated late Cambrian brachiopod-dominated assemblages in shallow-water environments. Many of these assemblages were limited in time and space but often formed high-density, high-dominance clusters in thin, locally extensive accumulations. In the Wachi La section, Bhutan, Billingsella occurs throughout the fossiliferous interval and dominates some, but not all, beds in which it occurs. Here and elsewhere, Billingsella may have pursued an opportunist life strategy, typified by thick, near-monospecific shell beds, exposed, for example, in the middle and upper Cambrian rocks of Iran and the Furongian strata of north-central Kazakhstan (Bassett et al. Reference Bassett, Popov, Holmer, Crame and Owen2002).
8. Detrital zircon geochronology
In addition to the fossil-bearing Cambrian rocks from the North Việt Nam block and the Sông Đà terrane, other rocks in Việt Nam have been interpreted as Cambrian even though they have not yielded fossils. Here we examine detrital zircon spectra from biostratigraphically constrained lower Palaeozoic rocks in the two northern areas, and two samples from the Trường Sơn terrane which lies immediately to the south of the Sông Mã suture (Figs 1, 3). Analyses were undertaken using a sensitive high-resolution ion microprobe (SHRIMP) at the Research School of Earth Sciences, Australian National University. Procedures are given in Williams (Reference Williams, McKibben, Shanks and Ridley1998 and references therein). Polished grain mounts of zircon grains analysed in this study were imaged using reflected and transmitted light microscopy, cathodoluminescence (CL) and scanning electron microscopy (SEM). Images were used to determine the internal structures of zircon grains and to ensure that the ∼20 μm SHRIMP spots were fully within the youngest single age component (i.e. the rims). The SQUID Excel macro (Ludwig, Reference Ludwig2001) was used to process the data. U/Pb ratios were normalized using the Temora reference zircon (value of 0.0668) with an age of 417 Ma (Black et al. Reference Black, Kamo, Allen, Aleinikoff, Davis, Korsch and Foudoulis2003). Uncertainties given for individual analyses (ratios and ages) are at the 1s level (see online Supplementary Material).
Correction for common Pb was made either using the measured 204Pb/206Pb ratio, or for grains younger than ∼800 Ma (or those low in U and so radiogenic Pb) the 207Pb correction method was used (see Williams, Reference Williams, McKibben, Shanks and Ridley1998). When the 207Pb correction is applied, it is not possible to determine radiogenic 207Pb/206Pb ratios or ages. The 207Pb–206Pb ages were generally used in the probability density spectra for analyses older than 800 Ma, whereas for zircons <800 Ma, the 206Pb–238U age was used. The concentration of U, and thereby radiogenic Pb, was also taken into account for selecting preferred ages. Some grain analyses were interpreted to be discordant, and this was based in part on the proximity to the concordia curve (using the total ratios, uncorrected for common Pb), and in part on whether the radiogenic 206Pb–238U age is part of a grouping of like ages, or a single outlier significantly younger than the inferred depositional age of the strata. Such interpreted discordant analyses were excluded from the age spectra.
8.a. Description
Sample DCV-1 was collected from upper Cambrian strata near Đình Cả, in the North Việt Nam block, ∼90 km north of Hà Nội (21° 44.800′ N, 106° 04.071′ N). Sixty-nine grains were analysed and these range in age from 486 ± 5 Ma to 2487 ± 5 Ma. The bulk of the grains range from ∼486 Ma to ∼1100 Ma. There are four grains between ∼1250 and ∼1350 Ma, a few scattered grains between ∼1825 and ∼2180 Ma, and another five grains that make a well-defined peak between ∼2445 and ∼2500 Ma (Fig. 16).
