


Introduction
Mosquitoes have been widely studied because of their role as vectors of several pathogens affecting humans and animals (Silver, Reference Silver2008). Mosquito population dynamics are of particular interest because mosquito abundance shapes the entomological risk of vector-borne disease transmission (Dye, Reference Dye1990). Specifically, vectorial capacity, the ability of a pathogen to be propagated by the action of insect vectors, linearly increases with vector-abundance (Garrett-Jones, Reference Garrett-Jones1964). Thus, the risk for disease transmission can be both spatially (Kitron, Reference Kitron1998) and temporally (Chaves et al., Reference Chaves, Hamer, Walker, Brown, Ruiz and Kitron2011) linked to vector abundance. Nevertheless, a detailed knowledge of factors regulating the abundance of mosquitoes, such as density-dependence and the impact of weather changes over a heterogenous landscape, have been relatively poorly studied for species other than dominant vectors (Chaves & Koenraadt, Reference Chaves and Koenraadt2010), with few exceptions (Yang et al., Reference Yang, Bradshaw, Whelan and Brook2008a ; Hoshi et al., Reference Hoshi, Higa and Chaves2014a ). Studies of mosquitoes over altitudinal gradients, in the context of climate change, allow to simultaneously evaluate mosquito abundance over the gradient of temperatures associated with elevation (Eisen et al., Reference Eisen, Bolling, Blair, Beaty and Moore2008), where, in general, temperature decreases with altitude. Similarly, information about mosquitoes with minor, or without any, medical importance is fundamental to understand possible evolutionary constraints on the response of mosquitoes, and the diseases they transmit, to climate change (Chaves & Koenraadt, Reference Chaves and Koenraadt2010).
The mosquito Armigeres subalbatus is a vector of filarial worms with medical importance, Brugia pahangi (Muslim et al., Reference Muslim, Fong, Mahmud, Lau and Sivanandam2013) and with veterinary importance, Dirofilaria repens (Lee et al., Reference Lee, Kim, Chong, Klein and Lee2007). Ar. subalbatus has also been found infected with Japanese Encephalitis Virus (JEV) (Tanaka et al., Reference Tanaka, Mizusawa and Saugstad1979) and has been incriminated as a JEV vector in settings without rice fields (Chen et al., Reference Chen, Dong, Chiou and Chuang2000). Regarding bloodfeeding, it has been reported that this mosquito was able to experimentally feed on humans, mice, chickens and reptiles, but not amphibians (Miyagi, Reference Miyagi1972), thus reinforcing the view that this mosquito can transmit zoonotic pathogens (Chaves et al., Reference Chaves, Harrington, Keogh, Nguyen and Kitron2010).
The mosquito Ar. subalbatus is widely distributed in Asia (Tanaka et al., Reference Tanaka, Mizusawa and Saugstad1979; Amerasinghe & Munasingha, Reference Amerasinghe and Munasingha1988a , Reference Amerasinghe and Munasingha b ) and has been observed across altitudinal gradients (Zea Iriarte et al., Reference Zea Iriarte, Tsuda, Wada and Takagi1991), yet no detailed study has assessed its population regulation. Studies on the natural history of Ar. subalbatus have shown that its larvae are common in nutrient enriched water (Senior-White, Reference Senior-White1925; Barr & Chellappah, Reference Barr and Chellappah1964), including septic tanks (Moriya et al., Reference Moriya, Harada and Yabe1967), especially with high ammonia concentration (Rajavel, Reference Rajavel1992b ), and the nutrient enriched water can be clear or turbid (Amerasinghe & Munasingha, Reference Amerasinghe and Munasingha1988b ). This mosquito also colonizes bamboo stumps (Kurashige, Reference Kurashige1963), artificial containers (Zea Iriarte et al., Reference Zea Iriarte, Tsuda, Wada and Takagi1991) and treeholes (Tsuda et al., Reference Tsuda, Takagi and Wada1994). Overwintering occurs in the larval stage (Mogi, Reference Mogi1996), and larvae seem to enter diapause because of a synergistic interaction between low temperature and short day length (Oda et al., Reference Oda, Wada, Kurokawa, Ueda and Ito1978). It has also been suggested, based on laboratory observations, that dry conditions may lead to egg diapause (Barr & Chellappah, Reference Barr and Chellappah1964). The phenology