Introduction
Ordovician cephalopods of the Xainza region, North Tibet, were first reported by the Tibet Geological Bureau General Survey Team (1980) during mapping of the Shigatse geological map sheet. Subsequently, many Ordovician cephalopods from this region were described by Lai (Reference Lai1982a, Reference Laib), Chen (Reference Chen1986, Reference Chen1987), Li and Cheng (Reference Li and Cheng1988), and in a series of works by Cheng et al. (Reference Cheng, Zhang and Zhang2005a, Reference Cheng, Zhang and Zhangb, Reference Cheng, Zhang and Zhangc, 2006). However, most of the recorded taxa were from the Upper Ordovician Keerduo and Gangmusang formations, which are both suggested as of Sandbian–Katian age by Xia (Reference Xia1983; Fig. 1.1), based on the outcrops mainly in the Xainza region. There is no conclusive evidence about the Middle Ordovician strata and the corresponding fossils in the Xainza area, northern Tibet (Cheng et al., Reference Cheng, Zhang and Zhang2004; Zhang et al., Reference Zhang, Cheng and Zhang2004).

Figure 1 (1) Location of the studied area in North Tibet, western China, with the tectonic sutures bounding the Lhasa Terrane (base map after Qu et al., Reference Qu, Zhang, Zheng and Wang2002; Xu et al., Reference Xu, Yang, Li, Ji, Zhang and Liu2011). (2) Lithostratigraphic and biostratigraphic subdivisions of the Ordovician in the Xainza Region in Tibet, western China; lithostratigraphic and biostratigraphic data from Xia (Reference Xia1983) and National Commission of Stratigraphy of China (2014). (3) Locality of the Lhasai section in Xainza County.
We have recovered nine species of actinocerid nautiloids, including one new species, from the Lhasai Formation, Middle Ordovician in Lhasai, Xainza County, northern Tibet. This is the first detailed taxonomic description of Middle Ordovician nautiloids from Xainza, which allows for more detailed stratigraphic correlation and better paleogeographic assessments.
Geological setting
The Xainza region is located in the northern part of the Lhasa Terrane, which is separated from the Himalaya Terrane by the Yarlung-Zangbo suture to the south and from the Qiangtang Terrane by the Bangon-Nujiang suture to the north (Fig. 1.2; Zhang et al., Reference Zhang, Cheng and Shen2010). The Middle Ordovician Lhasai Formation was first introduced during the geological investigations by the Jilin Institute of Geological Survey (Zhang et al., Reference Zhang, Qu and Zheng2003) in the Lhasai section, eastern Xiongmei town, in the sparsely populated northern part of Xainza County. The Ordovician in this section is represented, in ascending order, by the Middle Ordovician Lhasai Formation, the Middle to Upper Ordovician Keerduo and Gangmusang formations, and the overlying Silurian strata.
Among them, in the Lhasai Formation, abundant cephalopods, brachiopods, sponges, bryozoans, and conodonts are recorded (Qu et al., Reference Qu, Zhang, Zheng and Wang2002, Reference Qu, Wang, Zhang, Wang, Lü and Duan2004; Zhang et al., Reference Zhang, Qu and Zheng2003; Zheng, Reference Zheng2005), indicating a Middle Ordovician age. The unit is lithostratigraphically divided into three parts: (1) the lower part, consisting of limestone intercalated with some purple thin-bedded, silty, micritic limestone, with abundant cephalopods; (2) the middle part, consisting of limestone intercalated with yellow thin-bedded calcareous siltstone; and (3) the upper part, mainly consisting of bioclastic limestone, micritic limestone, and conglomeratic limestone.
Materials and methods
Most of the actinocerid specimens described herein were collected by some of the authors from the Lhasai section, which is located near Lhasai village, about 26 km east of Xiongmei town (31.2605°N, 89.1722°E to 31.2748°N, 89.1645°E), during the 2002 and 2012 investigations, which focused on the Middle Ordovician succession of the Xainza Region (Fig. 1.3).
All nautiloid shells are recrystallized and shell structures are poorly preserved. Due to poor preservation, only one specimen was found for each species. Most shells are slightly fragmented. All specimens were cut in median section, polished, and photographed with a Nikon D600 digital camera, Leica M125 microscope, and Leica DSC450C camera enhanced by Leica’s LAS software. Our identifications are based mainly on the position and shape of the siphuncle, the types of the septa and septal neck, the details of the connecting rings, the shape of the segment, and the endosiphuncular canal system, following Teichert et al. (Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erten, Moore and Zeller1964), Chao et al. (Reference Chao, Liang, Tsou, Lai and Chang1965), Flower (Reference Flower1968), Frey (Reference Frey1995), Zhu and Li (Reference Zhu and Li1996), and Niko and Sone (Reference Niko and Sone2014). The siphuncle segment compression ratio (SCR) is an important term introduced by Frey (Reference Frey1995) and is defined as the ratio of the maximum diameter of the connecting rings to the length of camerae.
To investigate quantitatively the paleogeographic affinity of the actinocerid nautiloids, the occurrence (presence or absence) data of 24 nautiloid genera from nine terranes or regions (Fig. 2; Table 1) are compiled into a binary data set. Cluster analysis (CA) methods in the PAST version 3.15 software (Hammer et al., Reference Hammer, Harper and Ryan2001) were adopted, using the Jaccard similarity coefficient (Jaccard, Reference Jaccard1901a, Reference Jaccardb), which is commonly used in paleobiogeographic studies (e.g., Shi, Reference Shi1993; Rong et al., Reference Rong, Li and Kul’Kov1995; Shen and Shi, Reference Shen and Shi2000, Reference Shen and Shi2004).

Figure 2 Global paleogeographic distribution of actinocerid nautiloids in the Middle Ordovician: 1. Xainza region, North Tibet; 2. Nyalam, South Tibet; 3. Inner Mongolia, North China; 4. Liaoning, North China; 5. Guizhou, South China; 6. Shan Plateau, Myanmar; 7. Southern Thailand; 8. Langkawi Islands, Malaysia; 9. Queensland, North Australia; 10. Tasmania (Tas. for short in the figure), Australia. Base map of paleogeographic reconstruction is modified from Burrett et al. (Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014). Light blue=relatively shallow platform and shelf seas; dark blue=relatively deep shelf seas; yellow=land or submarine highs.
