

The Lower Devonian Posongchong Formation (Wenshan, Yunnan Province, southwestern China) comprises a series of continental clastic deposits, from which 28 vascular plant genera have been described to date (Hao & Xue Reference Hao and Xue2013, table 5.1). This plant megafossil flora represents a key part of the Early Devonian diversity. Zhu et al. (Reference Zhu, Wang and Fan1994) considered the Posongchong Formation to be late Pragian in age on the basis of fish assemblages. This age assignation was later supported by Gerrienne (Reference Gerrienne1996) based on a biostratigraphic coefficient analysis applied to plant megafossils. The latest reviews of the Posongchong Formation also consider a Pragian age for those deposits, the middle–late Pragian time interval being the most probable datation (Hao & Xue Reference Hao and Xue2013, pp. 23–24, and references therein).
Spore data are good stratigraphic indicators for correlating marine–terrestrial deposits as well providing an accurate temporal assignation to plant megafossils. From the 1980s, this has been routinely done using the spore zonation schemes from the Old Red Sandstone Continent (Richardson & McGregor Reference Richardson and McGregor1986) and the Ardenne-Rhenish region (Streel et al. Reference Streel, Higgs, Loboziak, Riegel and Steemans1987). Before the present study, the spore evidence from the Posongchong and Pojiao formations at the Zhichang section (Gumu Town, Wenshan) came from Wang's (Reference Wang1994) investigation. The author concluded that the Apiculiretusispora plicata-Dictyotriletes emsiensis assemblage zone can be recognised in the Posongchong Formation, and assigned a ‘Siegenian' age (according to the original nomenclature) to this formation. To reach this conclusion, Wang (Reference Wang1994) correlated the relevant palynological zones of China and Laurussia (North America and Western Europe). However, the spore zonation of the Chinese formations is preliminary due to the limited data and the complexity of the geological series.
Here, we present a new spore assemblage recently collected from the Posongchong Formation from the Zhichang and Changputang sections (Wenshan area, Yunnan Province). A new spore species is described (Aneurospora posongchongensis sp. nov.). This palynoflora is of significance in that (i) evidence comes from northeastern Gondwana, a palaeogeographical area where the spore diversity is less known in comparison with Laurussia, (ii) the sampled deposits correspond to the most diverse Early Devonian plant megafossil assemblages and (iii) a complex early terrestrial landscape appears well represented in the Posongchong Formation. The aim of this study is to provide new insights into the known spore diversity and phytogeography of the Early Devonian floras, especially from northeastern Gondwana, and to reduce uncertainties about the temporal assignation of the Posongchong flora.
1. Stratigraphy, material and methods
The Lower Devonian strata of Wenshan area (southeastern Yunnan, Fig. 1a) are well exposed (Fig. 1b). The strata at the Zhichang section (Zhichang slope) include, in ascending order, the Posongchong, Pojiao, Bajiaoqing and Gumu formations (Fig. 1b). Details of the stratigraphic sequence of the Posongchong Formation are provided in Figure 2. The samples analysed in this study were mainly collected from this section (Figs 1b, 2). The Posongchong Formation at the Zhichang section unconformably overlies the quartzose sandstone of the marine Lower Ordovician Meitan Formation, and has a conformable contact with the overlying marine Pojiao Formation (Jin et al. Reference Jin, Shen, Chen, Lu, Wei, Wang and Xie2005; Hao & Xue Reference Hao and Xue2013). Various endemic Early Devonian plants have been discovered from this section, e.g., Adoketophyton subverticillatum (Hao et al. Reference Hao, Wang and Beck2003), Gumuia zyzzata (Hao Reference Hao1989), Guangnania cuneata (Wang & Hao Reference Wang, Hao and Liu2002), Ramoferis amalia (Hao & Xue Reference Hao and Xue2011) and Zhenglia radiata (Hao et al. Reference Hao, Wang, Wang and Xue2006; Fig. 2). The Changputang section (Tiechan slope) was also investigated (Fig. 1b). The stratigraphy and the lithology of this section are similar to those of the Zhichang section (Hao & Gensel Reference Hao and Gensel1998; Jin et al. Reference Jin, Shen, Chen, Lu, Wei, Wang and Xie2005). Several endemic plants have also been described from the Changputang section, e.g., Catenalis digitata (Hao & Beck Reference Hao and Beck1991) and Celatheca beckii (Hao & Gensel Reference Hao and Gensel1995).
