



Introduction
Extant medusozoans (phylum Cnidaria) commonly show four-fold radial (tetraradial) symmetry, and most species exhibit alternating polypoid and medusoid life stages (Barnes, Reference Barnes1987). By contrast, fossil medusozoans from the Cambrian Fortunian Stage and Cambrian Stage 2, namely the olivooids, carinachitiids, and hexangulaconulariids, were sedentary polyps having a non- or weakly mineralized periderm exhibiting two-, three-, four-, five-, or six-fold radial symmetry (Liu et al., Reference Liu, Li, Shao, Zhang, Wang and Qiao2014, Reference Liu, Shao, Zhang, Wang, Zhang, Chen, Liang and Xue2017; Steiner et al., Reference Steiner, Qian, Li, Hagadorn and Zhu2014; Han et al., Reference Han, Kubota, Li, Ou and Wang2016b, Reference Han, Li, Kubota, Ou and Toshinoc, Reference Han, Li, Wang, Yang, Guo, Sasaki and Komiya2018; Guo et al., Reference Guo, Han, Van Iten, Song, Qiang, Wang, Zhang, Li, Sun and Sun2020a, Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Lib). Tetraradial pelagic medusae as living jellyfishes first appear in Cambrian Stage 3 (Cartwright et al., Reference Cartwright, Halgedahl, Hendricks, Jarrard, Marques, Collins and Lieberman2007; Young and Hagadorn, Reference Young and Hagadorn2010; Han et al., Reference Han, Hu, Cartwright, Zhao, Ou, Kubota, Wang and Yang2016a, Reference Han, Guo, Ou, Song, Liu, Hao, Sun and Wang2020). The periderm of all previously described olivooids and hexangulaconulariids consists of more or less distinct apical and abapical regions (Van Iten et al., Reference Van Iten, Zhu and Li2010; Duan et al., Reference Duan, Dong, Porras, Vargas, Cunningham and Donoghue2017). The three species of previously described hexangulaconulariids, Arthrochites emeishanensis Chen, Reference Chen1982, Hexaconularia sichuanensis He and Yang, Reference He and Yang1986, and Septuconularia yanjiaheensis Guo et al., Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020, differ from the inverted-pagoda-like olivooids in having the periderm strongly lenticular (transversely) and in lacking a set of well-developed longitudinal ridges converging on the distal end of the apical region. In addition, the hexangulaconulariids show variation in the number of faces, with H. sichuanensis and A. emeishanensis each having a total of six faces and S. yanjiaheensis possessing fourteen faces. The nature of the boundary between the faces also varies, consisting, for example, of a longitudinal furrow in H. sichuanensis (Van Iten et al., Reference Van Iten, Zhu and Li2010) and collinear ridges and furrows in S. yanjiaheensis (Guo et al., Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020b). Indeed, it would not be surprising if additional variants, perhaps intermediate between previously described species in the number of faces and the anatomy of the interfacial boundaries (and other features), existed as well.
Bed 5 of the Yanjiahe Formation near the city of Yichang, Hubei Province, China, has previously yielded the hexangulaconulariid S. yanjiaheensis (Guo et al., Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020b) and the olivooid Octapyrgites elongatus (Guo et al., Reference Guo, Han, Van Iten, Song, Qiang, Wang, Zhang, Li, Sun and Sun2020a). Here we describe a new hexangulaconulariid, Decimoconularia isofacialis new genus new species, from this same bed. The new fossil possesses a total of ten faces bounded by collinear longitudinal ridges and furrows. Our discovery thus expands the known range of gross anatomical variation exhibited by hexangulaconulariids and provides additional data bearing on the evolution of biradial hexangulaconulariids and possible homologies between these fossils and olivooids.
