Isolated lower jaws are ones of the most common elements remaining from the Devonian tetrapodomorphs. This is probably due to their especially rigid construction in comparison to other skeletal parts. Findings presenting comparable mandibular elements are useful for morphological research and comparison. For this reason, the study of lower jaw transformations within the fish–tetrapod transition deserves special attention (e.g., Ahlberg & Clack Reference Ahlberg and Clack1998).
Each piece of newly obtained data becomes valuable for unearthing the earliest history of the fish–tetrapod transition. In 2005, an amateur palaeontologist Mikhail Yu. Kuleshov (Zoological Institute of the Russian Academy of Sciences, St Petersburg) found a lower jaw fragment in the Yam-Tesovo locality (Leningrad Region, northwestern Russia). He kindly donated this specimen to the A. A. Borissiak Palaeontological Institute of the Russian Academy of Sciences (PIN RAS) in Moscow. M. Yu. Kuleshov's specimen led to the revision of previously collected materials from this site. In 2015, Dr Aleksander O. Ivanov attracted our attention to a tetrapodomorph lower jaw specimen from the same locality in the L. A. Istratova's collection stored in the F. N. Chernyshev Central Scientific and Research Geological Survey Museum (TsNIGR Museum) in St Petersburg.
All specimens described herein originate from the Yam-Tesovo locality. It was discovered in 1928 by B. P. Asatkin and L. A. Istratova working in Geolkom (Geological Survey), Leningrad (now the A. P. Karpinsky Geological Research Institute (VSEGEI), St Petersburg), at that time. In 1929, R. F. Hecker and D. V. Obruchev (at that time both working in Geolkom), and D. V. Obruchev in 1959 (working at that time in PIN AN SSSR, Moscow), collected extensive materials there. Later, this locality was visited by O. A. Lebedev (PIN AN SSSR) in 1982, by A. O. Ivanov (Leningrad University) in 1984 and by these authors, jointly, in 2008. The authors of the present paper made a cooperative effort in the search for new material on Devonian tetrapodomorphs in the NW of Russia within the framework of the Joint French–Russian Centre National de la Recherche Scientifique – Russian Foundation for Basic Research (CNRS–RFBR) project, ‘Origin, ecology and environments of the earliest tetrapods from the Upper Devonian (Frasnian) and palaeogeography in the eastern Old Red Sandstone continent', in 2009–2011. The jaw collected by M. Yu. Kuleshov and the vomer collected by D. V. Obruchev in 1959 were figured, but not described by Ivanov & Lebedev (Reference Ivanov and Lebedev2011) in the guidebook for the field trip of the II International Obruchev Symposium, which took place in the Leningrad Region of Russia in 2011. In 2015, V. N. Glinskiy (from St Petersburg University) undertook another excavation in Yam-Tesovo that yielded a new tetrapodomorph skull bone (postfrontal), which he kindly presented to PIN RAS.
The mandibular elements described in the following section demonstrate a general ‘elpistostegalian' morphological pattern, together with early tetrapod characters. Upon similarity in their dermal ornamentation, these skeletal parts are suggested to belong to the same tetrapodomorph taxon for which we erect a new genus and species Rubrognathus kuleshovi n. gen. et sp. It is questionably referred here to ‘Elpistostegalia'. The postfrontal demonstrates peculiar ‘tetrapod-like' pits-and-ridges sculpturing known also in the presumed elpistostegalian Parapanderichthys stolbovi (Vorobyeva, 1960) (Vorobyeva Reference Vorobyeva1962, Reference Vorobyeva1992, Reference Vorobyeva, Novitskaya and Afanassieva2004), but it cannot be taxonomically assigned with certainty to any mandible specimen that bears different dermal ornamentation, nor with the vomer being incomparable to it. Determination of the taxonomic position of the postfrontal is thus limited to Tetrapodomorpha. The vomer is doubtfully related to the tetrapodomorph genus Livoniana Ahlberg et al., Reference Ahlberg, Lukševičs and Mark-Kurik2000 upon characteristic multiple tooth rows. Despite incomplete preservation of the material described herein, it provides important information on the morphology, phylogeny and diversity of the early tetrapodomorphs.
1. Geographical and geological setting
The Devonian vertebrate-bearing locality of Yam-Tesovo is situated on the left bank of the Oredezh River forming the Pristanskoye Lake water reserve downstream from the Yam-Tesovo village in the Luga District of the Leningrad Region (northwestern Russia) (Fig. 1).

Figure 1 Map showing the location of the Yam-Tesovo locality in the Leningrad Region, Russia, marked by a star.
The outcrop exposes the Yam-Tesovo Formation (Amata Regional Stage, ?lowermost Frasnian, Upper Devonian) of the regional stratigraphic scale of the East European Platform (Rzhonsnitskaya & Kulikova Reference Rzhonsnitskaya and Kulikova1990; Verbitsky & Vasilyeva Reference Verbitsky, Vasilyeva, Verbitsky, Verbitsky, Vasilyeva and Savanin2012). Deposits referred to the Amata Regional Stage crop out on a vast territory historically called the Main Devonian Field (MDF) in the NW of the East European Platform.
The Yam-Tesovo Formation overlies by unconformity the Lyuta Formation placed at the base of the Amata Regional Stage. The Yam-Tesovo deposits are red or brown, fine- and medium-grained cross-bedded sandstones, with intercalation of violet and greenish clays and clay pebbles. This formation is overlain by the marls of the Snetnaya Gora Formation referred to the Pļaviņas Regional Stage (Verbitsky & Vasilyeva Reference Verbitsky, Vasilyeva, Verbitsky, Verbitsky, Vasilyeva and Savanin2012).
These deposits formed in shallow-water nearshore conditions (Kuršs Reference Kuršs1992). Well-preserved vertebrate remains are found in the clayey beds. The list of taxa from Yam-Tesovo is only slightly shorter than that from the Amata Regional Stage on the whole (Ivanov & Lebedev Reference Ivanov and Lebedev2011) and, apart from the new forms described in the following section, includes: psammosteids: Placosteus undulatus Agassiz, 1844, Psammolepis venyukovi Obruchev, 1965, Psammosteus praecursor Obruchev, 1947, Ps. cf. Ps. cuneatus Obruchev, 1965, Ps. livonicus Obruchev, 1965 (Glinskiy Reference Glinskiy2013); acanthodians: Haplacanthus sp.; antiarchs: Asterolepis radiata Rohon, 1900, Bothriolepis obrutschewi Gross, Reference Gross1942; arthrodires: Plourdosteus livonicus (Eastman, 1896); porolepiform sarcopterygians: Glyptolepis baltica Gross, Reference Gross1930, Laccognathus panderi Gross, Reference Gross1941, Holoptychius cf. nobilissimus Agassiz, 1844; osteolepiform sarcopterygians: Eusthenopteron saevesoederberghi Jarvik, 1937, E. obruchevi Vorobyeva, Reference Vorobyeva1977; onychodontiform sarcopterygians: Onychodus sp.; actinistian sarcopterygians: ?Miguashaia sp.; elpistostegalians: Panderichthys rhombolepis (Gross, Reference Gross1930); and dipnoans: Dipterus sp. indet.