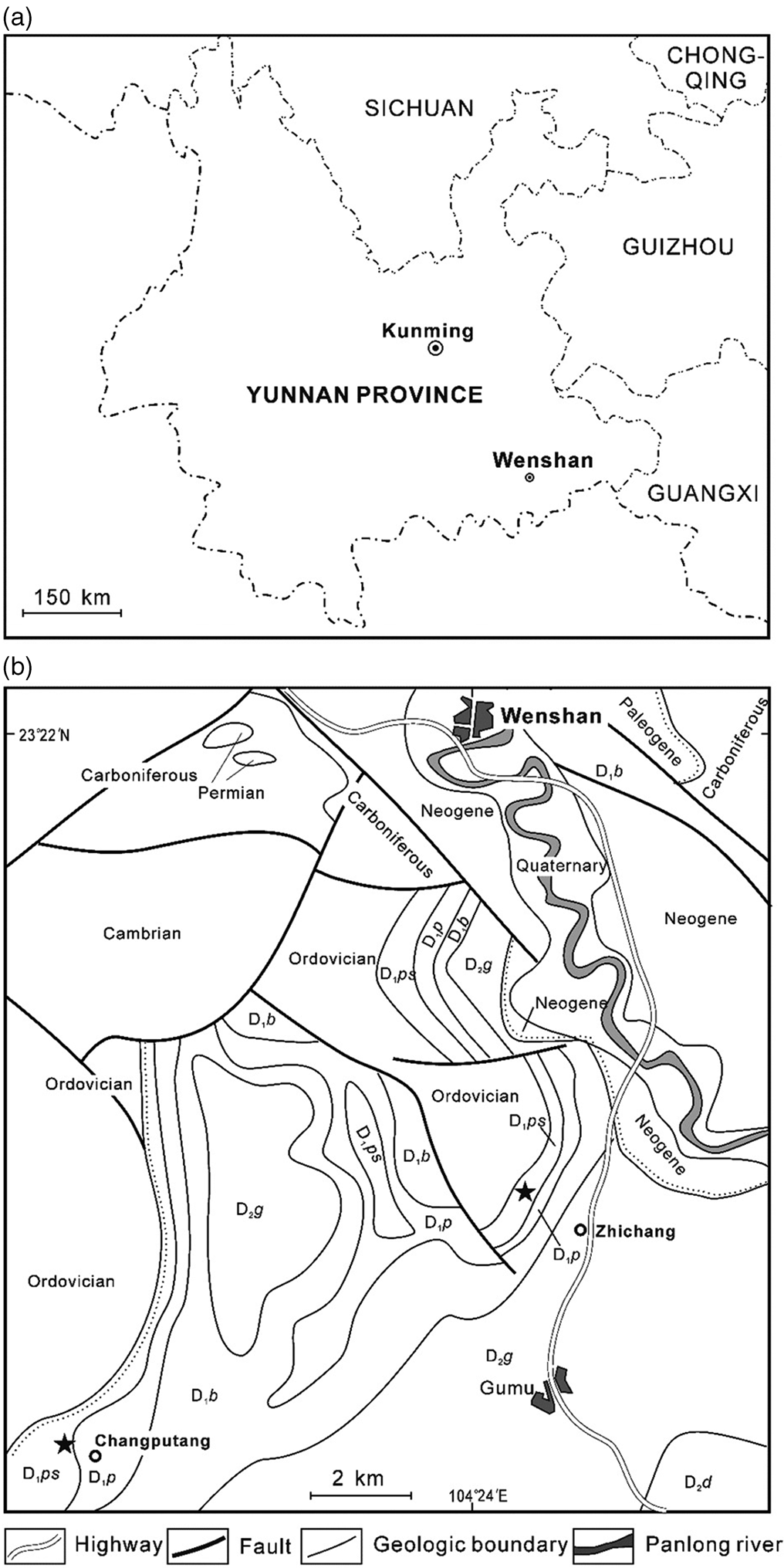
Figure 1 Map showing location of studied section: (a) general view of Yunnan Province (southwestern China) showing position of study area (Wenshan), modified from Hao & Xue (Reference Hao and Xue2013, fig. 2.1); (b) location of the Zhichang and Changputang sections in the Wenshan area, modified from Hao & Xue (Reference Hao and Xue2013, fig. 2.3). Abbreviations: D2d = Middle Devonian Donggangling Formation; D2g = Middle Devonian Gumu Formation; D1b = Lower Devonian Bajiaoqing Formation; D1p = Lower Devonian Pojiao Formation; D1ps = Lower Devonian Posongchong Formation.
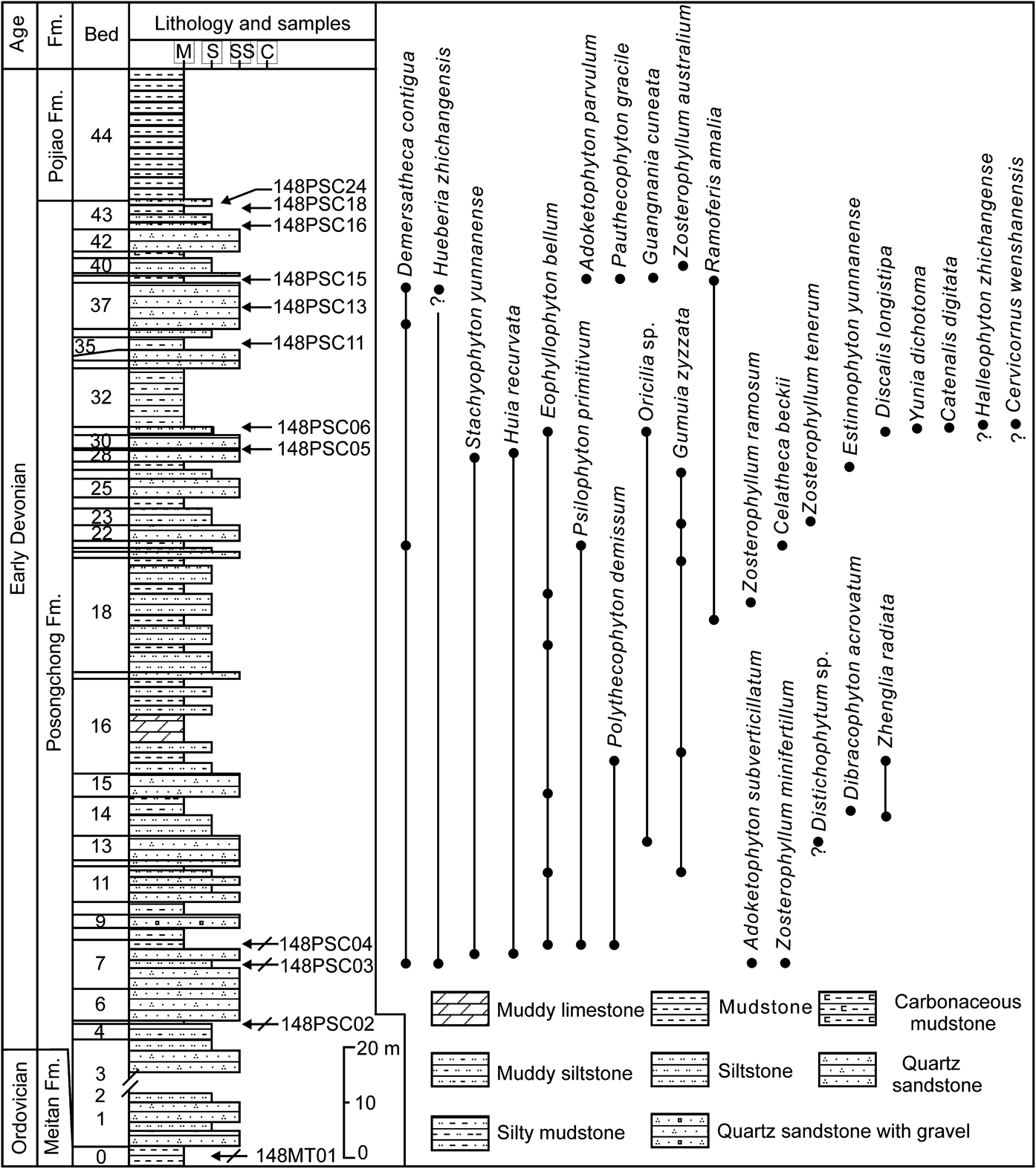
Figure 2 Stratigraphy of the Lower Devonian Posongchong Formation at the Zhichang section, Wenshan, Yunnan, southwestern China. Sampled levels indicated by arrows. Plant fossils recovered from this section are also indicated. Modified from Hao & Xue (Reference Hao and Xue2013, fig. 3.4).