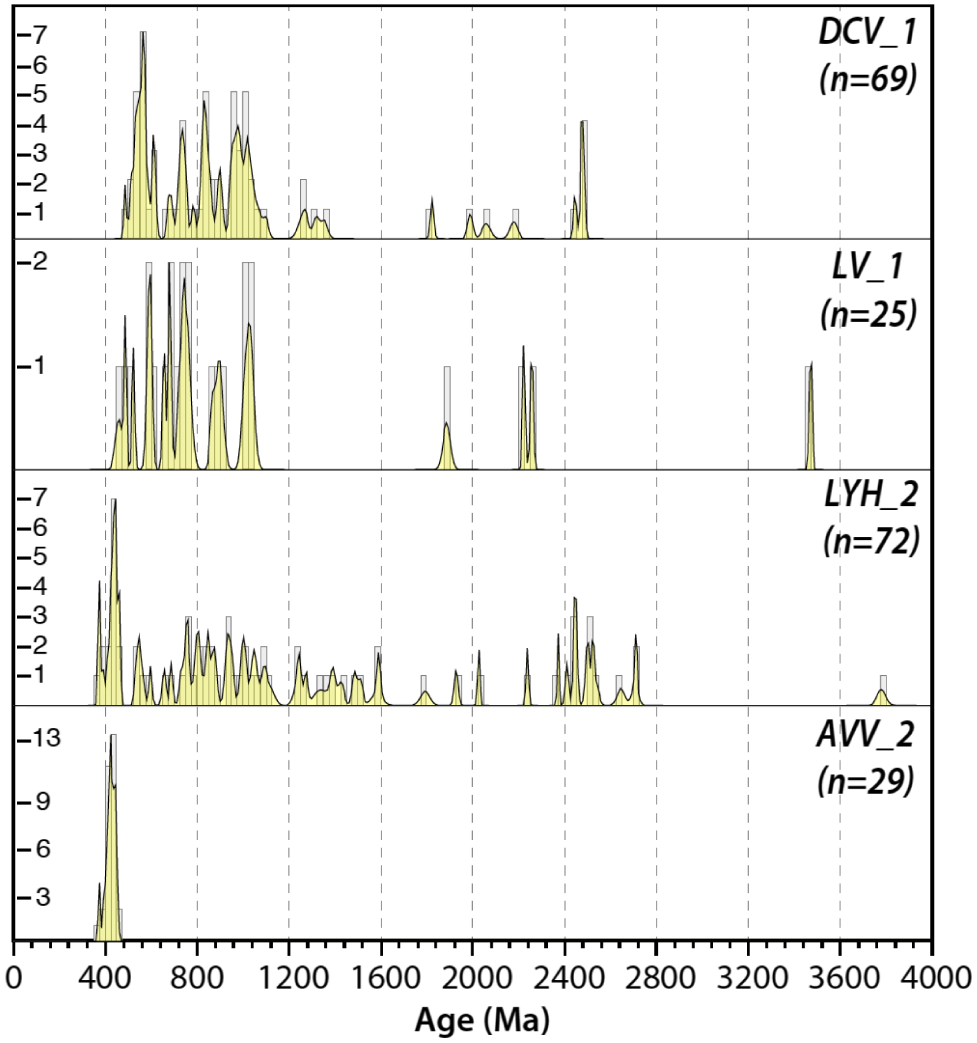
Fig. 16. Detrital zircon age spectra from Cambrian and Ordovician strata from Việt Nam. DCV_1 – Đinh Cà, Furongian sample in the North Việt Nam block; LV_1 – Làng Vạc, Tremadocian sample from the Sông Đà terrane; LYH_2 – Lý Hòa, Lower Devonian sample from the Trường Sơn terrane; AVV_2 – A Vương Formation, Trường Sơn terrane.
Sample LV-1 was collected from a sandstone bed, 2 m below the bed bearing the Tremadocian trilobite Troedssonia wimani in the Lower Ordovician Đông Sơn Formation near the village of Làng Vạc, northern Việt Nam, ∼100 km SSE of Hà Nội (20° 13.691′ N, 105° 22.376′ E). The sample yielded only 27 grains, of which one had very high discordancy, and another had an age (391 ± 6 Ma) well younger than the known depositional age and thus it likely suffered lead loss; these grains were thus not included in the analysis. The spectrum for this sample displays a wide range of grain ages, from 460 ± 15 Ma to 3472 ± 6 Ma. Most grains in the sample range from ∼460 to 1050 Ma. There is a small peak defined by three Palaeoproterozoic grains at ∼2220 Ma, and a single grain with an age of 3472 ± 6 Ma.