The position of this Regional Stage in the international chart of the Devonian is a matter of debate. These terrigenous sediments yield no conodonts, miospores are too scarce, and thus direct correlations to the standard conodont and miospore zonations are impossible. For this reason, the Givetian–Frasnian boundary is not clearly defined in the MDF and various authors had put forward different opinions on its position (Mark-Kurik Reference Mark-Kurik2000; Esin et al. Reference Esin, Ginter, Ivanov, Lebedev, Luksevics, Avkhimovich, Golubtsov and Petukhova2000; Ivanov et al. Reference Ivanov, Zhuravlev, Stinkulis, Evdokimova, Dronov, Sokiran, Shishlov, Broushkin and Myshkina2005; Ivanov & Lebedev Reference Ivanov and Lebedev2011; Lukševičs et al. Reference Lukševičs, Stinkulis, Ivanov and Tirzmale2018). Thus, the Amata Regional Stage and its vertebrate assemblage might be dated as uppermost Givetian or lowermost Frasnian, although the majority of the authors cited above supports the latter dating.
Bothriolepis obrutschewi Gross is an index fossil of the prima-obrutschewi Zone of the placoderm zonation in the MDF (Esin et al. Reference Esin, Ginter, Ivanov, Lebedev, Luksevics, Avkhimovich, Golubtsov and Petukhova2000). This makes possible a direct correlation of the Amata Regional Stage vertebrates to those in the Middle Timan in the NE of the East European Platform (Esin et al. Reference Esin, Ginter, Ivanov, Lebedev, Luksevics, Avkhimovich, Golubtsov and Petukhova2000; Lukševičs Reference Lukševičs2001; Ivanov & Lebedev Reference Ivanov and Lebedev2011). Mark-Kurik (Reference Mark-Kurik and Sorokin1981) correlated the Amata fish assemblages from Latvia, Estonia and Lithuania to those found in the Nairn Beds of Scotland. Schultze & Cloutier (Reference Schultze, Schultze and Cloutier1996) used the data on the Latvian locality Pasta Muiža of the same Amata age for faunistic comparative analysis with other regions of the world.
2. Materials and methods
Specimen PIN 1737/74, an incomplete left lower jaw with the missing anterior and posterior parts collected by M. Kuleshov in 2005, is designated here as the holotype of the new genus and species Rubrognathus kuleshovi sp. nov. Specimen TsNIGR Museum 3934/4 collected by L. A. Istratova in 1928 is a large but incomplete angular. According to the museum label, it had been originally identified as part of an ‘Asterolepis paired appendage'; this identification has not been revised since then. Here, this specimen is supposed to belong to Rubrognathus kuleshovi sp. nov., but an adult animal, by similar dermal sculpture and large rounded pores of the mandibular sensory canal. The postfrontal PIN 1737/86, described in Section 3.2 as Tetrapodomorpha indet., was discovered in 2015 by V. N. Glinskiy (Geological Faculty of the St Petersburg University) and presented to PIN RAS. Specimen PIN 1737/85 is an isolated vomer, somewhat rounded by abrasion. It is questionably assigned to the genus Livoniana. This piece was collected by D. V. Obruchev in Yam-Tesovo during his 1959 expedition and remained unidentified in the PIN RAS collection since that time.
All specimens had been found in loose sandstone and their light preparation was carried out manually by mounted needle. Specimens had been coated by ammonium chloride before photography.
Material is stored in the Borissiak PIN RAS, Moscow, Russia (coll. PIN 1737) and in the TsNIGR Museum, St Petersburg, coll. TsNIGR Museum 3934.
3. Systematic palaeontology
3.1. Subclass Tetrapodomorpha Ahlberg, 1991
Order Elpistostegalia Camp et Allison, 1961
Genus Rubrognathus Lebedev et Clément, gen. nov.
Derivation of name. Rubrum (Latin – red) after the characteristic red colour of the sand and enclosed vertebrate bones in the locality, and gnathus (Latin – jaw).
Type species. Rubrognathus kuleshovi sp. nov.
Diagnosis. Late Givetian–early Frasnian tetrapodomorph characterised by unusually shallow intercoronoid fossa, mostly formed by horizontal laminae of anterior and middle coronoids. Prearticular strongly convex in section, with its denticulated plate anteriorly reaching the anterior fang of the anterior coronoid. One or two rows of small teeth run along the dorsal margin of the prearticular with this denticulated area being separated from the ventral non-denticulated area by a well-developed ridge. Vertical coronoid laminae low. Coronoid fangs enter the coronoid tooth row. Middle coronoid fangs almost equal in size to anterior coronoid ones. Anterior part of lower jaw very slightly twisted. Mandibular sensory canal runs in open groove in the anterior half of mandible or enclosed in the bone and opens directly to the surface by large pores in its posterior half. Dermal ornament made of a network of small ridges forming tubercles in the nodes.
Rubrognathus kuleshovi n. gen. et sp.
Tetrapoda gen. et sp. nov.: Ivanov & Lebedev, Reference Ivanov and Lebedev2011, fig. 14i, j.
Derivation of name. After enthusiastic collector Mikhail Kuleshov (St Petersburg), who generously donated the specimen in 2008 to the A. A. Borissiak PIN RAS, Moscow, Russia.
Holotype. PIN 1737/74, an incomplete left mandible with the missing anterior and posterior parts.
Referred material. An incomplete right angular, TsNIGR Museum 3934/4.
Diagnosis. As for the genus.
Locality and horizon. An outcrop at the left bank of the Oredezh River by the western margin of the Yam-Tesovo village, Oredezh District, Leningrad Region, Russia; Yam-Tesovo Formation, Amata Regional Stage, ?Upper Givetian–Lower Frasnian, ?Middle–Upper Devonian.
Description. The symphysial part of the holotype PIN 1737/74 is damaged and missing. The posterior half of the jaw, posteriorly to the middle of the middle coronoid, is also broken off (Figs 2a, b, 3a, b). In comparison to the specimen TsNIGR Museum 3934/4 (Fig. 4a, b), the depth of the holotype specimen is approximately twice as small, suggesting that it probably belonged to a juvenile or subadult individual.

