


Introduction
Diverse invertebrate faunas have been described from the widely distributed methane seep deposits of the Upper Cretaceous Pierre Shale of the Western Interior (Landman et al., Reference Landman, Cochran, Larson, Brezina, Garb and Harries2012). Echinoderms, including crinoids, echinoids, ophiuroids, and asteroids, have been collected; the known asteroid specimens are treated here. A guide and background to the Western Interior seep ecosystems was furnished by Shapiro and Fricke (Reference Shapiro and Fricke2002), and an overview of ancient chemosynthetic settings was provided by Campbell (Reference Campbell2006). Although echinoderms are important deep-water benthic invertebrates (Gage and Tyler, Reference Gage and Tyler1991), they are of limited and incompletely understood significance in chemosynthetic settings. In the Gulf of Mexico, an extant species of the upper-slope predatory forcipulate asteroid Sclerasterias Perrier, Reference Perrier1891, was reported to be a seep colonist (Carney, Reference Carney1994), and a diversely fossiliferous Oligocene setting in Oregon provides another potential seep occurrence (Burns and Mooi, Reference Burns and Mooi2003, p. 95). Recently and for the first time, asteroids are reported from hydrothermal vents (Mah et al., Reference Mah, Linse, Copley, Marsh, Rogers, Clague and Foltz2015), these occurrences including a new family, a new genus, and two new species, the taxa emerging stemward in one major branch of forcipulatacean diversification.
All new Pierre Shale seep asteroid specimens, excepting a single fragment, are assigned to Betelgeusia brezinai new species, family Radiasteridae. The Radiasteridae today is known from relatively few but geographically widely separated largely deeper-water settings (448–2423 m, 245–1,325 fathoms; Clark, Reference Clark1946), and because of its somewhat problematic morphology, the genus and family long have been taxonomically difficult (Clark and Downey, Reference Clark and Downey1992, p. 89). Positioning of the Radiasteridae within crown-group diversification (see the following) argues to its phylogenetic significance.
A single small Pierre Shale fragment is assigned to the Forcipulatida. Forcipulate subdivisions of Spencer and Wright (Reference Spencer and Wright1966) are broadly typical of traditional morphology-based taxonomic arrangements whereas a recent molecular study (Mah and Foltz, Reference Mah and Foltz2011b) introduced significant changes placed in a historical biogeographic framework. Because of the limited data available for the single fossil, and because of taxonomic complexities, the fragment is assigned to the Pedicellasterinae in the traditional sense of Spencer and Wright (Reference Spencer and Wright1966); placement in the updated framework (Mah and Foltz, Reference Mah and Foltz2011b) is not attempted.
Previous reports of Upper Cretaceous Western Interior asteroids
Two asteroids were reported from the Upper Cretaceous of the Western Interior, one from the Turonian–lower Campanian Cody Shale of Wyoming (Weller, Reference Weller1905; Branson, Reference Branson1947) and another from the Pierre Shale of Colorado (Griffitts, Reference Griffitts1949, p. 2025). Gill and Cobban (Reference Gill and Cobban1966, p. A23) reported arms of an asteroid associated with hundreds of small echinoids in a concretion collected from near the top of the upper Campanian Baculites reesidei Zone; although echinoids were illustrated, the asteroid was not. An asteroid arm was included among echinoderm remains in a middle Campanian cold seep setting (Kato et al., Reference Kato, Oji and Shirai2017).
Geologic setting
Locations of the collecting localities are illustrated in Figure 1 and their biostratigraphic positioning in Figure 2; locality AMNH 3529 is illustrated in Figure 3, and locality 3418 is documented in Landman et al. (Reference Landman, Cochran, Larson, Brezina, Garb and Harries2012). Betelgeusia brezinai is illustrated in Figures 4.1–4.7, 5.1–5.13 and pedicellasterid? sp. in Figure 5.14, 5.15.