of adults also seems to be strongly seasonal, with latitudinal variation. For example, in Taiwan adult Ar. subalbatus were absent from mosquito traps during January and February (Sun, Reference Sun1964), at the higher latitude of Shandong province, China from November to early May (Zhang et al., Reference Zhang, Huang, Wang, Peng, Fu and Liu1992). At the even higher latitude of Kyoto, Japan adult mosquitoes have been only observed from August to October (Nakata & Ito, Reference Nakata and Ito1955). Rainfall has been suggested as an important factor for Ar. subalbatus population changes, with significant adult abundance increase during and after high rainfall (Amerasinghe & Munasingha, Reference Amerasinghe and Munasingha1988a ). Here, we present results from a season long study of Ar. subalbatus abundance patterns in Mt. Konpira in Nagasaki, Japan. We asked what factors of the landscape were correlated with Ar. subalbatus abundance across the altitudinal gradient of Mt. Konpira, and whether its population dynamics were under density dependence regulation while accounting for potential impacts of weather changes on its abundance.
Materials and methods
Study site
Larval and adult mosquitoes were biweekly sampled across 27 locations in Mt. Konpira (fig. 1). Mt. Konpira is located in Nagasaki city, northwest Kyushu, western Japan (fig. 1). Nagasaki has a temperate seasonal climate, with four well defined seasons. Temperature in the winter (from December to February) rarely drops below 0°C and snowfall is minimal, often absent. By contrast, the summer (mid July–mid September) has both high relative humidity, above 65%, and temperature, above 27°C (Isida, Reference Isida1969). Our sampling locations in Mt. Konpira followed those of a previous study in the same area, ranging from 109 to 330 m in altitude (Zea Iriarte et al., Reference Zea Iriarte, Tsuda, Wada and Takagi1991). The sampling setting consisted of three radial transects, across an altitudinal gradient, joined at a middle point (fig. 1). Our sampling locations were mainly located across a secondary forest, with diverse types of dominant vegetation according to the Japanese Ministry of the Environment (available at http://www.vegetation.biodic.go.jp/), but there were some urban sites in the periphery of all the sampling locations (fig. 1). Exact coordinates for each sampling location were measured with a Garmin Oregon 650 GPS (Garmin Ltd, KS, USA), which was also used to measure the site altitude, in metre. The altitude measures were compared with those from an ASTER digital elevation model (DEM), for Mt. Konpira (available at http://gdem.ersdac.jspacesystems.or.jp/), and given the low disagreement between the two measurements, we employed the DEM data in our analysis. At each sampling location we characterized the canopy cover in 25 May 2014 by following a standard methodology (Frazer et al., Reference Frazer, Fournier, Trofymow and Hall2001) where four photographs were taken with a fisheye ball lens located at ground level within a 1.5 m radius of a focal tree, i.e., a tree where an ovitrap was located. For the photos we used a EOS 40D camera (Canon Co., Tokyo, Japan) that had attached a normal lens (EF-S17-85 mm F4-5.6 IS USM; Canon Co., Tokyo, Japan) and a 180° fisheye lens (Nordward optronics 0.25X Super fisheye lens 180° G2; Revel Royal Inc, Aichi, Japan), which allowed to take hemispherical photos. The images were subsequently analyzed with the gap light analysis mobile application for android tablets (http://gap-light-analysis-mobile-app.android.informer.com/) to determine the average and standard deviation (SD) of canopy cover for each sampling location. We also quantified the ground cover by sampling twelve 2.5 m long transects radiating from each focal tree. Each transect had an approximate angular separation of 30°. We categorized each transect as: grass, leaf litter, bush, trees, concrete and terrace, based on the dominant group type among those six categories. A matrix with the percent of transects in each category at each sampling point was then analyzed with principal components analysis (Chaves et al., Reference Chaves, Hamer, Walker, Brown, Ruiz and Kitron2011), and the first principal component (online Table S1) was then used as a ground cover index.

