Introduction
Nitrogen (N) is a primary limiting factor of cropland productivity and application of N fertilizers is an important agricultural practice used to sustain and/or increase crop yields. However, N fertilization can also facilitate weed growth, reducing nutrient-use efficiency of crop plants as a result of enhanced interspecific competition (Blackshaw et al. Reference Blackshaw, Brandt, Janzen, Entz, Grant and Derksen2003). Many previous studies have examined the effects of N fertilization on weed–crop interactions (Blackshaw and Brandt Reference Blackshaw and Brandt2008; Chauhan and Johnson Reference Chauhan and Johnson2011; Knezevic and Datta Reference Knezevic and Datta2015), but as far as we are aware, no study has considered these effects from a stoichiometric perspective. Plant stoichiometry refers to the relative proportion of chemical elements in the biomass, which can be considered the indicator of plant nutrient requirements (Li et al. Reference Li, Liu, Fan, Ma, Ding, Zhong and Wang2015; Sterner and Elser Reference Sterner and Elser2002). The leaf is the primary photosynthetic organ of plants, and its stoichiometry often reflects nutrient requirements for plant growth (Güsewell Reference Güsewell2004; Wright et al. Reference Wright, Reich, Westoby, Ackerly, Baruch, Bongers, Cavender-Bares, Chapin, Cornelissen and Diemer2004). Leaf stoichiometry can be very sensitive to altered nutrient availability, such as increased soil N due to fertilization or deposition (Yuan and Chen Reference Yuan and Chen2015a, Reference Yuan and Chen2015b). Thus, knowledge of the effects of N fertilization on plant leaf stoichiometry is critical to understanding nutrient cycling and is of diagnostic value in predicting the competitive outcomes between crop and weed plants under different N input conditions in agroecosystems (Güsewell Reference Güsewell2004; Tilman Reference Tilman and Crawley1997).
The impact of N addition on leaf stoichiometry has been empirically examined in a variety of plant types. A majority of studies in forest and grassland ecosystems indicate that N inputs can increase N concentration [N], resulting in increased nitrogen:phosphorus (N:P) and nitrogen:potassium (N:K) ratios (Ågren and Weih Reference Ågren and Weih2012; Yuan and Chen Reference Yuan and Chen2015a). The effects of N fertilization on leaf stoichiometry were also observed in crops. For example, Ye et al. (Reference Ye, Liang, Chen, Li, Ji and Zhu2014) reported that rice (Oryza sativa L.) showed allometric increase of stem–leaf [N] and P concentration [P] in response to N fertilization, resulting in an increased N:P ratio. In contrast, N fertilization increased [N] and N:P ratio but reduced [P] in wheat (Triticum aestivum L.) leaves (Zhong et al. Reference Zhong, Yan, Xu and Shangguan2015). These inconsistent patterns of elemental concentrations suggest that different mechanisms may modulate the responses of leaf stoichiometry to N inputs in plants.
Based on the structure–function chain hypothesis, plant nutrient elements can be classified into three groups according to their biological functioning: growth-related elements, biomass-related elements, and limited-uptake elements (Ågren and Weih Reference Ågren and Weih2012). Following this hypothesis, two likely scenarios can be proposed to describe the responses of leaf [N], [P], and K concentration [K] and their ratios to addition of N. If plants exhibit stoichiometric flexibility in such a way that increased plant biomass is associated with changes in leaf stoichiometry, [N] and [P] would increase with increasing N fertilizer. Meanwhile, [P] has a weaker response than [N] due to the limited P available in soil (Yuan and Chen Reference Yuan and Chen2015a), whereas [K] would remain relatively constant, resulting in increased N:P and N:K ratios. Alternatively, if plants become more productive while maintaining stoichiometric homeostasis in leaves, there will be no significant variation in elemental concentrations and their ratios (Sterner and Elser Reference Sterner and Elser2002). However, which of these scenarios can predict the responses of plant (crops and weeds) leaf stoichiometry in agroecosystems remains unknown.
