


INTRODUCTION
The fin whale Balaenoptera physalus (Linnaeus, 1758) is the second largest member of the Balaenopteridae family. It has a cosmopolitan distribution, and is found in all major oceans. In the Mediterranean Sea, it represents the only mysticete regularly spotted during summer, especially in the north-western sub-basin within the Marine Protected Area, the Pelagos Sanctuary (Forcada et al., Reference Forcada, Notarbartolo di Sciara and Fabbri1995; Gannier, Reference Gannier1997, Reference Gannier1998a). This area forms one of the main cetacean habitats in the Mediterranean and is described as a likely feeding ground for large numbers of species (Forcada et al., Reference Forcada, Notarbartolo di Sciara and Fabbri1995; Gannier, Reference Gannier1998a; Laran & Drouot-Dualau, Reference Laran and Drouot-Dulau2007; Moulins et al., Reference Moulins, Rosso, Nani and Würtz2007). Whales are known to be affected by a number of anthropogenic problems, including ship collisions and the yearly estimated average for their non-natural mortality was approximately 1.7 whales from 1972 to 2001 (Panigada et al., Reference Panigada, Pesante, Zanardelli, Capoulade, Gannier and Weinrich2006). Most studies dedicated to this species describe its distribution, its abundance and its behaviour in the north-western Mediterranean Sea (Forcada et al., Reference Forcada, Notarbartolo di Sciara and Fabbri1995; Gannier, Reference Gannier2002; Panigada et al., Reference Panigada, Pesante, Zanardelli, Capoulade, Gannier and Weinrich2006). The abundance estimates during summer range from 0.019 to 0.024 individuals per km2 (with a respective coefficient of variation of 13% or 27%) (Forcada et al., Reference Forcada, Aguilar, Hammond, Pastor and Aguilar1996; Gannier, Reference Gannier1997). Mean water depth in the western Mediterranean Sea where sightings occur is 2360 m (Forcada et al., Reference Forcada, Aguilar, Hammond, Pastor and Aguilar1996) justifying that the whale is generally considered a pelagic animal. Its presence in waters deeper than 2000 m is because of up-welling caused by the local permanent frontal structure (Forcada et al., Reference Forcada, Aguilar, Hammond, Pastor and Aguilar1996; Gannier, Reference Gannier1998a; D'Amico et al., Reference D'Amico, Bergamasco, Carniel, Nacini, Zanasca, Portunato, Teloni, Mori and Barbati2003). Here, primary production is enhanced and could be a primer to the high recruitment of euphausiids, in particular the northern krill Meganyctiphanes norvegica (Labat & Cuzin-Roudy, Reference Labat and Couzin-Roudy1996), which is considered to be the main prey in this portion of the Mediterranean as indicated by faecal analysis (Astruc & Beaubrun, Reference Astruc, Beaubrun, Evans and O'Boyle2001). Likewise, it is suggested that whale movement patterns are closely related to the euphausiids' diurnal vertical migration, implying that fin whales may execute daylight deep dives exceeding 470 m (Panigada et al., Reference Panigada, Zanardelli, Canese and Jahoda1999).
The decrease in fin whale density occurring in the north-western Mediterranean from early autumn to the outset of the spring (Viale, Reference Viale and Barnes1985) suggests that whales migrate seasonally, but very little is known about their migration. Fin whales may stay within the Mediterranean Sea reaching some wintering grounds or may leave the semi-enclosed sea. Few studies envisaged a possible migration through the Strait of Gibraltar, the only passage possible to the Atlantic Ocean (Viale, Reference Viale1977; Guinet et al., Reference Guinet, Mate, Bentaleb, Mayzaud, Clotté, Taupier-Letage, Beaubrun, Dubroca and Monestiez2007). However, some outlines may confirm the yearly presence of fin whales in the Mediterranean Sea. The mitochondrial and nuclear loci differences point to a genetic isolation between the Atlantic and Mediterranean populations of fin whales (Bérubé et al., Reference Bérubé, Aguilar, Dendanto, Larsen, Notarbartolo di Sciara, Sears, Sigurjonsson, Urban and Palsboll1998). Moreover, winter aggregations have been reported in the southern central Mediterranean (Marini et al., Reference Marini, Villetti, Consiglio, Evans and Nice1995) more specifically around the island of Lampedusa. This area may be one of the Mediterranean winter feeding grounds (Canese et al., Reference Canese, Cardinali, Fortuna, Giusti, Lauriano, Salvati and Greco2006; Celona & Comparetto, Reference Celona and Comparetto2006).