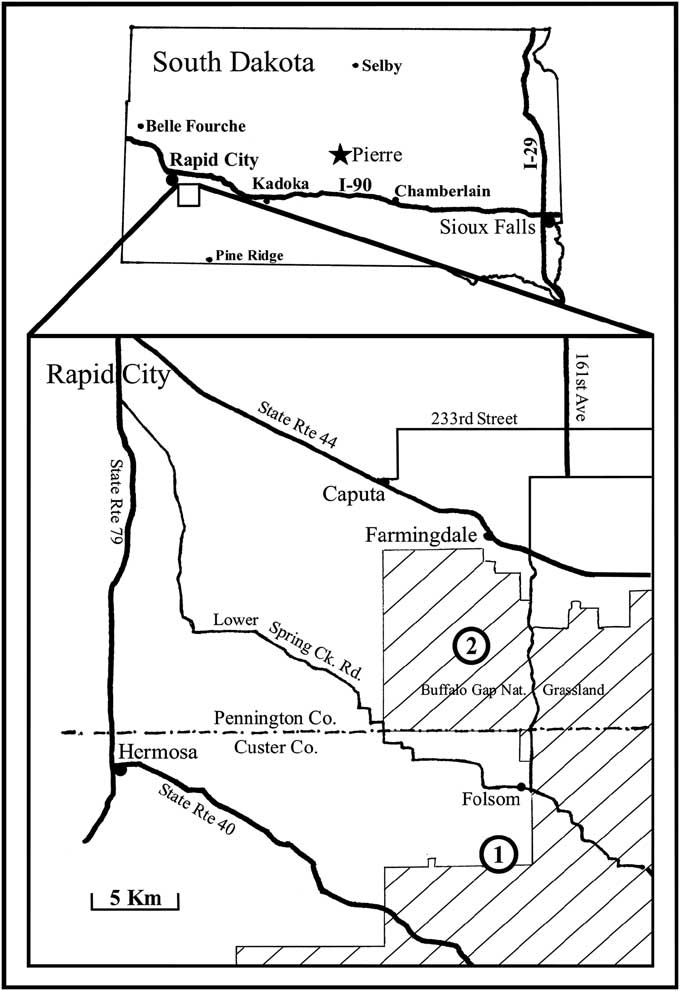
Figure 1 Map of South Dakota, USA, showing approximate asteroid collecting localities.
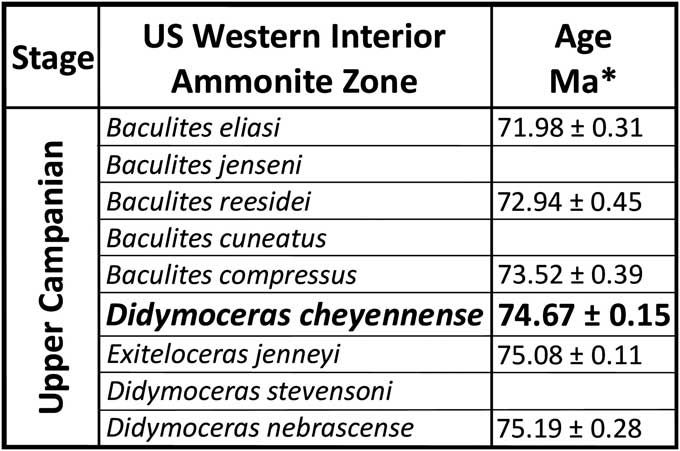
Figure 2 Upper Campanian ammonite zonal table of the Western Interior. *Age dates from Cobban et al. (Reference Cobban, Walaszczyk, Obradovich and McKinney2006). Zone of occurrence of the asteroid Betelgeusia brezinai n. sp. in bold.

Figure 3 AMNH loc. 3529. All asteroid specimens at this locality were found within the black outline, 2–3 m north of the micritic mass of the carbonate core (arrow).

Figure 4 Betelgeusia brezinai n. gen. n. sp. (1–4) American Museum of Natural History, holotype AMNH-FI 111823; (1, 2) dorsal and ventral views, the primary ossicles partially obscured by accessories and foreign shelly material; (3) interbrachium toward top of (2); mouth frame to right, many accessories remaining, fasciolar channeling between actinal series is not closely aligned with marginal series (to left); (4) marginal form, fascioles, and accessories. (5, 6) Paratype AMNH-FI 111824; (5) ventral lateral view of arm, superomarginals granulate, inferomarginal with robust edging spinelets, accessories more nearly granular on ventral surface; edges of actinal rows, these aligned, separated by fasciolar grooving; (6) ventral view showing adambulacral form, alignment of actinals, marginal form; comparatively few accessories remain. (7) Paratype AMNH-FI 111852, the only specimen derived from locality AMNH-FI 3418; MAO, form and positioning of actinals and marginals; see Fig. 5.1. (1, 2, 6, 7) Scale bars=10.0 mm; (3–5) scale bars=5.0 mm.