Different farming practices may also affect the responses of crop and weeds to N fertilization. In China, straw return has become a common agricultural practice. Some studies showed that straw return in fields can return a large amount of plant K to soil, while decreasing mineral N losses by causing N immobilization and increasing soil carbon (C) sequestration (Liu et al. Reference Liu, Lu, Cui, Li and Fang2014; Wang et al. Reference Wang, Yang, Liu, Wu, Chen, Wu, Zhu and Bian2015; Yang et al. Reference Yang, Yang, Dai, Xu, Koide, Wang, Liu and Bian2015; Zhao et al. Reference Zhao, He, Qiu, Jia, Liu, Jin and Johnston2014). Also, straw return may have interactive effects with N fertilization on leaf [P] by releasing available P from litter and subsequently alleviating the P limitation caused by N addition (Li et al. Reference Li, Liu, Fan, Ma, Ding, Zhong and Wang2015). Similarly, several studies indicated that an increase of plant litter inputs in forests may regulate the effects of N fertilizer on leaf stoichiometry (An et al. Reference An, Frenne, Staelens, Verstraeten, Muys, Vesterdal, Wuyts, Nevel, Schelfhout and Neve2012; Clarholm and Skyllberg Reference Clarholm and Skyllberg2013). Given the potential effects of straw return on soil properties, straw return may interact with N fertilization to alter leaf stoichiometry. No experiment has, however, examined how straw return alters the N effects on leaf stoichiometry.
Here, we conducted a field experiment to investigate the influence of N fertilizer application on leaf N:P:K stoichiometry in rice and four coexisting weeds (a variety of barnyardgrass [Echinochloa crus-galli (L.) P. Beauv.], Monochoria korsakowii Regel and Mack, alligatorweed [Alternanthera philoxeroides (Mart.) Griseb.], and Japanese mazus [Mazus pumilus (Burm. f.) Steenis]) in Liyang, Jiangsu Province, China. These weeds were selected because they are the most common and troublesome weeds in rice paddies (Zhang et al. Reference Zhang, Gu, Zhao, Yang, Peng, Li and Bai2017). The uncontrolled growth of these weeds, especially under nutrient enrichment, can exert suppressive effects on rice and lead to substantial reduction in rice yield. Further, we determined that straw return significantly affected the N effects. Our study may advance the understanding of the stoichiometric responses of rice and weeds to N addition and straw return and provides new insights into weed management in agroecosystems.
Materials and methods
The experimental site
This study was conducted at the experimental fields in Liyang City, Jiangsu Province, located south of the Yangtze River delta (119.28°E, 31.45°N). This region has a northern subtropical monsoon climate with an average annual temperature of 15.5 C (mean monthly temperatures range from 2.7 C in January to 28.1 C in July), and mean annual precipitation is 1,152.1 mm. The soil is classified as white soil, with a soil pH (extracted with ultrapure water, 1:2.5) of ~6.4. The organic matter, total N, and available [P] and [K] in the 0- to 20-cm soil layer were 21.12, 1.28, 0.0085, and 0.072 mg g−1, respectively. The farming system in this region is a winter wheat–summer rice rotation. The major weeds in this ecosystem are E. crus-galli, M. korsakowii, A. philoxeroides, and M. pumilus.
The experimental design
This experiment employed a randomized block design with a control; three N fertilization treatments: (1) 120 kg N ha−1, (2) 240 kg N ha−1, and (3) 360 kg N ha−1 addition; and one straw return treatment: 240 kg N ha−1. Because the cropping system is a wheat–rice rotation, 2,000 kg ha−1 wheat straw was applied when transplanting rice, while 5,000 kg ha−1 rice straw was applied when planting wheat. Each treatment had four replicates. Accordingly, four blocks were established in mid-June 2011, and each was divided into five plots (3 × 6 m with 2-m buffers). In this study, we focused on the recommended quantity of straw return and commonly used fertilization level (240 kg N ha−1) rather than all the fertilization levels. Within each block, one plot was left as an unfertilized control and the other four plots were randomly assigned to fertilization and straw return treatments. Phosphate fertilizer (P2O5, 75 kg ha−1) and one-half of the K fertilizer (K2O, 45 kg ha−1) were used as basal fertilizers, with the remaining half of the K fertilizer (45 kg ha−1) applied at the grain-filling stage of rice. Over the rice season, the total N fertilizer was divided into four parts, with 40% used as basal fertilizer, and the remaining three equal parts (20% each) applied at the initial, tillering, and grain-filling stages. The straw was ground and ditch-buried at a depth of 20 cm. Rice seedlings were raised in mid-May and transplanted manually in mid-June with 25 cm × 13 cm between hills and rows, and the rice density was 2,050 seedlings ha−1.