Table 1 Occurrences (presence or absence) of actinocerid nautiloids in the Middle Ordovician. MYS=Malaysia (Kobayashi, Reference Kobayashi1958, Reference Kobayashi1959; Stait and Burrett, Reference Stait and Burrett1982); MMR=Myanmar (Niko and Sone, Reference Niko and Sone2014); THA=Thailand (Brown et al., Reference Brown, Buravas, Charaljavanaphet, Jalichandra, Johnston, Sresthaputra and Taylor1951; Kobayashi, Reference Kobayashi1958, Reference Kobayashi1959; Kobayashi and Hamada, Reference Kobayashi and Hamada1964; Burton, Reference Burton1974; Ingavat et al., Reference Ingavat, Muenlek and Udomratn1975; Stait and Burrett, Reference Stait and Burrett1984); HIM=Himalaya (Chen, Reference Chen1975; Chen, Reference Chen1983, Reference Chen1984); LHA=Lhasa (this paper); TAS=Tasmania (Teichert and Glenister, Reference Teichert and Glenister1953; Stait, Reference Stait1984); N AUS=North Australia (Teichert and Glenister, Reference Teichert and Glenister1952; Wade, Reference Wade1977); NC=North China (Chao et al., Reference Chao, Liang, Tsou, Lai and Chang1965; Chen and Zou, Reference Chen and Zou1984; Zhu and Li, Reference Zhu and Li1996); SC=South China (Yang, Reference Yang1978, Reference Yang1980).

Repository and institutional abbreviation
All specimens are deposited in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (NIGP).
Age of the actinocerid nautiloids of Lhasai Formation
The actinocerid nautiloids of Lhasai Formation are comparable to those from other Asian terranes (e.g., Qiangtang Terrane, Sibumasu Terrane, South and North China). According to the latest detailed restudy of the Ordovician Huaiyuan Epeirogeny in North China and adjacent regions, two distinct phases of the tectonic event are distinguished. The early phase was initiated from the Floian to early Darriwilian and is manifested by an extensive diachronous hiatus in the platform facies across North China (Zhen et al., Reference Zhen, Zhang, Wang and Percival2016). According to a recent restudy of the conodonts from the Ordovician of North China, several stratigraphic units that were regarded as mainly of early Ordovician and Dapingian age, including the Zhuozishan, the Sandaokan, and the Majiagou formations, are revised as Darriwilian (late Middle Ordovician) (Wang et al., Reference Wang, Bergström, Zhen, Zhang and Wu2014a, Reference Wang, Bergström, Zhen, Zhang and Wub, Reference Wang, Zhen, Zhang and Wu2016; Zhen et al., Reference Zhen, Zhang, Wang and Percival2016).
Actinocerid nautiloids of the Lhasai Formation comprise nine species in six genera, including Pomphoceras nyalamense (Chen, Reference Chen1975), P. yaliense (Chen, Reference Chen1975), Wutinoceras cf. W. foerstei (Endo, Reference Endo1930), Mesowutinoceras cf. M. giganteum Chen in Chen and Zou, Reference Chen and Zou1984, Armenoceras tani (Grabau, Reference Grabau1922), A. teicherti Endo, Reference Endo1932, A. xizangense new species, Deiroceras globosom Zou and Shen in Chen and Zou, Reference Chen and Zou1984, and Discoactinoceras cf. D. multiplexum Kobayashi, Reference Kobayashi1927. They all belong to the order Actinocerida. A majority of the species from Xainza are diagnostic of the Darriwilian (Middle Ordovician) according to a comparison with those from North China. For example, Pomphoceras is found in the Zhuozishan Formation of the Ordos Region (Chen and Zou, Reference Chen and Zou1984), the Majiagou Formation of southern Jilin, North China (Zhu and Li, Reference Zhu and Li1996), and the Chiatsun Formation of Nyalam, Himalaya Terrane (Chen, Reference Chen1975), which are all restricted to the Darriwilian. Wutinoceras was originally described from the Majiagou Formation (Wuting Formation; Darriwilian) in Benxi (Pen-his-hu) and Nanpiao, Liaoning, North China (Endo, Reference Endo1930, Reference Endo1932; Kobayashi and Matumoto, Reference Kobayashi and Matumoto1942), and has also been recorded in the Kakit Bukit Formation (partly Darriwilian) in the Langkawi Islands, Malaysia (Stait and Burrett, Reference Stait and Burrett1982; Stait et al., Reference Stait, Wyatt and Burrett1987), and in the Whiterock (Darrwilian) of North America (Flower, Reference Flower1968). Armenoceras is very common in the Middle Ordovician of North China (Endo, Reference Endo1932; Chao et al., Reference Chao, Liang, Tsou, Lai and Chang1965; Zhu and Li, Reference Zhu and Li1996) and was discovered in the Thung Song Group and the Tha Manao Formation of Thailand (Kobayashi, Reference Kobayashi1958, Reference Kobayashi1959; Stait and Burrett, Reference Stait and Burrett1984), the Whiterock part of the Kakit Bukit Formation of Malaysia (Kobayashi, Reference Kobayashi1958, Reference Kobayashi1959; Stait and Burrett, Reference Stait and Burrett1984; Stait et al., Reference Stait, Wyatt and Burrett1987), the Wunbye Formation of Myanmar (Niko and Sone, Reference Niko and Sone2014), the Middle Ordovician in Queensland and Tasmania, Australia (Whitehouse, Reference Whitehouse1936; Teichert and Glenister, Reference Teichert and Glenister1952), and in North America, Greenland, and Siberia, Russia (Balashov, Reference Balashov1962). Therefore, the age of the actinocerid nautiloids from the Lhasai Formation of Xainza is Middle Ordovician, and most probably Darriwilian.
Paleobiogeographic implications
The Xainza region is tectonically located close to the northern margin of the Lhasa Terrane, which was part of the northeastern peri-Gondwana region during the Early–Middle Ordovician (Metcalfe, 2001; Burrett et al., Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014). The Middle Ordovician nautiloid faunas of the Lhasa Terrane and some other Chinese terranes (e.g., the Himalaya and North China terranes) are characteristically dominated by actinocerids (e.g., Chen, Reference Chen1975; Chen and Zou, Reference Chen and Zou1984) and indicate that the nautiloids of these regions have a strong affinity. Several global Darriwilian cephalopod biogeographic regions were identified, including Laurentian, Rheic, East Asian (i.e., southern Tibet, Tarim, South and North China), and Siberia regions (Kröger, Reference Kröger2013). Drawing on specimens from the Xainza region, the detailed paleogeographic reconstruction of northeastern peri-Gondwana during the Middle Ordovician may now be firmly established.
