1. Introduction
During Cenozoic times, European freshwater ecosystems hosted a number of crocodylian taxa. Many of them were ubiquitous across Europe and reported from Palaeocene and Eocene strata. Among these, the basal alligatoroid genus Diplocynodon has the longest stratigraphic record, ranging from the Early Eocene to the Middle Miocene. This genus has been recognized for a long time, its first description dating from the mid-1800s with the first report of Diplocynodon ratelii from the Early Miocene of Saint-Gérand-le-Puy in Allier, France (Pomel, Reference Pomel1847). Since then, more species were erected, and the last comprehensive review of the genus accepted eight species (Berg, Reference Berg1966). Recently, reports or attempts at taxonomic clarification were also presented (Vignaud et al. Reference Vignaud, Brunet, Guevel and Jehenne1996; Ginsburg & Bulot, Reference Ginsburg and Bulot1997; Brinkmann & Rauhe, Reference Brinkmann and Rauhe1998; Del Favero, Reference Del Favero1999; Hua, Reference Hua2004), and some efforts have concentrated on including species of Diplocynodon into a phylogenetic framework (Buscalioni, Sanz & Lourdes, Reference Buscalioni, Sanz and Lourdes1992; Brochu, Reference Brochu1999; Piras & Buscalioni, Reference Piras and Buscalioni2006). Despite that, the genus Diplocynodon still encompasses a fairly large number of species; because of this and its long stratigraphic record, the picture of its evolution is rather far from being fully resolved. A very large number of vertebrate-bearing localities are recorded throughout the Cenozoic, but few have yielded complete crocodylian remains, which would provide sufficient basis for taxonomic assessment. Many crocodylian remains are fragmentary and it is questionable whether efforts have concentrated mostly on small remains, namely mammal or herpetofaunistic assemblages to the detriment of larger vertebrates. Nevertheless, crocodylian skull fragments or isolated teeth and osteoderms are not commonly diagnostic beyond an ordinal level. Depending on the energetic nature of a deposit, such remains may still be abundantly recovered through sieving but will not yield sufficient taxonomic information in comparison to mammal teeth.
Thus, any additional material from a new or already known locality is of importance, as remains identified to the species level present a less complete stratigraphic picture among crocodylians than among diminutive mammals. Despite this fact, their fossil record is potentially promising, thanks to a depositional environment offering favourable conditions to their preservation. Moreover, in exceptional cases, articulated and exquisitely preserved specimens were recovered from Germany (Messel and the Geiseltal) and Italy (Monte Bolca). Such Lagerstätten provide excellent snapshots of diversity, but other types of preservation including accumulation in river channels are nevertheless informative. Landmark localities having yielded the best material are regionally limited to a handful of outcrops for a given time interval. This highlights one of the major problems regarding the continental fossil record: it is discontinuous, therefore, any new fossiliferous locality is of importance.
The present report describes a new Diplocynodon taxon from the terminal Eocene of Massif Central, France. The locality of Domérat, Allier (Fig. 1), which has yielded it, was dated on its mammal assemblage to the Mammal Palaeogene level MP18–20 (Meloux et al. Reference Meloux, Remy, Sudre, Turland and Vianey-Liaud1996).

Figure 1. Geographic position of the locality of Domérat, Massif Central in the ‘département de l'Allier’, France.
Despite several mentions from North America (Mook, Reference Mook1960; Grandstaff et al. Reference Grandstaff, Parris, Denton and Gallagher1992; Parris et al. Reference Parris, Grandstaff, Denton and Dobie1997) and from Asia (Efimov, Reference Efimov1988; Guanbang & Guikun, Reference Guanbang and Guikun1993), the genus Diplocynodon is, after reappraisal, thought to be endemic to Europe (Brochu, Reference Brochu1999; Piras & Buscalioni, Reference Piras and Buscalioni2006) and is known to have survived important climatic changes from a greenhouse Palaeogene to an icehouse period in the early Neogene. A change in crocodylian faunal composition was observed during this time interval. Ecosystems hosting several taxa shifted to ecosystems with a single taxon. However, any link between these observations remains unclear, and hypotheses for crocodylian faunal turnover at this period will be discussed below.
2. Geological setting
The département de l'Allier has rich vertebrate-bearing localities, including the Lower Miocene vertebrate fauna of Saint-Gérand-le-Puy in the Limagne basin, from where the holotype of Diplocynodon was recovered (Pomel, Reference Pomel1847). The older locality of Domérat is located in the western part of Allier in the smaller Montluçon–Domérat basin, separated from the Limagne basin by the northern crystalline and volcanic extension of the Massif Central horst (Fig. 1). The establishment of these basins is linked to the regional tectonic activity. The Domérat–Montluçon basin is delimited on its eastern margin by the Cher River fault with a N–S direction; therefore the basin follows this orientation.
A brief sedimentary study was provided by Thiry et al. (Reference Thiry, Schmitt, Trauth, Cojean and Turland1983), who characterized successions of palaeopedogenetic levels and detrital deposits. Among these deposits, lenses of dark green clay occur and have yielded the Late Eocene vertebrate fauna of Domérat. According to Meloux et al. (Reference Meloux, Remy, Sudre, Turland and Vianey-Liaud1996), the mammal assemblage, with Palaeotherium, Plagiolophus, Anoplotherium, Xiphodon and Amphimeryx, indicates a Ludian (Middle Priabonian) age corresponding to the MP18–MP20 reference level of the mammalian biochronological scale for the Palaeogene of Europe (BiochroM, Reference Aguilar, Legendre and Michaux1997).
Mammal remains reported from this locality essentially consist of isolated teeth, as well as rolled and often incomplete bones. The recovery of pebbles led Meloux et al. (Reference Meloux, Remy, Sudre, Turland and Vianey-Liaud1996) to suggest a flash-flood type of burial, which would have trapped the mammals at a drinking spot in a marshy environment. Vertebrate remains were eventually transported and sorted. The present report of a Diplocynodon skull does not seem to fit such a scenario. Despite poor preservation, it is complete and the recovery of bones in connection with the associated mandible is not in agreement with any long transport. The bones sometimes appear to be heavily weathered and turned into a yellowish oxidized dust. It is possible, however, that mammal bones were transported and sorted, as the depositional environment may correspond to a river channel. In any case, the presence of the complete Diplocynodon skull provides evidence that fossil remains recovered from that locality underwent different taphonomic processes.
Institutional abbreviations: Rhinopolis – Association Rhinopolis, Gannat, Allier, France; BMNH – British Museum of Natural History, London.
3. Systematic palaeontology
CROCODYLIA Gmelin, Reference Gmelin1789 (sensu Martin & Benton, Reference Martin and Benton2008)
EUSUCHIA Huxley, Reference Huxley1875
alligatoroidea Gray, Reference Gray1844
diplocynodontidae Hua, Reference Hua2004
Genus Diplocynodon Pomel, Reference Pomel1847
Type species. Diplocynodon ratelii Pomel Reference Pomel1847.