Then again, migration patterns of this species throughout its seasonal cycle are poorly studied due to the low monitoring conducted in the eastern Mediterranean basin. This work presents seasonal and spatial whale distribution data in the central Mediterranean Sea, in the vicinity where the western basin connects to the eastern basin and gives a detailed account of whale migration patterns outside the Pelagos Sanctuary. This study was carried out by analysing temporal sighting data collected over five years in different seasons in three diverse areas: (1) the northern part of the Pelagos Sanctuary; (2) the Strait of Messina; and (3) the waters surrounding the island of Lampedusa. Assessing some preliminary indications of regular whale presence, the study highlights the possible need to establish fin whale conservation plans outside of the Pelagos Sanctuary, where maritime traffic is intense.
MATERIALS AND METHODS
Study areas
From 2002 to 2006, three areas in the Mediterranean Sea were monitored for fin whales (Figure 1). The first is located in the north-western Mediterranean Sea and is part of the Pelagos Sanctuary. It extends from the coastline to 43°10′N and from 008°E to 009°52′E, representing 15,920 km2 (Figure 1A). The area is characterized by a heterogenic topographic distribution, notably canyons and seamounts. The Liguro–Provencal current is generated by the inflow of Atlantic Modified Waters and the Tyrrhenian Current, and constitutes a permanent cyclonic circulation with frontal zones (Astraldi et al., Reference Astraldi, Gasparini and Sparnocchia1994). This oceanic landscape provides large biomasses of Meganyctiphanes norvegica (Orsi Relini et al., Reference Orsi Relini, Relini, Cima, Fiorentino, Calandri, Relini and Torchia1992; McGehee et al., Reference McGehee, Demer and Warren2004).

Fig. 1. Location of the three Mediterranean Sea areas monitored for this study on a geographical context map of the Mediterranean Sea with some principal locations and fin whale distribution: (A) in the northern part of Pelagos Sanctuary with isobaths every 500 m; (B) in the Strait of Messina with isobaths every 500 m; and (C) around the island of Lampedusa with isobaths every 200 m.
The second area is the Strait of Messina in the south-western Mediterranean. This is an area of 1220 km2, extending from 015°30′E to 015°4′E and from 38°N to 38°30′N (Figure 1B). The strait is a narrow channel separating the Italian-facing peninsula of Sicily and connecting the Tyrrhenian Sea in the north with the Ionian Sea in the south. It is 32 km long and 3–16 km wide with a mean water depth of 80 m. In the southern part of the strait, the water depth increases rapidly. The 800 m isobaths are found approximately 15 km south of the sill. In the northern part, depth increases more gently: the 400 m isobath is found approximately 15 km north of the sill. The strait connects the western and the eastern Mediterranean basins and has an important role as a site of inflow of Modified Atlantic Waters and draw-out of the Atlantic Ionian Stream (Brandt et al., Reference Brandt, Rubino, Alpers and Backhaus1997). The complex hydrodynamic phenomena caused by the complex bathymetry allow 11 of 13 Mediterranean species of euphausiids (Brancato et al., Reference Brancato, Minatoli, Granato, Sidoti and Guglielmo2001) to reach high biomasses.
The third study area extends from 35°05′E to 35°45′E and from 012°N to 012°58′N (area of 7550 km2, Figure 1C) in the north-eastern Mediterranean surrounding the island of Lampedusa, the largest of the Pelagie Islands. Located in the open sea south of Sicily, the area is characterized by an important primary production regime (Lafuente et al., Reference Lafuente, Vargas, Criado, Garcia, Delgado and Mazzola2005), which develops as a result of up-welling created by the Atlantic Ionian Stream (Robinson et al., Reference Robinson, Sellschopp, Warner-Varnas, Leslie, Lozano, Haley, Anderson and Lermusiaux1999). Geologically, Lampedusa belongs to the continental African plate, and water depth in this area increases gently reaching a maximum of 250 m.