Fig. 1. Map of sampling locations. The inset figure shows the location of Mt. Konpira in Nagasaki, Japan. Contour lines indicate the elevation. The inset legend indicates the land use types and vegetation cover, and there is also a guide to the total abundance of Armigeres subalbatus adults and fourth instar larvae sampled during May–November 2014. A dashed red line indicates a significant spatio-temporal cluster.
Mosquito sampling
To sample immature mosquito stages we made ‘ovitraps’ (Zea Iriarte et al., Reference Zea Iriarte, Tsuda, Wada and Takagi1991) by using 350 ml Coca-Cola® cans painted black with an acrylic spray paint (Kanpe Hapio Co., Ltd., Osaka, Japan) inside and outside. We made one 5 mm opening to hold both the can to a tree, using a cord, and to drain excess water above 280 ml. All traps were uniformly set at 1.2 m above the ground level. We selected this height given that previous reports indicated that Ar. subalbatus preferentially oviposits in containers around this height (Amerasinghe & Alagoda, Reference Amerasinghe and Alagoda1984). Ovitraps were filled with rain water collected during April 2014 and were set in 18 May, and then biweekly surveyed from 1 June until 29 November of 2014. During each survey we determined the presence of first to third instar mosquito larvae of any species, and we counted the number fourth instar larvae of Tripteroides bambusa Yamada and Ar. subalbatus, due to their easy identification in the field (Tanaka et al., Reference Tanaka, Mizusawa and Saugstad1979), and removed all other fourth instar larvae and all pupae for laboratory rearing and identification of emerged adults (Hoshi et al., Reference Hoshi, Higa and Chaves2014a ).
Adult mosquitoes were sampled using a sweep net (36 cm diam; Model 61-1B; Shiga Insect Co., Tokyo, Japan), a common tool to sample adult mosquitoes (Hoshi et al., Reference Hoshi, Imanishi, Higa and Chaves2014b ). Sampling started at sunrise. We selected this sampling time given previous reports that indicated this mosquito is active at sunrise (Berlin et al., Reference Berlin, Dwarakanath and Pandian1975). Also sweep nets are appropriate to sample this mosquito, which is predominant at ground level (Tsuda et al., Reference Tsuda, Maekawa, Saita, Hasegawa and Takagi2003). The use of sweep nets was standardized by staying at a fixed point within a 2.5 m radius from the focal tree where the ovitraps were set up. After arriving to a sampling location the net was swept around the body of the person sampling for 2 min, after this time period the person sampling stopped for 1 min and then swept the net for another 2 min. During each sampling session the order of the sampling locations was determined randomly in order to avoid any possible bias due to sampling the same locations always at the same time. Adult mosquitoes were biweekly sampled between 4 May and 15 November of 2014. We selected a biweekly sampling to have a sampling interval that coincided with the pre-adult developmental time of Ar. subalbatus (Weathersby, Reference Weathersby1962). Similarly, we started to collect mosquitoes in May, given that in Shandong Province, China, at the same latitude of Nagasaki, adult mosquitoes have only been caught after 10 May (Zhang et al., Reference Zhang, Huang, Wang, Peng, Fu and Liu1992).
Weather variables
At each sampling session we recorded the air temperature and relative humidity using a portable O-230 termohygrometer (Dretec Co., Saitama, Japan). The water temperature in the ovitraps was measured using an AD-5617WP infrared thermometer (A&D Co., Tokyo, Japan). Data on daily records for rainfall from 1 May to 1 December 2014 were obtained for Nagasaki city from the Japanese Meteorological Agency Website (http://www.jma.go.jp/jma/index.html). For the analyses we employed the cumulative amount of rainfall on the 14 days prior to the mosquito sampling. The weather station is located within a 5 km radius from our study site.