Altogether, we collected 12 palynological samples from the Zhichang section and one from the Changputang section (see sample details in Table 1). Rock samples (approximately 30g each) were treated using standard HF-HCL-HF acid maceration. Following maceration, the remaining residue was briefly oxidised in HNO3 and KClO3 and sieved through a 12μm mesh to remove particles of organic matter and fine mineral matter. Afterwards, a 25% HCl hot bath was used to eliminate the remaining fine mineral particles. All samples were finally rinsed through a 12μm mesh. The remaining organic residue was rich in well-preserved palynomorphs dominated by spores and phytodebris. No acritarchs were found.
Table 1 Details of studied samples from the Posongchong Formation.

2. Systematic palaeontology
Genus Aneurospora Streel emend. Richardson et al., Reference Richardson, Streel, Hassan and Steemans1982
Type species. Aneurospora goensis Streel, Reference Streel1964
Aneurospora posongchongensis sp. nov.
(Plate 1, 4–8)

Plate 1 Posongchong spore diversity (I). 1–2. Ambitisporites avitus Hoffmeister, Reference Hoffmeister1959 (74474 V36/1, 74469 M39/2). 3. Aneurospora conica (Lu & Ouyang) Wellman et al., Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012 (74473 N36/2, 74474 H45/2). 4–8. Aneurospora posongchongensis sp. nov. (74482 H39/0, 74482 C50/0, 74482 H43/1, 74482 D38/2, 74482 M43/2-Holotype). 9. Aneurospora xuchiachongensis Wellman et al., Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012 (74480 D49/3). 10–11. Aneurospora sp. A (74473 E29/1, 74474 P47/1, 74474 H45/2). 12. Aneurospora sp. B (74475 E37/0). 13–14. Apiculiretusispora plicata (Allen) Streel, Reference Streel1967 (74468 G52/0–4, 74470 G41/0). 15. Apiculiretusispora cf. arabiensis Al-Ghazi, Reference Al-Ghazi2009 (74473 S5/0 4). 16. Archaeozonotriletes chulus (Cramer) Richardson & Lister, Reference Richardson and Lister1969 (74477 U40/2). 17–21. Biornatispora cf. dubia (McGregor) Steemans Reference Steemans1989 (74473 L51/0, 74473 R35/4, 74474 D44/1, 74480 H47/0, 7448/0 D48/4). 22. Camarozonotriletes parvus Owens, Reference Owens1971 (sensu Steemans, Reference Steemans1989) (74477 J32/3). 23–24. Camarozonotriletes sp. A (74469 G35/4, 74473 O41/4). Slides housed in the Palaeobiogeology–Palaeobotany–Palaeopalynology Unit, Liege University collections. Scale bar = 20μm.
Holotype. Plate 1, 8 (slide 74482, England Finder M43/2).
Paratype. Plate 1, 7 (slide 74482, England Finder H43/1).
Type locality and horizon. Zhichang section, Wenshan, Yunnan Province, southwestern China; upper part of the Posongchong Formation.
Age. Lower Devonian; Pragian (but not early).
Derivation of name. From the formation name.
Diagnosis. Trilete spore ornamented distally by regularly distributed baculae with a conical top. The base of the baculae is polygonal, 0.5–1.0μm in diameter and separated by more than 0.5–3.0μm. The cingulum is 2.0–3.5μm wide.
Description. Trilete spore with subcircular to subtriangular amb, with an equatorial cingulum of 2–3.5μm. Laesurae straight, simple, 2/5 to 7/10 of the length of spore radius. Proximal face laevigate. Distal and equatorial regions are sculptured with evenly distributed conical baculae, 0.5–1.0μm wide and 1.0–1.5μm high. The base of the baculae is polygonal and the distance between sculptures is irregular, ranging from 0.5 to 3.0μm. The sculptures tend to be concentrated in the centre of the distal face. On some specimens, the cingulum is not clearly visible.
Dimensions. 27–(33)–43μm; 20 specimens measured.
Comparison and remarks. Some specimens are similar in size and number of baculae to Aneurospora conica (Lu & Ouyang) Wellman et al., Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012. However, the new species bears a cingulum and the base of the baculae is polygonal. Aneurospora posongchongensis sp. nov. is close to Aneurospora cf. tojoides (Cramer) Steemans, Reference Steemans1989, but it is smaller, with a smaller number and a lower density of baculae; furthermore, it lacks prominent labra.
Aneurospora sp. A
(Plate 1, 10–11)
Description. Trilete spore with sub-circular amb. Equatorial cingulum up to 2μm wide. Laesurae straight and simple, and extending up to the margin of the amb. Proximal face laevigate. Distal and equatorial regions are sculptured with irregularly distributed coni separated by up to 5μm. The base of the cone is polygonal/rounded, 1.0–1.5μm wide and 1.0–1.5μm apart.
Dimensions. 30–(33)–37μm; three specimens measured.
Comparison. Cymbosporites proteus McGregor & Camfield, Reference McGregor1976 has more dense and regular ornamentation than Aneurospora sp A. Aneurospora cf. tojoides (Cramer) Steemans, Reference Steemans1989 has prominent labra and the separation between the coni is regular. Aneurospora sp. A and Aneurospora posonchongensis are similar in amb and size, but differ in the ornamentation. The base of the coni in Aneurospora sp. A is slightly larger (1.0 to 1.5μm) than in Aneurospora posonchongensis (0.5 to 1.0μm).
Dimensions. 30–(33)–37μm; three specimens measured.