Pomphoceras is a key element of the Zhuozishan Formation in Inner Mongolia (North China) and in the Chiatsun Formation in Nyalam (Himalaya Terrane). In addition, Pomphoceras nyalamense is present in both the Lhasa and Himalaya terranes (Chen, Reference Chen1975), and Pomphoceras yaliense is found in the North China, Lhasa, and Himalaya terranes (Chen, Reference Chen1975; Chen and Zou, Reference Chen and Zou1984). Pomphoceras resembles Ordosoceras in external form, inner construction, and systematic phylogeny; the latter is the Middle Ordovician index fossil of North China (Chang, Reference Chang1959; Chen and Zou, Reference Chen and Zou1984). Ordosoceras is also recorded from Myanmar (Sibumasu Terrane; Niko and Sone, Reference Niko and Sone2014), which indicates that the actinocerids from Sibumasu have close affinities with those of North China. Discoactinoceras is recorded in Inner Mongolia (Zou, Reference Zou1981) and Liaoning (Kobayashi, Reference Kobayashi1978), both from North China, so its occurrence in the Xainza Region of Tibet also supports a close relationship of the two terranes. Wutinoceras and Armenoceras are widespread genera and are common in the Himalaya Terrane (Chen, Reference Chen1975), Sibumasu (Stait and Burrett, Reference Stait and Burrett1982; Niko and Sone, Reference Niko and Sone2014), Australia (Stait, Reference Stait1984), and North America (Flower, Reference Flower1968). Some common elements of the actinocerids from Xainza are shared with North Australia (Teichert and Glenister, Reference Teichert and Glenister1952; Wade, Reference Wade1977), but few with Tasmania (Stait, Reference Stait1984). Nevertheless, there are few forms shared with Lhasa and South China during the Middle Ordovician (Chen and Liu, Reference Chen and Liu1976; Yang, Reference Yang1978, Reference Yang1980). Among them, the dominant form Meitanoceras is an endemic genus, which is only reported in the Middle Ordovician Meitan Formation from Chongqing, Guizhou, and Yunnan, South China. Ormoceras is a widespread genus, distributed in most parts of the world during this period. However, in general, there are few resemblances between the nautiloids of South and North China (Yang, Reference Yang1980).
The Middle Ordovician actinocerids of the Lhasa Terrane show close affinity with those of the Himalaya Terrane, North China, and the Sibumasu Terrane (Fig. 3). The similarities of the Middle Ordovician actinocerid nautiloids can be interpreted as reflecting similar paleogeographic situations in low paleolatitude; all these terranes are located in a northern tropical region. This also supports the Middle Ordovician reconstructions that place North China close to Australia and place South China close to the Middle East (e.g., Burrett and Stait, Reference Burrett and Stait1987; Burrett et al., Reference Burrett, Long and Stait1990, Reference Burrett, Khin, Meffre, Lai, Khositanont, Chaodumrong, Udchachon, Ekins and Halpin2014; Metcalfe, Reference Metcalfe2011) rather than the reconstructions that have these positions reversed (e.g., PaleoGIS 4.0 for ArcGIS, 2011). The affinities of nautiloids from these regions have a similar paleobiogeographic relationship to those of other marine organisms. For example, brachiopod affinities between Tibet and Sibumasu have been recognized from 230 genera of rhynchonelliform brachiopods from 65 localities during the Dapingian and Darriwilian (Harper et al., Reference Harper, Rasmussen, Liljeroth, Blodgett, Candela, Jin, Percival, Rong, Villas and Zhan2013).