Mosquito identification
Fourth instar larvae and adult mosquitoes were identified using the taxonomic key of Tanaka et al. (Reference Tanaka, Mizusawa and Saugstad1979). Voucher specimens are available in the Entomological Collection in the Institute of Tropical Medicine of Nagasaki University, Japan and in the Mosquito Collection in the Walter Reed Biosystematics Unit – Smithsonian Institute, Washington DC, USA.
Statistical analysis
Larvae data of Ar. subalbatus were not analyzed given that we found them at two localities, once at one site, inside a vacant lot bordering a prickly forest, and two times at another other site, inside an area whose vegetation was dominated by acorns (fig. 1).
To detect spatio-temporal clusters, i.e., hotspots, of Ar. subalbatus adults, we employed an elliptical cluster detection mode in a SCAN spatio temporal Poisson model (Kulldorff & Nagarwalla, Reference Kulldorff and Nagarwalla1995; Kulldorff et al., Reference Kulldorff, Heffernan, Hartman, Assunção and Mostashari2005). We constrained the maximum cluster size up to 50% of the samples while scanning the whole study site. We chose the elliptical shape over a circular one given that our sampling followed transects (Kulldorff et al., Reference Kulldorff, Huang, Pickle and Duczmal2006). For statistical inference we employed 999 Monte Carlo simulations. We then proceeded with a spatial analysis of Ar. subalbatus abundance in Mt. Konpira, Nagasaki. We fitted Poisson generalized linear models (Faraway, Reference Faraway2006) to the total abundance per site. The total abundance per site was obtained by adding the abundance through the 14 biweeks of sampling at each sampling location (fig. 1). We started by building a model that included elevation (m), mean and SD canopy openness (%), mean, SD and kurtosis of temperature (°C) and relative humidity (%), and the ground cover index. This model was simplified by a combined backward-elimination and forward-addition algorithm (Kuhn & Johnson, Reference Kuhn and Johnson2013) that minimized the Akaike Information Criterion (AIC). Briefly, AIC is a model selection criterion that weighs the trade-off between the goodness of fit in a model and the number of parameters, and the best model is chosen by minimizing the difference between a function of a model log-likelihood and the number of parameters (Kuhn & Johnson, Reference Kuhn and Johnson2013). Moran's I test (Venables & Ripley, Reference Venables and Ripley2002) was employed to test the spatial independence of residuals in the model selected as best, in order to ensure that inferences from the best Poisson generalized linear model were sound (Chaves, Reference Chaves2010).
For the temporal analysis of Ar. subalbatus abundance we generated a 14 observations adult abundance time series, given the absence of adult mosquitoes during 4 May 2014. The time series was generated by adding the biweekly records across the 27 sampled sites per sampling session. We used this time series for some preliminary time series statistical analysis. We estimated the autocorrelation- (ACF) and partial autocorrelation (PACF)-functions to describe the patterns of temporal autocorrelation in Ar. subalbatus abundance (Chaves et al., Reference Chaves, Morrison, Kitron and Scott2012, Reference Chaves, Higa, Lee, Jeong, Heo, Kim, Minakawa and Lee2013). The ACF presents the correlation of a time series with itself through different time lags, while PACF shows a similar profile that only considers the correlation between consecutive time lags (Shumway & Stoffer, Reference Shumway and Stoffer2011). We also estimated the cross-correlation functions (CCFs), i.e., the temporal correlation function between two time series for different time lags (Shumway & Stoffer, Reference Shumway and Stoffer2011), between the Ar. subalbatus abundance time series and rainfall, temperature and relative humidity. We estimated the 95% confidence limits (CL), of the ACFs, PACFs and CCFs to test if the correlations departed from what would be expected by random, with only correlations outside the 95% CL being considered statistically significant (Shumway & Stoffer, Reference Shumway and Stoffer2011). Then, we proceeded to study the density-dependence regulation of Ar. subalbatus adults. We started by plotting the per-capita growth rate (r) of this mosquito as function of its total adult abundance (N t ). The per-capita growth rate is defined by (Turchin, Reference Turchin2003):