Materials and methods
Six specimens of Decimoconularia isofacialis n. gen. n. sp. (Figs. 1, 2) were collected from Bed 5 of the well-studied Yanjiahe Formation (Chen, Reference Chen1984; Guo et al., Reference Guo, Li, Han, Zhang, Zhang, Ou, Liu, Shu, Maruyama and Komiya2008, Reference Guo, Li and Li2014, Reference Guo, Han, Van Iten, Song, Qiang, Wang, Zhang, Li, Sun and Sun2020a, Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Lib, Reference Guo, Li, Qiang, Song, Zhang, Han and Wangc; Chang et al., Reference Chang, Clausen, Zhang, Feng, Steiner, Bottjer, Zhang and Shi2018, Reference Chang, Zhang, Clausen and Feng2020; Steiner et al., Reference Steiner, Yang, Hohl, Zhang and Chang2020) in the measured Yanjiahe section near the city of Yichang (Guo et al., Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020b, fig. 1.1–1.4). Bed 5 is composed of phosphatic dolostone and belongs to Cambrian Stage 2 (ca. 528 Ma; Zhu et al., Reference Zhu, Yang, Yuan, Li, Zhang, Zhao, Ahn and Miao2019). The specimens were picked under a binocular microscope from insoluble residues produced by digestion of rock samples with 10% acetic acid. The fossils were coated with gold and then imaged using a FEI Quanta 400 FEG scanning electron microscope (SEM) at an accelerating voltage of 15 eV (secondary electron mode). The holotype specimen (CUBar172-3) was imaged using the Zeiss Xradia 520 at the State Key Laboratory of Continental Dynamics, Northwest University, Xi'an, China, at a beam current/voltage of 80 μA/40 kV with a voxel size of 2.23 μm. The micro-computed tomography data were processed using VG Studio 2.2 Max for three-dimensional (3D) volume rendering and movies.
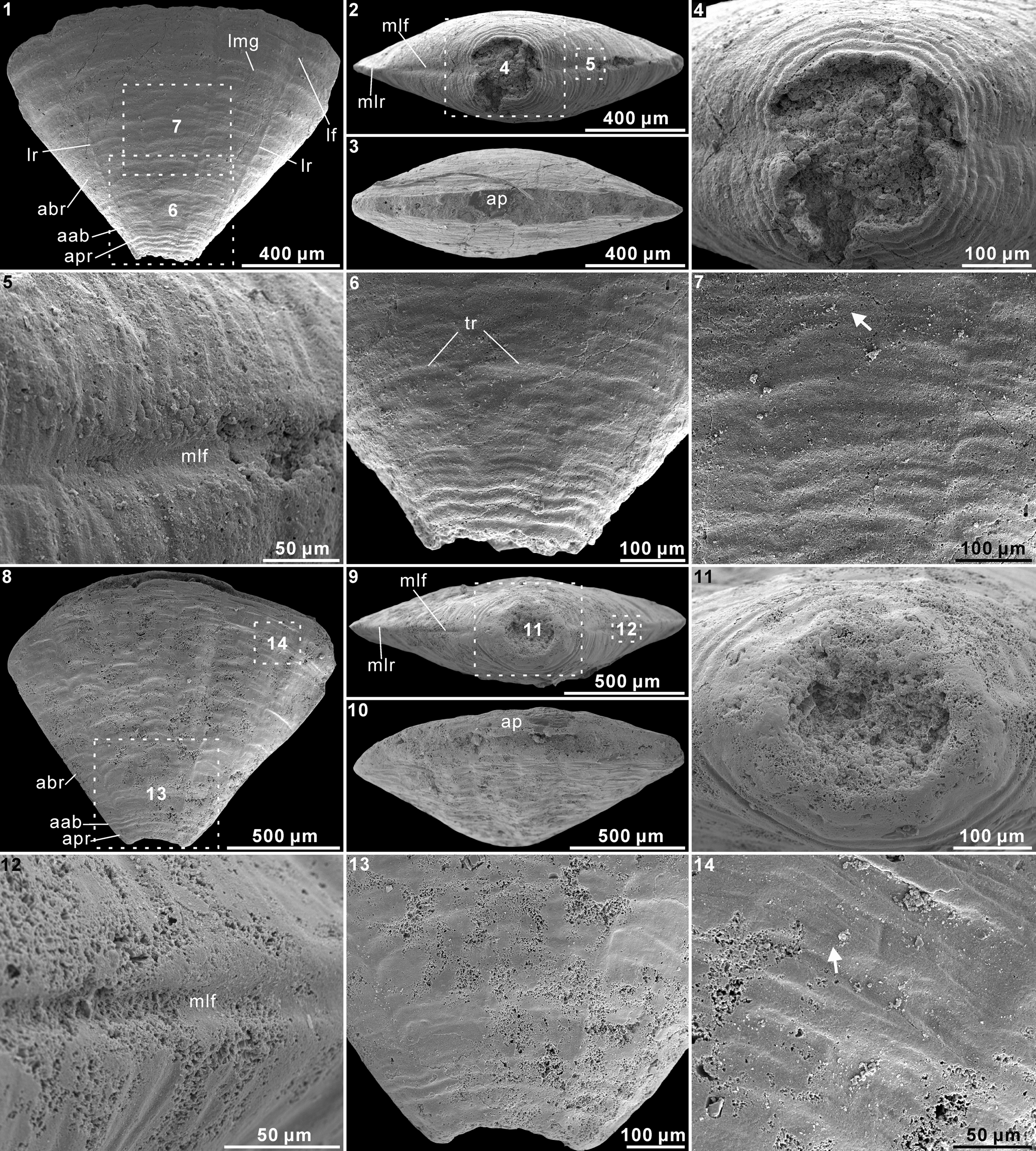
Figure 1. Decimoconularia isofacialis n. gen. n. sp. from the Cambrian Stage 2 Yanjiahe Formation (Bed 5) in the Three Gorges area near Yichang, Hubei Province, China. (1–7) Holotype CUBar172-3: (1) side view; (2) apical end view, showing the collinear marginal longitudinal ridges and furrows; (3) apertural end view; (4) close-up end view of the apical region, showing the continuous transverse ribs crossing the two marginal furrows, indicated by the number 4 in (2); (5) close-up view of one of the marginal longitudinal furrows, indicated by the number 5 in (2); (6) close-up side view of the apical region, showing the longitudinal ridges and continuous transverse ribs, indicated by the number 6 in (1); (7) close-up side view of the abapical region, showing