Cetacean sightings
Daily year-round sighting expeditions were carried out in both the Ligurian Sea and the Strait of Messina. Surveys were conducted aboard an 11 m long vessel with a mean cruising speed ranging from 9 to 35 km h−1 during the on-effort period. Transect lines were defined according to a random stratified design. In the north of the Pelagos Sanctuary, stratum 1 was delimited by depths between 800 and 1500 m, and stratum 2 was delimited by depths between 1500 and 2700 m. Meanwhile in the Strait of Messina, stratum 1 was delimited by depths between 20 and 500 m, and stratum 2 by depths between 500 and 1000 m. In both areas, the two strata were surveyed equally in distance per day, but the starting position in each stratum was randomly selected. The observation platform was 4 m above sea level and occupied by four trained observers, who scanned 90° quadrants by naked eye or binoculars. Scanning quadrants were switched every hour.
In Lampedusa island the monitoring covered winter and spring seasons. Surveys were conducted from semi-rigid 5 m long vessels that cruised at maximum speeds of 20 km h−1. Tracks were designed to maximize encounters with whales, according to some indications supplied by fishing boats in the area. Some other sightings were made from fishing boats to achieve photographic identification, but they were not included in these results.
In all studied areas, observations were made from 0900 to 1800 in summer, and from 0800 to 1500 in winter corresponding to Greenwich Mean Time, preferentially in favourable working weather conditions, defined as calm seas, wind speeds never exceeding 3 on the Beaufort scale and the sky not totally cloudy. Time and geographical positions of the boat were recorded with a global positioning system, and environmental conditions (wind speed, wind direction, sea state, sea colour, cloud level and boats in the area), were registered every 30 minutes. Whales were approached upon detection to determine group size and verify presence of calves. Individual rates were calculated by dividing the number of fin whale individuals by the total distance covered under favourable conditions.
Whales' behaviour
Fin whale activity was determined by recording respiration pattern and analysing locomotion behaviour, for approximately one hour from distances of 200 m or greater. Travelling behaviour is displayed by a quasi-linear trajectory with sub-surface constant swimming velocity reaching 5 knots with a single to pair blow counts (Lafortuna et al., Reference Lafortuna, Jahoda, Azzellino, Saibene and Colombini2003; Gannier, Reference Gannier2005). Whereas, feeding behaviour was characterized by a quiet swimming velocity and an extremely convoluted trend well suited to the capture of the prey. In the northern Pelagos Sanctuary, the foraging activity is recognized also by long dive duration and blow counts less than 8 (Lafortuna et al., Reference Lafortuna, Jahoda, Azzellino, Saibene and Colombini2003; Gannier, Reference Gannier2005) while resting behaviour is specified by the very quietly swimming (1–2 knots) and short sub-surface dives, however, socializing is revealed by group individuals' physical contact, and synchrony swimming.
Topographic indicators
Three topographic variables were used as conditioning physical oceanographic indicators of fin whale habitat in the three different study areas. The first two were depth and depth gradient (sea-floor slope) and the third was distance from the coast. Our bathymetry has been borrowed from the US Navy dataset with a grid unit of 1 × 1 nautical miles. Depth gradient and distance from the coast are derived from the depth grid, using a 5 × 5 pixel gradient and are displayed in m/km for depth gradient and km for distance to coast.
RESULTS
During the five-year survey a total of 41,270 km was covered on-effort with acceptable sighting conditions. Ninety-eight fin whale sightings were recorded which amounted to 198 individuals (Table 1).
Table 1. Summary results of the total km surveyed, number of sightings, number of whales, mean group size and individual rate referred to each study area.

Pelagos Sanctuary area
A total of 21,860 km was surveyed on-effort, resulting in 43 sightings of 71 fin whales (Figure 1B). Mean group size was 1.65 (standard deviation (SD) =1.09, range = 1–5); 65% of sightings were single individuals and 18% pairs. Groups with more than paired individuals were quite unusual (13% of sightings) (Figure 2). As shown in Figures 3 and 4, whales were highly abundant in water deeper than 2000 m, far from the coast (15 km away of the shoreline). The individual rate over 100 km in the first strata (depth interval = 800–1500 m) was 0.105 individual whereas in the second strata (depth interval = 1500–2700 m) was 0.493 individual (Table 2). Seasonal distributions of individuals point to a high occurrence from the end of spring to the beginning of autumn as revealed by seasonal individual rates over 100 km spaced out from 0.054 in winter to 0.530 in summer. Meanwhile, there was an approximately equal value in spring and autumn, respectively 0.202 and 0.186 (Figure 5).

Fig. 2. Group size–frequencies of fin whale sightings obtained in the northern part of the Pelagos Sanctuary, in the Strait of Messina and around the island of Lampedusa.