 $$r = \ln \left({N_t}\right) - \ln \left({N_{t - 1}}\right)$$
$$r = \ln \left({N_t}\right) - \ln \left({N_{t - 1}}\right)$$
Based on the results from this preliminary analysis, we fitted the Ricker model to the Ar. subalbatus time series data. The Ricker model has been widely used to study mosquito populations (Yang et al., Reference Yang, Bradshaw, Whelan and Brook2008a , Reference Yang, Brook, Whelan, Cleland and Bradshaw b ; Hoshi et al., Reference Hoshi, Higa and Chaves2014a ) and is defined by the following equation:
 $$N_t = \lambda_0 N_{t - {\rm 1}} {\exp} \left( { - bN_{t - {\rm 1}}} \right)$$
$$N_t = \lambda_0 N_{t - {\rm 1}} {\exp} \left( { - bN_{t - {\rm 1}}} \right)$$
Where λ0 is the intrinsic rate of population growth and b is a density-dependence coefficient (Turchin, Reference Turchin2003). When −b < 0, it can be affirmed that a population undergoes density-dependent regulation (Turchin, Reference Turchin2003). Further details about this model are presented elsewhere, including its derivation (Turchin, Reference Turchin2003; Mangel, Reference Mangel2006) and several stochastic versions (Melbourne & Hastings, Reference Melbourne and Hastings2008).
To fit the models to our data we assumed biweekly adult counts had a negative binomial (NegBin) distribution, given our observations had a variance larger than their mean, i.e., they were over-dispersed (Bolker, Reference Bolker2008). This assumption for the model presented in (2), when also assuming the model has an observation error, i.e., without feedback on future states (Bolker, Reference Bolker2008), coincides with the assumption that mosquito abundance follows a negative binomial environmental stochastic Ricker model (Melbourne & Hastings, Reference Melbourne and Hastings2008):
 $$\eqalign{N_t \sim NegBin({\rm mean} = \lambda _0 N_{t - 1} {\rm exp}\left( { - bN_{t - {\rm 1}}} \right),\;{\rm overdispersion} = k)}$$
$$\eqalign{N_t \sim NegBin({\rm mean} = \lambda _0 N_{t - 1} {\rm exp}\left( { - bN_{t - {\rm 1}}} \right),\;{\rm overdispersion} = k)}$$
which implies stochasticity affects individuals, in this case Ar. subalbatus mosquitoes, independently of their density. We also fitted the model assuming stochasticity was demographic, i.e., a function of mosquito density (Melbourne & Hastings, Reference Melbourne and Hastings2008):
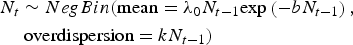 $$\eqalign{&N_t \sim NegBin({\rm mean} = \lambda_0 N_{t - 1} {\exp}\left({ - bN_{t - {\rm 1}}} \right),\cr &\quad\;{\rm overdispersion} = kN_{t - 1} )}$$
$$\eqalign{&N_t \sim NegBin({\rm mean} = \lambda_0 N_{t - 1} {\exp}\left({ - bN_{t - {\rm 1}}} \right),\cr &\quad\;{\rm overdispersion} = kN_{t - 1} )}$$
We expanded the model presented in (2) to account for the potential impacts of weather variables (WV) on the dynamics, as follows:
 $$N_t = \lambda_0 N_{t - {\rm 1}} {\exp}\left( { - bN_{t - {\rm 1}} + \gamma {\rm WV}_{t - z}} \right)$$
$$N_t = \lambda_0 N_{t - {\rm 1}} {\exp}\left( { - bN_{t - {\rm 1}} + \gamma {\rm WV}_{t - z}} \right)$$
Where γ is a coefficient for the impact of a WV at any time lag z. The lag z for the WVs was estimated with the CCFs previously described. To incorporate the forcing of equation (5) into equations (3) and (4), we simply changed the mean of the negative binomial to the following:
 $${\rm mean} = \lambda_0 N_{t - {\rm 1}} {\exp}\left( { - bN_{t - {\rm 1}} + \gamma {\rm WV}_{t - z}} \right)$$
$${\rm mean} = \lambda_0 N_{t - {\rm 1}} {\exp}\left( { - bN_{t - {\rm 1}} + \gamma {\rm WV}_{t - z}} \right)$$
To fit the model presented in (3) and its forced version (6), we employed a negative binomial generalized linear model (Faraway, Reference Faraway2006), using the natural logarithm of N t−1 as an offset, and N t−1 and WV t−z as covariates. Nevertheless, to fit the model presented in (4) and its forced version (6), we wrote an R function that is available as a supplementary online material (online Appendix S1). We computed the AIC of the models for comparison.