Figure 3 Cluster analysis of the Middle Ordovician actinocerid nautiloids of peri-Gondwana using PAST (Hammer et al., Reference Hammer, Harper and Ryan2001) based on the Jaccard similarity coefficient.
Systematic paleontology
Class Cephalopoda Cuvier, Reference Cuvier1797
Subclass Nautiloidea Agassiz, Reference Agassiz1847
Order Actinocerida Teichert, Reference Teichert1933
Family Meitanoceratidae Kobayashi, Reference Kobayashi1977
Genus Pomphoceras Zou and Shen in Chen and Zou, Reference Chen and Zou1984
Type species
Pomphoceras wuhaiense Zou and Shen in Chen and Zou, Reference Chen and Zou1984
Pomphoceras nyalamense (Chen, Reference Chen1975)

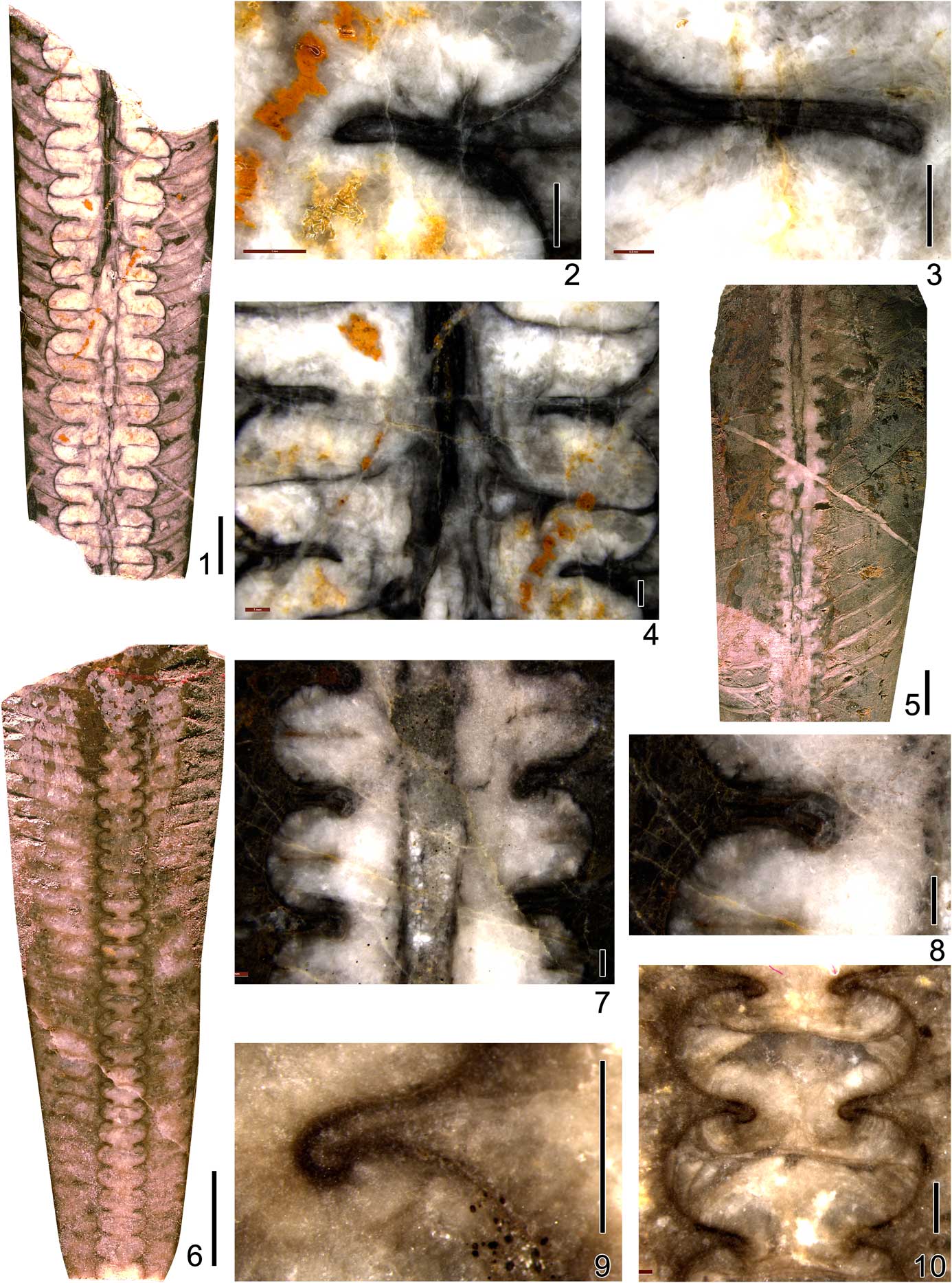
Figure 4 Illustration of siphuncular segments of actinocerid nautiloids from Lhasai Formation in Xainza, Tibet, showing the characteristics of different species: (1) Pomphoceras yaliense (Chen, Reference Chen1975), NIGP166263 (P25Hs-10-13-1); (2) Wutinoceras cf. W. foerstei (Endo, Reference Endo1930), NIGP166261 (qD2226-b1); (3) Pomphoceras nyalamense (Chen, Reference Chen1975), NIGP166262 (PM4-9-DH6); (4) Armenoceras tani (Grabau, Reference Grabau1922), NIGP166268 (qD0274-b1); (5) Armenoceras xizangense n. sp., NIGP166267 (PM4-9-DH3); (6) Armenoceras teicherti Endo, Reference Endo1932, NIGP166269 (PM4-9-DH4); (7) Deiroceras globosom Zou and Shen in Chen and Zou, Reference Chen and Zou1984, NIGP166264 (P25Hs-9-1); (8) Mesowutinoceras giganteum Chen in Chen and Zou, Reference Chen and Zou1984, NIGP166266 (6283-11); (9) Discoactinoceras cf. D. multiplexum Kobayashi, Reference Kobayashi1927, NIGP166265 (P25Hs-16-4). (1–3, 6, 8) Scale bars=5 mm; (4, 5) scale bars=1 cm; (7, 9) scale bars=1 mm.
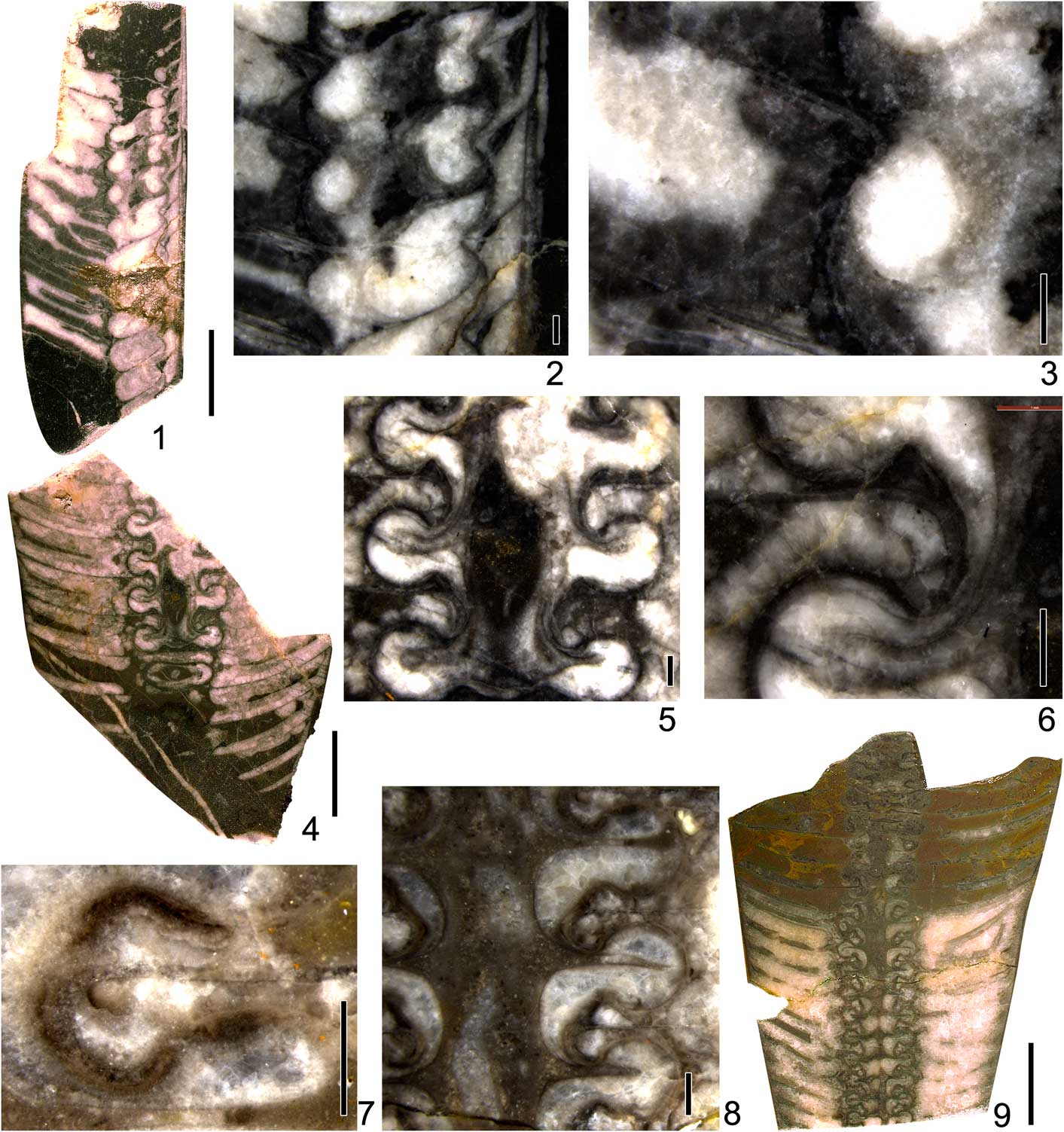
Figure 5 Photographs of Wutinoceras and Pomphoceras species from the Lhasai Formation in Xainza, Tibet. (1–3) Wutinoceras cf. W. foerstei (Endo, Reference Endo1930), NIGP166261 (qD2226-b1); (1) longitudinal polished section, side view; (2) general siphuncle shape and endosiphuncular deposits; (3) details of septal necks, connecting rings, and cameral deposits. (4–6) Pomphoceras nyalamense (Chen, Reference Chen1975), NIGP166262 (PM4-9-DH6); (4) longitudinal polished section, side view; (5) general siphuncle shape, vascular system, and endosiphuncular deposits; (6) details of septal necks, connecting rings, and radial canals. (7–9) Pomphoceras yaliense (Chen, Reference Chen1975), NIGP166263 (P25Hs-10-13-1); (7) details of septal necks and connecting rings; (8) general siphuncle shape, vascular system, and endosiphuncular deposits; (9) longitudinal polished section, side view. (1, 4, 9) Scale bars=1 cm; (2, 3, 5–8) scale bars=1 mm.