Figure 5 (1–13) Betelgeusia brezinai n. gen. n. sp. (1) Paratype AMNH-FI 111852, disk in ventral aspect, aligned series of imbricate actinals separating two mouth angle ossicle (MAO) pairs (upper left) from inferomarginal series (far right); compare (10, 11). (2, 3) Paratype AMNH-FI 111825, arm tip in dorsal and ventral aspect; (2) the terminal is large, as is common among paxillosidans (including the Radiasteridae) with attenuated arm tips; (3) arm tip upturned as is common in life, an orientation enabling extension of sensory distal tube feet into the environment; marginals with aligned, transverse accessory pustules; marginals abut spinose adambulacrals. (4–6) Paratype AMNH-FI 111826, marginal series and associated ossicles in dorsal, lateral, and ventral views; (4) ossicular debris including few abactinals and actinals that were displaced dorsally; (5) angular openings at abradial edge of superomarginals suggest pedicellariae openings, these otherwise unknown among radiasterids; (6) inferomarginal pustule configuration and a few remaining accessories; actinals largely disrupted. (7–9) Paratype AMNH-FI 111827; (7) superomarginals and displaced small paxilliform abactinals; (8) both marginal series, lateral surfaces showing fasciolar ridges; (9) inferomarginals above with spinelet pustules; displaced actinals below. (10, 11) Paratype AMNH-FI 111828, actinal and marginal series in ventral and inclined ventral views; the strongly imbricating and aligned actinals are characteristic of radiasterids, here potentially partially displaced taphonomically, but of overall orientation likely similar to those assumed during normal body flexure. (12) Paratype AMNH 111829, dorsal aspect near juncture of arm and disk, compare (1); superomarginal form, inferomarginals extending laterally beyond superomarginals; abactinal form. (13) Paratype AMNH-FI 111830, fragment in ventral aspect, shingled actinal rows (arrows) extend adradially from two inferomarginals. (14, 15) Pedicellasterinae? gen. indet. sp. indet., figured specimen AMNH-FI 111853, two views of the single available fragment; (14) dorsal ossicular views, cross-shaped body-wall ossicles are characteristic of forcipulates; a single ambulacral (arrow) is comparatively large, the figure ‘8’ or wasp-shape indicates a single row of podia rather than offset podial series of the Asteriidae; (15) dominantly ventral ossicular views, adambulacral (arrows) form is consistent with a pedicellasterid assignment. (1–13) Scale bars=1.0 mm; (14, 15) scale bars=2.0 mm.
Cretaceous seep deposits were first reported by Gilbert and Gulliver (Reference Gilbert and Gulliver1895), who provided detailed description of carbonate occurrences northeast of Pueblo, Colorado. The transition from the roughly cylindrical carbonate ‘core’ to the surrounding shale was found to be abrupt, with differential weathering and erosion yielding a distinctive conical pattern resembling Native American lodges, the structures therefore dubbed ‘tepee buttes.’ Several possible explanations for carbonate core origins were offered, but in the end it was concluded that “our present knowledge is not sufficient” (Gilbert and Gulliver, Reference Gilbert and Gulliver1895, p. 340), and butte genesis remained enigmatic for 100 years. The diverse molluscan fauna of the carbonate cores includes the index ammonites Didymoceras nebrascense Meek and Hayden, Reference Meek and Hayden.1856 and Didymoceras cheyennense Meek and Hayden, Reference Meek and Hayden.1856, whereas the adjacent shales are devoid of fossils.
Chemosynthetic communities were first discovered in a modern cold seep setting in the Gulf of Mexico in 1983 using the manned deep submergence vehicle (DSV) Alvin (Paull et al., Reference Paull, Hecker, Commeau, Freeman-Lynde, Newmann, Corso, Golubic, Hook, Sikes and Curray1984); since then, cold seeps and their faunas have been studied from locations around the globe (e.g., Levin, Reference Levin2005). Extant seep sites were compared to the tepee butte structures and found to provide a compelling match (e.g., Kauffman et al., Reference Kauffman, Arthur, Howe and Scholle1996). Further studies measuring δ13C and δ18O levels in the carbonate core rocks support the methane (cold) seep hypothesis for tepee buttes and other carbonate mound structures, some retaining rich Late Cretaceous faunas (Landman et al., Reference Landman, Cochran, Larson, Brezina, Garb and Harries2012; Cochran et al., Reference Cochran, Landman, Larson and Meehan2015).