bifurcation of a transverse rib (arrow), indicated by the number 7 in (1). (8–14) Paratype CUBar173-5: (8) side view; (9) apical end view, showing the collinear marginal longitudinal ridges and furrows; (10) apertural end view; (11) close-up end view of the apical region, showing the possibly concave base, indicated by the number 11 in (9); (12) close-up view of a marginal longitudinal furrow, indicated by the number 12 in (9); (13) side view of the apical region, showing the longitudinal ridges and continuous transverse ribs, indicated by the number 13 in (8); (14) close-up side view near the apertural margin of the abapical region, showing bifurcation of a transverse rib (arrow), indicated by the number 14 in (8). aab = apical–abapical boundary; abr = abapical region; ap = aperture; apr = apical region; lf = longitudinal furrow; lmg = longitudinal median groove; mlf = marginal longitudinal furrow; mlr = marginal longitudinal ridge; lr = longitudinal ridge; tr = transverse rib.

Figure 2. Decimoconularia isofacialis n. gen. n. sp. from the Cambrian Stage 2 Yanjiahe Formation (Bed 5) in the Three Gorges area near Yichang, Hubei Province, China. (1–4) Paratype CUBar146-4: (1) side view; (2) apical end view, showing the slightly concave base; (3) apertural end view; (4) close-up side view of the apical region, showing the longitudinal ridges, indicated by the number 4 in (1). (5–8) Paratype CUBar38-4: (5) side view, showing the longitudinal median groove on one of the faces; (6) apical end view, showing the collinear marginal longitudinal ridges and furrows; (7) apertural end view, showing the aperture; (8) close-up view of a longitudinal furrow and transverse rib, indicated by the number 8 in (5). (9–13) Paratype CUBar209-8: (9) side view; (10) apical end view; (11) apertural end view; (12) close-up end view of the apical region, showing the continuous transverse ribs crossing the two marginal furrows, indicated by the number 12 in (10); (13) close-up view of one of the lateral sides, showing the collinear marginal longitudinal ridge and furrow, indicated by the number 13 in (10). (14–16) Paratype CUBar209-7: (14) side view; (15) apical end view; (16) apertural end view. aab = apical–abapical boundary; abr = abapical region; ap = aperture; apr = apical region; lf = longitudinal furrow; lmg = longitudinal median groove; mlf = marginal longitudinal furrow; mlr = marginal longitudinal ridge; lr = longitudinal ridge; tr = transverse rib.