Aneurospora sp. B
(Plate 1, 12)
Description. Trilete spore with subtriangular amb. Equatorial cingulum up to 2.5μm wide. Laesurae straight, simple, extending 2/5 to 7/10 of the length of the spore radius. Proximal face laevigate. Distal and equatorial regions sculptured with grana. The base of the sculptures is rounded to polygonal, 0.5μm wide, up to 0.5μm high and 0.5μm apart. The verrucae are connected at their base, forming convoluted muri, up to 4.0μm long.
Dimensions. 35μm; one specimen measured.
Genus Camarozonotriletes Naumova ex Naumova, Reference Naumova1953
Type species. Camarozonotriletes devonicus Naumova, Reference Naumova1953
Camarozonotriletes sp. A
(Plate 1, 23–24)
Description. Trilete spores with amb sub-circular with rounded corners. Laesurae visible in one of the specimens, simple, straight and extending to the inner margin of the equatorial cingulum, 2.5μm wide. The exine is equatorially and distally granulate. The sculptural elements are less than 1.0μm high. Some sculptures are connected, forming rugulate ornaments of no more than five elements.
Dimensions. 30–(31)–32μm; two specimens measured.
Genus Chelinospora Allen emend. McGregor & Camfield, Reference McGregor and Camfield1976
Type species. Chelinospora concinna Allen, 1965
Chelinospora sp. A
(Plate 2, 2)

Plate 2 Posongchong spore diversity (II). 1. Camarozonotriletes? cf. luii Wellman et al., Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012 (74481 Q43/2). 2. Chelinospora sp. A. (74481 L44/1). 3. Concentricosisporites agradabilis (Rodríguez) Rodríguez, Reference Rodríguez1983 (74649 H39/1). 4. Convolutispora sp. A (74474 G33/2). 5. Convolutispora? sp. B (74474 L46/3). 6–7. Dibolisporites echinaceus (Eisenack) Richardson, Reference Richardson1965 (74470 M36/3, 74471 F33/2). 8. Dibolisporites sp. A (74482 Q37/1). 9. Dibolisporites sp. B (74482 K30/0). 10. Emphanisporites rotatus (McGregor) McGregor, Reference McGregor1973 (74482 E41/2). 11–12. Gneudnaspora divellomedia (Chibrikova) Balme, Reference Balme1988 (74468 T51/1, 74473 O39/4). 13–14. Grandispora sp. A (74481 E49/2, 74481 Q37/4). 15–16. Latosporites ovalis Breuer, Reference Breuer2007 (74480 K40/4, 74470 P42/0). 17–18. Leiozonospora xichongensis Wellman et al., Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012 (74481 Q43/2, 74474 E36/03). 19–20. Retusotriletes triangulatus (Streel) Streel, Reference Streel1967 (74473 D28/0, 74471 H28/1). Slides housed in the Palaeobiogeology–Palaeobotany–Palaeopalynology Unit, Liege University collections. Scale bar = 20μm.
Description. Amb sub-circular. Laesurae thick, extending to the end of the inner body. Patina sculptured with broad reticulum. Muri 0.5μm wide and 1.0μm high. At junctions, the muri widen and form rounded verrucae.
Dimensions. 21.5μm; one specimen measured.
Genus Convolutispora Hoffmeister et al., Reference Hoffmeister, Staplin and Malloy1955
Type species. Convolutispora subtilis Owens, Reference Owens1971
Convolutispora sp. A
(Plate 2, 4)
Description. Amb circular. Distal face with convoluted ornamentation. The ornamentation is formed by grana closely attached together and forming rugulae, 0.5μm apart and 1.0μm high.
Dimensions. 42μm; one specimen measured.
Convolutispora? sp. B
(Plate 2, 5)
Description. Amb circular. Distal face densely ornamented. The sculpture is formed by grana closely attached together of no more than five or six elements forming rugulae, up to 0.5μm wide, up to 0.5μm apart and 1.0μm high.
Comparison. Convolutispora? sp. B and Convolutispora sp. A are similar in size, but in Convolutispora? sp. B, the size of the grana is larger and the number of attached elements does not exceed six.
Dimensions. 44μm; one specimen measured.
Genus Dibolisporites Richardson, Reference Richardson1965
Type species. Dibolisporites echinaceus (Eisenack)
Richardson, Reference Richardson1965
Dibolisporites sp. A
(Plate 2, 8)
Description. Trilete spores with amb sub-triangular. The exine is 3.0–3.5μm thick equatorially. Equatorial and distal region is ornamented. The sculptures consist of biform ornaments with a cone 3.5–4.0μm wide at base, 2.5–3.0μm high and surmounted by a spine 0.5μm long.
Dimensions. 50μm; one specimen measured.
Dibolisporites sp. B
(Plate 2, 9)
Description. The preservation of the spore does not allow for the precise defining of the amb. Equatorial and distal regions ornamented. The sculptures consist of biform ornaments: cones 1.0μm wide at the base, 2.0–3.0μm high and surmounted by spines 0.5μm high. The specimen presents zones where the ornamentation is widely separated or absent.
Comparison. Dibolisporites sp. A is covered by a dense ornamentation and the sculptures are wider at their base. It is possible that Dibolisporites sp. A and Dibolisporites sp. B represent the same species because they show comparable size and sculptures, but this is not possible to ascertain because there is currently only one specimen for each.
Dimensions. 50μm; one specimen measured.
Genus Grandispora Hoffmeister et al. emend.
Neves & Owens, Reference Neves and Owens1966
Type species. Grandisposa spinosa Hoffmeister et al., Reference Hoffmeister, Staplin and Malloy1955
Grandispora sp. A
(Plate 2, 13–14)
Description. Camerate spore with a sub-circular amb. Laesurae not visible. The inner body diameter is 2/3 to 4/7 of the total diameter. Inner body is laevigate. The outer layer is sculptured by grana. Elements are up to 1.0μm wide, round to polygonal at the base and up to 0.5μm apart.
Comparison. Grandispora sp. A as described in Steemans et al. (Reference Steemans, Rubinstein and Melo2008) is similar but smaller in size (25–(29)–32μm).
Dimensions. 30–(40)–50μm; two specimens measured.
Genus Retusotriletes Naumova emend. Streel, Reference Streel1964
Type species. Retusotriletes simplex Naumova, Reference Naumova1953
Retusotriletes sp.