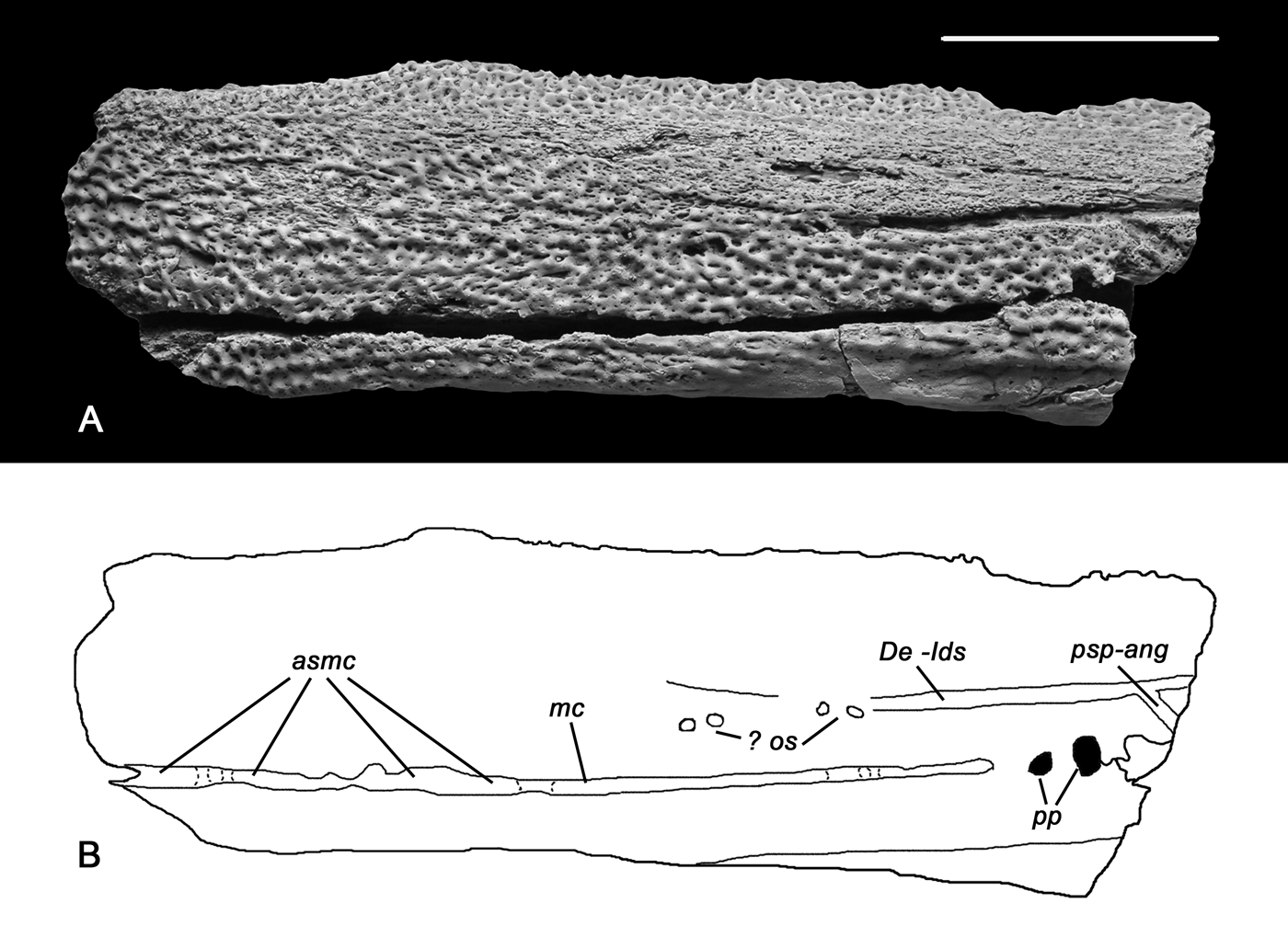
Figure 2 Rubrognathus kuleshovi sp. nov., holotype PIN 1737/74. (A, B) Photograph and specimen drawing of incomplete left ramus of the lower jaw in lateral view. Rostral end of the jaw to the left. Abbreviations: asmc=abraded segments of the mandibular sensory canal; De-Ids=dentary/infradentaries suture; mc = mandibular sensory canal; ? os=pits probably marking the obliterated sutural part; pp=large pores in the posterior part of the mandibular canal; psp-ang=postsplenial/splenial suture. Scale bar=1cm.

Figure 3 Rubrognathus kuleshovi sp. nov., holotype PIN 1737/74. (A, B) Photograph and specimen drawing of incomplete left ramus of the lower jaw in mesial view. Abbreviations: Co I = anterior coronoid; Co II = middle coronoid; De = dentary; Mc = Meckelian ossification; Pra = prearticular; Pra-Sp = sutural prearticular–splenial contact; Sp = splenial; amcCo = anteromesial corner of the horizontal lamina of anterior coronoid; covl = vertical lamina of the coronoids; ecs = external coronoid sulcus; icf = intercoronoid fossa; idf = infradentary flange; ldr = longitudinal dental row of teeth on the prearticular; lpf = lateral parasymphysial foramen; mcf = fenestra in the floor of the intercoronoid fossa showing Meckelian ossification; mf = Meckelian foramina; mgh = m. geniohyoideus insertion; mpf = mesial parasymphysial foramen; pcf = precoronoid fossa. Scale bar = 1cm.

Figure 4 Rubrognathus kuleshovi sp. nov., specimen TsNIGR Museum 3934/4, incomplete angular. (A) Lateral view. (B) Mesial view. Abbreviations: mc=mandibular sensory canal; oc=ossification centre; sac=surface overlapped by the surangular. Scale bar=1cm.
The anterior part of the mandible is only slightly twisted counter-clockwise in respect to the rest of the jaw in contrast to strong twisting known in the ‘elpistostegalians' Panderichthys rhombolepis (Vorobyeva & Schultze Reference Vorobyeva, Schultze, Schultze and Trueb1991), Elpistostege watsoni (Schultze &Arsenault Reference Schultze and Arsenault1985; Schultze Reference Schultze, Cloutier, Schultze and Cloutier1996) and Tiktaalik roseae (Daeschler et al. Reference Daeschler, Shubin and Jenkins2006).
A network of small ridges forming tubercles at the nodes ornaments the external face of the lower jaw (Fig. 2a). Pit bottoms between ridges bear variously sized, but generally small, vascular pores. Along the dorsal margin of the dentary, the ornamentation is finer than that on the infradentaries. However, the ornamentation is similar on the dentary and infradentaries in the anteriormost part of the jaw. A wide area running above the dentary/infradentary suture, and tapering anteriorly, is devoid of ornamentation, which may be partially due to moderate abrasion.
A well expressed furrow marks the suture between the dentary and the infradentaries. In the rostral direction, it runs only as far as to the level of the posterior half of the precoronoid (Fig. 2a, b, De-Ids) and does not suggest a kinetic zone described in Obruchevichthys, Elginerpeton (Ahlberg et al. Reference Ahlberg, Lukševičs and Mark-Kurik2000) and Tiktaalik (Daeschler et al. Reference Daeschler, Shubin and Jenkins2006). The course of the suture might be more anteriorly deduced by several pits probably marking the obliterated sutural part (? os). It is possible that the suture is widened due to its combination with the horizontal part of the mandibular pit-line, as discussed by Borgen & Nakrem (Reference Borgen and Nakrem2017). The partly preserved postsplenial/angular suture groove (psp-ang) is placed at the broken end of the jaw fragment.
The mandibular sensory canal (Fig. 2a, b, mc) occupied a deep furrow running almost all along the preserved fragment. Opposite to the middle coronoid it becomes enclosed in the bone and opens externally by two large pores (pp). It is possible that some of the bridges between openings have been destroyed during the original cleaning of the specimen (asmc), but examination at high magnification reveals large intact parts of the groove edges, meaning that a large part of the canal was originally open. The diameter of pores close to the posterior broken margin is larger than the width of the canal groove anteriorly, suggesting a more or less intact state of preservation of the latter.
In mesial view (Fig. 3a, b), the depth of the infradentary flange (idf) decreases posteriorly to a point opposite to the anterior coronoid fang. The mesial lamina of the postsplenial is only slightly developed.
The most prominent feature of the mesial side of the lower jaw is its long and large precoronoid fossa (Fig. 3a, b, pcf) which is almost twice longer than wide and deep. Its floor is formed by the Meckelian ossification (Mc) and its lateral rim is formed by the long and thin anterior process of the anterior coronoid (Co I ) and the mesial process of the dentary (De). The mesial portion of the splenial forms a deep and robust ridge bounding the precoronoid fossa mesially (Sp). The anterior coronoid contacts the posteriormost tip of the posteromesial process of the splenial (or the posterior part of the anterior process of the prearticular) by its anteromesial corner of the horizontal lamina (amcCo), thus forming the posterior wall of the precoronoid fossa. Denticulation of the vertical coronoid lamina extends anteriorly as far as the middle of the precoronoid fossa. The following narrow anterior process is toothless. The insertion scar for the m. geniohyoideus is placed at the dorsal surface of the anteromesial process of the splenial and is small and obliquely elongated (mgh).