Repository and institutional abbreviation
All six specimens of D. isofacialis n. gen. n. sp. are housed in the paleontological collections of Chang'an University (CU) in Xi'an, China.
Systematic paleontology
Terminology
Anatomical terminology used herein mostly follows precedents to be found in Conway Morris and Chen (Reference Conway Morris and Chen1992), Van Iten (Reference Van Iten1992), and Han et al. (Reference Han, Li, Wang, Yang, Guo, Sasaki and Komiya2018).
Phylum Cnidaria Verrill, Reference Verrill1865
Subphylum Medusozoa Petersen, Reference Petersen, Larwood and Rosen1979
Subclass ?Conulata Moore and Harrington, Reference Moore and Harrington1956
Family Hexangulaconulariidae He, Reference He1987
Included genera
Arthrochites Chen, Reference Chen1982; Hexaconularia He and Yang, Reference He and Yang1986; Septuconularia Guo et al., Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020b; and Decimoconularia n. gen.
Emended diagnosis
Moderately to strongly tapered, laterally compressed, biradially symmetrical periderm bearing from six to fourteen faces exhibiting transverse ribbing and bounded by longitudinal furrows and/or ridges; wide oral end of the periderm open, apical end originally bluntly rounded. Relatively small apical region weakly to strongly differentiated from the rest of the periderm. Interspaces between the transverse ribs crossed by closely spaced, somewhat irregular longitudinal ridges.
Genus Decimoconularia new genus
Type species
Decimoconularia isofacialis n. gen. n. sp. from the Aldanella attleborensis Assemblage Zone (equivalent to the Watsonella crosbyi Assemblage Zone; Guo et al., Reference Guo, Li, Qiang, Song, Zhang, Han and Wang2020c) of the Yanjiahe Formation (Bed 5; Cambrian Stage 2) in the Yanjiahe section near the city of Yichang, Hubei Province, China.
Diagnosis
As for the species by monotypy.
Occurrence
The Yanjiahe Formation (Bed 5; Cambrian Stage 2) in the Yanjiahe section near the city of Yichang, Hubei Province, China; the Zhongyicun Member of the Zhujiaqing Formation (Cambrian Fortunian Stage), Huize, Yunnan Province, China (Qian et al., Reference Qian, Li, He and Xie2001).
Etymology
Decimo, Latin, referring to the ten faces of the periderm; conul-, Latin, cone-shaped periderm.
Remarks
The new genus currently comprises the type species only. Decimoconularia n. gen. differs from Arthrochites, Hexaconularia, and Septuconularia in having ten equally wide faces.
Decimoconularia isofacialis new species
Figures 1, 2
- Reference Qian, Li, He and Xie2001
Paradoxiconus typicalis (Type IV) Qian et al., p. 489, pl. I, figs. 9, 10.
Holotype
CUBar172-3 (Fig. 1.1–1.3; see supplemental data Movie 1); the A. attleborensis Assemblage Zone of the Yanjiahe Formation (Bed 5; Cambrian Stage 2) in the Yanjiahe section near the city of Yichang, Hubei Province, China.
Paratypes
Five relatively well-preserved specimens (CUBar173-5 [Fig. 1.8–1.10], CUBar146-4 [Fig. 2.1–2.3], CUBar38-4 [Fig. 2.5–2.7], CUBar209-8 [Fig. 2.9–2.11], CUBar209-7 [ Fig. 2.14–2.16]) from the same stratigraphic level and locality as the holotype.
Diagnosis
Periderm exhibits ten equally wide faces crossed by somewhat irregularly spaced, adaperturally curving, trochoidal (longitudinally) transverse ribs; five faces on each of the two major sides of the periderm bounded by a longitudinal furrow in about the apertural half of the periderm and by a collinear longitudinal ridge in about the apical half. Two major sides separated by a marginal ridge in about the apertural half of the periderm and by a collinear marginal furrow in about the apical half. Subdued, longitudinal median groove (midline) present on some of the faces. Short apical region weakly differentiated from the rest of the periderm and lacking lateral constrictions.
Occurrence
As for the type species.