Sample LYH-2 was collected from Lower Devonian (Lochkovian to Pragian stages) sandstone strata at coastal outcrops at Lý Hòa, Quảng Bình Province, Việt Nam (17° 39.452′ N, 106° 31.053′ E), and is part of the Trường Sơn terrane. Seventy-two grains were analysed from this sample. Grain ages range from 372 ± 4 Ma to 3780 ± 12 Ma. The probability density plot shows a wide range of ages with many between ∼350 and ∼1650 Ma, and a peak at ∼445 Ma. There is a cluster of latest Archaean to earliest Palaeoproterozoic ages between ∼2370 and ∼2710 Ma, and then only a single older grain.
Sample AVV-2 was collected from low-grade metasedimentary rocks of the A Vương Formation, 40 km southwest of Thừa Thiên Huế, central Việt Nam (16° 14.580′ N, 107° 16.100′ E), an area that is part of the Trường Sơn terrane. The sample yielded a small number of grains (n = 30). Grain ages range from 373 ± 4 Ma to 453 ± 5 Ma, defining a relatively narrow and unimodal distribution. One grain, which yielded an age of 99 ± 2 Ma, is clearly much younger that the depositional age of the rock and likely records lead loss, and thus this grain was not included in the plotted spectrum (Fig. 16).
8.b. Interpretation
All the samples have relatively few grains, limiting the inferences that can be drawn from them. The A Vương Formation has commonly been correlated with the Cambrian of northern Việt Nam (Phạm & Lương, Reference Phạm and Lương1996, table 1; Phạm, Reference Phạm2008, fig. 14). Although the AVV-2 sample is particularly sparse in detrital zircon grains, the consistent age of c. 400 Ma shows that it is substantially younger than Cambrian in age, and its correlation with rocks in more northern Việt Nam requires reconsideration. The sample from Lý Hòa indicates a depositional age considerably younger than Cambrian and its large peak of grains at ∼445 Ma and slightly older indicate derivation from widespread Cambrian–Ordovician felsic intrusions around the equatorial Gondwanan margin (LeFort et al. Reference Lefort, Debon, Pêcher, Sonet and Vidal1986; Liu et al. Reference Liu, Hu, Gao, Feng, Huang, Lai, Yuan, Liu, Coulson, Feng, Wang and Qi2009; Mao et al. Reference Mao, Yin, Tang, Wang, Liao and Xiong2014; Shi et al. Reference Shi, Anderson, Wu, Yang, Li and Ding2016). This profile recalls those of several sectors of equatorial Gondwana, particularly from North China (McKenzie et al. Reference McKenzie, Hughes, Myrow, Choi and Park2011), Bhutan (Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011) and Sibumasu (Burrett et al. Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014; McKenzie et al. Reference McKenzie, Hughes, Gill and Myrow2014), and also those from Silurian – Middle Devonian samples from the North Việt Nam block (Königshof et al. Reference Königshof, Linnemann and Tạ2017, fig. 10). Our two samples from northern Việt Nam show an abundance of grain ages extending back to ∼1 Ga, with scattered peaks of older grains that are typical of lower Palaeozoic equatorial Gondwanan samples. Overall, these samples are of consistent derivation from sources in the North Việt Nam block and the Sông Đà terrane, both being part of equatorial Gondwana in Cambrian–Ordovician times, as was the Trường Sơn terrane in Devonian time (Usuki et al. Reference Usuki, Lan, Wang and Chiu2013). None of the samples show the prominent peaks of ∼0.9 and 1.1 Ga commonly present in profiles of rocks of this depositional age from South China (e.g. McKenzie et al. Reference McKenzie, Hughes, Gill and Myrow2014; Xu et al. Reference Xu, Cawood, Du, Zhong and Hughes2014; Yao et al. Reference Yao, Li, Li and Yang2014). However, with such a small number of dated grains in the sample, the significance of this absence, if any, is unclear.