(Plate 3, 2)

Plate 3 Posongchong spore diversity (III). 1. Retusotriletes cf. rotundus Streel emend. Lele & Streel, Reference Lele and Streel1969 (74475 H38/3). 2. Retusotriletes sp. (74474 G37/04). 3–4. Tetrahedraletes medinensis Strother & Traverse emend. Wellman & Richardson, Reference Wellman and Richardson1993 (74474 N39/4, 74477 R35/4). 5. Verrucosporites polygonalis Lanninger, Reference Lanninger1968 (74482 H50/2). 6–7. Verrucosisporites cf. polygonalis Lanninger, Reference Lanninger1968 (74480 N47/4, 74480 H33/1). 8–9. Verrucosisporites sp. (74470 V30/0, 74473 Q40/0–4). 10. Specimen 74483 X48/4 (zonate spore). 11–12. Fragment of spore showing distinct types of ornament on two distinct parts (74474 F47/2, 74474 Q51/1). Wellman et al. (Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012, fig. 8N) illustrated and commented on a similar phenomenon. Slides housed in the Palaeobiogeology–Palaeobotany–Palaeopalynology Unit, Liege University collections. Scale bar = 20μm.
Description. Amb sub-circular. Laesurae straight and thin, up to 0.5μm wide, extending up to the end of the amb. Proximal and distal faces are laevigate.
Dimensions. 27–(32.5)–40μm; six specimens measured.
Genus Verrucosisporites Ibrahim emend. Smith, Reference Smith, Alpern and Neves1971
Type species. Verrucosisporites verrucosus (Ibrahim)
Ibrahim, 1933
Verrucosisporites sp.
(Plate 3, 8–9)
Description. Amb sub-triangular. Laesurae curved and 2.5μm thick, 2/5 to 3/5 of the length of the amb radius. Proximo-equatorial and distal regions are sculptured with verrucae and pilae, 2.5 to 5.0μm wide, 2.0 to 3.5μm high and up to 0.5μm apart.
Dimensions. 30–(31)–32μm; two specimens measured.
3. Spore diversity from the Posongchong Formation
Nine of the 13 samples were productive (Table 1). The palynological analysis shows a high spore diversity, with 18 genera and 32 species. The dispersed spores include: Ambitisporites avitus (Plate 1, 1–2), Aneurospora conica (Plate 1, 3), Aneurospora posongchongensis sp. nov. (Plate 1, 4–8), Aneurospora xuchiachongensis (Plate 1, 9), Aneurospora sp. A (Plate 1, 10–11), Aneurospora sp. B (Plate 1, 12), Apiculiretusispora plicata (Plate 1, 13–14), Apiculiretusispora cf. arabiensis (Plate 1, 15), Archaeozonotriletes chulus (Plate 1, 16), Biornatispora cf. dubia (Plate 1, 17–21), Camarozonotriletes parvus (Plate 1, 22), Camarozonotriletes sp. A (Plate 1, 23–24), Camarozonotriletes? cf. luii (Plate 2, 1), Chelinospora sp. A. (Plate 2, 2), Concentricosisporites agradabilis (Plate 2, 3), Convolutispora sp. A (Plate 2, 4), Convolutispora? sp. B (Plate 2, 5), Dibolisporites echinaceus (Plate 2, 6–7), Dibolisporites sp. A (Plate 2, 8), Dibolisporites sp. B (Plate 2, 9), Emphanisporites rotatus (Plate 2, 10), Gneudnaspora divellomedia (Plate 2, 11–12), Grandispora sp. A (Plate 2, 13–14), Latosporites ovalis (Plate 2, 15–16), Leiozonospora xichongensis (Plate 2, 17–18), Retusotriletes triangulatus (Plate 2, 19–20), Retusotriletes cf. rotundus (Plate 3, 1), Retusotriletes sp. (Plate 3, 2), Tetrahedraletes medinensis (Plate 3, 3–4), Verrucosporites polygonalis (Plate 3, 5), Verrucosisporites cf. polygonalis (Plate 3, 6–7) and Verrucosisporites sp. (Plate 3, 8–9). Interestingly, a zonate spore (Plate 3, 10) and several small fragments of spores showing distinct types of ornamentation are also observed (Plate 3, 11–12) – a phenomenon previously illustrated and commented upon by Wellman et al. (Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012, fig. 8N).
Wang (Reference Wang1994) previously documented 27 spore taxa from the Posongchong Formation (Table 2). Our palynoflora shares three species (Apiculiretusispora plicata, Dibolisporites echinaceus, Verrucosporites polygonalis) with Wang's result. Amongst the spores described by Wang (Reference Wang1994), we think that seven of his taxa could be re-interpreted as follows: Brochotriletes sp. B (=cf. Biornatispora dubia in the present study), Camptozonotriletes cf. caperatus (=?Camptozonotriletes cf. caperatus), Camptozonotriletes sp. G (=cf. Camptozonotriletes macrospinosus), Crissisporites guangxiensis (=?Crissisporites guangxiensis), Cyclogranisporites sp. (=?Cyclogranisporites sp.), Cymbosporites raistrickiaeformis (=Dibolisporites wetteldorfensis) and Punctatisporites sp. (=Retusotriletes spp.). The spore diversity of the Posongchong Formation also includes 17 other taxa from Wang's (Reference Wang1994) study, namely: Apiculiretusispora pygmaea, Apiculiretusispora weenshanensis, Brochotriletes? foveolatus, Calamospora cf. microrugosa, Calamospora cf. panucea, Camarozonotriletes sextantii, Cymbosporites echinatus, Dibolisporites eifeliensis, Dictyotriletes emsiensis, Dictyotriletes gorgoneus, Dictyotriletes subgranifer, Emphanisporites cf. decoratus, Emphanisporites cf. neglectus, Raistrickia sp., Retusotriletes cf. triangulatus, Retusotriletes rotundus and Retusotriletes warringtonii. Thus, combining the results of this study and Wang (Reference Wang1994), the currently known Posongchong spore diversity is composed of at least 56 different morphological species belonging to 28 genera (see supplementary material for the list (available at https://doi.org/10.1017/S1755691018000233)), which is consistent with the impressive plant diversity and disparity documented from this formation (Fig. 2).
Table 2 Spore taxa reported by Wang (Reference Wang1994) from the Posongchong Formation, Wellman et al. (Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012) from the Xujiachong Formation and this study (including those common to Wang Reference Wang1994).