Diplocynodon elavericus sp. nov. Figures 2–9

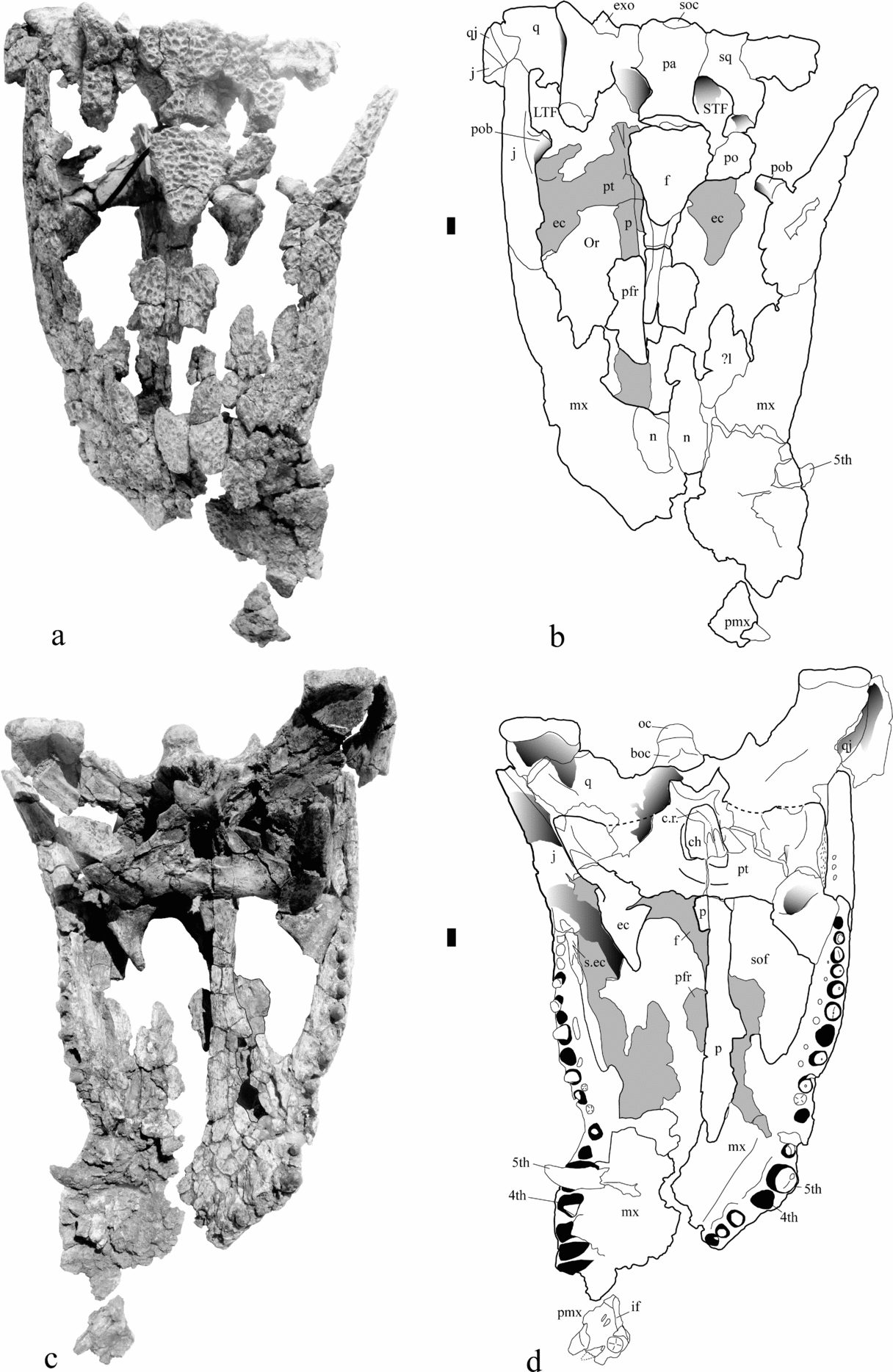
Figure 2. The skull of Diplocynodon elavericus (Rhinopolis B3) in dorsal (a, b) and ventral (c, d) views. Abbreviations: boc – basioccipital; ch – choana; c.r. – choanal recess; ec – ectopterygoid; exo – exoccipital; f – frontal; if – incisive foramen; LTF – lower temporal fenestra; j – jugal; l – lacrimal; mx – maxilla; n – nasal; oc – occipital condyle; Or – orbit; p – palatine; pa – parietal; pfr – prefrontal; po – postorbital; pob – postorbital bar; pt – pterygoid; pmx – premaxilla; q – quadrate; qj – quadratojugal; s.ec. – suture for ectopterygoid; sof – suborbital fenestra; sq – squamosal; STF – supratemporal fenestra; soc – supraoccipital; 4th, 5th – fourth and fifth maxillary tooth. Scale bar represents 1 cm.

Figure 3. The skull of Diplocynodon elavericus (Rhinopolis B3) in right lateral (a, b) and left lateral (c) views. boc – basioccipital; bsph – basisphenoid; ec – ectopterygoid; f – frontal; if – incisive foramen; j – jugal; mx – maxilla; n – nasal; pa – parietal; pfr – prefrontal; pmx – premaxilla; pob – postorbital bar; pt – pterygoid; q – quadrate; qj – quadratojugal; sq – squamosal; STF – supratemporal fenestra. Scale bar represents 1 cm.

Figure 4. The skull of Diplocynodon elavericus (Rhinopolis B3) in occipital (a, b) view. Abbreviations: boc – basioccipital; ec – ectopterygoid; exo – exoccipital; f.a. – foramen aëreum; f.c.p. – foramen caroticum posterius; fm – foramen magnum; f.pt – posttemporal fenestra; f.v. – foramen vagi; j – jugal; meu – median eustachian foramen; pa – parietal; pt – pterygoid; q – quadrate; qj – quadratojugal; soc – supraoccipital; sq – squamosal. Scale bar represents 1 cm.

Figure 5. The posterior skull table of Diplocynodon elavericus (Rhinopolis B3): (a) detail of the right supratemporal fenestra in anterior view. The white arrows delimit the parietal–quadrate suture; (b) dorsal view of the skull table with the black arrow pointing to the detailed region in (a). Abbreviations: otf – orbitotemporal foramen; pa – parietal; q – quadrate. Scale bar represents 1 cm.