9. Discussion
The geological term ‘Indochina’ is here considered to apply to an entity that had amalgamated during Permian time at the latest and made up of the Trường Sơn, Kon Tum and Loei-Pechabun terranes, in addition to the smaller entity in southern Việt Nam, Cambodia and southern Laos that now includes the Khorat Plateau (and marked as ‘Indochina’ in Burrett et al. Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014, fig. 1 and Loydell et al. Reference Loydell, Udchachon and Burrett2019, fig. 1) (Fig. 17). This geographical concept of Indochina is thus akin to that of Metcalfe given in multiple papers (e.g. Metcalfe, Reference Metcalfe1994, Reference Metcalfe2011, Reference Metcalfe2013 a,b), except that it excludes what that author refers to as the Simao subterrane (which here we consider approximately equivalent to the Ailaoshan terrane) that extends between the South China platform and the Baoshan portion of the Sibumasu terrane (Fig. 17) northwards towards the Yunling collage (Hughes et al. Reference Hughes, Peng and Luo2002) or Gongshan terrane (Song et al. Reference Song, Ji, Wei, Su, Zheng, Song and Zhang2007). The exclusion of the Simao terrane from Indochina follows recent findings that suggest that the Simao and Indochina terranes independently collided with South China early in the Triassic period, before they themselves annealed late during the same period (Roger et al. Reference Roger, Jolivet, Maluski, Respaut, Müncha, Paquette, Vũ and Nguyễn2014; Rossignol et al. Reference Rossignol, Bourquin, Poujol, Hallot, Dabard and Nalpas2016), implying a pre-Triassic history for the Simao terrane that was independent of other parts of Indochina (in this case the Trường Sơn terrane).
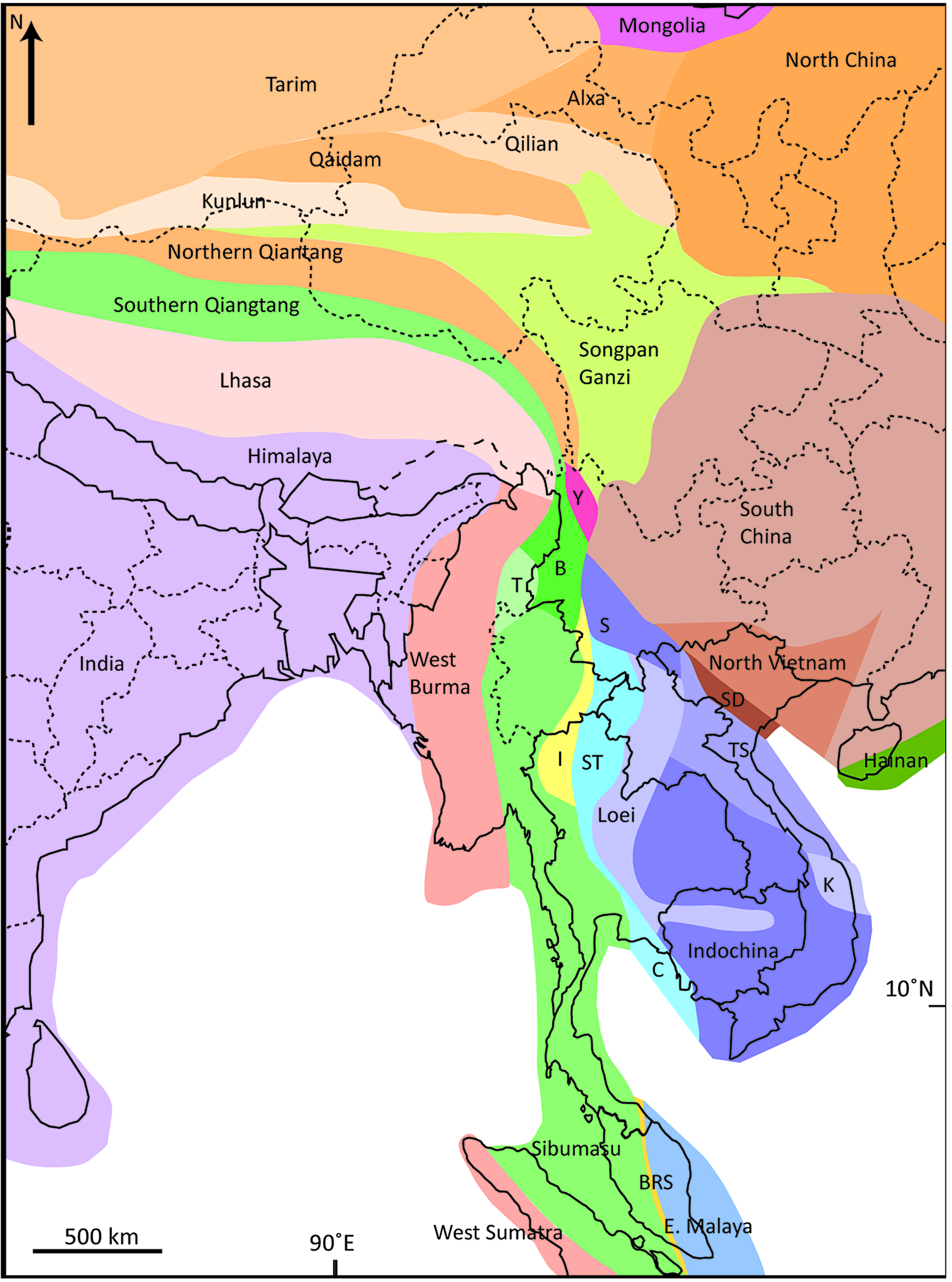