*Retusotriletes rotundus (Streel) Streel Reference Streel1967 in original source.
Wellman et al. (Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012) recently studied the spore diversity of the Xujiachong Formation from Yunnan Province (see Table 2), which is more or less coeval with the Posongchong Formations (Hao & Xue Reference Hao and Xue2013, table 2.1). Moreover, outcrops of the Xujiachong and Posongchong formations (Quijing and Wenshan, respectively) are located in adjacent zones within the South China Plate (Fig. 3). At least 12 spore species (Ambitisporites avitus, Aneurospora conica, Aneurospora xuchiachongensis, Apiculiretusispora plicata, Archaeozonotriletes chulus, Camarozonotriletes? cf. luii, Latosporites ovalis, Leiozosterospora xichongensis, Retusotriletes cf. triangulatus, Retusotriletes cf. rotundus, Tetrahedraletes medinensis and Verrucosisporites polygonalis), which represents around 52% of observed spore diversity from the Xujiachong Formation, have also been collected from the Posongchong Formation.

Figure 3 Palaeogeographical map showing the distribution of the major outcrops of fossil floras (Whensan and Guangnan, Posongchong Formation; Quijing, Xujiachong Formation; Yanmenba, Pingyipu Group) of the South China Plate. Green, blue and yellow colours represent land, shallow seas and estuarine conditions, respectively. Modified from Edwards et al. (Reference Edwards, Geng and Li2016).
4. Description of palynomorph assemblages
The distribution and abundance of the spore taxa within the studied samples are shown in Table 3. The spore assemblages are dominated by crassitate apiculate spores (Aneurospora spp.). This spore group represents the maximum observed values of abundances (e.g., 77.5% in sample 148PSC24). Three Aneurospora species are present, Aneurospora conica, Aneurospora posongchongensis sp. nov. and Aneurospora xujiachongensis, but only one of them (Aneurospora posongchongensis) shows high abundance values (25.5% and 29.5% in samples 148PSC16 and 148PSC18, respectively). The second dominant group includes laevigate retusoid spores (Retusotriletes spp.). Because of its morphological simplicity, this spore group is difficult to discriminate. Indeed, only a well-defined species (Retusotriletes triangulatus) was identified. It shows high abundance values (14.5–24% in samples 148PSC11, 148PSC13 and 148PSC14). The third most characteristic group consists of the apiculate retusoid spores (Apiculiretusispora spp.). Two species are recognised (Apiculiretusispora cf. arabiensis and Apiculiretusispora plicata), but only the latter is quantifiable. Dibolisporites, Verrucosisporites or Archaeozonotriletes are uncommon. For instance, Archaeozonotriletes chulus is only quantifiable in a single sample (2.5% in sample 148PSC13). Interestingly, some specimens of the simple laevigate monolete spore, Latosporites ovalis, are present. They occur in four samples (148PSC5, 148PSC6, 148PSC11, 148PSC15; 0.5–1.5%). Cryptospores are also present, but the only representatives are hilate cryptospores (Gneudnaspora divellomedia) and permanent tetrads (Tetrahedraletes medinensis).
Table 3 Distribution and abundance (in percentage) of the spore taxa in the studied samples. P, present but not featured in counts.

5. Biostratigraphical interpretation
The Apiculiretusispora plicata–Dictyotriletes emsiensis (PE) zone recognised from the Posongchong Formation by Wang (Reference Wang1994) correlates with the Pragian of the Old Red Sandstone Continent (Streel Reference Streel1967; Steemans Reference Steemans1981, Reference Steemans1989; Streel et al. Reference Streel, Fairon-Demaret, Otazo-Bozo and Steemans1981, Reference Streel, Higgs, Loboziak, Riegel and Steemans1987; Richardson & McGregor Reference Richardson and McGregor1986; Fig. 4). Definitively, the PE zone according to Wang (Reference Wang1994) correlates with the polygonalis–emsiensis Spore Assemblage Biozone (PE) sensu Richardson & McGregor (Reference Richardson and McGregor1986) and the equivalent polygonalis–wetteldorfensis Oppel Zone (PoW) sensu Streel et al. (Reference Streel, Higgs, Loboziak, Riegel and Steemans1987) (Fig. 4). The PE/PoW spore assemblage is considered early (but not earliest) Pragian to? earliest Emsian in age (Wellman Reference Wellman2006; Wellman et al. Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012).

Figure 4 Comparison of the uppermost Silurian–lowermost Middle Devonian spore and plant zonations from Laurussia and South China. *Miospore zones based on Streel et al. (Reference Streel, Higgs, Loboziak, Riegel and Steemans1987): MN = micrornatus–newportensis Oppel Zone; BZ = breconensis–zavallatus Oppel Zone; PoW = polygonalis–wetteldorfensis Oppel Zone; AB = annulatus–bellatulus Oppel Zone; FD = foveolatus–dubia Oppel Zone; AP = apiculatus–proteus Oppel Zone. **Miospore zones based on Gao (Reference Gao1981): I = Synorisporites verrucatus–Streelispora newportensis Assemblage Zone; II = Streelispora granulate–Archaeozonotriletes chulus Assemblage Zone; III = Emphanisporites neglectus–Brochotriletes sp. Assemblage Zone; IV = Emphanisporites annulatus–Dictyotriletes emsiensis Assemblage Zone; V = Rhabdosporites langii–Grandispora velata Assemblage Zone. ***Miospore zones according to Fang et al. (Reference Fang, Cai, Wang, Li, Wang, Geng, Wang, Gao, Wang and Li1994): SN = Apiculiretusispora spicula–Emphanisporites neglectus Assemblage Zone, Yulongsi Formation, Qujing (Yunnan); NC = Emphanisporites micrornatus–Streelispora newportensis Assemblage Zone, Xiaxishacun Formation, Qujing (Yunnan). ****Miospore zones according to Wang (Reference Wang1994): PE = Apiculiretusispora plicata–Dictyotriletes emsiensis Assemblage Zone, Posongchong Formation, Wenshan (Yunnan); ES = Dibolisporites eifeliensis–Camarozonotriletes sextantii Assemblage Zone, Pojiao Formation, Wenshan (Yunnan). *****Miospore zone according to Hsü & Gao (Reference Hsü and Gao1991): VL = Calytosporites velatus–Rhabdosporites langii Assemblage Zone, Chuandong Formation, Qujing (Yunnan). Spore and plant biozones from Laurussia are based on Edwards et al. (Reference Edwards, Fairon-Demaret and Berry2000, fig. 2). Abbreviations: DEV = Devonian.