All statistical analysis was implemented with the statistical software R version 3.2.0, with the exception of the SCAN cluster analysis that was performed with SaTScan version 9.3.1.
Results
We collected a total of 114 adults and 21 fourth instar larvae of Ar. subalbatus during the duration of our study (fig. 1). Adults of Ar. subalbatus were present through all the land cover uses and vegetation types that we sampled, and were only absent from one of the points that we sampled (fig. 1). The average (±SD) number of adults per sampling location was 4.22 ± 2.35, ranging from 0, i.e., a place where Ar. subalbatus was never detected, to 9 individuals (fig. 1). Figure 1 also shows that adults of Ar. subalbatus were clustered across the northernmost sampling locations, which were all inside an area of the forest dominated by acorns.
The best spatial model showed that elevation and ground cover (table 1) were the best covariates explaining the spatial patterns of Ar. subalbatus abundance. The lack of significance for the Moran's I index (table 1) indicates spatial independence in the residuals, thus fulfilling the assumptions for a sound statistical inference (Chaves, Reference Chaves2010). Figure 2 clearly shows how the number of adult Ar. subalbatus decreased with elevation and increased with the ground cover index, where positive values indicate a dominance of leaf litter over bushes and/or concrete (Table S1 online only), while negative values indicate an abundance of bushes and/or concrete over leaf litter.

Fig. 2. Spatial patterns of Armigeres subalbatus abundance. Dot size is proportional to abundance of Ar. subalbatus, ranging from 1 to 9, as function of ground cover and altitude. The grey gradient indicates the abundance estimated with the model presented in Table 1.
Table 1. Parameter estimates for the best spatial Poisson Generalized Linear model explaining Armigeres subalbatus abundance across an altitudinal gradient. This model was selected by a combined backward-elimination and forward-addition algorithm, from a model that included elevation (m), mean and SD canopy openness, mean, SD and kurtosis of temperature (°C) and relative humidity (%), and a ground cover index. The Akaike Information Criterion (AIC) of the starting model was 124.05, and the AIC for the model presented here is 111.25.

* Statistically significant (P < 0.05).
Temporal patterns of Ar. subalbatus (fig. 3) showed that its abundance peaked during the 5th, 7th and 9th sampling biweek, i.e., 12 July, 9 August and 6 September, respectively, reaching a maximum of 34 individuals (fig. 3a). From the 13th biweek of sampling we did not catch any Ar. subalbatus. The average (±SD) number of adults per sampling biweek was 8.14 ± 10.23. In general, Ar. subalbatus abundance time series had temporally independent observations, as indicated by the lack of significant lags in its ACF (fig. 3b) and PACF (fig. 3c). Regarding fluctuations in the weather variables, it can be seen that peaks in mosquito abundance (fig. 3a) overlapped with those of rainfall (fig. 3d), but not those of temperature (fig. 3e) nor relative humidity (fig. 3f), nevertheless, these two last factors seem to be related with the presence of adult mosquitoes, which were only captured when their values were high. CCF analysis showed that Ar. subalbatus abundance was significantly associated with rainfall without a temporal lag (fig. 3g), but not with temperature (fig. 3h), nor with relative humidity (fig. 3i).