Fig. 3. Depth distribution of fin whale occurrences in the northern part of the Pelagos Sanctuary, in the Strait of Messina and around the island of Lampedusa.

Fig. 4. Distribution of shoreline distance from fin whale positions in the northern part of the Pelagos Sanctuary, in the Strait of Messina and around the island of Lampedusa.
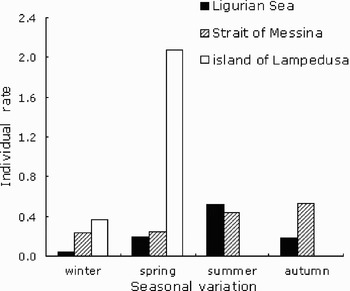
Fig. 5. Variability of the seasonally individual rate on each study area.
Table 2. Distribution of the total km surveyed related to each strata, number of sightings, number of whales and the encounter rate on the Ligurian Sea and Messina Strait.

km survey, on-effort km surveyed; No. sight, number of sightings; No. ind, number of individuals; Ind km−1, individual rate.
Feeding behaviour was observed throughout the study area mainly for single individuals or pairs. It was observed in 68.4% of the total sightings, largely common during summer with 82.8% of the total seasonal sightings (Table 4). Whale course trajectory was convoluted with a slow swimming velocity. The shorter surface duration counts were 2–6 blows and come after a long dive duration sometimes exceeding 8 minutes. Socializing behaviour was mostly recorded for groups composed of 3–4 individuals representing only 3.4% of the total sightings. Whales displayed a synchrony cycle of surfacing-diving activity near to the boat, crossing the boat's heading with a long sub-surface linear trajectory with physical contacts between individuals. During one sighting, we assisted a very close parallel side by side individual swimming with one turn on its side exposing its abdomen.
Strait of Messina area
A total of 14,830 km was covered on-effort and 39 positions were recorded, which represented 60 sightings of fin whales (Figure 1C). The mean group size was about 1.53 (SD = 0.6, range = 1–3), with 51% of sightings being single individuals, and only 3% pairs (Figure 2). Specimens were observed in a narrow area indicating a high distribution in shallow water reaching the coast within the 15 m depth (Figure 3). The mean shoreline distance was about 50 km (Figure 4). The individual rate on the first stratum (depth interval = 20–500 m) was twice as much as the one in the second stratum (500–1000 m) (Table 2). Seasonal presence of individuals is major from the end of the summer to the end of autumn (Figure 5). Winter and spring individual rates over 100 km were quite constant, respectively at 0.237, 0.251 and 0.532 in autumn (Table 3).
Table 3. Summary results on seasonal on-effort km surveyed, number of sightings, number of whales, and individual rate referred to each study area.

LS, Ligurian Sea; MS, Strait of Messina; LI, Lampedusa Island; km survey, on-effort km surveyed; No. sight, number of sightings; No. ind, number of individual; Ind km−1, individual rate.
Only two behaviours (feeding and travelling) have been equally observed all year around. However, seasonal sightings were largely favouring feeding activity during spring (87.5%) and travelling in autumn (80%) (Table 4). During travel activity, whales' course trajectory was quasi-linear, with a long sub-surface distance follow-up of a single blow count. Compared to vessel speed, their swimming velocity was 4–5 knots.
Table 4. Summary results on percentage of seasonal sightings of fin whale behaviour on each study area.

LS, Ligurian Sea; MS, Strait of Messina; LI, Lampedusa Island; Feed, feeding; Trav, travelling; Soci, socializing; Rest, resting.
Lampedusa area
Near the island of Lampedusa where surveys covered 4580 km, 16 whale groups representing 67 individuals were recorded during the 5 y of study (Figure 1B). Figure 2 indicates the preference of the species for large groups of individuals. Twelve per cent of the sightings were of single mysticetes, 25% were couples and 56% were groups of more than three individuals. Mean group size was 4.18 (SD = 2.8, range = 1–10). In this area, individuals were seen near to the coast up to 5 km (Figure 4), and were encountered in waters as shallow as 10 m in depth. Seasonally high individual rates were recorded (2.082 individuals over 100 km) and whale presence in this area seems to be confined to from late winter until early spring (Table 3). The periodicity is logically due to the restriction of our surveys to this period, nevertheless no records have been reported by fishing boats outside of this sampling window.