The temporal distribution of the key Posongchong spores is presented in Figure 5. Our results suggest that the time interval covered by the Posongchong spore assemblages corresponds to the Pragian. This age assignation is supported by the presence of index species of the PoW zone, such as Verrucosporites polygonalis, Dictyotriletes subgranifer and Camarozonotriletes parvus (sensu Steemans, Reference Steemans1989) (Table 2; supplementary material). Importantly, Camarozonotriletes parvus (sensu Steemans, Reference Steemans1989) has only been described from the Pragian of Belgium and Germany, and is characteristic of the Pa Interval Zone of Streel et al. (Reference Streel, Higgs, Loboziak, Riegel and Steemans1987), i.e., the Pragian (but not early) horizons of the PoW zone. This view is also in agreement with the presence of Aneurospora conica, Aneurospora xujiachongensis and Leiozonospora xichongensis, three spore species documented only from the Xujiachong Formation of Qujing, Yunnan, a sequence correlative to the PE/PoW spore assemblage from Euramerica (Wellman et al. Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012). To summarise, the presence of the aforementioned taxa, together with Latosporites ovalis, a taxon that ranges from the Pragian to the Emsian (Breuer & Steemans Reference Breuer and Steemans2013; Cascales-Miñana et al. Reference Cascales-Miñana, Gerrienne, Moreno-Domínguez, Xue, Valenzuela-Ríos, Diez, Rial and Steemans2016; Breuer, pers. comm. 2016), is consistent with a middle–late Pragian age for the Posongchong Formation (Fig. 5).

Figure 5 Stratigraphic ranges of key spore taxa encountered in this study for dating the Posongchong Formation. Grey boxes indicate the assigned temporal interval. Absolute ages according to the International Chronostratigraphic Chart (v2016/12). Abbreviations: PRA = Pragian. *Camarozonotriletes parvus (sensu Steemans, Reference Steemans1989).
The Posongchong spore assemblage is very similar to that of the PE/PoW (Steemans Reference Steemans1981, Reference Steemans1989; Streel et al. Reference Streel, Fairon-Demaret, Otazo-Bozo and Steemans1981, Reference Streel, Higgs, Loboziak, Riegel and Steemans1987; Richardson & McGregor Reference Richardson and McGregor1986; see also Wellman Reference Wellman2006 and references therein). The Posongchong assemblage is dominated by crassitate apiculate spores, laevigate retusoid spores (Retusotriletes), with a noticeable presence of apiculate forms (Apiculiretusispora) and with biform sculpture (Dibolisporites) (Table 3). Some species of the PE/PoW spore assemblage from Euramerica, such as Verrucosisporites polygonalis, Dictyotriletes emsiensis and Dibolisporites wetteldorfensis, are present in South China from Xujiachong and Posongchong assemblages (Wellman et al. Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012; Table 2; supplementary material). A close spore comparison with the previous study from the Xuajiachong Formation shows that the main difference is in the spore dominance: retusoid spores in all of the levels in the Xujiachong Formation (Wellman et al. Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012) and crassitate apiculate spores for this study. Morphotypes such as Dictyotriletes and Brochotriletes present in Wang's (Reference Wang1994) study have not been found in the assemblage, and the diversity of Emphanisporites in the Posongchong Formation is limited to Emphanisporites rotatus only (Table 2). This fact supports the observations of Wellman et al. (Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012) from the Xujiachong Formation, who showed that (i) Dictyotriletes, Brochotriletes and Emphanisporites are rare in South China, whereas they are common in Euramerica; (ii) other common taxa in Euramerica (e.g., Clivosispora and Breconisporites) are absent from the Posongchong spore assemblages; and (iii) cryptospore dyads that are often observed in Euramerica assemblages have also not been found in the Posongchong assemblages.
In the Wenshan area, the Posongchong Formation is overlain by the Pojiao and Bajiaoqing formations (Jin et al. Reference Jin, Shen, Chen, Lu, Wei, Wang and Xie2005; Hao & Xue Reference Hao and Xue2013, table 3.1). Updated studies of the faunas from the marine Pojiao and Bajiaoqing formations (Lu & Chen Reference Lu and Chen2016; Lu et al. Reference Lu, Qie and Chen2016) provide further constraints for dating the Posongchong spores. Lu & Chen (Reference Lu and Chen2016) re-examined the conodonts obtained from the Bajiaoqing Formation and concluded that the previously reported Polygnathus dehiscens abyssus and Polygnathus dehiscens dehiscens from the base of the formation should rather be identified as Polygnathus excavatus. This means that the Bajiaoqing Formation may be correlated with the Polygnathus excavatus Zone (i.e., the second Emsian conodont zone). The level of the boundary of the first Emsian conodont zone, the Polygnathus kitabicus Zone, remains unknown. The Pojiao Formation is characterised by the Acrospirifer tonkinensis, now called Rostrospirifer tonkinensis brachiopod fauna (Jin et al. Reference Jin, Shen, Chen, Lu, Wei, Wang and Xie2005). This fauna has also been found from the lower part of the Yujiang Formation at the Liujing section of Guangxi, China (Wang & Rong Reference Wang and Rong1986), where the Pragian–Emsian boundary most probably is in the lower part of the Yujiang Formation (Shizhou Member; Lu et al. Reference Lu, Qie and Chen2016). This means that, at Liujing, the Rostrospirifer tonkinensis fauna probably ranges from the latest Pragian to early Emsian. In northern Vietnam, similar brachiopods, called Euryspirifer tonkiensis (=Rostrospirifer tonkinensis) fauna, have also been reported from the Pragian Mia Le Formation (Racheboeuf & Tong-Dzuy Reference Racheboeuf and Tong-Dzuy2000). Thus, faunal evidence suggests that the Pojiao Formation ranges from the late (or latest) Pragian to earliest Emsian in age. This scenario indicates that the Posongchong spore assemblages are, therefore, older than Emsian, which reinforces a Pragian assignation for the Posongchong Formation.