Field sampling
In the present study, we focused solely on leaf stoichiometry of weeds and rice at the rice grain-filling stages after water flooding, which represent the mature period of rice and the critical period determining the crop yield (Yang and Zhang Reference Yang and Zhang2010). Understanding the nutrient demands of rice and weeds is extremely important for management at this stage. At the end of September 2016, five surface-soil cores (0- to 10-cm depth) were randomly collected from each plot using a 2.5-cm soil auger. Soil cores were pooled to form a composite soil sample for each plot. The soil samples were air-dried in a shaded and ventilated environment for approximately 1 mo to achieve a constant weight. The soil samples were sieved to remove organic material and rocks and then ground down to a size that enabled them to be passed through a 0.425-mm sieve using a glass mortar.
We collected the plant samples at the same time. Healthy plants were harvested and kept in a cooler during transport to laboratory. In the laboratory, we separated mature leaves from shoots. All the leaves were oven-dried at 65 C for 48 h and then ground to a 0.15 mm grain diameter with a Wiley mill (Thomas Model 4 Wiley® Mill, Thomas Scientific, Swedesboro, NJ, USA) and kept cool and dry for subsequent chemical analysis.
Chemical analyses
The [N] in both soils and leaves was determined using the following methods: Subsamples (0.1 g of plant tissues and 0.5 g of soil) were weighed to the nearest 0.001 mg for [N] analyses; [N] was determined using an elemental analyzer (2400II CHNS/O Elemental Analyzer, Perkin-Elmer, Waltham, MA, USA). The P in leaves was digested in a mixture of trace metal–grade H2SO4–HClO4, and [P] was determined using a modified Olsen method (according to the [P] in leaves) using a spectrophotometer. The available P (mg g–1) in soil was measured using a modified Olsen method after extraction with 0.5 mol L−1 NaHCO3. The [K] in leaves and available K in soil were measured using a flame spectrophotometer (Sherwood M410, Sherwood, Cambridge, UK) after being digested in a mixture of trace metal–grade H2SO4–HClO4 and extracted with 1 mol L−1 NH4OAC, respectively. Soil pH was measured in a 1:2.5 ratio of soil to deionized water.
Statistical analyses
We quantified the response of [N] of weeds and rice in response to N inputs using response ratios (ln RR), where ln RR was calculated as follows:
 $${\rm{In}}{{{X_{\rm{t}}}} \over {{X_{\rm{c}}}}} = {\rm{In}}{X_{\rm{t}}} - {\rm{In}}{X_{\rm{c}}}$$
$${\rm{In}}{{{X_{\rm{t}}}} \over {{X_{\rm{c}}}}} = {\rm{In}}{X_{\rm{t}}} - {\rm{In}}{X_{\rm{c}}}$$
where X t and X c are values of [N] in the experimental treatment and in the control within the same block.
All data were transformed when necessary to meet the parametric test assumptions of normality (Bartlett’s test) and homogeneous variances, and two-way mixed-effects ANOVA was used to explore the effects of N fertilizer and species on leaf stoichiometry. One-way ANOVA was used to determine the effects of N fertilizer treatment on soil nutrients and pH and leaf stoichiometry. Linear regression was employed to analyze the changes of [P], [K], N:P ratio, or N:K ratio with [N] in leaves. Data analyses were performed with R v. 3.0.2 (R Development Core Team 2005).