Fig. 17. Asian Equatorial Gondwanan terranes in their modern configuration. Modified from Metcalfe (Reference Metcalfe2011, Reference Metcalfe2017), Burrett et al. (Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014) and Loydell et al. (Reference Loydell, Udchachon and Burrett2019). B – Baoshan; BRS – Bentong-Raub Suture Zone; C – Chanthaburi; I – Inthanon; K – Kon Tum; S – Simao/Ailaoshan; ST – Sukhothai; TS – Trường Sơn; SD – Sông Đà; T – Tengchong; Y – Yunling. Here we show Indochina in the sense of Burrett et al. (Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014) with the Loei-Petchabun foldbelt included. The Palaeozoic terranes considered here to constitute Indochina include Loei, Trường Sơn, Kon Tum and the Khorat plateau region. We treat Simao as a different terrane (see text for explanation).
The pre-Permian history of Indochina is the subject of debate, with some authors seeing it as a coherent entity prior to that time (e.g. Metcalfe, Reference Metcalfe2017, fig 18; Torsvik & Cocks, Reference Torsvik and Cocks2017; Domeier, Reference Domeier2018, fig. 19; Li et al. Reference Li, Zhao, Liu, Cao, Yu, Li, Somerville, Yu and Suo2018, figs 7a, 20) while others stress an extended interval of terrane amalgamation during Palaeozoic time (e.g. Trần et al. Reference Trần, Khin, Halpin, Manaka, Meffre, Lai, Lee, Lê and Đinh2014, fig. 12; Shi et al. Reference Shi, Lin, Fan, Deng, Cong, Trần, Zhu and Wang2015, fig. 11; Loydell et al. Reference Loydell, Udchachon and Burrett2019, fig. 9; Nguyễn et al. Reference Nguyễn, Feng, Zi, Zhao, Trần, Ngô, Trần and Nguyễn2019, fig. 9) Accordingly, when discussing the pre-Permian history of the region that later became Indochina, we draw attention to the particular terrane in question (Fig. 17).