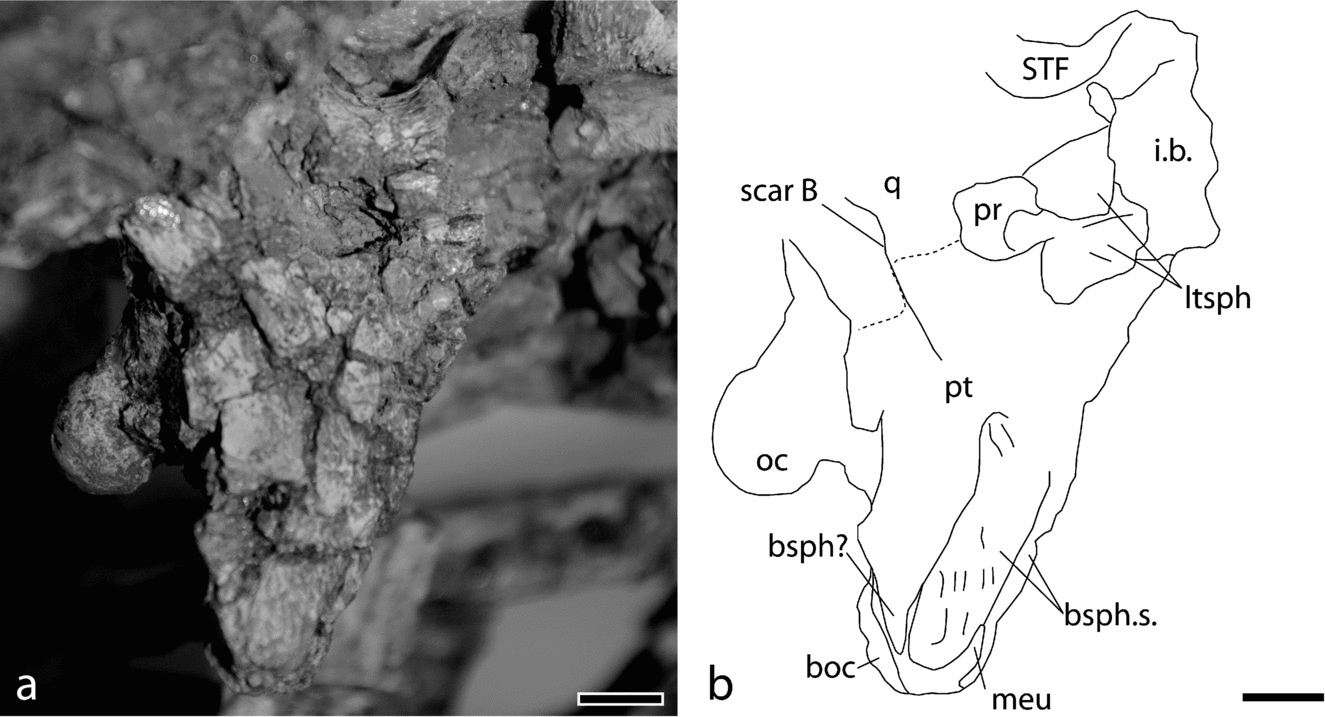
Figure 6. The braincase of Diplocynodon elavericus (Rhinopolis B3) in right lateral view. Abbreviations: boc – basioccipital; bsph – basisphenoid; bsph.s. – suture for basisphenoid; i.b. – internal braincase; ltsph – laterosphenoid; meu – median eustachian foramen; oc – occipital condyle; pr – prootic; pt – pterygoid; q – quadrate; scar B – muscle scar B of Iordansky (Reference Iordansky, Gans and Parson1973); STF – supratemporal fenestra. Scale bar represents 1 cm.

Figure 7. The left dentary of Diplocynodon elavericus (Rhinopolis B3) in occlusal (a), lateral (b) and medial (c) views. Abbreviations: m.g. – meckelian groove; sp. s. – splenial symphysis; sym – symphysis; 3rd – position for third confluent alveolus. Scale bar represents 1 cm.

Figure 8. The right dentary of Diplocynodon elavericus (Rhinopolis B3) in lateral (a), medial (b) and occlusal (c) views. Abbreviations: art – articular; den – dentary; f.a. – foramen aëreum; san – surangular; sp.s. – suture for splenial. Scale bar represents 1 cm.

Figure 9. Dorsal osteoderm of Diplocynodon elavericus (Rhinopolis B3) in dorsal (a) and ventral (b) views. Scale bar represents 1 cm.
Holotype. Rhinopolis B3, an almost complete skull with mandibular fragments and some osteoderms.
Etymology. ‘elavericus’ refers to the Latin name of the Allier River: elaver. Note that Bravard (Reference Bravard1843) used ‘Crocodilus elaveris’ to designate various crocodylian remains from Allier but did not provide any description, nor did he figure any specimens. As Vaillant (Reference Vaillant1872) pointed out, this early mention cannot be considered as having any right of priority over Diplocynodon ratelii, given the absence of a description or figure. ‘Crocodilus elaveris’ should be considered as a nomen nudum. Here, to prevent any ambiguity, a slightly different species name is chosen for the new taxon described below.
Locality and horizon. Middle Priabonian, Late Eocene of Domérat, Allier, France.
Diagnosis. Among the genus Diplocynodon, elongated quadrate rami and a pterygoid–palatine suture located at the posterior angle of the suborbital fenestra are autapomorphic for D. elavericus. This taxon differs from D. hantoniensis in lacking an elevated rim around the supratemporal fenestra and by having a horizontal skull table and a slender palatine projection in the maxillae. It also differs from D. tormis, D. muelleri and D. darwini in having a longer rostrum. The rostrum of D. elavericus is, however, shorter than the rostrum of D. gervaisi, D. ratelii and D. styriacus.
4. Description
Preservation. The remains of a single specimen were recovered in loose connection. Bones were not plastically deformed, but the skull is heavily fragmented and all the bones are displaced along cracks and sutures where they are cemented by clay. Therefore, the clay keeps the skull elements in a conformation close to the original one, but most of the contact between the bones has been destroyed either by clay swelling or during the excavation. Bones were prepared piece by piece and glued back together when connections were found. However, most of the medial and anterior region of the rostrum is missing. In some places, the bone and teeth are altered and turn into a yellowish powder.
Description. The premaxilla is very fragmentary. The only recognizable piece is an anterior fragment of the left premaxilla bearing two teeth. The anteriormost margin is straight. In ventral view, the first alveolus is broken and may have been small. A large occlusal pit separates the first alveolus from the second alveolus. The third alveolus is almost three times larger than the second. The premaxilla is then broken behind that alveolus. A foramen perforates the base of the alveolar wall. The margin of the external naris is smooth and vertical. The anterior extension of the incisive foramen is visible and was excluded from the tooth row margin.
Both maxillae are preserved but their palatal suture and the connection with the nasal is not preserved. The suture with the premaxilla is preserved on the right maxilla and does not show a marked notch for the passage of the mandibular caniniform dentition. Each maxilla bears 17 alveoli. The largest alveoli are as in other Diplocynodon, the fourth and fifth. At their level, the lateral margin of the maxilla is convex, and then posteriorly behind the sixth alveolus, a constriction marks the lateral margin of the bone. The occlusion pattern is well discernable from the presence of pits along the tooth row. The first pit is visible between the sixth and seventh alveoli. This is the deepest of all. Pits are then observed between the seventh and eighth up to the last alveoli. However, their position in the anterior portion progressively shifts from a nearly interfingering position to a complete medial position in the posterior region. Moreover, from the eleventh to the fifteenth alveolus, the pits turn into a groove medial to the tooth row. Small foramina perforate this groove. The maxilla forms most of the lateral and all the anterior margin of the suborbital fenestra. In this lateral margin, the maxilla is completely vertical. A foramen pierces this lateral wall. Posteriorly, the maxilla does not send any outgrowth between the tooth row and the ectopterygoid. Instead, the ectopterygoid is adjacent to the last two alveoli. The right maxilla offers some information about the medial wall of the cavichonchal recess. However, it is extensively fractured and further preparation could not be carried out. The cavichoncal recess is deep and its medial wall is smooth. Despite the fractures, no caecal recesses could be detected near the ventral lamina of the maxilla.
The nasal is very fragmentary and the few available fragments do not present any connection with the bones of the rostrum. However, given the conformation of the maxilla, the nasals seem to have been excluded from the external naris. The nasals seem to expand laterally near their mid-region. They are dorsally sculptured with small circular pits.
The lacrimals are very fragmentary and their anterior extension is impossible to assess.