Results and discussion
Effects of N fertilizer on soil properties
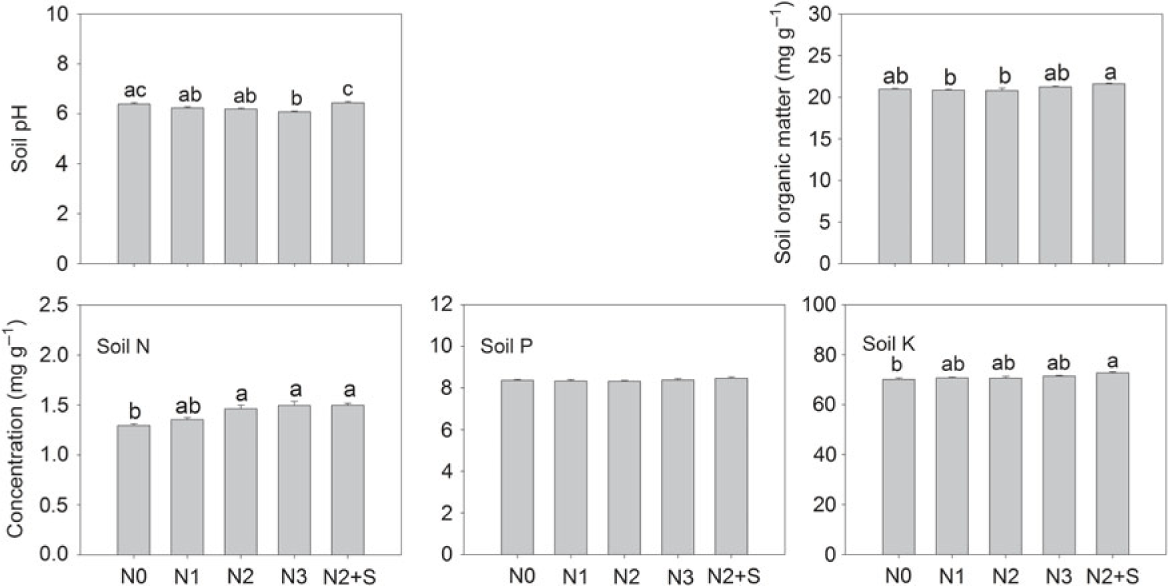
High level of N addition (360 kg N ha−1) significantly decreased soil pH, whereas straw return significantly dampened the negative effects of N fertilizer (Figure 1). N fertilizer had no significant effects on soil organic C, [P], and [K], but significantly increased total N in soil at addition rates of 240 and 360 kg N ha−1 (Figure 1). The combined treatment of N and straw return significantly increased [N] and [K] in soil but had no effects on [P] (Figure 1).
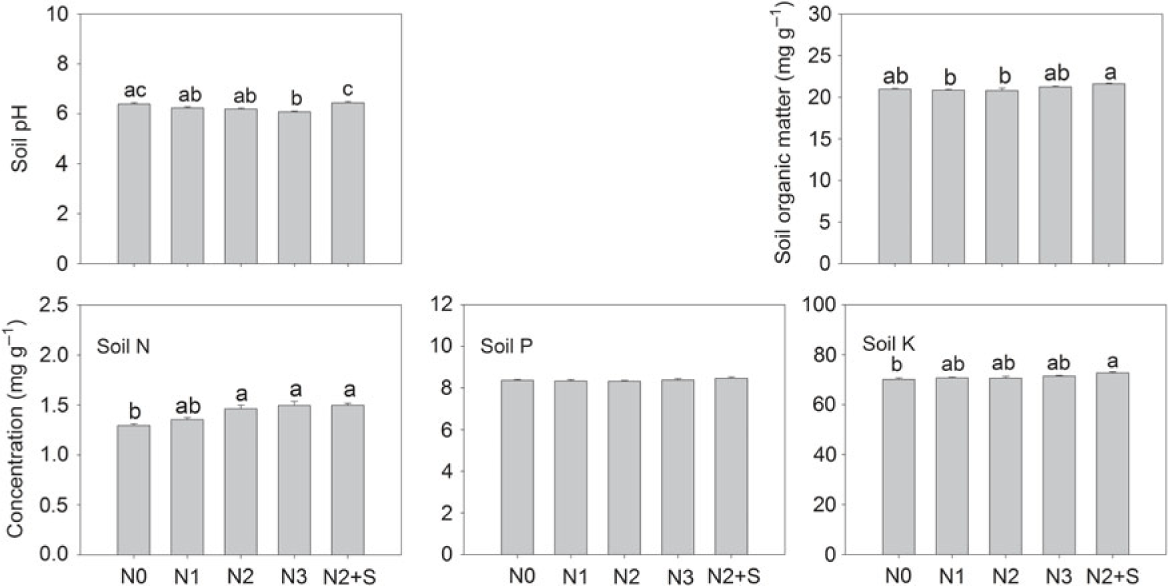
Figure 1. Changes in pH, organic matter, nitrogen (N), phosphorus (P), and potassium (K) concentrations in soil (total N, available P and K) in response to increased N fertilizer. N0, ambient N; N1, 120 kg N ha−1; N2, 240 kg N ha−1; N3, 360 kg N ha−1; and N2+S, 240 kg N ha−1 with straw. Different letters indicate a significant difference among treatments (Tukey’s HSD; P < 0.05).