Description
Strongly tapered, fan-shaped periderm with a narrowly lenticular transverse cross section. Specimens 1.132–1.456 mm wide and 0.982–1.198 mm long; width/length ratio approximately 6/5; apical angle in the plane of greatest width approximately 70°–80° (Table 1). External ornament defines a short, weakly differentiated apical region (apr) and a larger abapical region (abr) terminating along a slit-like apertural margin that appears to be incomplete (Figs. 1.1, 1.6, 1.8, 1.13, 2.1, 2.4, 2.9, 3.1). Distal tip of the apical region missing, although remnants of the periderm near the tip appear to be smooth. External ornament of the paratype specimens partially effaced, owing possibly to abrasion.
Table 1. Measurements of Decimoconularia isofacialis n. gen. n. sp.


Figure 3. 3D reconstructions of Decimoconularia isofacialis n. gen. n. sp. with hypothetical aperture, highlighting key anatomical features and terms used herein. (1–4) Side, marginal, apical end, and apertural end views, respectively. aab = apical–abapical boundary; abr = abapical region; am = apertural margin; ap = aperture; apr = apical region; cf = central face; lf = longitudinal furrow; lfs = lateral faces; lmg = longitudinal median groove; mf = marginal face; mlf = marginal longitudinal furrow; mlr = marginal longitudinal ridge; lr = longitudinal ridge; tr = transverse rib.
Two major sides of the periderm meet along a marginal longitudinal ridge (mlr; Figs. 1.2, 1.9, 2.2, 2.6, 2.10, 2.13, 2.15, 3.2, 3.3) in about the apertural half of the periderm and along a collinear marginal longitudinal furrow (mlf; Figs. 1.2, 1.5, 1.9, 1.12, 2.6, 2.10, 2.13, 2.15, 3.2, 3.3) in about the apical half, with the furrow being deep and extending far into the apical region. Each major side consists of five equally wide, adapically tapering faces (two marginal faces [mf], two lateral faces [lfs], and one central face [cf] [Fig. 3.1]) bounded by a longitudinal furrow (lf) in about the apertural half of the region (Figs. 1.1, 2.5, 2.8, 2.9, 3.1) and by a collinear longitudinal ridge (lr) in about the apical half (Figs. 1.1, 2.5, 2.9, 3.1); longitudinal ridges and faces extend far into the apical region, which, however, differs from the adapertural region in having the transverse ribs more pronounced (Figs. 1.1, 2.8, 2.9, 3.1). Angle of expansion of the central faces measures ~17°–20° (Table 1), with the weakly lobate apertural end of the faces being bent slightly toward the opposite side of the periderm (Figs. 1.3, 1.10, 2.3, 2.7, 2.11, 2.16, 3.2, 3.4). Some faces subdivided into two equal halves by a subdued, longitudinal median groove (lmg, midline) that extends from near the aperture to a level about midway between the aperture and the apical–abapical boundary (Figs. 1.1, 2.5, 2.9, 3.1). Faces crossed by irregularly spaced, trochoidal (longitudinally), adaperturally arching transverse ribs (tr) that number approximately 12–13 per 1 mm near the aperture; height of the transverse ribs variable but generally lower than that of the longitudinal ridges; most ribs extend across the entire width of the face (Fig. 1.6, 1.7), but some ribs bifurcate toward the midline (Fig. 1.7, 1.14). The continuous transverse ribs cross the two marginal longitudinal furrows in the apical region (Figs. 1.2, 1.4, 1.9, 1.11, 2.10, 2.12, 2.13).
Etymology
From the Latin, iso-, equi-, and faciem, face; isofacialis, equal-faced, denoting the ten equally wide faces of the fossil.
Remarks
Two additional specimens, both similar to D. isofacialis, were first described as Paradoxiconus typicalis (Type IV) by Qian et al. (Reference Qian, Li, He and Xie2001) from the Zhongyicun Member of the Zhujiaqing Formation (Fortunian Stage) at Huize, Yunnan Province, China (Qian et al., Reference Qian, Li, He and Xie2001; pl. I, figs. 9, 10). The photographs of these incomplete specimens show them to consist of a laterally compressed, biradially symmetrical periderm bearing ten equally wide faces exhibiting irregular transverse ribs and bounded by collinear longitudinal furrows and ridges. Therefore, we reinterpret these two specimens as being conspecific with the specimens here described. The other three types (Types I, II, and III) in Paradoxiconus differ from P. typicalis (Type IV) in being conical in shape, in having well-defined, closely spaced longitudinal ridges or sulci, and in exhibiting a surface ornament consisting of small spines or discontinuous nodes. It is therefore difficult to accept the hypothesis that P. typicalis (Type IV) and the other three types belong to the same species or even genus.