1975 Ordosoceras nyalamense Reference ChenChen, p. 272, text fig. 1, pl. 1, figs. 14, 15.
Occurrence
Lhasai Formation, Darriwilian (Middle Ordovician), near Lhasai village, about 26 km east of Xiongmei town, Xainza County, North Tibet.
Description
The orthocone is composed of nine camerae. It is about 47 mm in length and 38 mm in width, with a large expansion angle of 28.6°; conch cross section is dorsoventrally slightly depressed; external shell wall is not preserved. Septal curvature and cameral length are moderate; there are seven to eight camerae within the length of the corresponding lateral conch diameter; siphuncle is large, probably central in position. Siphuncular wall consists of cyrtochoanitic septal necks and thin connecting rings. Diameter of septal foramina is probably 4.2 mm. Septal necks are long and straight and brims are recumbent. Lengths of necks and brims are 1.8 mm and 0.8 mm, respectively. Maximum diameter of connecting rings is 11.6 mm, while the corresponding conch diameter is 37.8 mm. The anteriors of the connecting rings are curved and one-third part of the posterior is inflated and then progresses straight down obliquely; adnation area is narrow. SCR is 2.2. Cameral deposits well developed as episeptal-mural and hyposeptal deposits. The endosiphuncular canal system consists of a large central canal, radial canals, and perispatium. Central canal is probably 3.4 mm in diameter and distal ends of the radial canals are nearly horizontal.
Material
An incomplete orthocone cut and polished in median section. NIGP166262.
Remarks
According to Chen and Zou (Reference Chen and Zou1984), the conch of the genus Pomphoceras is slightly curved adapically, circular in cross section; siphuncle is small, central or subcentral. The segment shape of Pomphoceras is similar to that of Meitanoceras and Ordosoceras, but differs from Meitanoceras in having small septal foramina and from Ordosoceras in that its radial canals seem to be straight and directly transverse.
Pomphoceras nyalamense is similar to the type species of Pomphoceras, P. wuhaiense Zou and Shen in Chen and Zou (Reference Chen and Zou1984, p. 73, text fig. 23, pl. 15, figs. 3, 4) from Inner Mongolia, in its general siphuncular shape and central position of the siphuncle, but differs in that the former has a larger expansion rate.
Pomphoceras yaliense (Chen, Reference Chen1975)
1975 Ordosoceras yaliense Reference ChenChen, p. 274, pl. 1, figs. 5, 6.
1984 Pomphoceras yaliense; Reference Chen and ZouChen and Zou, p. 74, pl. 15, fig. 5, pl. 19, fig. 8.
Occurrence
Lhasai Formation, Darriwilian (Middle Ordovician), near Lhasai village, about 26 km east of Xiongmei town, Xainza County, North Tibet.
Description
An orthocone with 16 camerae is approximately 48.5 mm in length and 40.5 mm in width, with a high expansion rate and a dorsoventrally circular cross section. External shell wall is not available. Septa are curved and cameral length is moderate. There are 14 camerae in length of corresponding lateral conch diameter. Siphuncle is relatively large, central in position. Siphuncular wall consists of cyrtochoanitic septal necks and thin connecting rings. Diameters of septal foramina range from 3.2 to 3.8 mm. Septal necks are long and straight and brims are recumbent. The neck and brim length are 0.9–1.1 mm and 0.5–0.7 mm, respectively. The maximum diameters of connecting rings are 8.3–9.5 mm, which is approximately one-third of the corresponding conch diameters; the anterior of connecting rings are curved and one-third part of the posterior is inflated and then straight down obliquely. SCR is large and ranges from 3.1 to 3.5. Cameral deposits are well developed as episeptal-mural and hyposeptal deposits. The endosiphuncular canal system consists of a large central canal, radial canals, and perispatium. Central canal diameters are probably 0.7–2.5 mm; they are located centrally in the siphuncle. Distal ends of radial canals are nearly horizontal.
Material
An incomplete orthocone cut and polished in median section. NIGP166263.
Remarks
Chen (Reference Chen1975) erected Ordosoceras yaliense on the basis of an incomplete specimen from the Chiatsun Formation of Nyalam, southern Tibet. Its small septal foramina and straight and directly transverse radial canals indicate this species should be placed in the genus Pomphoceras (Chen and Zou, Reference Chen and Zou1984). P. yaliense has shorter septal necks and denser camerae than other species of Pomphoceras.