Effects of N fertilizer on leaf N:P:K stoichiometry
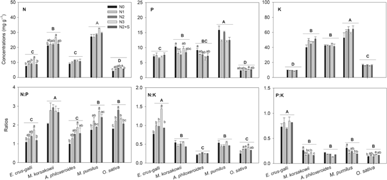
We only found that N addition at a rate of 360 kg N ha−1 significantly increased [N] in leaves of E. crus-galli and M. korsakowii (P < 0.05). By contrast, [N] in rice leaves was significantly increased at all fertilization levels (P < 0.05), but there were no significant differences among the three N fertilization levels (Figure 2). N addition showed no significant influence on [P] in leaves of rice, E. crus-galli, and M. pumilus, but decreased [P] in M. korsakowii and A. philoxeroides at rates of 240 and 360 kg N ha−1, respectively (Figure 2). There were no significant effects of N fertilizer on [K] in leaves of all species (Figure 2). These results were consistent with the first scenario that was proposed in terms of the structure–function chain hypothesis.

Figure 2. Effects of nitrogen (N) fertilizer addition on the concentrations of N, phosphorus (P), and potassium (K) and their ratios in mature leaves of four weed species and rice. N0, ambient N; N1, 120 kg N ha−1; N2, 240 kg N ha−1; N3, 360 kg N ha−1; and N2+S, 240 kg N ha−1 with straw. The uppercase letters show the different stoichiometric characteristics among species, while the lowercase letters show the different stoichiometric characteristics among N fertilization levels in each species (weeds: Echinochloa crus-galli, Monochoria korsakowii, Alternanthera philoxeroides, Mazus pumilus; crop: Oryza sativa). Different letters indicate a significant difference (Tukey’s HSD; P < 0.05).
The highest rate of N addition (360 kg N ha−1) significantly increased N:P ratio in E. crus-galli, A. philoxeroides, and M. pumilus, whereas the N:P ratio in rice leaves significantly increased under all N addition rates (Figure 2). Combined with straw return, N had no significant effects on leaf N:P ratios in weeds and rice (Figure 2). Although N at 360 kg N ha−1 significantly increased [N] in M. korsakowii, leaf N:P ratios did not change across the treatments (Figure 2). Surprisingly, N addition not only changed the N-related stoichiometry in leaves, but also significantly decreased P:K ratios in M. korsakowii and M. pumilus leaves and increased leaf P:K ratios in rice at the highest N addition rate (Figure 2).