The two behaviours observed were feeding and resting. The feeding activity was largely common throughout this area within groups and individuals, representing 75% of the total sightings. Dissimilar to the Ligurian Sea, this behaviour was characterized by quickly swimming closely parallel to the surface so that throats were completely visible due to their open mouths engulfing water.
Calves were seen three times in two different areas. Two sightings occurred in the Ligurian Sea in the summer 2005 and both were estimated to measure less than 10 m long. During one of the observations, the newborn was socializing with its supposed mother. The third sighting occurred in the Strait of Messina in the autumn 2002; the newborn was estimated to be less than 9 m.
DISCUSSION
In the north-western Ligurian Sea, individual rates reveal peak abundance from late spring to the late summer and the lowest abundance during winter. In addition, a major fraction of these sightings were for isolated animals in deep water (with a mean depth of 2043 m) which possessed a deep water feeding behaviour (in 82.8% of seasonal sightings). This seasonal distribution is in agreement with previous studies which found favourable summer feeding habitats in the Mediterranean Sea (Forcada et al., Reference Forcada, Notarbartolo di Sciara and Fabbri1995; Gannier, Reference Gannier1998a,Reference Gannierb, Reference Gannier2002). This activity is correlated to the relative high abundance of Meganyctiphanes norvegica, the main prey of fin whales as revealed by faecal analysis (Astruc & Beaubrun, Reference Astruc, Beaubrun, Evans and O'Boyle2001). This euphausiid is a cosmopolitan species that, like other plankton, shows a strong diel vertical migration. It tends to concentrate within a layer under the thermocline during daytime and to disperse in the upper layer at night-time (Anderson et al., Reference Anderson, Sardou and Nival1992). The absence of the prey at the surface justifies the absence of surface feeding behaviour in this area. Compared to the outcome observed by Gannier et al. (2002) and Panigada et al. (Reference Panigada, Notarbartolo di Sciara, Zanardelli, Airoldi, Borsani and Jahoda2005), our results are consistent, with respectively 2026 and 2317 m. The infrequency of sightings starting at the end of autumn has been explained by the decreases and sometimes disappearance of the biomass of fin whale prey (Andersen et al., Reference Andersen, Vilan, Ruellet and Gubanova2001). Abundance sightings of single individuals represent 65%. Indeed single whales are frequent in the entire western area; Forcada et al. (Reference Forcada, Aguilar, Hammond, Pastor and Aguilar1996) found 68% of all fin whales were swimming as single individuals during a zigzag survey that covered the area from the Strait of Gibraltar to the island of Corsica.
This study is the first to investigate whale occurrence in the Strait of Messina. Sightings showed a year round distribution with a remarkably high concentration from the end of summer to the end of autumn. Group size was similar to the one observed in the Pelagos Sanctuary, manifesting a dominant presence of single individuals (51%) and pairs (43%). The mean group size (1.53) obtained in this study is not significantly different from the one obtained in the southern Tyrrhenian Sea (1.33) (Gannier, Reference Gannier2002). Whales were found mostly in shallow waters less than 500 m, mainly feeding during both the spring and summer seasons. The area should therefore be classified as a probable seasonal feeding ground. Based on the work of Brancato et al. (Reference Brancato, Minatoli, Granato, Sidoti and Guglielmo2001), the small connection area between both Mediterranean basins was considered as a site for the annual distribution of 11 species of euphausiid where M. norvegica is abundant due to the intense and complex hydrodynamic phenomena caused by up-welling. The year round presence of M. norvegica can therefore explain the year round fin whale distribution in this area.
Nonetheless, travelling behaviour was largely common from late autumn to early winter, which may indicate a migration from the southern area to other contiguous zones. It has been hypothesized that fin whales migrate between areas where krill is highly abundant such as the Ionian Sea (Catlano et al., 2001). Indeed one fin whale has been identified in both the Ionian Sea and the Strait of Messina (Tringali et al., Reference Tringali, Caltavuturo, Gurrieri and Di Martino1999).