The prefrontals are divided by the frontal. They are elongate and form the anteromedial margin of the orbits. Laterally, they are slightly inflated near the sutural region with the lacrimal.
The frontal sends a long anterior and finely sculptured process onto the rostrum. From what is presently preserved, this process attains the anterior level of the prefrontal extension. This process descends gently from the skull table but is not separated from it by a preorbital ridge. The skull table portion of the frontal is completely flat and sculptured with large circular and shallow pits. The frontal seems to enter into the supratemporal fenestrae. No contact between parietal and postorbital seems to occur.
The parietal forms the median and posterior region of the skull table. It is completely flat and shows the same patterns of sculptures on its dorsal surface. Between the supratemporal fenestrae, the bone margins are not straight but constricted. There is no ridge elevated along their rim. On the left side, the anteriormost margin of the supratemporal fenestra presents a fossa formed by the parietal. The parietal forms the ventral portion of the orbitotemporal foramen, therefore preventing the ascending process of the quadrate from attaining this foramen. The parietal contacts the squamosal lateral to that foramen. The posterior margin of the skull table is uneven and marked by two small processes.
The jugal forms the lateral boundary of the infratemporal fenestra. It is heavily sculptured on its dorsal surface with deep pits. It also participates in the anterolateral boundary of the orbit where it projects over the maxilla as a thin plate. The jugal contributes to the postorbital bar by sending a process, which is inset from the dorsal margin of the bone. While it is inset, the process is, however, continuous with the medial smooth margin of the jugal. Ventrally, the suture for reception of the ectopterygoid does not extend onto the posterior jugal rod. Laterally, the jugal does not reach the quadrate but leaves the quadratojugal visible.
The quadratojugal is very fragmentary and can only be observed from the right side. The preserved part is reduced to a lamina between the quadrate and jugal, where it is sculptured and appears on the lateral side. It does not send any process along the lower temporal bar.
Both quadrate bones are preserved, but the right one is in a better state of preservation. Due to fractures and deformation, the quadrate rami are almost vertically oriented. Despite such unnatural verticalization, the quadrate rami are unusually elongate and are as long as the descending squamosal prongs. As a comparison, the quadrate rami are shorter than the squamosal prongs in Crocodylus or Caiman. The medial quadrate hemicondyle is smaller than the massive lateral hemicondyle. The foramen aëreum appears on the dorsomedial margin of the quadrate.
The left postorbital is preserved. Its lateral margin is convex and gives to the anterior skull table a gently continuous shape. The postorbital bar is mediolaterally compressed and is therefore expanded anteroposteriorly.
The squamosal is best preserved on the right side. Here, a prominent prong is visible. This prong projects far posteriorly behind the level of the occipital condyle and beyond the lateral margin of the skull table. The squamosal extends far ventrally on the paroccipital process. Its suture with the quadrate lies on the ventral margin of the otic notch. The dorsal margin of the otic notch is not straight but bowed. The squamosal extensively overhangs the occipital region.
The supraoccipital participation to the posterior portion of the skull table is limited to a small incursion in dorsal aspect. The supraoccipital is, however, massive and extensively overhangs the occiput. A strong median crest projects ventrally and is framed by two massive spines that project posteriorly beyond the posterior margin of the skull table. A pair of grooves delimits the spines from the median crest.
The exoccipital is severely fractured but is nevertheless almost complete on the right side. It covers most of the occipital portion of the skull, forming all the margin of the foramen magnum. It laterally expands and creates a developed process overhanging the cranioquadrate passage. However, the passage, while delimited by a depression, is not well preserved. A large foramen vagus pierces the bone at the level of the foramen magnum. The ventral process of the exoccipital projects beyond the foramen caroticum posterius but does not attain the ventralmost level of the basioccipital.
The basioccipital is divided into two regions. First, the occipital condyle presents a foramen in its dorsal part. Second, the vertical basioccipital plate presents two shallow pillars, then a strong descending single pillar in its ventralmost portion. The basioccipital plate is very long in proportion compared to that of extant Crocodylus or Alligatoridae.
The basisphenoid is incomplete but its suture can be observed at the base of the basioccipital plate. It forms the posterior margin of the large median Eustachian opening.
The foramen ovale is large and its organization shows that the prootic is well visible inside this foramen for the entire posterior margin. It is not obscured by any other bones but does not participate in the wall. The suture between quadrate and pterygoid is not obvious but may have been straight from the foramen ovale to the posterior margin of the braincase. The laterosphenoid is heavily fragmented but it is recognizable as forming the anterodorsal portion of this foramen.
The pterygoid wings appear vertical, but it is impossible to assess the contribution of deformation. The Torus transiliens is thick and finely pitted. It corresponds to the attachment of tendons for the M. pterygoideus posterior. The posterior margin of the wing is uneven and slightly concave on its lateral side. Medially, the pterygoid is notched at the level of the median Eustachian passage. The lateral walls of this notch send two processes posteriorly. The choanae are large and square. Their posterior margin is almost continuous with the pterygoid surface. They are slightly more elongate in the anteroposterior direction. They pierce the pterygoids near their posterior margin and face anteroventrally. A lamina divides the choanae and enters the dorsal wall of the cavity. The pterygoid forms the posterior margin of the suborbital fenestra and is wide there. It is slightly notched at this level.
The ectopterygoid sends a posterior process onto the pterygoid wing but does not reach its posteriormost margin. The suture with the pterygoid is almost straight. Dorsally, the posterior process of the ectopterygoid on the jugal is almost absent. The anterior process, however, projects far along the tooth row and is contiguous with the last five alveoli and contributes directly to the last alveolus.
The palatine is a long bone that participates in the medial margin of the suborbital fenestra. The right side is preserved in its entirety for the ventral part. It is anteroposteriorly straight and its lateral wall is vertical in its posterior portion. The suture with the pterygoid occurs at the posteriormost level of the suborbital fenestra. Near the anterior level of the fenestra, the suture with the maxilla shows a mediolateral direction. A small foramen pierces the bone near the median suture. More anteriorly, the palatine penetrates into the maxilla by sending a long and slender pointed process reaching the level of the seventh maxillary alveolus.
The mandible is very fragmentary. Pieces of both dentary bones are present and indicate the presence of at least twelve alveoli in the left bone. The right dentary is broken at the level of the possibly confluent and largest alveoli, as evidenced by the size of their remaining wall. At least eleven alveoli can be counted in this bone, therefore providing an estimate of at least fifteen alveoli in this ramus. Numerous foramina pierce the base of the alveoli. The splenial is not preserved but its suture on the dentary indicates that it was excluded from the symphysis. The profile of the tooth row is slightly constricted behind the fifth alveolus with the presence of small alveoli. Alveoli then become larger and the profile becomes elevated from the ninth or tenth alveolus. The surangular consists of a fragment from the right posterodorsal portion. The smooth attachment for M. adductor mandibulae superficialis and medius is large and inclined laterally. A small fossa is present lateral to the dorsal contribution of the surangular to the articular jaw joint. The corresponding articular consists of the articular capsule as well as the descending anterior process. The surangular–articular suture is simple, but due to preservation, it is not possible to properly assess the position of the visible foramen relative to the suture. There is no sulcus between the articular and surangular. The foramen aëreum is visible and is laterally shifted.
The dentition of this taxon consists of conical teeth in the premaxilla and in most of the maxilla. From the eleventh alveolus, the teeth are slightly mediolaterally compressed and blunter. Two ridges run from the mesial face to the apex. The enamel is smooth.