Differences in leaf stoichiometry between weeds and rice
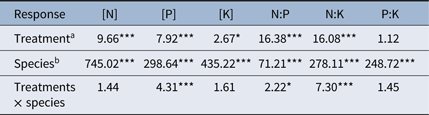
The leaf [N], [P], [K], and their ratios significantly varied among species (Table 1). The [N] and [P] in weed leaves were significantly higher than those in rice leaves, with highest [N] in M. pumilus (Figure 2). Mazus pumilus also had highest leaf [K], followed by M. korsakowii and A. philoxeroides, and then rice (Figure 2). Monochoria korsakowii had the highest N:P ratio, followed by M. pumilus and rice, whereas E. crus-galli and A. philoxeroides had similar lower leaf N:P ratios (Figure 2). The leaf N:K ratio was highest in E. crus-galli, but lowest in A. philoxeroides and rice (Figure 2). Echinochloa crus-galli also had a higher P:K ratio than the other three weeds and rice (Figure 2).
Table 1. F-value from two-way mixed-effects ANOVA relating nitrogen (N), phosphorus (P), and potassium (K) concentrations and their ratios in rice and weed leaves to levels of N fertilization and species

a Treatments: addition of (1) 120, (2) 240, and (3) 360 kg N ha−1, and (4) 240 kg N ha−1 with straw (2,000 kg ha−1 wheat straw, 5,000 kg ha−1 rice straw).
b Species: Echinochloa crus-galli, Monochoria korsakowii, Alternanthera philoxeroides, Mazus pumilus, and rice.
* P < 0.05.
** P < 0.01.
*** P < 0.0001.
The response rate of E. crus-galli leaf [N] to N addition was similar to that of rice leaves across fertilization levels, but the response rates of the other three weeds were lower (Figure 3). The response rates of the leaf [N] in some of weed species became more intense under high N fertilization (with the ln RR of leaf [N] in response to 360 kg N ha−1 being more than double the ln RR of leaf [N] in response to 120 kg N ha−1), whereas the response rate in rice leaves was similar at all N addition levels (the ln RR for response of rice leaf [N] to 360 kg N ha−1 only increased 38.9% compared with the ln RR for response of rice leaf [N] to 120 kg N ha−1) (Figure 3).

Figure 3. The response ratios (ln RR) of nitrogen (N) concentration in mature leaves of each species (weeds: Echinochloa crus-galli, Monochoria korsakowii, Alternanthera philoxeroides, Mazus pumilus; crop: Oryza sativa) to N additions. Different letters indicate a significant difference among treatments (Tukey’s HSD; P < 0.05).
Relationships among N, P, and K in leaves
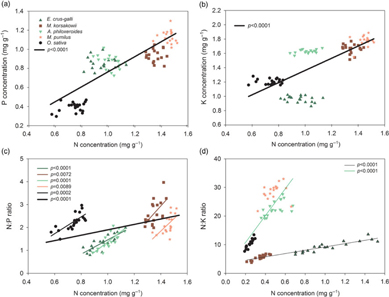
For each species, we did not find significant relationships among [N] and [P] or [K], but across the species, the [P] or [K] significantly increased with [N] (Figure 4A and B). The N:P ratio showed a significant positive relationship with increasing [N] for each species and across all species (Figure 4C). Although the N:K ratio also showed a positive relationship with [N] for each species, it remained unchanged with increasing [N] across all species. Furthermore, the four weeds and rice can be divided into two distinct groups: significant linear relationships between N:K ratios and [N] were seen across E. crus-galli and M. korsakowii, whereas the slope of the linear regression was much smaller than across the other two weed species and rice (Figure 4D).

Figure 4. The linear relationships between nitrogen (N) concentration and phosphorus (P) and potassium (K) concentrations and N:P and N:K ratios in mature leaves within each species (weeds: Echinochloa crus-galli, Monochoria korsakowii, Alternanthera philoxeroides, Mazus pumilus; crop: Oryza sativa), and across species.