Most authors consider the Sông Mã suture to be the boundary between the South China block and the Trường Sơn terrane, i.e. part of what became Indochina (Carter & Clift, Reference Carter and Clift2008; Faure et al. Reference Faure, Lepvrier, Nguyễn, Vũ, Lin and Chen2014; Halpin et al. Reference Halpin, Trần, Lai, Meffre, Crawford and Khin2016; Wang et al. Reference Wang, Mo, Wang and Ye2016) (Fig. 1). This view is not universally accepted, with some preferring to locate that boundary south of the Trường Sơn terrane (Findlay & Phan, Reference Findlay and Phan1997; Findlay, Reference Findlay1998; Trần et al. Reference Trần, Khin, Halpin, Manaka, Meffre, Lai, Lee, Lê and Đinh2014; Phạm et al. Reference Phạm, Yang, Đỗ, Nguyễn, Lê and Chen2015) (Fig. 17). Others, who accept the Sông Mã fault to be the boundary between the South China block and the Trường Sơn terrane, also argue that the region immediately to the north of the Sông Hồng fault, i.e. the North Việt Nam block, was part of Indochina during Mesozoic time. According to this model, the Sông Đà terrane was a slice of South Chinese material that was emplaced via complex lateral movements associated with lateral slip during the Tertiary period (Cai & Zhang, Reference Cai and Zhang2009; Faure et al. Reference Faure, Lepvrier, Nguyễn, Vũ, Lin and Chen2014; Halpin et al. Reference Halpin, Trần, Lai, Meffre, Crawford and Khin2016). The Sông Mã suture is a collision zone that is typically dated as Permian–Triassic (Faure et al. Reference Faure, Lepvrier, Nguyễn, Vũ, Lin and Chen2014; Halpin et al. Reference Halpin, Trần, Lai, Meffre, Crawford and Khin2016; Wang et al. Reference Wang, Mo, Wang and Ye2016), although others see this as the reactivation of a collision that took place much earlier, possibly in Early Devonian time (Carter & Clift, Reference Carter and Clift2008).
Palaeontological evidence potentially offers important constraints on palaeographical reconstructions by providing evidence of association between regions that is independent of particular tectonic scenarios (Fortey & Cocks, Reference Fortey, Cocks, Hall and Holloway1998). While our results do suggest that the Sông Đà terrane and North Việt Nam block were conjoined during Cambrian time (cf. Cai & Zhang, Reference Cai and Zhang2009; Faure et al. Reference Faure, Lepvrier, Nguyễn, Vũ, Lin and Chen2014; Halpin et al. Reference Halpin, Trần, Lai, Meffre, Crawford and Khin2016), the similar depositional histories and comparable faunas of the Cambrian of the Sông Đà terrane and the East Bắc Bộ region of the North Việt Nam block provide no evidence of separation at that time. Rather, the most striking regional difference is the shelf–slope transition within the North Việt Nam block itself.
Palaeontological data constraining the early Palaeozoic positions of parts of Indochina are presently sparse and their interpretation is not straightforward. Based on the assumption that the Simao terrane was part of Palaeozoic Indochina, strong biotic ties between this region and South China have been suggested from the Ordovician (Zhou et al. Reference Zhou, Dean and Luo1998) through the early Silurian (Wang & Zhang, Reference Wang and Zhang2010), with sufficient proximity for conspecific land plant spores in both Simao and South China. Wang et al. (Reference Wang, Deng, Li, Li, Yu and Qiao2014) suggested an early Palaeozoic link between Simao and the Tethyan Himalaya based on detrital zircon age distributions, and here we note that their figure 3 shows a marked unconformity between the Cambrian and Ordovician, as also evident in the Himalaya, the Lhasa block and in Baoshan, but not in the remainder of Sibumasu (Wernette et al. Reference Wernette, Hughes, Myrow and Aye2021). However, as noted above, Simao apparently docked with the Trường Sơn terrane in the Triassic period, and so its earlier history is not directly relevant to the early location terranes that ultimately became part of Indochina.