Decimoconularia isofacialis n. gen. n. sp. differs from other hexangulaconulariids in having ten equally wide faces. The new taxon is similar to Septuconularia yanjiaheensis in having the faces equal in size and devoid of longitudinal striations, but the latter species exhibits fourteen faces. Hexaconularia sichuanensis differs from D. isofacialis in being substantially smaller and in having only three faces on each major side of the periderm, with the central face being distinctly wider than the two faces on either side of it (Van Iten et al., Reference Van Iten, Zhu and Li2010, text-fig. 1; Steiner et al., Reference Steiner, Qian, Li, Hagadorn and Zhu2014). Also in H. sichuanensis, differentiation of the apical and abapical regions is much more pronounced than in D. isofacialis, with the regularly spaced transverse ridges in the abapical region of H. sichuanensis contrasting sharply with the irregular transverse wrinkles in the apical region. Furthermore, the longitudinal furrows of H. sichuanensis are wider and deeper than those of D. isofacialis, and in H. sichuanensis the marginal longitudinal ridges extend the full length of the abapical region as opposed to being restricted to about its apical half. In addition, the constrictions (Van Iten et al., Reference Van Iten, Zhu and Li2010; Duan et al., Reference Duan, Dong, Porras, Vargas, Cunningham and Donoghue2017) seen in the apical region of Hexaconularia are completely absent in Decimoconularia.
Finally, the longitudinal furrows in the abapical region of Hexaconularia become longitudinal ridges near the apical–abapical boundary. Septuconularia yanjiaheensis also exhibits an inconspicuous ridge–furrow transformation, which lies in level closer to the apertural margin (Guo et al., Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020b, fig. 2.4, 2.6, 2.14).
Discussion
Eventually, the problem of the phylogenetic relationships among hexangulaconulariids and their putative nearest relatives (carinachitids, conulariids, and olivooids; Guo et al., Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020b, fig. 8) will have to be investigated, as will the question of the significance of differences between these taxa in the morphology of the interfacial boundaries (simple furrow or ridge versus collinear furrow and ridge). While at present we are not prepared to address these problems at length, we do offer a modification of the preliminary phylogenetic hypothesis proposed by Guo et al. (Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020b, fig. 8). Olivooids differ from other sedentary medusozoans in having a flexible periderm without differentiation of face and sulcus. Specifically, we suggest that biradial symmetry and possession of a laterally compressed periderm are synapomorphies of hexangulaconulariids (Fig. 4). Biradial symmetry is interpreted as being derived relative to the more nearly radial symmetry of olivooids, carinachitids, and conulariids, although of course the symmetry of some Paleozoic conulariids (e.g., Conulariella) is strongly biradial (Van Iten et al., Reference Van Iten, Muir, Botting, Zhang and Lin2013). Hexaconularia and Arthrochites are interpreted as sister groups on the basis of the shared presence of six faces, two of which are centrally located and broader than the four faces flanking them. Decimoconularia and Septuconularia are united by possession of ten or more faces, faces devoid of longitudinal striations, as well as collinear longitudinal ridges and furrows. In keeping with the preceding diagnosis of Decimoconularia, the presence of ten faces is here reinterpreted (Fig. 4) as an autapomorphy of that genus.

Figure 4. Phylogenetic hypothesis for olivooids, carinachitiids, conulariids, and hexangulaconulariids (including Decimoconularia n. gen.) (modified from Guo et al., Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020b).