The vertical laminae of both preserved coronoids (Fig. 3a, b, covl) are very low; the anterior and posterior processes of the anterior coronoid do not project dorsally much above the external coronoid sulcus (ecs), and slightly rise above it only opposite to the fang pairs. The tooth row of the vertical laminae of the coronoids interrupts opposite to the coronoid fang pairs, giving space for fangs. The sockets of the coronoid fangs suggest that these teeth on the anterior and middle coronoids had been of almost equal size, like in Panderichthys rhombolepis (PIN 3547/18), but in contrast to Tiktaalik roseae (Daeschler et al. Reference Daeschler, Shubin and Jenkins2006) in which the anterior ones are much larger than the posterior.
The floor of the shallow intercoronoid fossa (Fig. 3a, b, icf) is almost completely closed by the horizontal laminae of the anterior coronoid and the anterior part of the middle coronoid. The fenestra demonstrating the ossified portion of the Meckelian cartilage (mcf) is a small, elongated, oblique dumb-bell-shaped fissure exposing the passage of the canal possibly transmitting a branch of n. mandibularis V, a. mandibularis externa and v. mandibularis superior.
A small lateral parasymphysial foramen (Fig. 3a, b, lpf; sensu Ahlberg Reference Ahlberg1995) is placed in the external coronoid sulcus, anteriorly to the anterior process of the first coronoid. An also small mesial parasymphysial foramen (mpf) is found ventromesially from the anterior process of the anterior coronoid opposite to the lateral parasymphysial foramen. According to Vorobyeva (Reference Vorobyeva1977), these foramina transmitted a branch of n. mandibularis, as well as a sinus of v. mandibularis superior. This sinus is also observed in Obruchevichthys, Lamprotolepis (Vorobyeva Reference Vorobyeva1977; Clément & Lebedev Reference Clément and Lebedev2014), Livoniana (Ahlberg et al. Reference Ahlberg, Lukševičs and Mark-Kurik2000), Ichthyostega, Acanthostega, Metaxygnathus, Ventastega and Elginerpeton (Ahlberg & Clack Reference Ahlberg and Clack1998).
The dorsal margin of the prearticular (Fig. 3a, b, Pra) is straight. The acute anterior tip of its denticulated plate does not elevate above the dorsal surface of the bone and reaches the level of the anterior fang of the anterior coronoid. In Tiktaalik roseae (Daeschler et al. Reference Daeschler, Shubin and Jenkins2006) and Panderichthys rhombolepis (Vorobyeva Reference Vorobyeva1962) the denticulated area only reaches the anterior margin of the intercoronoid fossa. The prearticular denticulated plate bears a longitudinal row of small but well-marked separate teeth on its narrow dorsal surface (ldr). This row bifurcates posteriorly at the level of the anterior intercoronoid fossa. The diameter of the prearticular tooth sockets suggests that their size was only twice smaller than that of the dentary marginal teeth. Mesially to the main tooth row, a very narrow area bears few very small denticle sockets. Ventrally, the prearticular presents a reduced shagreen area and then no ornamentation. The dorsal and mesial planes of the prearticular form a right angle. This condition contrasts with that known in osteolepiforms (Borgen & Nakrem Reference Borgen and Nakrem2017), in which the prearticular plate is more or less vertical and flat, but agrees with that in Panderichthys. The suture between the anterior end of the prearticular and the posterior process of the splenial is hardly visible and marked only as an area of zigzag striations (Pra-Sp).
The width of the Meckelian bone exposure in the middle of the jaw fragment approximately equals that of the prearticular. The Meckelian cartilage (Fig. 3a, b, Mc) is partly ossified in this area to form the walls of two preserved Meckelian foramina (mf). The small anterior foramen is placed far rostrally, at the level of the first fang of the anterior coronoid fang pair; the posterior larger foramen is located at the level of the posterior process of the anterior coronoid. A wide groove, probably marking a vein passage, runs by the mesial side of the ventral flange of the postsplenial from the anterior Meckelian foramen towards the posterior one.
Marginal teeth of the dentary were closely spaced as shown by their ovoid, rostrocaudally compressed tooth sockets. No accessory lateral dentition is observed on the dentary in contrast to Panderichthys and Elginerpeton (Ahlberg & Clack Reference Ahlberg and Clack1998). The mesial margin of the dentary tooth row is a ridge forming the lateral wall of a deep and narrow external coronoid sulcus (ecs).
The specimen TsNIGR Museum 3934/4 is the anterior part of a right angular (Fig. 4a, b). This element is proportionally about twice as large as the holotype lower jaw. It is considered here as belonging to an adult individual. All margins of the specimens, except the dorsal one showing a large overlapped area for the surangular (Fig. 4a, sac), are more or less damaged, especially the rostral and caudal ones. This prevents reconstruction of the bone proportions and its contacts with the postsplenial and dentary. The preserved fragment is a middle part of the bone; it clearly demonstrates the ossification centre on the mesial side of the specimen (Fig. 4b, oc). The most conspicuous feature seen on the lateral surface (Fig. 4a) is an oblique anteroventrally running mandibular sensory canal (mc) that opens to the surface with very large rounded and ovoid pores fusing in the area of the ossification centre of the bone into elongated openings. Similarly large pores are present in the posterior part of the holotype. This pattern of pores arrangement and size is observed in the tetrapods Acanthostega (Ahlberg & Clack Reference Ahlberg and Clack1998), Ymeria (Clack et al. Reference Clack, Ahlberg, Blom and Finney2012) and Ventastega (Ahlberg et al. Reference Ahlberg, Lukševičs and Lebedev1994), but is different from what is known in Panderichthys rhombolepis in which the pores are slit-like and irregularly curved (Gross Reference Gross1941).
The dermal ornamentation and pattern of the sensory canal are very similar to those of the Rubrognathus holotype and, despite significant size difference, we assign this specimen to Rubrognathus kuleshovi n. gen. et n. sp.
3.2. Tetrapodomorpha indet
Material. An isolated postfrontal PIN 1737/86 from the Yam-Tesovo locality.
Description. This postfrontal PIN 1737/86 is not related to Rubrognathus kuleshovi n. gen. et sp., differing from it by its pits-and-ridges ornamentation well expressed in the area mesially to the orbital margin (Fig. 5a), unless findings of more complete and associated material prove the opposite. The area adjoining the orbital margin forms a thick supraorbital swelling (‘brow ridge') (sos). The rest of the external bone surface shows pitted sculpturing expressed to a smaller degree, well-marked tuberculation or no ornamentation, especially along the parietal contact and in the depressed caudal area. This absence of regular sculpturing is comparable to the ornamentation known in the interorbital depression of the ‘elpistostegalians' Elpistostege (Schultze Reference Schultze, Cloutier, Schultze and Cloutier1996 and pers. obs. of the MHNM 538 cast in PIN RAS) and Tiktaalik (Daeschler et al. Reference Daeschler, Shubin and Jenkins2006). The pitted type of sculpturing observed on this isolated postfrontal is shared by the presumed ‘elpistostegalian' Parapanderichthys (Vorobyeva Reference Vorobyeva1962, Reference Vorobyeva, Novitskaya and Afanassieva2004) and by Devonian and Carboniferous tetrapods (e.g., Ahlberg et al. Reference Ahlberg, Lukševičs and Lebedev1994; Clack Reference Clack2002).