6. Palaeobotanical implications
A comparison of the most representative Early Devonian floras is shown in Table 4, including the Posongchong, Xujiachong and Pingyipu floras from South China. With ca. 33 species, the Posongchong flora is the most diverse Early Devonian flora from the fossil record (Table 4). However, only three Posongchong species (Guangnania cuneata, Hedeia sinica and Zosterophyllum australianum) are shared with the Xujiachong Formation (Table 4), and a single species (Zosterophyllum australianum) together with Zosterophyllum sp. are shared with the upper Baragwanathia flora from Victoria, Australia (Table 4). Indeed, the Posongchong flora does not share any species with other coeval floras from the Old Red Sandstone Continent, nor with any other floras from Laurussia, which shows a high level of endemism of the South China fossil floras during the Early Devonian. The absence of paratracheophytes (plants with S-type water-conducting cells, formerly called Rhyniaceae; Gerrienne et al. Reference Gerrienne, Dilcher, Bergamaschi, Milagres, Pereira and Rodrigues2006) and the diversity of euphyllophytes from the Posongchong and Xujiachong floras are also noticeable (Wellman et al. Reference Wellman, Zhu, Marshall, Wang, Berry and Xu2012; Table 4). These data are quite difficult to reconcile with the evidence supplied by the dispersed spore fossil record. The different palynoflora comparators contain similar spore morphotypes, especially the PE palynofloras (i.e., the coeval Posongchong, Xujiachong, Anglo-Welsh and Rhynie spore assemblages), independently of the known plant diversity. Furthermore, the Xujiachong and Posongchong palynofloras are dominated by Retusotriletes spp. and Apiculiretusispora spp. or Aneurospora spp. respectively, which also occur in Euramerican assemblages. It has to be noted that some spore species are only documented from the Lower Devonian of South China, as in the case of Aneurospora conica, Aneurospora posongchongensis, Aneurospora xujiachongensis or Leiozonospora xichongensis. This fact also suggests an isolated palaeogeographic position of the South China Plate during the Early Devonian, as do the plant megafossil data. Likewise, differences of species-level abundance between taxa would also support this view (e.g., the poor presence of Camarozonotriletes, Emphanisporites or Verrucosisporites in South China compared to their high abundance in Euramerica).
Table 4 Comparison of the main Early Devonian floras from northeastern Gondwana and Laurussia. Modified from Hao & Xue (Reference Hao and Xue2013, table 5.2).
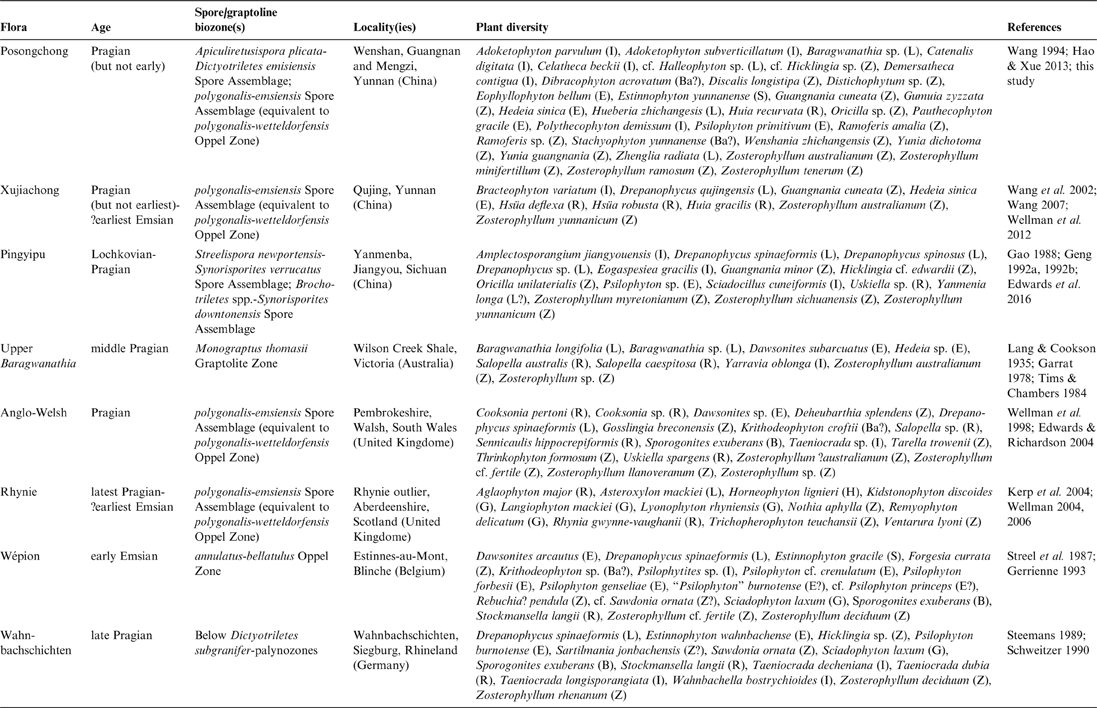
Notes: The capital letter in brackets indicates taxonomic or morphological groups as most appropriate. B: Bryophyte; Ba: Barinophyte; E: Euphyllophyte; G: Gameotophyte; H: Horneophytopsid; I: Incertae sedis; L: Lycopsid; R: Rhyniopsid and related plants; S: Sphenopsid; Z: Zosterophyllopsid. Northeastern Gondwanan floras: Posongchong, Xujiachong, Pingyipu, Upper Baragwanathia. Laurussian floras: Anglo-Welsh, Rhynie, Wépion, Wahnbachschichten.
7. Acknowledgements
We thank Charles Wellman and Deming Wang for their valuable comments on the earlier version of this manuscript. BCM thanks the support provided by a Marie Curie COFUND Postdoctoral Fellowship, University of Liege (grant no. 600405). JZX and PH are supported by the National Natural Science Foundation of China (no. 41722201) and the Yunnan Key Laboratory for Palaeobiology, Yunnan University (no. 2015DG007-KF04). PG and PS are FRS–FNRS Senior Research Associates.
8. Supplementary material
Supplementary material is available online at https://doi.org/10.1017/S1755691018000233.