Family Wutinoceratidae Shimizu and Obata, Reference Shimizu and Obata1936 emend Flower, Reference Flower1968
Genus Wutinoceras Shimizu and Obata, Reference Shimizu and Obata1936
Type species
Nybyoceras foerstei Endo, Reference Endo1930
Wutinoceras cf. W. foerstei (Endo, Reference Endo1930)
1930 Nybyoceras foerstei Reference EndoEndo, p. 298, pl. 60, figs. 1a–c.
1932 Nybyoceras foerstei; Reference EndoEndo, p. 79, pl. 25, figs. 3–5, pl. 26, figs. 9, 10.
1933 Nybyoceras foerstei; Reference TeichertTeichert, p. 145, pl. 10, figs. 9, 10.
1936 Wutinoceras foerstei; Reference Shimizu and ObataShimizu and Obata, p. 263.
1957 Nybyoceras foerstei; Reference Yü and ChaoYü and Chao, p. 233, pl. 128, figs. 9, 10.
1965 Wutinoceras foerstei; Reference Chao, Liang, Tsou, Lai and ChangChao et al., p. 62, pl. 15, figs. 1, 2.
Occurrence
Lhasai Formation, Darriwilian (Middle Ordovician), near Lhasai village, about 26 km east of Xiongmei town, Xainza County, North Tibet.
Description
Specimen NIGP166261 is a 101 mm long fragment of an incomplete straight chamber, 65 mm in diameter, with conch cross section nearly circular. External shell wall is not preserved. There are 4.8 camerae in the length of the corresponding conch. The siphuncle is ventral in position. The siphuncle wall consists of long recumbent cyrtochoanitic septal necks and thick connecting rings. The ratios of maximum diameter of connecting ring per corresponding conch diameter are about 0.36. SCR ranges from 2.2 to 2.9. The diameter of the septal foramina is 4.6 mm. The brims are short, 0.4 mm in length, and septal necks are 0.8 mm in length. The segments connect with the bottom of the septa on the dorsal side and connect with the top of the septa on the ventral side. Cameral deposits are better developed at the episeptal-mural and hyposeptal positions in the anterior. Siphuncular deposits developed in the central canal. The central canal is probably 2.0 mm in diameter. Distal ends of radial canals are nearly horizontal with perispatial deposits.
Material
An incomplete orthocone cut and polished in median section. NIGP166261.
Remarks
The external construction, conch shape, and siphuncular characteristics of this specimen are the same as those of the type material of Wutinoceras foerstei (Endo, Reference Endo1930), but this specimen has the longer septal necks and narrower brim, which may be the result of bad cutting and polishing. Consequently, this specimen has been identified as Wutinoceras cf. W. foerstei (Endo, Reference Endo1930). The species is most similar to Wutinoceras lui Chang, Reference Chang1959 with regard to the position and construction of the siphuncle, except that the segments of W. lui are not in contact with the top of the septa on the ventral side. W. remotum Chen, Reference Chen1975 has sparser camerae with 3–3.5 camerae in the length of the corresponding conch and segments are not in contact with the bottom of the septa on the dorsal side. W. moeseini (Thein, Reference Thein1968) has a subventral siphuncle and longer brims, which differs from W. foerstei. W. robustum (Kobayashi and Matumoto, Reference Kobayashi and Matumoto1942) from the Darriwllian of Malaysia (Stait and Burrett,Reference Stait and Burrett1982) was assigned to W. moeseini (see Niko and Sone, Reference Niko and Sone2014) because the SCR of this specimen is approximately 2.0, which is smaller than that of W. foerstei.
Genus Mesowutinoceras Chen, Reference Chen1976
Type species
Mesowutinoceras discoides Chen, Reference Chen1976
Mesowutinoceras giganteum Chen in Chen and Zou, Reference Chen and Zou1984

Figure 6 Photographs of Deiroceras, Discoactinoceras, and Mesowutinoceras from the Lhasai Formation in Xainza, Tibet. (1–3) Deiroceras globosom Zou and Shen in Chen and Zou, Reference Chen and Zou1984, NIGP166264 (P25Hs-9-1); (1) longitudinal polished section, side view; (2) general siphuncle shape, central canals, cameral deposits, and endosiphuncular deposits; (3) details of septal necks and connecting rings. (4, 5) Discoactinoceras cf. D. multiplexum Kobayashi, Reference Kobayashi1927, NIGP166265 (P25Hs-16-4); (4) longitudinal polished section, side view; (5) details of septal necks and connecting rings. (6) Mesowutinoceras giganteum Chen in Chen and Zou, Reference Chen and Zou1984, NIGP166266 (6283-11), longitudinal polished section, side view. (1, 4, 6) Scale bars=1 cm; (2, 3, 5) scale bars=1 mm.
1984 Mesowutinoceras giganteum Chen in Reference Chen and ZouChen and Zou, p. 75, text-fig. 24, pl. 5, fig. 3, pl. 6, figs. 13, 14.
Occurrence
Lhasai Formation, Darriwilian (Middle Ordovician), near Lhasai village, about 26 km east of Xiongmei town, Xainza County, North Tibet.
Description
A large orthoconic fragment is available that is 29 mm in length and 20 mm in width, with only its dorsal side preserved. Siphuncle is subcentral in position. Siphuncular wall consists of long recumbent cyrtochoanitic septal necks and thick connecting rings. Brims are relatively long and adnation area is broad. Ratios of maximum diameter of connecting ring per corresponding conch diameter are about 0.48; segment is ellipsoid. Cameral deposits weakly developed at episeptal and hyposeptal positions. Siphuncular deposits are filling siphuncle. Central canal is unknown and perpendicular to radial canal. Distal ends of radial canals are anastomosing with perispatium near the most inflated portions of connecting rings.
Material
A large orthocone cut and polished in median section. NIGP166266.
Remarks
This specimen is very similar to M. giganteum described by Chen in Chen and Zou (Reference Chen and Zou1984) from the Ordos area because of its large size, subcentral siphuncle position, ellipsoidal segment, and recumbent septal necks, except that only the dorsal side of the specimen studied herein is preserved. This species differs from the type species M. discoides (Chen, Reference Chen1976) from the Shandong area in being larger and having a wider body size and wider segments.
Family Armenoceratidae Troedsson, Reference Troedsson1926
Genus Armenoceras Foerste, Reference Foerste1924
Type species
Actinoceras hearsti Parks, Reference Parks1913.