Interpretation of data on Palaeozoic fossils from the Trường Sơn terrane is also challenging. A recent study (Loydell et al. Reference Loydell, Udchachon and Burrett2019) argued that during early Silurian time this terrane was likely situated towards the western end of the equatorial Gondwanan margin. Graptolites from Sepon mine in Laos included species commonly present at higher southern latitudes that are as yet unrecognized in other equatorial peri-Gondwanan terranes. In the later Silurian, shared occurrence of the shallow-water Retziella marine faunal assemblage in both the North Việt Nam terrane and the Trường Sơn of the late Silurian suggests their proximity (Tống et al. Reference Tống, Boucot, Rong and Fang2001), as does the shared occurrence of Early Devonian freshwater fish in both the Trường Sơn terrane and South China (Tống et al. Reference Tống, Janvier and Tạ1996, Reference Tống, Tạ, Boucot, Goujet and Janvier1997), but also see Loydell et al. (Reference Loydell, Udchachon and Burrett2019) for a considered review of these arguments. Early Palaeozoic coalescence of geologic fragments that were to become Indochina is suggested by evidence for Cambrian–Devonian suturing of the Trường Sơn terrane to the Kon Tum Massif (Usuki et al. Reference Usuki, Lan, Yui, Iizuka, Vũ, Trần, Okamoto, Wooden and Liou2009; Trần et al. Reference Trần, Khin, Halpin, Manaka, Meffre, Lai, Lee, Lê and Đinh2014; Thassanapak et al. Reference Thassanapak, Udchachon and Burrett2018; Nguyễn et al. Reference Nguyễn, Feng, Zi, Zhao, Trần, Ngô, Trần and Nguyễn2019; Wang et al. Reference Wang, Zhang, Qian, Senebouttalah, Wang, Wang, Gan and Khin2021), although others date this event as late Permian (Shi et al. Reference Shi, Lin, Fan, Deng, Cong, Trần, Zhu and Wang2015). Evidently, the earlier Palaeozoic history of the Indochinese terranes is not yet well resolved. Later Silurian and Devonian biotic similarities explain why in some recent regional tectonic reconstructions Indochina is shown joined to South China throughout early Palaeozoic time (Torsvik & Cocks, Reference Torsvik and Cocks2017; Domeier, Reference Domeier2018, figs 18–26). Presumably rifting produced a short-lived ocean basin between them that opened during early Carboniferous time (Torsvik & Cocks, Reference Torsvik and Cocks2017, fig. 9.4) and closed in late Triassic time (Torsvik & Cocks, Reference Torsvik and Cocks2017, fig. 11.2). This scenario may reconcile palaeontological evidence with structural data from the Sông Mã suture.
Evidence of early Palaeozoic magmatism and thermal events across various parts of what was to become Indochina (Roger et al. Reference Roger, Leloup, Jolivet, Lacassin, Phan, Brunel and Seward2000, Reference Roger, Maluski, Leyreloup, Lepvrie and Phan2007; Carter & Clift, Reference Carter and Clift2008; Zelázniewicz et al. Reference Zelázniewicz, Trần and Larionov2013; Nguyễn et al. Reference Nguyễn, Phạm, Trần, Bùi, Nguyễn and Đặng2014; Shi et al. Reference Shi, Lin, Fan, Deng, Cong, Trần, Zhu and Wang2015; Metcalfe, Reference Metcalfe2017) might relate in some way to the ‘Guangxi’ or ‘Kwangsian’ event (Carter et al. Reference Carter, Roques, Bristow and Kinny2001; Ou et al. Reference Ou, Lai, Carvalho, Zi, Jiang, Wang and Liu2021) that is also well represented in the eastern part of South China (Xu et al. Reference Xu, Cawood and Du2016). Zhan et al. (Reference Zhan, Jin, Rong, Zhu and Han2010) noted that the Cambrian strata of the Guole area of Guangxi Province, which borders northernmost Việt Nam, comprise shallow-water carbonate facies, although with respect to the general palaeogeography of South China, the area is surrounded by slope facies. This prompted these authors, building on a previous suggestion (Wu, Reference Wu2000), to propose that the southeastern Yunnan–south Guangxi–north Việt Nam area might be a fragment of continental material distinct from the South China block. This may be consistent with the view that the Dian–Qiong belt marks a major suture between northern Việt Nam and the South China block (Cai & Zhang, Reference Cai and Zhang2009). However, we do not see evidence that the Sông Đà