Postcranial elements are represented by a few osteoderms from the dorsal shield. The best preserved of all is rectangular and presents large and shallow rounded pits on its dorsal surface. A shallow median ridge is present and runs along most of its length, except anteriorly where the smooth articulation facet is present. This ridge slightly overhangs the posterior margin. Laterally and medially, the osteoderm is sutured. Its ventral surface is smooth with criss-crossing striation for epaxial muscle insertion.
5. Comparison with other species of the genus
The quadrate rami are longer than in any other species of Diplocynodon or alligatoroids. Discarding deformation, the quadrate rami would have projected well beyond the posterior margin of the skull table, whereas among other species of Diplocynodon, the quadrate rami are located close to the level of the posterior skull table margin, when observation is made from a dorsal point of view. In contrast to D. darwini, D. ratelii, D. muelleri, D. tormis, D. hantoniensis or D. styriacus, the palatine–pterygoid suture is located at the posterior level of the suborbital fenestra in D. elavericus.
Diplocynodon elavericus is younger than Diplocynodon hantoniensis from the Headon Hill formation (Early Priabonian: Hooker, Cook & Benton, Reference Hooker, Cook, Benton, Benton, Cook and Hooker2005) of the United Kingdom. These two species are very similar in general aspect. They both show a broad and stout rostrum, well-developed squamosal prongs that project far posteriorly, well-developed supraoccipital spines projecting posteriorly, the absence of a preorbital ridge and comparable head size and proportions (see Table 2 for comparison). However, a number of differences are to be observed, including the completely flat dorsal surface of the parietal in D. elavericus. D. elavericus and D. hantoniensis are among the largest known species of the genus. Such a concave parietal can be observed in D. hantoniensis, as well as in D. cf. hantoniensis from the Oligocene of Dordogne (Vignaud et al. Reference Vignaud, Brunet, Guevel and Jehenne1996). Other differences include the presence of an elevated rim on the medial margin of the supratemporal fenestrae in D. hantoniensis. Also, the shape of the palatine projection within the maxilla is different. In D. hantoniensis, the anterior palatine projection is wide and squared off and reaches the level of the ninth maxillary alveolus. In D. elavericus, the palatine projection is slender and pointed and reaches further anteriorly between the level of the sixth and seventh alveoli.
Diplocynodon tormis from the Late Eocene of Salamanca, Spain, is older than Diplocynodon elavericus and Diplocynodon hantoniensis as it is Bartonian in age (Buscalioni, Sanz & Lourdes, Reference Buscalioni, Sanz and Lourdes1992). It is, however, very different in size and skull proportions, and the differences exhibited render it less comparable to D. elavericus than D. hantoniensis. The basalmost member of Diplocynodontidae, D. darwini, has a relatively short rostrum together with D. muelleri, while D. ratelii, D. gervaisi and specimens referred to D. styriacus show the most slender morphology. The current interrelationship hypothesis proposed here reveals that the general skull proportions do not have any phylogenetic meaning, D. elavericus and D. hantoniensis not being closely related to each other. Their contemporaneous occurrence shows that several lineages inhabited Europe with probably similar ecologies or feeding habits. This could explain such morphological convergence despite their different phylogenetic positions.
Vignaud et al. (Reference Vignaud, Brunet, Guevel and Jehenne1996) recognized that the posterior region of the maxillary tooth row is mesially bound by the ectopterygoid in D. ratelii, D. gervaisi and D. tormis but not in D. hantoniensis. I disagree, however, about the condition in D. hantoniensis, and the preceding description shows that the same is true for D. elavericus. Such a character might be added to the generic diagnosis of Diplocynodon pending observations on other species. Piras & Buscalioni (Reference Piras and Buscalioni2006) described the maxilla of D. muelleri as sending an outgrowth between the tooth row and the ectopterygoid. In D. tormis (Buscalioni, Sanz & Lourdes, Reference Buscalioni, Sanz and Lourdes1992), the ectopterygoid is described as running beside the alveolar border; however, the interpretive drawing seems to indicate otherwise. In specimens attributed to D. styriacus (Ginsburg & Bulot, Reference Ginsburg and Bulot1997), there is no outgrowth of the maxilla separating tooth row from ectopterygoid in the photos of the authors (fig. 2, p. 115; fig. 3, p. 118), although their drawings indicate otherwise.
6. Phylogenetic analysis
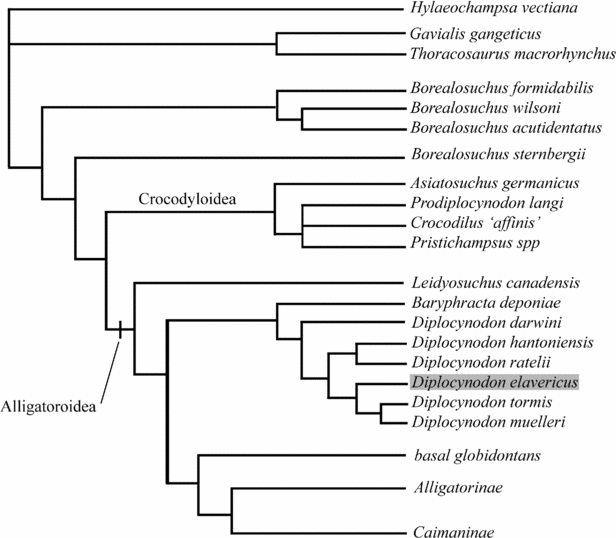
In order to compute a maximum of alligatoroid taxa, Diplocynodon elavericus was coded (Table 1) and included in the matrix of Brochu (Reference Brochu1999, Reference Brochu2004), together with the recently coded Diplocynodon muelleri and Diplocynodon tormis by Piras & Buscalioni (Reference Piras and Buscalioni2006). Nine other taxa were taken from the matrix of Salisbury et al. (Reference Salisbury, Molnar, Frey and Willis2006), and their codings were adjusted to the present matrix. Because it has been demonstrated that Borealosuchus was different from the alligatoroid Leidyosuchus (Brochu, Reference Brochu1997), four taxa of the genus Borealosuchus were included in the analysis (B. sternbergii, B. acutidentatus, B. wilsoni and B. formidabilis) to test its monophyly. Two gavialoids (Gavialis gangeticus and Thoracosaurus macrorhynchus), as well as two other crocodyloids (Prodiplocynodon langi and ‘Crocodylus’ affinis), were added to the analysis and permitted wide sampling of the Eusuchia. The basal eusuchian Hylaeochampsa vectiana was defined as the outgoup taxon. A total of 49 ingroup taxa were treated under multiple heuristic searches with Paup* (Swofford, Reference Swofford2002) for 100 random-seed replicates. The analysis yielded 2432 equally most-parsimonious trees with a length of 397 steps (CI, excluding uninformative characters = 0.4557; RI = 0.7721).
Table 1. Character codings for Diplocynodon elavericus

Key to characters in Brochu (Reference Brochu1999, Reference Brochu2004).