Armenoceras tani (Grabau, Reference Grabau1922)

Figure 7 Photographs of Armenoceras from the Lhasai Formation in Xainza, Tibet. (1–4) Armenoceras xizangense n. sp., holotype NIGP166267 (PM4-9-DH3); (1) longitudinal polished section, side view; (2, 3) details of septal necks; (4) general siphuncle shape, vascular system, and endosiphuncular deposits. (5, 7, 8) Armenoceras tani (Grabau, Reference Grabau1922), NIGP166268 (qD0274-b1); (5) longitudinally cut and polished section, side view; (7) general siphuncle shape, vascular system, and endosiphuncular deposits; (8) details of septal necks. (6, 9, 10) Armenoceras teicherti Endo, Reference Endo1932, NIGP166269 (PM4-9-DH4); (6) longitudinal polished section, side view; (9) details of septal necks and connecting rings; (10) general siphuncle shape, vascular system, and endosiphuncular deposits. (1, 5, 6) Scale bars=1 cm; (2–4, 7–9) scale bars=1 mm.
1922 Actinoceras tani Reference GrabauGrabau, p. 80, pl. 7, figs. 4–7.
1932 Armenoceras hatai Reference EndoEndo, p. 86, pl. 18, figs. 1–3.
1932 Armenoceras tani; Reference EndoEndo, p. 99, pl. 21, fig. 7, pl. 14, fig. 2, pl. 40, fig. 3.
1957 Armenoceras tani; Reference Yü and ChaoYü and Chao, p. 232, pl. 127, figs. 8, 9.
1959 Armenoceras cf. A. tani; Reference ChangChang, p. 265, pl. 1, fig. 4, pl. 3, figs. 2, 3.
1965 Armenoceras tani; Reference Chao, Liang, Tsou, Lai and ChangChao et al., p. 66, pl. 15, figs. 13, 14, pl. 18, figs. 5, 6.
Occurrence
Lhasai Formation, Darriwilian (Middle Ordovician), near Lhasai village, about 26 km east of Xiongmei town, Xainza County, North Tibet.
Description
Longiconic orthocone with gradual shell expansion and dorsoventrally depressed cross section. It is a phragmocone 98.1 mm in length and 48 mm in lateral diameter. External wall and body chamber are not preserved. There are nearly 8–9 camerae per corresponding lateral conch diameter. Siphuncle is large and subcentral in position. The diameters of foramina range from 5.6 to 7.1 mm at lateral conch diameter 38.2 to 4.79 mm. Septal necks are recumbent cyrtochoanitic and connecting rings are thick. SCR is approximately 2.4–2.6. The ratios of diameters of connecting rings to corresponding conch diameters are about 0.3. Brims are relatively long at 1.6 mm with 1.3 mm septal necks. Connecting rings show strong inflation with a very broad adnation area. Cameral deposits not well developed; siphuncle is filled with siphuncular deposits except for the vascular system, which is differentiated into central canals, radial canals, and perispatium.
Material
A longiconic orthocone cut and polished in median section. NIGP166268.
Remarks
Armenoceras tani (Endo, Reference Endo1932) appears similar to A. myanmarense Niko and Sone (Reference Niko and Sone2014) from the Shan Plateau, Myanmar, but the former differs in having more depressed segments and a narrower siphuncle, which is one-third of the corresponding conch diameter. A. manchurense Kobayashi (Reference Kobayashi1927) from Liaoning area is similar to A. tani except for the large siphuncle size and complex cameral deposits. A. tani shares several characteristics with A. coulingi (Grabau, Reference Grabau1922) from Shandong area (e.g., siphuncle diameter, cameral density, and septal concavity), but A. tani has a subcentral siphuncle and more-depressed siphuncular segments.
Armenoceras teicherti Endo, Reference Endo1932
1932 Armenoceras teicherti Reference EndoEndo, p. 89, pl. 19, fig. 1, pl. 40, fig. 6.
1965 Armenoceras teicherti; Reference Chao, Liang, Tsou, Lai and ChangChao et al., p. 67, pl. 16, fig. 8.
Occurrence
Lhasai Formation, Darriwilian (Middle Ordovician), near Lhasai village, about 26 km east of Xiongmei town, Xainza County, North Tibet.
Description
Specimen NIGP166269 is an almost complete orthoconic phragmocone with circular cross section; it is 67 mm in length and 24 mm at maximum lateral diameter within 29 camerae. It has a moderate expansion rate, with 11° in apical part. External features and body chamber are not preserved. There are 6, 8, and 10 camerae of corresponding conch diameter in the apical, middle, and adoral parts, respectively. The siphuncle is central in position. The siphuncular wall consists of recumbent cyrtochoanitic septal necks and thin connecting rings. Ratios of maximum diameter of connecting ring to corresponding lateral conch diameter are about 0.3. Siphuncular segment is discoidal and SCR is 1.8–2.2. Septal necks are 0.5 mm in length where brims are 0.8 mm and adnation area is broad. Cameral deposits are well developed in episeptal-mural and hyposeptal positions. Siphuncle is filled with endosiphuncular deposits. The vascular system is unknown.
Material
A complete orthocone cut and polished in median section. NIGP166269.
Remarks
Armenoceras tani (Grabau, Reference Grabau1922) is most similar to A. teicherti, but differs in having a depressed circular cross section and a subcentral siphuncle. A. coulingi (Grabau, Reference Grabau1922) has a larger body size than A. teicherti. A. concavum (Endo, Reference Endo1932) is different from A. teicherti in having higher camerae and smaller SCR.
Armenoceras xizangense new species
Holotype
Holotype NIGP166267 (A complete orthocone cut and polished in median section).
Diagnosis
Large siphuncle, occupying two-thirds of the corresponding later conch diameter, small SCR of approximately 4.
Occurrence
Lhasai Formation, Darriwilian (Middle Ordovician), near Lhasai village, about 26 km east of Xiongmei town, Xainza County, North Tibet.
Description
A large, longiconic orthocone with gradual shell expansion and dorsoventrally depressed cross section is available for study. Phragmocone is 101 mm in length and 37.2 mm in lateral conch diameter. External wall and body chamber are not preserved. There are 3.6–4.5 camerae per corresponding lateral diameter. Siphuncle is large and subcentral in position. The ratio of lateral diameter to length is 3.5. The siphuncular wall consists of recumbent cyrtochoanitic septal necks and thin connecting rings. Diameters of septal foramina are 7.5 and 9.3 mm at lateral diameters of 28.7 and 32.3 mm, respectively, where maximum diameters of connecting rings are 16.8 and 21 mm. Ratios of maximum diameter of connecting rings to corresponding lateral conch diameter are about 0.67. SCR is 2.2–2.3. Brims are relatively long, 1.7 mm in length where septal neck is 0.9 mm and adnation area is broad. Cameral deposits well developed in episeptal-mural and hyposeptal positions. Siphuncle is filled with deposits except for the vascular system. Radial canals are perpendicular to central canal. Distal ends of radial canals anastomosing with perispatium.