Figure 5 Tetrapodomorpha indet., specimen PIN 1737/86, left postfrontal. (A, B) Dorsal view. (C, D) Lateral view (rostral margin to the left). (E, F) Mesial view (rostral margin to the right). (G, H) Ventral view. Abbreviations: ?apsos=incompletely obliterated suture presumably between the anterior and posterior anlagen of supraorbital elements; ?aso-psoc=presumed contact of the anterior supraorbital element over the processes of the posterior supraorbital element bearing vascular grooves; on=orbital notch; pocb=postorbital commissure branching from the supraorbital canal; pocco=lateral opening of postorbital commissure canal; pocs=contact surface for the postorbital; ?prfc=depression which possibly housed the overlapped process of the prefrontal; rosoc=rostral opening of the supraorbital canal; soc=supraorbital canal; soco=posterior opening of the supraorbital canal; sos=supraorbital swelling. Scale bars=1cm.
The specimen is abraded rostrally, caudally and very slightly mesially. Its contact surfaces with the prefrontal and frontal are not clear, but the posterior edge of the contact for the frontal may be cautiously reconstructed on the basis of the rostral opening of the supraorbital canal (Fig. 5g, h, rosoc). The remaining part of the mesial bone margin (Fig. 5e, f) is a contact for the parietal. It consists of a thick rugose dorsal area and a thin lamellar edge, slightly damaged at the base. The contact for the parietal probably occurred only by this thin lamellar edge as far as the thick dorsal part shows no sutural structure and bears foramina of the supraorbital sensory canal running along the margin and enclosed in the bone.
A well-preserved lateral margin (Fig. 5c, d) demonstrates the orbital notch (on) and, caudally from it, a distinct contact surface for the postorbital (pocs) separated in two parts: a rostral one facing ventrolaterally, and a caudal one facing posterolaterally. These surfaces are subdivided by a projection wedging into the postorbital like in Tiktaalik (Daeschler et al. Reference Daeschler, Shubin and Jenkins2006).
A few sensory canal pores (Fig. 5a, b) mark the postorbital commissure branching from the supraorbital canal (pocb). Further, the postorbital commissure gives no superficial branches. Rostromesially, a row of supraorbital canal pores runs close to the mesial margin of the bone (Fig. 5e, f). The canal itself may be observed on a short distance in the mid-length of the bone; it is exposed here by the bone breakage (soc). Further rostrally, its route may be traced by a row of pores lined along and above the postfrontal–frontal contact. The rostral opening of the canal (rosoc) is present on the ventral edge of this contact due to post-mortem wearing (Fig. 5g, h). The postorbital commissure canal (pocco) opens at the base of the obtuse projection on the contact surface for the postorbital (Fig. 5d, h). The posterior opening of the supraorbital canal (soco) is observed on the caudal, abraded end of the bone.
The ventral surface of the postfrontal (Fig. 5g, h) is subdivided in two areas: a mesial one roofing the interorbital division and a lateral one forming the dorsal margin of the orbit. The former is S-shaped and smooth. Rostrally, between the two areas is a deep depression which possibly housed the overlapped process of the prefrontal (?prfc). Walls of this depression bear grooves marking the course of branching blood vessels. In the mid-length of the lateral area the orbital margin is interrupted by a short oblique slit, which is interpreted here as an incompletely obliterated suture between the anterior and posterior margins of supraorbital elements anlagen (?apsos). Connection of these anterior and posterior elements may be also traced in the roof of the depression described above as an overlap of the anterior supraorbital element over thin projecting processes of the posterior element bearing vascular grooves (?aso-psoc). No sutures are marked on the external surface of the bone. This twofold structure of the postfrontal might be a trace of an ancestral two- or even threefold state and further incomplete fusion of the supraorbital series of bones (Jarvik Reference Jarvik1944; Borgen & Nakrem Reference Borgen and Nakrem2017).
3.3. ?Livoniana sp
Material. An isolated specimen PIN 1737/85 identified here as a right vomer from the Yam-Tesovo locality. Figured but not described by Ivanov & Lebedev Reference Ivanov and Lebedev2011, fig. 14k (Tetrapodomorpha gen. et sp. nov.).
Description. The specimen PIN 1737/85 is distinguished by its multiple tooth rows (of which only sockets are preserved) on its occlusal surface (Fig. 6a–f). In this respect, it is comparable to the tetrapod-like Livoniana multidentata Ahlberg et al., Reference Ahlberg, Lukševičs and Mark-Kurik2000, known only by the anterior fragments of lower jaws.

Figure 6 ?Livoniana sp. specimen PIN 1737/85, right vomer. (A, B) Ventral (occlusal) view. (C, D) Anterolateral (labial) view. (E, F) Dorsal view. Abbreviations: avlm = anteroventrolateral (labial) margin; csPmx = contact surface for the premaxilla; fts = fang-sized tooth sockets; oDpl = presumable surface overlapped by the process of the dermopalatine; ollDsph = area overlapped by the lateral lobe of the rostral part of parasphenoid; rp = replacement pits. Scale bars = 1cm.
The specimen is slightly rounded by abrasion and is probably incomplete mesially. As in Livoniana multidentata, the new specimen demonstrates up to six rows of variously sized teeth (Fig. 6a, b). According to the size of the sockets, the teeth seem to have been proportionally smaller in PIN 1737/85 than in Livoniana. For instance, the sockets tend to diminish labially to microscopic size in contrast to the Livoniana dentary dentition in which even the smallest marginal teeth are rather large. In Livoniana, the tooth rows run parallel to the jaw margin. In contrast to that, the vomer PIN 1737/85 shows two larger fang pairs of tooth sockets (fts) and replacement pits (rp) at the posteromesial (lingual) margin anterolaterally. The fangs and replacement pits were semi-encircled by smaller teeth. Each of the two ‘fang' sockets is accompanied by a smaller one entering the semicircular row. Labially to the semicircular rows, there is only one row running all along the vomer length and composed of medium-sized teeth. A patch of tiny sockets is placed labially in the mid-length of the specimen. A small area devoid of teeth is situated in the posteromesial corner of the denticulated plate, and another one in its posterolateral corner.
The labial margin is the deepest and tapers lingually (Fig. 6c, d). The rounded anteroventrolateral (labial) margin (avlm) is swollen. It slightly protrudes and limits the posterior margin of the presumably slit-like apical fossa. A well-marked contact surface for the premaxilla (csPmx) is present posterolaterally. This contact surface is wide and low, but only slightly overhangs the anterolateral bone margin, in contrast to rhipidistians in which this well expressed process forms the posterolateral margin of a large apical fossa.
The dorsal surface of the bone is separated into two contact surfaces by a prominent ridge (Fig. 6e, f). A narrow area placed between the ridge and the labial margin of the bone becomes slightly deeper and wider posteriorly. This area is considered as the overlap area for the dermopalatine bone (oDpl). The wider mesial area bears radial grooves and is the overlap area for the rostral part of the parasphenoid (ollDsph).