Table 2. Comparative measurements (in mm) of the skull of Diplocynodon elavericus (Rhinopolis B3) and Diplocynodon hantoniensis

The topology of the strict consensus tree (Fig. 10) is mostly consistent with the results of Salisbury et al. (Reference Salisbury, Molnar, Frey and Willis2006), with Alligatoroidea and Crocodyloidea being sister clades, although Pristichampsus spp. is presently incorporated in a polytomy with Prodiplocynodon langi and ‘Crocodylus’ affinis, whereas it is basal to the Brevirostres in the results of Salisbury et al. (Reference Salisbury, Molnar, Frey and Willis2006). Also here, Borealosuchus is not monophyletic: B. sternbergii sits in a more derived position than the clade composed of the three other species of Borealosuchus. The limited resolution at the base of Eusuchia can be interpreted as a result of the small taxon sampling in the analysis and may also explain why Gavialoidea is not sister-group to Borealosuchus but forms a polytomy with Hylaeochampsa vectiana. However, the purpose of this sampling was to polarize characters in order to stabilize the relationship within Alligatoroidea. The topology of this clade is consistent with the results of Brochu (Reference Brochu1999, Reference Brochu2004) and is also consistent with the results of Piras & Buscalioni (Reference Piras and Buscalioni2006) in the basal arrangement of the Diplocynodontidae with Baryphracta deponiae as the basalmost member, and then D. darwini as the basalmost diplocynodontine. Moreover, the internal nodes are here better resolved; D. hantoniensis and D. ratelii are sister taxa and form the sister group to the three remaining taxa: D. elavericus is the sister taxon to D. muelleri and D. tormis, which form a clade. A Bremer decay analysis tested the robustness of the arrangement; one step longer than the optimal tree length did not alter the major divisions. Internal relationships are polytomic, but Leidyosuchus canadensis, Diplocynodontidae, basal globidontans, Alligatorinae and Caimaninae are well defined.

Figure 10. Hypotheses of relationships for Diplocynodon elavericus included in the matrix of Brochu (Reference Brochu1999, Reference Brochu2004) with a total number of seven diplocynodontids. Relationships with more derived members of Alligatoroidea were simplified for clarity of reading but the results do not differ from that of Brochu (Reference Brochu1999, Reference Brochu2004).
Diplocynodontidae are supported by the presence of paired osteoderms that suture together (39–2) and a linear frontoparietal suture (86–1), although these traits are unknown in D. elavericus. Also unknown in D. elavericus, the nasal excluded from the external naris (95–2) as well as the foramen opening on the angular–surangular suture, are still considered ambiguous because they are also present in Borealosuchus (Brochu, Reference Brochu1999). The linear quadrate–pterygoid suture from the basisphenoid to the exposure of the foramen ovale (127–1) remains ambiguous as it is originally only observed in D. ratelii (Brochu, Reference Brochu1999) but is now found in D. elavericus. The phylogenetic results unambiguously present the splenial as excluded from the symphysis as a synapomorphy for Diplocynodontinae (43–1), and this is again confirmed here with D. elavericus. The presence of confluent dentary teeth of similar size (3+4) in D. elavericus remains ambiguous as it is also shared with Leidyosuchus canadensis, Borealosuchus and also Deinosuchus (Brochu, Reference Brochu1999). Piras & Buscalioni (Reference Piras and Buscalioni2006) defined the genus Diplocynodon as possessing a large medial jugal foramen (120–1), but this trait is coded as small for D. darwini (Brochu, Reference Brochu1999) and is unknown in D. elavericus. Piras & Buscalioni (Reference Piras and Buscalioni2006) also include in this definition the trait ‘largest dentary alveolus immediately caudal to fourth is 10, 11 or 12 (167–2)’, which is consistent with observations made on D. elavericus.
The fragmentary nature of D. elavericus does not help to define further characters for the genus Diplocynodon. Nevertheless, its inclusion triggers a resolution within Diplocynodontidae. This arrangement does not show a continuous succession of species through time. Instead, old records show more affinities with recent ones, as exemplified by sister taxa such as D. tormis + D. muelleri, or D. hantoniensis + D. ratelii. This topology remains tentative. Indeed, the Bremer decay analysis demonstrated a weak support and it might be preferable to include more complete taxa that would bring more robustness into the internal relationship of the Diplocynodontidae. Interestingly, the sister group relationship of D. hantoniensis implies a long fossil record for D. ratelii from the Late Eocene up to the Early Miocene. For this reason, it might be worth testing the relationship of the specimen from the Early Oligocene of Céreste, attributed to D. ratelii (Brinkmann & Rauhe, Reference Brinkmann and Rauhe1998). Finally, the most reliable aspect of the inclusion of D. elavericus in a phylogenetic framework permits confirmation of the monophyly of the Diplocynodontidae as well as the endemic nature of this clade to Europe.
7. Diplocynodon in the context of the Late Eocene climate change
The Palaeogene and Neogene fossil record of crocodylians in Europe is not continuous, but a broad pattern of faunal succession has been identified. Buffetaut (Reference Buffetaut1982) gave a first brief review in which he emphasized a decline in diversity during the Eocene period. A similar pattern can be observed when looking at the Eocene–Oligocene fossil record of the Duero Basin, Spain (Martín de Jesús et al. Reference Martín de Jesús, Jimenez Fuentes, Fincias, Del Prado and Mulas Alonso1987), or that of Italian crocodylians as listed by Kotsakis, Delfino & Piras (Reference Kotsakis, Delfino and Piras2004). Despite these observations, European crocodylians have received little attention in this respect. In his review of crocodylian diversity from the Cretaceous to the present, Markwick (Reference Markwick1998a) identified such a diversity decline during the Late Eocene over a global scale and invoked the influence of climate variation on crocodylian diversity. However, he pointed out that a sampling effect could influence such results. Due to the incompleteness of the fossil record, it is challenging to link patterns of faunal turnover with the evolution of climatic conditions. Although this problem is inherent to the fossil record of vertebrates, sparse but well-dated occurrences can provide important pieces of evidence, especially because semi-aquatic crocodylians are ectothermic and thus are dependent on environmental conditions (Berg, Reference Berg1965; Hutchison, Reference Hutchison1982; Antunes, Reference Antunes1994; Markwick, Reference Markwick1998a). Therefore, their occurrence implies sub-tropical or tropical conditions. Diplocynodon elavericus is Middle Priabonian in age and represents one of the youngest known records of crocodylians in the Eocene period of Europe (Fig. 11). Of similar age, isolated teeth and osteoderms have been attributed to Diplocynodon from the Priabonian of the Basque country, Spain (Astibia et al. Reference Astibia, Aranburu, Suberbiola, Murelaga, Sesé, Cuesta, Moya-Sola, Baceta, Badiola and Köhler2000). Nevertheless, Diplocynodon elavericus is recovered at a higher latitude than the Spanish remains and introduces the discussion about the evolution of the genus in the temperature decline of the Late Eocene.