Around the island of Lampedusa, our study was limited to both the winter and spring seasons. Sightings show an important individual rate from late winter to early spring with an indisputable connection to shallow waters of depths shallower than 250 m close to the coast. This result is consistent with sighting data reported by other studies (e.g. Canese et al., Reference Canese, Cardinali, Fortuna, Giusti, Lauriano, Salvati and Greco2006). In this area, only 12.5% of all sightings were single individuals; 62.5% were groups composed of more than two whales. Mean group size in this study is twice that reported by Canese et al. (Reference Canese, Cardinali, Fortuna, Giusti, Lauriano, Salvati and Greco2006) and can result from the short period of monitoring (8 d) which allowed 14 sightings with a research vessel and six with an opportunistic vessel. While feeding behaviour was similarly observed in both the northern Pelagos Sanctuary and the Strait of Messina area, the strategy differs around the island of Lampedusa. Whales were engaged in surface feeding with synchronic collaboration in groups of ten individuals. Similar behaviour was reported previously with 5–20 whales (Marini et al., Reference Marini, Villetti, Consiglio, Evans and Nice1995; Canese et al., Reference Canese, Cardinali, Fortuna, Giusti, Lauriano, Salvati and Greco2006). This new described strategy may be a possible indication of the presence of another prey other than the deep vertically migrating M. norvegica. According to Canese et al. (Reference Canese, Cardinali, Fortuna, Giusti, Lauriano, Salvati and Greco2006), zooplankton sampling collected where fin whales fed indicate the presence of Nyctiphanes couchi. These observations support the idea that prey distribution is the driving force influencing fin whale habitat (Woodley & Gaskin, Reference Woodley and Gaskin1996). It highlights the relative importance of physiographic parameters such as depth preference that can play on the fin whale distribution, which are frequently used in cetacean studies (Davis et al., Reference Davis, Ortega-Ortiz, Ribic, Evans, Biggs, Ressler, Cady, Leben, Mullin and Würsing2001). In some way, in the Ligurian Sea, whales were more abundant in deep water (1500–2700 m) while in the Strait of Messina and around the island of Lampedusa whales were mostly observed in shallow water (10–500 m).
The decreasing density in the Ligurian Sea from late summer coincides with the highest distribution in the Strait of Messina where whales seem to be mostly in a transfer phase from autumn to early winter. Then whales aggregate around the island of Lampedusa until the beginning of the spring season especially the month of March. This seasonal dispatching and the related activities can be the result of possible cyclic migration of some Mediterranean individuals between both western and eastern basins.
In our study, the schematic seasonal pattern displayed by the Mediterranean population can be represented as aggregation in summer in the north-western Mediterranean Sea, one of the main summer feeding grounds. Afterwards, some individuals migrate through the Strait of Messina during autumn and winter to reach the eastern basin where whales are clustered until spring. Moreover, based on preliminary photo-identification results obtained with the three different monitoring programmes, one individual was identified in the Pelagos Sanctuary on two occasions in late May and early September 2005 and around the island of Lampedusa in February 2005. This whale was clearly recognizable due to its scars demonstrating a previous collision with a propeller, running from behind its blowhole to its dorsal fin. The detection of the same individual in the two areas characterized by two different feeding behaviours reveals the role food availability can play in structuring whale distributions.
To conclude, fin whale distributions observed in the central part of the Mediterranean Sea indicate a probable presence year round. However, the results are still insufficient to prove year round residency of a large portion of the fin whale Mediterranean population. A small part of the population at least seems to use different feeding strategies in different areas with various topographies. The depth related to distribution of whales changes, so topography seems to be an insufficient indicator to describe it. Furthermore, fin whales may have a wider range of prey than a single-type-prey specialization (M. norvegica) as previously thought. Fin whales may adapt their foraging activity according to the abundance of potential preys. Whale attendance may be largely dependent on prey availability and distribution. The identification of one individual in different Mediterranean areas indicates the necessity to conduct more surveys on a larger scale in order to better comprehend fin whale ecology. If some individuals remain inside the Mediterranean year-round, it underlines the importance of effective management strategies which will support healthy fin whale protection outside of the Pelagos Sanctuary, especially where threats are present.
ACKNOWLEDGEMENTS
The work was partially funded by the Italian ‘Ministero dell'Ambiente e della Tutela del Territorio’ and ACCOBAMS. Surveys conducted by the Biology Department of the University of Genoa were sponsored by Yanmar and SACS Boatyard. We are grateful to the many students involved in the data collection with special thanks to Agostino Corsi, to Massimo D'incà and to Massimiliano Rosso. Tomah Phenix and Andrew K. Sweetman are also thanked for English corrections. Special thanks also to the anonymous referees for their useful comments and their considerable help in improving the manuscript.