No trace of the choana margin is observed. This is probably due to the intense post-mortem wearing. It can also be assumed that the contribution of the vomer to the choana margin was very reduced, as known in Tulerpeton curtum in which the anterior edge of the choana forms only a small vertical area on the posterolateral corner of the vomer (Lebedev & Clack Reference Lebedev and Clack1993).
4. Discussion
4.1. Rubrognathus
Due to the patchy, rare and usually incomplete material of Late Devonian tetrapodomorphs, it is crucial to establish a set of cranial characters distinguishing ‘true tetrapods' from finned tetrapodomorphs. The outstanding preservations of Panderichthys, Tiktaalik, Ichthyostega and Acanthostega excepted, the key taxa of the tetrapodisation process are generally represented by isolated skull and pectoral girdle elements. One of the most frequently preserved skeletal elements is the lower jaw. This element, although incomplete, is important as it provides information on the dermal ornament, the nature of the sensory canal, the morphology of the snout reflected in the structure of the anterior part of the mandible, the character of dentition and other features. These characters are informative on the ecology of the Devonian tetrapodomorphs, including the mode of prey capture, sensory perception, skin physiology, breathing mode, etc., and can reflect innovative steps from aquatic to terrestrial living modes.
Ahlberg et al. (Reference Ahlberg, Lukševičs and Lebedev1994, Reference Ahlberg, Lukševičs and Mark-Kurik2000) and Ahlberg & Clack (Reference Ahlberg and Clack1998) presented evolution scenarios of the lower jaws during the so-called fish–tetrapod transition. For instance, they studied the different state characters of the mandibular sensory canal, dentition, structure of the ventral margin of the mandible and orientation of the glenoid facet of the articular. In their studies, special attention is paid to the establishment of a list of features unique to tetrapods.
Due to its incomplete preservation, Rubrognathus kuleshovi n. gen. et sp. presents only some of the characters that are shared with tetrapods: 1) the mandibular sensory canal running partly in an open groove and partly enclosed in the bone, with large rounded pores opening directly to the surface; 2) the reduction of fangs and their lateral shift in combination with the reduction of the vertical lamina of coronoids; and 3) the presence of lateral and mesial parasymphysial foramina.
While discussing the mandibular sensory line structure, Vorobyeva (Reference Vorobyeva1977) mentioned the presence of a ‘stegocephalian-like' position of the sensory mandibular canal in an open groove in Panderichthys and Lamprotolepis, but did not refer to any specimen numbers or describe or illustrate this statement. A restudy of the lower jaws of Panderichthys rhombolepis with additional preparation shows that the pores pattern of the sensory mandibular canal can be slit-like, vermiculate posteriorly (Gross Reference Gross1941, text-fig. 15a) in the P. rhombolepis specimens from the Ērmaņi locality in Latvia, or they can be rounded and lined in a row, as previously illustrated by Gross in the anterior part (Gross Reference Gross1941, text- fig. 19b) of jaw specimens from the Lode locality. However, a fragment of the middle part of the mandible of P. rhombolepis from Lode (collection PIN 3547/49) presents a variation in which these slit-like foramina are lined in a single row and form an almost completely opened groove, sometimes bridged by narrow connections, giving an impression of a completely opened sulcus in unprepared state (Fig. 7a, b).

Figure 7 Various patterns of the openings of the anterior section of the mandibular sensory canal. (A, B) Panderichthys rhombolepis (Gross, Reference Gross1930): (A) specimen PIN 3547/18; (B) specimen PIN 3547/49. (C) Obruchevichthys gracilis Vorobyeva, Reference Vorobyeva1977, holotype PIN 1491/51. (D) Rubrognathus kuleshovi gen. et sp. nov., holotype PIN 1737/74. (E) Acanthostega gunnari Jarvik, Reference Jarvik1952. (E) Redrawn from Ahlberg & Clack (Reference Ahlberg, Lukševičs and Lebedev1998). Not to scale.
The earlier character state is then recorded in the early ‘elpistostegalian' tetrapodomorph Panderichthys, showing possible morphological transition between the two patterns. In Obruchevichthys (Fig. 7c), the anterior part of the mandibular sensory canal enclosed into the bone opens by large pores or short sections of open grooves (Clément & Lebedev Reference Clément and Lebedev2014).
In Rubrognathus kuleshovi n. gen. et sp. holotype PIN 1737/74 (Fig. 7d), presumably existing bridges that became broken during initial preparation are fewer than in Panderichthys and Obruchevichthys. In the posteriormost part of the specimen, there are two rounded pores with a diameter larger than the width of the mandibular sensory groove. This suggests that in Rubrognathus the anterior part of the mandibular sensory canal is placed within the sulcus, whilst the posterior part opened through large rounded pores. The same is seen in the specimen TsNIGR Museum 3934/4. This character, regarded as a tetrapod marker, is now documented in the early tetrapodomorph Rubrognathus, suggesting an earlier appearance of this anatomical character than previously thought.
The passage of a sensory canal in an opened groove, or in a closed canal with large openings, cannot be solely used to attribute a specimen to limbed or finned tetrapods. The general case in the Devonian tetrapods is that the course of the canal on the postfrontal is not expressed superficially; this may result from the deep position of the canal itself and/or the small size of the pores opening to the surface, which are obscured by rough sculpturing. New finds of cranial remains might help to solve this issue.
As discussed by Ahlberg & Clack (Reference Ahlberg and Clack1998) and Ahlberg et al. (Reference Ahlberg, Lukševičs and Mark-Kurik2000), one of the most important changes during the ‘fish–tetrapod transition' is the size reduction of fangs and their shift into the lateral row of denticles on the vertical lamina of the coronoids. The first steps of this evolution can be observed in Tiktaalik (Daeschler et al. Reference Daeschler, Shubin and Jenkins2006) and Rubrognathus (Fig. 3). As noted by Ahlberg & Clack (Reference Ahlberg and Clack1998), the coronoid fangs retain their large size in the Devonian tetrapods Obruchevichthys, Ventastega and Metaxygnathus. The presence of large coronoid fangs and their incorporation into the vertical coronoid lamina tooth row, in combination with the reduction of the vertical lamina itself, is an intermediate state between the condition in tetrapods sensu stricto and the condition known in Panderichthys (Vorobyeva Reference Vorobyeva1962, Reference Vorobyeva, Novitskaya and Afanassieva2004) and many other rhipidistians (Borgen & Nakrem Reference Borgen and Nakrem2017).
The presence of mesial and lateral parasymphysial foramina is of doubtful evolutionary signal. These foramina transmitted branches of n. mandibularis, innervating the anterior part of the dental lamina, and the venous sinus of v. mandibularis superior (Vorobyeva Reference Vorobyeva1977). The enlargement of these foramina in the lower jaws of the Devonian tetrapods might reflect the increase of blood supply in the anterior part of the head, in comparison to osteolepiforms.
The phylogenetic or evolutionary signals of some features present in Rubrognathus are difficult to evaluate. The anterior end of the vertical lamina of the anterior coronoid is toothless in Rubrognathus (Fig. 3), thus matching the condition in Ichthyostega, but in contrast to other Devonian tetrapods (Ahlberg & Clack Reference Ahlberg and Clack1998). In the finned sarcopterygians this feature is variable. A toothed anterior coronoid process is observed in numerous ‘osteolepidids' and Lamprotolepis, but not in Megalichthys, tristichopterids and elpistostegalians (Vorobyeva Reference Vorobyeva1977; Borgen & Nakrem Reference Borgen and Nakrem2017); thus, it cannot be used to unequivocally define limbed tetrapods.