Effects of N fertilizer on leaf N:P:K stoichiometry in rice and weed plants
We found that leaf [N] in weeds did not change at low (120 kg N ha−1) and regular (240 kg N ha−1) N fertilization levels but significantly increased at high N inputs (360 kg N ha−1). This result agrees with the two scenarios outlined in the “Introduction,” which suggest that weeds can maintain stoichiometric homeostasis under relatively low N supply levels, but not under high N levels. Compared with weeds, leaf [N] in rice was significantly increased by low and regular N fertilization but not further increased by high N input. These results suggest that rice can take up N more efficiently when available N is relatively low, whereas an excess supply of N in the soil would be more efficiently used by weeds. Similarly, a previous study reported that high N fertilization increased competition of a high N-demand weed with wheat (Blackshaw and Brandt Reference Blackshaw and Brandt2008). We also found [P] unchanged, even reduced, with increasing N input in rice and two weeds (i.e., M. korsakowii and A. philoxeroides), which is consistent with our assumption proposed in the “Introduction” (i.e., unchanged [P] in plant leaves is expected under higher N inputs). The decreased [P] can likely be ascribed to the dilution effect, which is caused by the rate of P uptake failing to keep pace with the rate of biomass increase under a higher N regime (Deng et al. Reference Deng, Hui, Dennis and Reddy2017; Yuan and Chen Reference Yuan and Chen2015b). The [K] on the basis of mass showed no response to N inputs (Figure 2), which is consistent with the structure–function chain prediction (Ågren and Weih Reference Ågren and Weih2012).
Our results also showed divergent effects of N input on leaf N:P:K stoichiometry. Specifically, the weeds maintained N:P and N:K homeostasis, but rice significantly increased N:P and N:K ratios in leaves when the supply rate of N fertilization was ≤240 kg N ha−1 (Figure 2). This result suggests that rice may take up more N than weeds when the N supply rate is ≤240 kg N hm−2. Under high fertilization conditions (360 kg N hm−2), however, the N:P and N:K ratios significantly increased in three weed species (Figure 2) because of the accelerating increase of [N] (Yuan and Chen Reference Yuan and Chen2015a). In contrast, there was no further increase of leaf N:K, and in fact even a decrease of N:P ratio, in rice at the highest N supply rate. This result further reflects that excess N may not be sufficiently used by rice. Meanwhile, high N inputs may simultaneously increase the competition of weeds for other elements, such as K. Moreover, N fertilization decreased the P:K ratio in M. korsakowii and M. pumilus, whereas it increased the P:K ratio in rice leaves, which may be caused by allometric responses in these species that affect [P] and [K] under N fertilization. These results suggest that N fertilization may not only alter coupled relationships between N and P, but may also change the coupled relationships between other elements. Because no significant changes in soil P and K were found among N treatments, the distinct responses of these P:K ratios to N addition were probably not caused by available nutrients in soil, but were driven by different physiological mechanisms in reaction to altered N supplies.
In terrestrial and aquatic plants, [N] and [P] always correlate positively with each other (Sterner and Elser Reference Sterner and Elser2002). In our study, significant relationships between [P], [K], and [N] were not found within species, but were detected across species (Figure 3). The N:P ratios across species were stable compared with those within species with increasing [N], while the N:K ratios remained unchanged across all species because of the distinct responses of the two species groups (Figure 3). These results suggest that the stoichiometric homeostasis of a plant community can be maintained by regulating species composition or individual plant stoichiometry response to N inputs (Ye et al. Reference Ye, Liang, Chen, Li, Ji and Zhu2014). In this case, N:P and N:K homeostasis at the community level can also be maintained by increasing the growth of high N-demand plants under excess N fertilization or individual plant stoichiometry, suggesting that the decoupling of N and P induced by N inputs in terrestrial plants at the community level can be dampened and offset by the changes in species growth.
Straw return effects on the responses of leaf stoichiometry to N fertilization
Our results showed that straw return dampened the influences of N fertilization on soil acidification, mainly by increasing basic cation nutrients, such as K+ (Figure 1). Straw return also weakened the positive effects of N fertilization on rice leaf N:P:K stoichiometry due to increased availability of P and K under straw return conditions (Wang et al. Reference Wang, Yang, Liu, Wu, Chen, Wu, Zhu and Bian2015), while it enhanced the positive effects on the N:P ratio in leaves of M. pumilus, reflecting that N released from straw was used by weeds. Similar results were previously found in grassland. For example, litter addition significantly dampened the effects of N inputs on leaf N:P (Li et al. Reference Li, Liu, Fan, Ma, Ding, Zhong and Wang2015).The different effects of straw returns on N:P ratios in rice and M. pumilus leaves may reflect different N demands by species, that is, lower N demand in rice at the mature stage, but higher N demand in weeds at the rapid growth stage. Thus, extra available N from straw was mainly used by weeds. These results suggest that we should return straw to alleviate the effects of N fertilization on rice leaf stoichiometry. However, future studies should consider the appropriate N fertilization to apply in combination with straw returns to decrease the N uptake by weeds.