terrane was more strongly allied with South China than with the North Việt Nam block, which is a claim of the Cai & Zhang (Reference Cai and Zhang2009) model. A thorough review of Vietnamese Cambrian faunas described by Mansuy (Reference Mansuy1915, Reference Mansuy1916), along with new collections, and faunal and detrital zircon comparisons of these with Cambrian sedimentary rocks of southeastern Yunnan (Luo et al. Reference Luo, Hu, Hou, Gao, Zhan and Li2009), Baoshan (Sun & Xiang, Reference Sun and Xiang1979; Luo, Reference Luo1982, Reference Luo1983, Reference Luo1985) Sibumasu (Shergold et al. Reference Shergold, Burrett, Akerman and Stait1988; Wernette et al. Reference Wernette, Hughes, Myrow and Sardsud2020 a,b), Bhutan (Hughes et al. Reference Hughes, Myrow, McKenzie, Harper, Bhargava, Tangri, Ghalley and Fanning2011), the Lesser (Hughes et al. Reference Hughes, Peng, Bhargava, Ahulwalia, Walia, Myrow and Parcha2005) and Tethyan Himalaya (Peng et al. Reference Peng, Hughes, Heim, Sell, Zhu, Myrow and Parcha2009; Hughes, Reference Hughes2016) and any fossil-bearing Cambrian or Ordovician rocks from Cambodia (Fromaget & Bonelli, Reference Fromaget and Bonelli1932) and Laos (Fromaget, Reference Fromaget1927, Reference Fromaget1941) will be instructive in evaluating this idea further. As several taxa from southeastern Yunnan, Sibumasu including Baoshan, the Himalaya and the northern Việt Nam region are congeneric, other informative associations among these areas may emerge.
10. Conclusions
The description of the fauna, facies and detrital zircon ages from lower Palaeozoic rocks of the Sông Đà terrane allows assessment of its regional geological history. Although data are limited, similarities between the later Cambrian and Tremadocian histories of the Sông Đà and North Việt Nam terranes do not suggest that the two areas were parts of different continental fragments, as some recent models suggest. Whether the Dian–Qiong suture marks an important Palaeozoic distinction between the South China block and northern Việt Nam (including southeastern Yunnan and south Guangxi provinces) merits further consideration.
Acknowledgements
Fieldwork in the region was supported by the US National Science Foundation EAR-053868 and EAR-1849963 to N.C.H. and by EAR-054334 and EAR-1849968 to P.M.M. This paper is a contribution to IGCP668, Equatorial Gondwanan History and Early Palaeozoic Evolutionary Dynamics. N.C.H. acknowledges receipt of Fulbright Academic and Professional Excellence Award 2019 APE-R/107 and thanks the Geological Studies Unit, Indian Statistical Institute, Kolkata for kindly hosting him, and Bernice Ridgeway of UCR Orbach Library for continual, prompt assistance. Dr Clive Burrett and another reviewer provided thoughtful, informative reviews that significantly strengthened the paper. Dr Nguyễn Dịu Hương kindly assisted with Vietnamese script. D.A.T.H. acknowledges support from the Leverhulme Trust. We thank Dr Võ Xuân Định for facilitating field arrangements and accompanying us. S.-C.P. and X.-J.Z. are funded by the State Key Laboratory of Palaeobiology and Stratigraphy, Chinese Academy of Sciences (XDB26000000, 20191101, 20190009) and National Natural Science Foundation of China (41672002, 41672028) and X.-J.Z.’s visit to UCR was funded by the Chinese Government Overseas Study Programme organized by Chinese Academy of Sciences. We thank Drs Mark Fanning and Ryan McKenzie for help with detrital zircon analysis. S.J.W. acknowledges the support of Geological Society of America, the American Museum of Natural History (Lerner-Gray Memorial Fund), the Evolving Earth Foundation, the American Association of Petroleum Geologists (David Worthington Named Grant) and the Paleontological Society (Allison R. ‘Pete’ Palmer Grant). We thank Dr Trương Quang Quý, director of the Bảo Tàng Địa Chất (DMR Geological Museum), and Drs Brenda Hunda and Cameron Schwalbach of the Cincinnati Museum Center for help with specimen collection and curation. Prof. Richard Fortey kindly commented on a taxonomic determination.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/S0016756821000844



