The dermal ornamentation in Rubrognathus (Fig. 8a, b) differs strongly from the ‘starburst' pattern reaching the margins of each separate bone in tetrapods (Ahlberg et al. Reference Ahlberg, Lukševičs and Lebedev1994, Reference Ahlberg, Lukševičs and Mark-Kurik2000; Ahlberg & Clack Reference Ahlberg and Clack1998), but is similar to that observed in Panderichthys rhombolepis (Fig. 8c) and Livoniana multidentata (Ahlberg et al. Reference Ahlberg, Lukševičs and Mark-Kurik2000).

Figure 8 Dermal ornamentation in various tetrapodomorphs. (A, B) Rubrognathus kuleshovi gen. et sp. nov.: (A) lower jaw holotype PIN 1737/74; (B) lower jaw specimen TsNIGR Museum 3934/4. (C) Panderichthys rhombolepis (Gross, Reference Gross1930), lower jaw specimen PIN 3547/49. (D) Tetrapodomorpha indet., postfrontal PIN 1737/86. (E, F) Parapanderichthys stolbovi (Vorobyeva, 1960), holotype PIN, 54/169, dorsal and ventral parts of the left premaxilla, respectively. Scale bars=2mm (A, B); 4mm (C–F).
4.2. Comments on the postfrontal bone
The pits-and-ridges dermal ornamentation of the bone (Figs 5a, 8d) is very similar to that known in early tetrapods, including the Devonian ones. This type of ornamentation is used as one of the features defining the tetrapods (Ahlberg et al. Reference Ahlberg, Lukševičs and Lebedev1994, Reference Ahlberg, Lukševičs and Mark-Kurik2000; Daeschler et al. Reference Daeschler, Shubin and Jenkins2006). However, the elpistostegalian fish Parapanderichthys stolbovi (Vorobyeva 1960, Reference Vorobyeva1962; Fig. 8e, f) also presents the same type of ornamentation on the snout and lower jaw. Rubrognathus kuleshovi n. gen. et sp. shows an ornamentation of the lower jaw different from that of the isolated postfrontal. Thus, the latter probably belongs to another taxon than Rubrognathus, but sculpturing on the skull roof and lower jaw may differ in the same animal. The presence of a supraorbital ridge is characteristic of early tetrapods (Clack Reference Clack2002), but is also present in ‘elpistostegalians' (Panderichthys rhombolepis: Vorobyeva & Schultze Reference Vorobyeva, Schultze, Schultze and Trueb1991; Elpistostege watsoni: Schultze Reference Schultze, Cloutier, Schultze and Cloutier1996; Tiktaalik roseae: Daeschler et al. Reference Daeschler, Shubin and Jenkins2006).
The postfrontal posterolaterally contacts the postorbital by a more or less short, straight suture (Fig. 9a) in all Devonian tetrapods in which the postfrontal is known: Ichthyostega (Jarvik Reference Jarvik1952, Reference Jarvik1996), Acanthostega (Clack Reference Clack2003), Ventastega (Ahlberg et al. Reference Ahlberg, Lukševičs and Lebedev1994) and ?Tulerpeton (Lebedev & Clack Reference Lebedev and Clack1993), except Jakubsonia, in which the bones in this area are fused and show no sutures (Lebedev Reference Lebedev2004). The same pattern is observed in Elpistostege (Schultze Reference Schultze, Cloutier, Schultze and Cloutier1996) and Panderichthys (Vorobyeva & Schultze Reference Vorobyeva, Schultze, Schultze and Trueb1991) (Fig. 9b). In contrast to this pattern, in Tiktaalik (Daeschler et al. Reference Daeschler, Shubin and Jenkins2006) the postfrontal presents a lateral projection forming a long suture with the postorbital (Fig. 9c). This character is also observed in the postfrontal PIN 1737/86 (Fig. 9d), although the projection is not so much expressed.

Figure 9 Shape and contacts of the postfrontals. (A) Acanthostega gunnari Jarvik, Reference Jarvik1952. (B) Panderichthys rhombolepis (Gross, Reference Gross1930). (C) Tiktaalik roseae Daeschler et al., Reference Daeschler, Shubin and Jenkins2006. (D) Tetrapodomorpha indet., postfrontal PIN 1737/86. (A) Redrawn from Clack (Reference Clack2003); (B) redrawn from Vorobyeva & Schultze (Reference Vorobyeva, Schultze, Schultze and Trueb1991); (C) redrawn from Daeschler et al. (Reference Daeschler, Shubin and Jenkins2006). Abbreviations: Fr=frontal; It=intertemporal; Pa=parietal; Po=postorbital; Pof=postfrontal; Prf=prefrontal. Not to scale.
4.3. Comments to ?Livoniana sp. vomer
In Devonian and Carboniferous osteolepiform fishes, the horizontal lamina of the vomer bears a fang pair while the vertical one is set by a row of denticles. Panderichthys rhombolepis follows this pattern (Vorobyeva & Schultze Reference Vorobyeva, Schultze, Schultze and Trueb1991) (Fig. 10a). Smaller teeth anteriorly from this tooth row are registered in Megalichthys hibberti and Ectosteorhachis (Schultze Reference Schultze1974) and in Megalichthys sp. (Borgen & Nakrem Reference Borgen and Nakrem2017). Multiple tooth rows on osteolepiform vomers are unknown.

Figure 10 Vomer structure and its relation to the adjacent bones in: (A) Panderichthys rhombolepis (Gross, Reference Gross1930); (B) ?Livoniana sp., PIN 1737/85; (C) Acanthostega gunnari Jarvik, Reference Jarvik1952; (D) Tulerpeton curtum Lebedev, 1984. (A) Redrawn from Vorobyeva & Schultze (Reference Vorobyeva, Schultze, Schultze and Trueb1991); (C) redrawn from Clack (Reference Clack2002); (D) redrawn from Lebedev & Clack (Reference Lebedev and Clack1993). Abbreviations: Pmx = premaxilla; Vo = vomer; af = apical fossa; ch = choana. Not to scale.
In those Devonian tetrapods in which vomers are recognised (Ichthyostega: Jarvik Reference Jarvik1952, 1996; Tulerpeton: Lebedev & Clack Reference Lebedev and Clack1993; Ventastega: Ahlberg et al. Reference Ahlberg, Lukševičs and Lebedev1994; and Acanthostega: Clack Reference Clack2002), the small fang pairs enter the lateral tooth row (Fig. 10c, d). This tooth row is set on the anterolateral (labial) margin of the vomer; the vertical lamina itself is not expressed. The horizontal lamina is, to some extent, ornamented by fields of small denticles mesially (e.g., in Tulerpeton: Lebedev & Clack Reference Lebedev and Clack1993).
In the specimen PIN 1737/85, described above as ?Livoniana sp., the distribution of teeth on the horizontal plate contrasts osteolepiform, Panderichthys and early tetrapod patterns (Fig. 10b). The vertical lamina is almost absent; the largest teeth on the horizontal lamina are placed mesially rather than laterally. This construction suggests that the lateral tooth rows in ?Livoniana sp. are homologous to the tooth row set upon the vertical lamina of the osteolepiforms and Panderichthys, but present a multiplicated pattern extending anterolaterally. This is different from the multiplicated pattern extending anteromesially observed in Tulerpeton (Fig. 10d).