Fig. 3. Temporal patterns of Armigeres subalbatus abundance. (a) Time series of Ar. subalbatus (TSAS), biweekly abundance from 18 May to 15 November 2014. (b) Auto-correlation function (ACF), of TSAS, (c) partial ACF, PACF, of TSAS. Time series of weather variables: (d) rainfall, (e) temperature and (f) relative humidity (RH). Cross-correlation functions between TSAS and: (g) rainfall, (h) temperature and (I) RH.
As expected in a population undergoing density-dependent regulation (fig. 4) we found that the per capita growth rate of Ar. subalbatus decreased with its population size.

Fig. 4. Per capita population growth rate (r) of Armigeres subalbatus as function of its density, (N t−1), in Mt. Konpira, Nagasaki, Japan.
Estimates of the environmental negative binomial Ricker model (table 2) outperformed those of the demographic one (Table S2, online only). Table 2 shows the fit considering the forcing by rainfall was better, i.e., with a lower AIC, than an autonomous model. The effect of rainfall on the density dependent regulation of Ar. subalbatus (fig. 5) was to increase the number of mosquitoes, nevertheless, in all cases the abundance of Ar. subalbatus was under a strong density dependence, with the per capita growth rate diminishing with adult abundance in all instances.

Fig. 5. Temporal abundance of Armigeres subalbatus (N t ) as function of its previous abundance (N t−1) and rainfall. In the plot dots are proportional to the biweekly abundance of Ar. subalbatus (N t ), which ranged from 1 to 34. The grey gradient indicates the expected number of Ar. subalbatus according to the Ricker model forced by rainfall (for parameters see Table 2).
Table 2. Parameter estimates for the negative binomial environmental stochastic Ricker model fitted to the time series of Armigeres subalbatus biweekly abundance. Models were fitted with (Forced) and without (Autonomous) rainfall as covariate. AIC stands for the Akaike Information Criterion.

NS, not significant (P > 0.05), *Statistically significant (P < 0.05), **Statistically significant (P < 0.001).
Discussion
We found that temporal changes in adults of Ar. subalbatus were autonomous from changes in temperature and relative humidity. Nevertheless, these two factors may be important for the phenology of Ar. subalbatus adult activity, since we only found the species when temperatures were above 14°C and relative humidity above 65%. In fact, we found that phenology of adults of Ar. subalbatus at our study site was similar to observations at a similar latitude in China, where adult mosquitoes were only observed from May to October (Zhang et al., Reference Zhang, Huang, Wang, Peng, Fu and Liu1992). Moreover, our spatial model indicated that abundance of Ar. subalbatus adults decreased with altitude, further supporting that temperature might be important to the phenology and distribution of this mosquito, as suggested by observations across different latitudes (Nakata & Ito, Reference Nakata and Ito1955; Sun, Reference Sun1964; Zhang et al., Reference Zhang, Huang, Wang, Peng, Fu and Liu1992) and studies on its overwintering (Oda et al., Reference Oda, Wada, Kurokawa, Ueda and Ito1978). However, spatially we also found that adults of Ar. subalbatus were more common on grounds with abundant leaf litter, suggesting that organic matter necessary for enriching potential larval habitats (Senior-White, Reference Senior-White1925; Rajavel, Reference Rajavel1992b ) is a factor that might modulate habitat use by adults. This ground cover also resembles the habitats where this mosquito has been found transmitting filarial worms (Muslim et al., Reference Muslim, Fong, Mahmud, Lau and Sivanandam2013). Therefore, ground cover might be important to understand landscape heterogeneities in pathogen transmission risk by Ar. subalbatus.