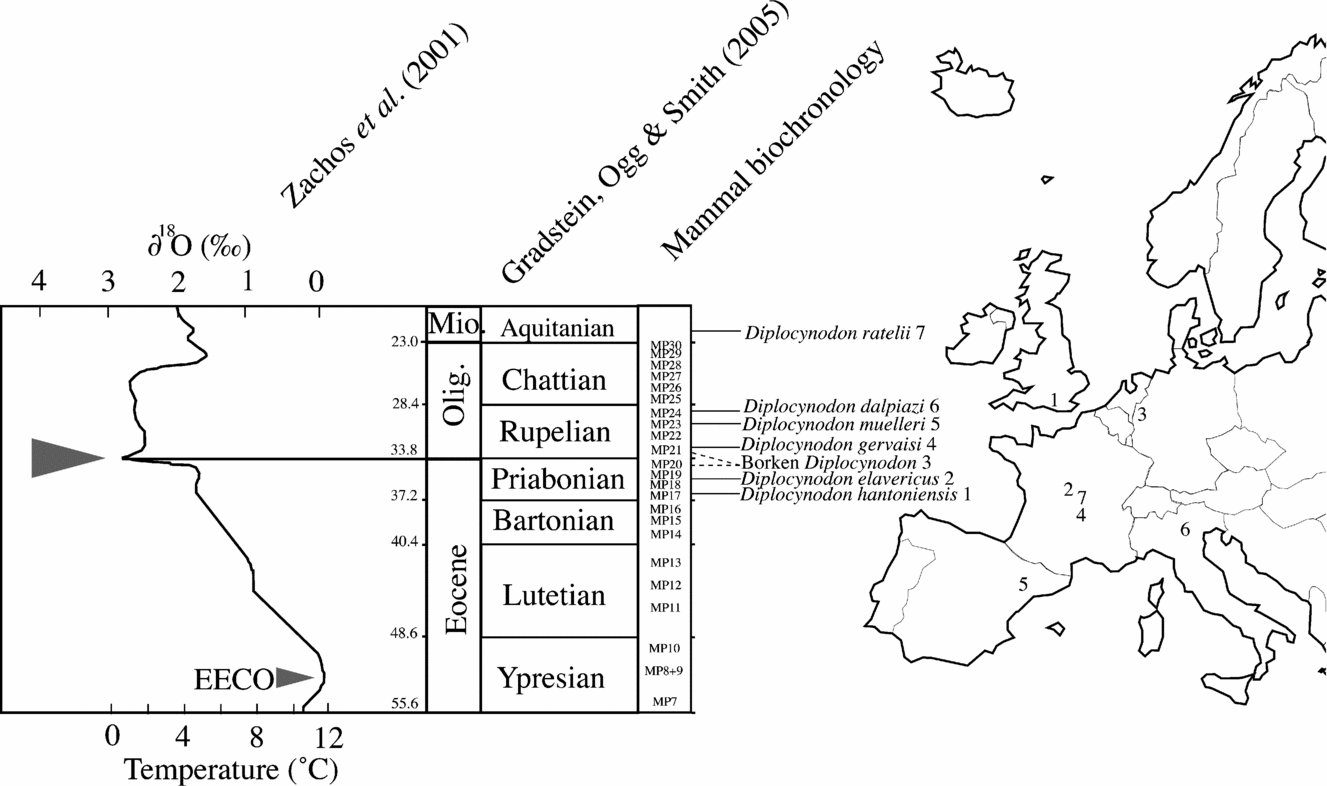
Figure 11. Geographic occurrences of six Diplocynodon species compared to their stratigraphic position (numerical ages according to Gradstein, Ogg & Smith, Reference Gradstein, Ogg and Smith2005, and European Land Mammal Ages according to BiochroM, Reference Aguilar, Legendre and Michaux1997) and the global temperature curve across the Late Eocene and Oligocene (after Zachos et al. Reference Zachos, Shackleton, Revenaugh, Palike and Flower2001). Numbers refer to the localities around the Eocene–Oligocene boundary having yielded Diplocynodon: 1 – Hordwell; 2 – Domérat; 3 – Borken; 4 – Ronzon; 5 – Lleida; 6 – Monteviale; 7 – St-Gérand-le-Puy. EECO – Early Eocene Climatic Optimum. Note the unsettled stratigraphic position of the remains from Borken as highlighted by the dashed line.
Evolution of global temperatures through the Cenozoic has received particular attention during the last decade thanks to quantitative analyses. Isotopic composition of benthic foraminifera (Zachos et al. Reference Zachos, Shackleton, Revenaugh, Palike and Flower2001) and biomarkers (Schouten et al. Reference Schouten, Eldrett, Greenwood, Harding and Sinninghe Damsté2008) revealed a negative shift in global temperature at the Eocene–Oligocene transition, a climatic phenomenon that was previously hypothesized to account for turnover in mammalian communities of Europe and known as ‘La Grande Coupure’ (Stehlin, Reference Stehlin1910; Legendre, Reference Legendre1989 for a review; Hooker, Collison & Sille, Reference Hooker, Collinson and Sille2004). This climatic transition marks the end of a greenhouse world, which peaked in temperature after the Palaeocene–Eocene boundary. During most of the Eocene period, continental mean annual air temperature seems to have been relatively constant (Mosbrugger, Utescher & Dilcher, Reference Mosbrugger, Utescher and Dilcher2005; Schouten et al. Reference Schouten, Eldrett, Greenwood, Harding and Sinninghe Damsté2008), with progressively increasing seasonality from the end of the Middle Eocene, as revealed by floras from Central Europe (Mosbrugger, Utescher & Dilcher, Reference Mosbrugger, Utescher and Dilcher2005). However, the present resolution of the climatic record is not sufficient to allow a precise climatic reconstruction, and although it has become widely admitted that a permanent ice sheet settled on Antarctica from the earliest Oligocene (Miller, Wright & Fairbanks, Reference Miller, Wright and Fairbanks1991; Lear, Elderfield & Wilson, Reference Lear, Elderfield and Wilson2000; Zachos et al. Reference Zachos, Shackleton, Revenaugh, Palike and Flower2001; Coxall et al. Reference Coxall, Wilson, Palike, Lear and Backman2005; Ivany et al. Reference Ivany, Van Simaeys, Domack and Samson2006), there is some evidence, including reports of ice-rafted debris, to suggest that a continental ice sheet and glaciers were already present in the northern hemisphere as early as the Middle Eocene (Moran et al. Reference Moran, Backman, Brinkhuis, Clemens, Cronin, Dickens, Eynaud, Gattacceca, Jakobsson, Jordan, Kaminski, King, Koc, Krylov, Martinez, Matthiessen, McInroy, Moore, Onodera, O'Regan, Pälike, Rea, Rio, Sakamoto, Smith, Stein, St John, Suto, Suzuki, Takahashi, Watanabe, Yamamoto, Farrell, Frank, Kubik, Jokat and Kristoffersen2006; Eldrett et al. Reference Eldrett, Harding, Wilson, Butler and Roberts2007; Tripatti et al. Reference Tripatti, Eagle, Morton, Dowdeswell, Atkinson, Bahé, Dawber, Khadun, Shaw, Shorttle and Thanabalasundaram2008). Such climatic evolution must have had an influence, for example, on the distribution of ectothermic vertebrates.