A vomer with a flattened general morphology showing no clearly enlarged fang pairs and with tooth rows decreasing laterally in size has not been described before. The only analogue for this model is the anterior part of the dentary of the near-tetrapod Livoniana (Ahlberg et al. Reference Ahlberg, Lukševičs and Mark-Kurik2000). A dissimilarity is the disposition of the lingually placed dental elements, which form two arches (around the each fang) in ?Livoniana rather than parallel rows. This is explained by retention of the semicircular teeth arrangement on the vertical lamina of the vomer.
The anterolateral extension of the tooth rows in ?Livoniana suggests that the apical fossa should have been reduced and narrow, as exemplified by the anteromesial extension of the vomer in some tetrapods (e.g., Tulerpeton; Fig. 10d). At the jaw closure, the small anterior dentary tooth rows contacted the premaxillary dentition. In those osteolepiforms and elpistostegalians in which the anterior dentary teeth are enlarged, the dentary fangs entered the apical fossa. Teeth on the parasymphysial plate abutted the tooth row of the vertical lamina of the vomers. By analogy, in Livoniana-like fishes the smaller external rows of the dentary should have contacted the premaxillary teeth whilst the tips of enlarged teeth of the lingual row contacted vomerine surface labially from the tooth rows. In this case, the apical fossa should be reduced and very narrow.
4.4. Tetrapodomorphs in the Yam-Tesovo vertebrate assemblage
Some isolated tetrapodomorph remains collected in the Yam-Tesovo locality are difficult to assign with certainty. They show interesting derived features but cannot be related to each other. The dermal ornamentation of the postfrontal PIN 1737/86 is very different from that seen on the new taxon Rubrognathus kuleshovi n. gen. et sp. The mandible dentition of the latter is quite different from that seen in the vomer of ?Livoniana sp. PIN 1737/85. No taxonomic relation can be made between the postfrontal and the ?Livoniana vomer.
The situation becomes even more complicated taking into account Vorobyeva's (Reference Vorobyeva1962) descriptions of a lower jaw fragment (PIN 54/182) and of an isolated coronoid (PIN 54/201) assigned by her to Panderichthys rhombolepis. ‘Panderichthys'-type rhombic tuberculated scales had been also mentioned by this author from the Yam-Tesovo locality. However, the dermal ornamentation of the jaw fragment PIN 54/182 is quite different from that known in Panderichthys rhombolepis and the isolated coronoid does not give any further information on its taxonomic attribution. These problems may only be solved by new material hopefully found during future excavations, particularly at the Yam-Tesovo and other localities of the Amata age.
Vorobyeva & Lyarskaya (Reference Vorobyeva, Lyarskaya and Obruchev1968) described and figured (pl. 11, fig. 1) an incompletely preserved lower jaw from the Amata deposits of the Latvian locality Pasta Muiža, which they identified as Panderichthys sp. This jaw demonstrates some characters similar to those seen in the Rubrognathus kuleshovi n. gen. et sp. holotype specimen, such as strongly convex denticulated lamina of the prearticular, the dorsal margin of the prearticular separated from the ventral part by a well-developed ridge, low vertical coronoid laminae and coronoid fangs entering the coronoid tooth row. The specimen is described and photographed from the mesial side only, thus suggesting poor preservation of its lateral side, and it is hard to say whether this specimen might be related to the coeval R. kuleshovi n. gen. et sp. from Russia.
5. Conclusions
Lower jaw morphology is important in the early tetrapodomorph studies as it concentrates information on the phylogenetic relationships and anatomical characters that reflect physiology and adaptations to the environment, including hunting and capturing prey, sensory perception, skin physiology and possibly change of breathing mode during fish–tetrapod transition.
Rubrognathus kuleshovi n. gen. et sp., questionably referred to here as ‘Elpistostegalia', demonstrates a combination of characters seen in limbed tetrapods, ‘near-tetrapods' and ‘elpistostegalian' fishes at the earliest stage of the tetrapodisation process in vertebrates. The morphology of the sensory mandibular canal, the lateral shift of coronoid fangs and the presence of parasymphysial foramina relate Rubrognathus to tetrapods sensu stricto in modern definition of this group. However, the structure of the precoronoid fossa precludes its direct attribution to these animals.
The isolated postfrontal demonstrates peculiar ‘tetrapod-like' pits-and-ridges ornamentation. This cannot be related to the mandible parts of Rubrognathus kuleshovi n. gen. et sp., which bear different dermal sculpturing, or to the isolated vomer. Its taxonomic attribution is thus limited to Tetrapodomorpha.
The isolated vomer shows a unique pattern of multiple tooth rows and is doubtfully related to the tetrapodomorph genus ?Livoniana Ahlberg et al., Reference Ahlberg, Lukševičs and Mark-Kurik2000. Despite its incomplete preservation, material from the uppermost Givetian–lowermost Frasnian Yam-Tesovo locality provides important information on the morphology, phylogeny and diversity of early tetrapodomorphs. With the description of new ‘proto-' and ‘near-tetrapod' material during the last two decades, the mosaic nature of the appearance of tetrapod characters in various tetrapodomorph lineages grows increasingly evident and the boundary between ‘limbed' and ‘finned' tetrapodomorphs becomes significantly blurred. This suggests a much more complex evolutionary history of the fish–tetrapod transition than previously thought.
6. Acknowledgements
The authors are grateful to M. Yu. Kuleshov, St Petersburg, Russia, for his donation of the specimen to A. A. Borissiak PIN RAS, Moscow, Russia, and for providing information on the Yam-Tesovo locality. V. N. Glinskiy (St Petersburg University) joined the teams of the 2010–2011 field trips and symposium excursion, discovered a new skull bone in 2015 and kindly donated it to PIN RAS, and photographed the material in the TsNIGR Museum in St Petersburg. Alexander Ivanov (St Petersburg University) was one of the main coordinators of the Russian–Franco–Latvian expedition to the Leningrad Region. He attracted our attention to the Rubrognathus specimen in the TsNIGR Museum collection in St Petersburg. We are also pleased to thank the other 2009–2011 members of the expedition who helped us with excavations: G. V. Zakharenko, Yu. V. Shuvalova and E. E. Deitchman (PIN RAS, Moscow) and E. M. Kirilishina (Moscow University) from Russia; A. Blieck (University of Lille, France) and S. Olive (Natural History Museum, Brussels, Belgium); E. Lukševičs, V. Sturis (University of Latvia) and I. Zupiņš (Latvian Natural History Museum), from Riga, Latvia; and the participants of the field trip of the 2nd International Obruchev Symposium. We thank the reviewers E. Lukševičs (University of Latvia, Riga, Latvia) and P. Beznosov (Institute of Geology, Syktyvkar, Russia) for providing numerous corrections to the text and for offering useful advice, for which we are grateful. Photographs were taken by A. V. Mazin and S. V. Bagirov in the photo laboratory of PIN RAS. The research, including the expedition, was supported by the Joint French–Russian project of CNRS–RFBR ‘Programmes Internationaux de Coopération Scientifique (PICS)' and by the Russian Foundation for Basic Research no. 09-05-91059-CNRS entitled ‘Origin, ecology and environments of the earliest tetrapods from the Upper Devonian (Frasnian) and palaeogeography in the eastern Old Red Sandstone continent' and ‘Expedition Research' project of the Presidium of the Russian Academy of Sciences.