Etymology
The species name ‘xizangense’ is in reference to the origin of the holotype, Tibet, also called ‘Xizang’ in Chinese.
Remarks
This new species shares several characteristics with A. teicherti and A. submarginale (Grabau, Reference Grabau1922), for example, the moderate expansion and large siphuncle, but A. teicherti has a circular cross section and A. submarginale has a submarginal siphuncle, which distinguish them from A. xizangense. A. xizangense is most similar to A. magnitubulatum Endo (Reference Endo1932) with its large siphuncle size, but A. xizangense has a wider siphuncle occupying two-thirds of the lateral conch diameter rather than three-fifths as in A. magnitubulatum. A. magnitubulatum also has larger foramina and endocones. A. xizangense has 3.6–4.5 camerae per corresponding lateral diameter, less than the five camerae of A. centrale (Kobayashi and Matumoto, Reference Kobayashi and Matumoto1942), and A. centrale has a smaller siphuncle, which is less than one-half of the lateral conch.
Family Ormoceratidae Saemann, Reference Saemann1853
Genus Deiroceras Hyatt, Reference Hyatt1884
Type species
Orthoceras python Billings, Reference Billings1857
Deiroceras globosom Zou and Shen in Chen and Zou, Reference Chen and Zou1984
1984 Deiroceras globosum Zou and Shen in Reference Chen and ZouChen and Zou, p. 77, pl. 13, figs. 14, 15.
Occurrence
Lhasai Formation, Darriwilian (Middle Ordovician), near Lhasai village, about 26 km east of Xiongmei town, Xainza County, North Tibet.
Description
Specimen NIGP166264 is a longiconic orthocone with subcircular cross section; it is 31 mm in length and 11 mm in lateral conch diameter. It has moderate expansion, with 9° angle in apical part. External shell wall and body chamber are not well preserved. There are 3–4 camerae per corresponding lateral conch diameter. The concavity is nearly equal to the length of one camera. Siphuncle is subcentral in position. Ratios of maximum diameters of connecting rings to corresponding lateral conch diameter are 0.26. Siphuncular wall consists of short, recumbent cyrtochoanitic septal necks and thick connecting rings. Septal foramina are 0.7–0.9 mm. Septal necks are comparatively short and brims are weakly recumbent. Connecting rings are weakly inflated in the anterior part of each siphuncle segment. Siphuncular segment is gently rounded, with SCR approximately 0.8. Cameral deposits well developed as episeptal-mural and hyposeptal deposits in ventral side and as episeptal deposits on dorsal side. Siphuncular deposits well developed. Central canal is straight and weakly subdorsal; notches developed in the endosiphuncular deposits toward the posterior of the upper of the siphuncular segments, indicating the radial canals are better developed adoral to the septal foramina rather than apically.
Material
A longiconic orthocone cut and polished in median section. NIGP166264.
Remarks
According to Teichert et al. (Reference Teichert, Kummel, Sweet, Stenzel, Furnish, Glenister, Erten, Moore and Zeller1964) and Dzik (Reference Dzik1984), Deiroceras is most similar to Ormoceras except for the siphuncular segment shape, and Deiroceras has elongate connecting rings that are longer than wide.
This specimen has several similarities with the type specimen described by Zou and Shen (in Chen and Zou, Reference Chen and Zou1984) from Inner Mongolia , such as 3–4 camerae per corresponding lateral conch diameter, the concavity equal to one camerae length, and moderate siphuncle size.
Family Discoactinoceratidae Kobayashi, Reference Kobayashi1978
Genus Discoactinoceras Kobayashi, Reference Kobayashi1927
Type species
Discoactinoceras multiplexum Kobayashi, Reference Kobayashi1927
Discoactinoceras cf. D. multiplexum Kobayashi, Reference Kobayashi1927
1927 Discoactinoceras multiplexum Reference KobayashiKobayashi, p. 202, pl. 22, figs. 7a, b.
1931 Discoactinoceras multiplexum; Reference KobayashiKobayashi, p. 56, pl. 4, figs. 1a, b.
1965 Discoactinoceras multiplexum; Reference Chao, Liang, Tsou, Lai and ChangChao et al., p. 61, pl. 14, figs. 3–6.
1978 Discoactinoceras multiplexum; Reference KobayashiKobayashi, p. 226, text-figs. 1, 2, pl. 31, figs. 1–5.
1981 Discoactinoceras multiplexum Kobayashi in Reference ZouZou, p. 358, pl. 2, fig. 7.
Occurrence
Lhasai Formation, Darriwilian (Middle Ordovician), near Lhasai village, about 26 km east of Xiongmei town, Xainza County, North Tibet.
Description
The siphuncle is preserved and the orthoconic phragmocone with circular cross section is 21 mm in length. Siphuncle is large and central in position. Siphuncular foramina are broad. Siphuncle segments are about four times as wide as long. Septal necks are short and brims are short. Siphuncle is filled with deposits, and vascular systems are unknown.
Material
An orthoconic fragment cut and polished in median section. NIGP166265.
Remarks
Kobayashi (Reference Kobayashi1978) proposed a new family Discoactinoceratidae because the genus Discoactinoceras developed intrasiphuncular stereoplasmic deposits and recumbent cyrtochoanitic septal necks. This specimen (NIGP166265) is most similar to the specimen described by Zou (Reference Zou1981) from the Inner Mongolia area in its body size and large siphuncle.
Acknowledgments
We are indebted to C. Li (Jilin University) for providing several specimens and stratigraphic information. We are grateful to D. Evans and an anonymous reviewer for their constructive suggestions of the manuscript. Financial support from the Chinese Academy of Sciences (XDPB05, XDB10010100), the National Natural Science Foundation of China (grant No. 41290260, 41521061), and the Ministry of Science and Technology of China (2013FY111000) is acknowledged. This is a contribution to the IGCP 653 project (‘The Onset of Great Ordovician Biodiversification Event’).