The origin of interfacial boundaries consisting of a collinear ridge and furrow in hexangulaconulariids remains an open question. Presumably they were derived from an ancestor in which the boundaries consisted of a simple ridge, a sulcus, or a collinear ridge and sulcus. Additional insight into this problem may be obtained by examining more closely the interfacial boundaries of other Cambrian sedentary medusozoans. For example, olivooids almost always exhibit sharp-crested longitudinal ridges on the apical embryonic part (see Steiner et al., Reference Steiner, Qian, Li, Hagadorn and Zhu2014); occasionally, however, an abnormal longitudinal ridge in Quadrapyrgites quadratacris Li, Reference Li1984 (Liu et al., Reference Liu, Li, Shao, Zhang, Wang and Qiao2014) exhibits a narrow groove in the apertural region of the periderm (e.g., the single corner in fig. 1.1 of Guo et al., Reference Guo, Han, Van Iten, Song, Qiang, Wang, Zhang, Li, Sun and Sun2020a). A single specimen of Octapyrgites elongatus exhibits apical ridges continuing adapertureward as inconspicuous linear structures crossing the annulation crests (Guo et al., Reference Guo, Han, Van Iten, Song, Qiang, Wang, Zhang, Li, Sun and Sun2020a, fig. 3.6, 3.10). Carinachitids consistently exhibit a pronounced interfacial (corner) sulcus, but unfortunately the apical region of these fossils is invariably missing (Conway Morris and Chen, Reference Conway Morris and Chen1992; Han et al., Reference Han, Li, Wang, Yang, Guo, Sasaki and Komiya2018). Whether the interfacial boundaries had any functional significance, for example in opening and closing the short apertural lappets (see the following), also remains an open question.
Last, Septuconularia and Decimoconularia from Bed 5 of the Yanjiahe Formation differ markedly in abundance, with 70 specimens of Septuconularia having been found thus far (Guo et al., Reference Guo, Han, Van Iten, Wang, Qiang, Song, Wang, Zhang and Li2020b) versus only six for Decimoconularia. This difference is unlikely to be taphonomic in origin as these two genera are mutually similar in size, structure, and composition and therefore probably also in preservation potential. It thus seems safe to infer that Septuconularia dominated Decimoconularia numerically in the original, shallow-shelf marine environment. Furthermore, it is tempting to speculate that this pattern may have had some causal basis in functional morphology. Specifically, each of the short facial lappets may have been associated with a longitudinal retractor muscle similar to that seen in lappet-bearing olivooids preserving relic soft tissues (Han et al., Reference Han, Kubota, Li, Yao and Yang2013, Reference Han, Li, Kubota, Ou and Toshino2016c). If indeed this was the case, then perhaps one reason for the apparent dominance of Septuconularia over Decimoconularia is that the more numerous and therefore smaller lobate folds of Septuconularia were easier to bend than the larger lobes of Decimoconularia. Greater ease of bending of the lappets may have made it easier to avoid predation or other disturbances through rapid retraction of the oral region of the soft polyp into the interior of the periderm, the strongly lenticular shape of which may itself have been an adaptation for life in a relatively high-energy bottom environment (Guo et al., Reference Guo, Han, Van Iten, Song, Qiang, Wang, Zhang, Li, Sun and Sun2020a).
Conclusions
The present discovery expands the known range of gross anatomical variation exhibited by members of the family Hexangulaconulariidae, which occurs exclusively in strata of early Cambrian age. The new genus Decimoconularia, distinguishable from other hexangulaconulariids in having a total of ten equally wide faces, may be the sister genus of Septuconularia. The periderm of the most recent common ancestor of hexangulaconulariids may have exhibited tetraradial or hexaradial symmetry, an apical region with four or six longitudinal ridges or furrows, and lobate apertural folds or lappets separated by radial grooves. Further prospecting in early Cambrian strata bearing small shelly fossils may well yield additional variants of this distinctive and intriguing group of ancient medusozoans.
Acknowledgments
We thank X. Liu and J. Sun (Northwest University, China) for preparation of the 3D drawings and microtomographic reconstruction of Decimoconularia. We are grateful to G. Young and one anonymous reviewer for their constructive comments and suggestions. This investigation was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (no. XDB26000000), the Natural Science Foundation of China (nos. 41890844, 41890840, 41621003, 41772010, 41720104002), Key Scientific and Technological Innovation Team Project in Shaanxi Province, State Key Laboratory of Palaeobiology and Stratigraphy (Nanjing Institute of Geology and Palaeontology, CAS) (nos. 203106, 163107), and the Ministry of Science and the Overseas Expertise Introduction Centre for Discipline Innovation Project (111) of the Ministry of Education of China (no. D17013).
Data availability statement
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.sqv9s4n38.