Late Eocene and Oligocene continental crocodylian faunas consist of only a single taxon at any given locality. This contrasts significantly with Early or Middle Eocene ecosystems, in which niche partitioning allowed several taxa to co-occur. For example, the Lutetian localities of Messel or Monte Bolca include terrestrial taxa such as the ziphodont Pristichampsus, the blunt-snouted ‘Allognathosuchus’, as well as semi-aquatic forms such as Asiatosuchus-like taxa (sensu Delfino & Smith, Reference Delfino and Smith2009), or Diplocynodon (Berg, Reference Berg1966; Kotsakis, Delfino & Piras, Reference Kotsakis, Delfino and Piras2004). A bias in sampling cannot be excluded for younger faunas as they are not preserved as Lagerstätten, as is the case at Messel or Monte Bolca. However, from the Priabonian onward, ziphodont crocodylians are no longer found, whereas their remains are not so rare in Lutetian or Bartonian fluvial deposits, their youngest European occurrence being from the Bartonian (MP16, BiochroM, Reference Aguilar, Legendre and Michaux1997) of La Badayré/Sables du Castrais (Berg, Reference Berg1966) and a single tooth from the Quercy (imprecise date–Bartonian after Buffetaut, Reference Buffetaut1982). The last record of Asiatosuchus-like taxa is from the Bartonian of Camburg, Germany (Vasse, Reference Vasse1992). The latest basal alligatorine from Europe is Arambourgia gaudryi from Quercy and may represent an exception as it is as old as or slightly older than the new taxon from Domérat (MP18 or MP19) (Rage, Reference Rage2006). While semi-aquatic fossil crocodylians were certainly ectothermic, as this represents a great advantage in such environment (Seymour et al. Reference Seymour, Bennet-Stamper, Johnston, Carrier and Gordon2004 for a review), terrestrial crocodylians may have had a different metabolic regime and thus, their climatic requirements may have differed from that of semi-aquatic crocodylians. Extant crocodylians live in a specific temperature window and any variation outside of it has consequences for their life (Lang, Reference Lang1979; Markwick, Reference Markwick1998b): either they are pushed to migrate or they eventually die. It is, however, impossible to quantify the range of temperatures that the extinct crocodylians could have sustained, and thus comparison with the known increasing seasonality in continental ecosystems of Europe is limited. It is clear, however, that the Eocene–Oligocene negative temperature shift post-dates the crocodylian diversity decline and cannot be considered as a possible cause.
As was proposed by Buffetaut (Reference Buffetaut1982), progressive temperature decline from the middle Late Eocene may have entered the range in which most crocodylian taxa could no longer survive. In this light, the latitudinal distribution of the genus Diplocynodon through time is interesting (Fig. 11). Diplocynodon hantoniensis is recorded from the base of the Priabonian stage in southern England. Diplocynodon elavericus is recorded at a more southern latitude in France later on (MP18–20). In any case, these occurrences are recorded prior to the negative temperature shift of the Eocene–Oligocene boundary, which is considered to take place at the limit between the mammal zones MP20 and MP21 (Escarguel, Marandat & Legendre, Reference Escarguel, Marandat, Legendre, Aguilar, Legendre and Michaux1997; but see Hooker, Collinson & Sille, Reference Hooker, Collinson and Sille2004). Subsequent to the dramatic drops in temperatures, the next record of Diplocynodon is that from Ronzon (Berg, Reference Berg1966), which is dated as basal Rupelian (MP21) and contains Diplocynodon gervaisi basking in sub-tropical to tropical conditions. Another lower Oligocene (Rupelian, MP24) French locality near Céreste produced a specimen attributed to Diplocynodon ratelii (Brinkmann & Rauhe, Reference Brinkmann and Rauhe1998). It is more or less of the same age as Diplocynodon dalpiazi from Monteviale, Italy (Del Favero, Reference Del Favero1999). Indeed, it cannot be ruled out that Diplocynodon suffered from temperature decline during ‘La Grande Coupure’ and that low latitude regions of Europe may have constituted climatic refugia before recolonization of Europe became possible during the following Oligocene warming, with D. gervaisi as an example. A progressive southward migration of Diplocynodon during this climatic deterioration would represent a tempting explanation, nevertheless the continental record is not continuous and the absence of crocodylians at higher latitude localities of Europe needs to be tested, especially near the Eocene–Oligocene boundary. In this regard, a mandibular fragment was reported from the Melanienton formation at Borken, northern Germany (Gramann, Reference Gramann1958). This formation is currently attributed a Late Eocene age on the basis of mammals (MP20; BiochroM, Reference Aguilar, Legendre and Michaux1997), but the locality of Borken has never yielded any mammals and was usually considered Early Oligocene in age (Gramann, Reference Gramann1958). Recent calibration using nannoplankton places the Melanienton formation across the Eocene–Oligocene boundary (Martini & Reichenbacher, Reference Martini and Reichenbacher2007). As no information was provided concerning the position of the specimens from Borken within the formation, the Diplocynodon remains from that locality could be either Late Eocene or Early Oligocene. A southward shift in relation to climatic deterioration has already been proposed for Diplocynodon during the Miocene (Böhme, Reference Böhme2003) and a similar shift in distribution may well have taken place at the Eocene–Oligocene transition when temperatures dropped more rapidly than during the gradual Late Miocene cooling. However, two hypotheses remain to be tested. Diplocynodon may have been able to endure several months below 0°C, as is the case for the extant Alligator sinensis. Such an ability might have helped it to survive the Eocene–Oligocene climatic deterioration and would explain the success of Diplocynodon in continental ecosystems of Europe over other crocodylian taxa. While Markwick (Reference Markwick1998b) presented all extant crocodylians as having similar thermal preferences, Lang (Reference Lang1979) measured differences of body temperature between Alligator mississippiensis and Crocodylus acutus, and suggested these might reflect different thermoregulatory strategies. That extant Alligator is more cold-tolerant than extant Crocodylus still requires a quantitative assessment. Although cold-tolerance in Diplocynodon has to be tested isotopically (oxygen from enamel apatite), it may extend the optimization of this character from Alligatoridae (Alligatorinae + Caimaninae) down to basal Alligatoroidea. A second option, which does not exclude the former, is a possible southward migration of Diplocynodon during the Late Eocene period. Isolation of populations by climatic barriers would have promoted speciation within this genus. Coeval localities are not abundant, but the recognized high diversity of Diplocynodon species might be the result of such speciation mechanisms. A comprehensive review of the relationships of all the known Diplocynodon species is pending and may provide an interesting comparative basis for the reconstruction of possible migratory events.
8. Conclusions
Relatively complete remains of a new Diplocynodon taxon allow comparison with other members of the genus. Diplocynodon elavericus is more closely related to the Spanish Diplocynodon (D. muelleri and D. tormis) than to Diplocynodon hantoniensis or Diplocynodon ratelii from more northern localities of Europe. The new taxon adds to the diversity of this successful group in continental ecosystems of Europe. It is widely recognized that a global climatic event led to a decrease in temperatures near the Eocene–Oligocene boundary. Biotic evidence, including the evolution of crocodylian assemblages, has the potential to provide reliable proxies insofar as it may record such a climatic deterioration, namely by presenting a diversity decline. At that time, the presence of continental ice in the northern hemisphere is consistent with such a hypothesis. In any case, the extensive fossil record of Diplocynodon in Europe throughout the Palaeogene and early Neogene might be an indirect evidence for the ability of this genus to sustain low temperatures.
Acknowledgements
The author expresses his gratitude to E. Buffetaut for discussion and advice during the completion of this work. S. Jouve and one anonymous reviewer significantly helped to improve the content of the manuscript. Former labmates K. Travouillon and G. Suan, as well as doctoral students in geology, Lyon 1, provided entertaining support during the long preparation of the specimen. F. Escuillié brought the present specimen to my attention. S. Hervet provided much help in making the specimen accessible from the Association Rhinopolis. The author was funded by Le Conseil Général de l'Allier.