The positive impact of rainfall on adult Ar. subalbatus abundance can be related to two aspects of its biology: (i) under dry conditions, eggs can enter diapause (Barr & Chellappah, Reference Barr and Chellappah1964) and (ii) the low variability in oviposition above the water surface, where more than 75% of the eggs were within a ‘threshold’ of 16 mm above the waterline (Amerasinghe & Alagoda, Reference Amerasinghe and Alagoda1984), suggest that any rainfall above the oviposition ‘threshold’ has the potential to synchronously trigger egg hatching in Ar. subalbatus larval habitats, which can lead to an eventual ‘outbreak’ or sudden large change in mosquito abundance (Chaves et al., Reference Chaves, Scott, Morrison and Takada2014), as we observed in our study site, a pattern also observed in Sri Lanka (Amerasinghe & Munasingha, Reference Amerasinghe and Munasingha1988a ). Unfortunately, the scarcity of larval samples did not allow us to connect any potential changes in larval density with adult abundance. However, a previous study (Rajavel, Reference Rajavel1992a ) suggests that at high larval density Ar. subalbatus keeps its numbers in check by cannibalizing larvae, especially if Ar. subalbatus does not co-occur with other mosquito species, and in that regard we can mention that we found fourth instar larvae of Ar. subalbatus only co-existing with larvae of Tp. bambusa, the most common and abundant mosquito in our ovitraps, following a similar pattern also observed by Zea Iriarte et al. (Reference Zea Iriarte, Tsuda, Wada and Takagi1991) in our study area. However, we were unable to asses any potential interaction between these two species given the scarce Ar. subalbatus larval data that we collected.
Our results clearly indicate that there is density dependence regulation in Ar. subalbatus. The density dependence parameter b in the Ricker model was negative as expected under density dependence regulation (Turchin, Reference Turchin2003), and in congruence with patterns observed in several mosquitoes (Makiya, Reference Makiya1973, Reference Makiya1974, Yang et al., Reference Yang, Bradshaw, Whelan and Brook2008a , Reference Yang, Brook, Whelan, Cleland and Bradshaw b , Chaves et al., Reference Chaves, Morrison, Kitron and Scott2012; Hoshi et al., Reference Hoshi, Higa and Chaves2014a ). However, although rainfall can transiently increase the number of adults, population growth still decreases with population abundance, highlighting the importance of density dependence regulation. It is also interesting to note that for the autonomous environmental stochastic Ricker model, the natural logarithm of the intrinsic rate of population growth parameter, i.e., ln(λ0) = 1.98, was close to the threshold value for weather independent oscillations, a.k.a., bifurcations, which is ln(λ0) ≥ 2 (Mangel, Reference Mangel2006). Nonetheless, when considering rainfall it became clear that any potential periodicities likely reflected rainfall patterns. Finally, we can conclude that population dynamics of adult Ar. subalbatus follows Schmalhausen's law, the prediction that systems are more sensitive to environmental variables more unpredictable around their mean (Chaves & Koenraadt, Reference Chaves and Koenraadt2010; Chaves et al., Reference Chaves, Morrison, Kitron and Scott2012). Rainfall, the only weather variable associated with abundance changes in Ar. subalbatus adults, undergoes less extreme changes in its mean behaviour than temperature or relative humidity at our study site, which although more extreme, have a more predictable pattern of change and seem to impact Ar. subalbatus phenology, but not its abundance dynamics.
Supplementary Material
The supplementary material for this article can be found at http://www.journals.cambridge.org/BER
Acknowledgements
We thank Sai Zaw Min Oo, Dr Hanako Iwashita and Prof. Mark Wilson (The University of Michigan) for their help measuring the ground cover and canopy openness at each sampling point. Trang T. T. Huynh also kindly helped us measuring diverse aspects of the sampling locations and with the larval surveys. Ms Junko Sakemoto provided valuable administrative help.