Differences in leaf stoichiometry between weeds and rice
The different stoichiometric responses among species may primarily be explained in terms of their diverse nutrient demands (Güsewell Reference Güsewell2004). In our study, compared with rice at grain-filling stages, weed leaves contained higher elemental concentrations at all fertilization levels, especially M. korsakowii and M. pumilus. These differences are likely ascribed to the rapid growth of weeds at this stage, resulting in high nutrient requirements according to the growth rate hypothesis: plants require more nutrients, especially P, to support metabolism for a high growth rate, resulting in changes in the N:P ratio in leaves (Sterner and Elser Reference Sterner and Elser2002). Another possible explanation is that weeds were less able to compete for N compared with rice when N inputs were under 240 kg N ha−1. Under higher N inputs (>240 kg N ha−1), excess N would be used by weeds, resulting in disproportionally increased growth and competitive abilities in weeds. In contrast with weeds, rice had a relatively constant growth rate and lower nutrient requirements (Yan et al. Reference Yan, Guan, Han, Han, Guo and Fang2016). Therefore, high levels of N addition satisfy the requirements of weeds, especially M. korsakowii and M. pumilus, and then contribute to the high competitiveness of these weed species (González et al. Reference González, Kominoski, Danger, Ishida, Iwai and Rubach2010; Tilman Reference Tilman1982, Reference Tilman and Crawley1997). A previous study showed that the increase of competitive capacity for ruderal species with high N requirement was related to increased N availability, but for those with low N requirement, the effect of N fertilization was weak (Blackshaw and Brandt Reference Blackshaw and Brandt2008). These results, combined with the previous studies (Blackshaw and Brandt Reference Blackshaw and Brandt2008; Zhang et al. Reference Zhang, Gu, Li and Yang2016), further suggest that low N availability in soil may be more beneficial for rice competition, while high N availability in soil may be more beneficial for weeds. Moreover, low K and P fertilization at the grain-filling stage of rice might also be preferred to reduce weed competition.
In addition, N fertilization promotes planktonic and herbaceous species’ competitive abilities with high N:P ratios (except for legumes) (Mamolos et al. Reference Mamolos, Veresoglou and Barbayiannis1995; Tilman Reference Tilman and Crawley1997). In our study, the leaf N:P ratio was highest in M. korsakowii, whereas the leaf N:P ratios in rice were in the middle range of all measured plants. Given the strong influence of weeds on nutrient accumulation of rice under high N conditions (Zhang et al. Reference Zhang, Gu, Li and Yang2016), our results suggest that N supply should be applied in coordination with the requirements of the crop, as indicated by leaf stoichiometry, and controlled at a relatively low level to reduce nutrient uptake by weeds.
Significance for N management
These results suggest that N supply should be managed at a relatively low level at grain-filling stages to reduce the nutrient acquisition and competitive abilities of weeds without simultaneously decreasing the efficiency of rice N utilization. Certainly, future studies under field conditions will be needed to investigate the seasonal variation in stoichiometric demand of rice and weeds and how to manage fertilizer applications according to the requirements of rice. Meanwhile, other agricultural measures such as straw return should be considered to offset the effects of N on leaf stoichiometry. From a stoichiometric perspective, this study highlights the importance of N management in combination with straw return in controlling weeds and increasing the efficiency of nutrient use by the crop.
Author ORCID
Acknowledgments
We thank X. Xu, X. Yan, X. Luo, and J. Zhang for assistance in the field and lab. This work was supported financially by the National Natural Science Foundation of China (NSFC 600030), the Natural Science Foundation of Jiangsu Province (BK20161445), and the Key Project of NAU (0306J0887). No conflicts of interest have been declared.