Ancient methane seep deposits ranging in age from late Cenomanian to early Maastrichtian are widely distributed throughout the Upper Cretaceous of the Western Interior (Metz, Reference Metz2008; Landman et al., Reference Landman, Cochran, Larson, Brezina, Garb and Harries2012; Larson et al., Reference Larson, Brezina, Landman, Garb and Handle2014; Hunter et al., Reference Hunter, Larson, Landman and Oji2016) and have been further documented from east-central Montana, south-central Colorado, western Kansas, northwestern Nebraska, western South Dakota, eastern Wyoming (Metz, Reference Metz2010; Landman et al., Reference Landman, Cochran, Larson, Brezina, Garb and Harries2012; Larson et al., Reference Larson, Brezina, Landman, Garb and Handle2014), and southern Utah (Kiel et al., Reference Kiel, Wiese and Titus2012).
The seep deposits from the Black Hills of South Dakota to southern Colorado were formed between 76 Ma and 70 Ma (Larson et al., Reference Larson, Brezina, Landman, Garb and Handle2014) in response to methane seepage through seafloor faults (Metz, Reference Metz2010). Larson et al. (Reference Larson, Brezina, Landman, Garb and Handle2014) hypothesized that they developed along fault lines due to tectonic activity, allowing for dewatering of the thick Cretaceous shales prior to and coincident with the Black Hills uplift. At any one locality, the seeps typically occur in groups of three or more, representing either closely connected conduits or movement of a single seep through time. Howe (Reference Howe1987) reported that most fossil seep core dimensions near Pueblo, Colorado, range from 1 to 8 m in width and up to 30 m in height, whereas many seep deposits around the Black Hills are larger (Larson et al., Reference Larson, Brezina, Landman, Garb and Handle2014).
The lithology of ancient seeps has been described by a number of authors (Kauffmann et al., 1996; Metz, Reference Metz2008; Hendricks et al., Reference Hendricks, Gong, Jones, Hsiung, Tao, Metz, Raymond and Pope2011). Using the seep structure model of Handle (Reference Handle2014), Meehan and Landman (Reference Meehan and Landman2016) related faunal expressions to seep architecture, with variation among seep occurrences resulting from variation among causal factors of original depth, lateral extent of seepage, methane flux, oxygen levels, water temperature (Levin, Reference Levin2005), seep longevity (Larson et al., Reference Larson, Brezina, Landman, Garb and Handle2014), and perhaps as yet unrecognized factors; additional work is needed.
Regardless of the lithology and architecture, most of the carbonate core intervals are micritic and were formed by bacterial-mediated anaerobic oxidation of methane (AOM) coupled with sulfate reduction, the overall chemical reaction being: CH4+SO4 −2→HCO3−+HS−+H2O (Levin, Reference Levin2005). Carbonate precipitation commonly occurred from the seafloor surface to several meters within the substrate (Hendricks et al., Reference Hendricks, Gong, Jones, Hsiung, Tao, Metz, Raymond and Pope2011; Cochran et al., Reference Cochran, Landman, Larson and Meehan2015). Inoceramid bivalve accumulations together with fine clastics provided a varied substrate that supported a diverse primary autotrophic fauna of tube worms and bivalves nourished by methane and/or sulfide chemosymbionts. The autotrophs were prey for predatory gastropods, crabs (MacAvoy et al., Reference MacAvoy, Carney, Fisher and Macko2002), and cephalopods (Landman et al., Reference Landman, Cochran, Larson, Brezina, Garb and Harries2012). Cemented ‘shell hash’ and seep-associated concretions can be present at the periphery of the main micritic mass (‘core’; Hunter et al., Reference Hunter, Larson, Landman and Oji2016, fig. 3).
All the seeps from Colorado to the Black Hills occur in the Pierre Shale, a thick, homogenous deposit of black shale more than 1,000 m thick deposited from the early Campanian through early Maastrichtian in a seaway generally 300 m deep or less (Gill and Cobban, Reference Gill and Cobban1966). Pierre Shale ammonite paleontology and biostratigraphy were summarized by Larson et al. (Reference Larson, Jorgensen, Farrar and Larson1997), the pertinent Campanian zonation summarized in Figure 2. The fossil methane seeps encircling the Black Hills are from the Didymoceras nebrascense Zone (base of upper Campanian) through the Baculites grandis Zone (lower Maastrichtian) (Larson et al., Reference Larson, Brezina, Landman, Garb and Handle2014), the asteroids from two sites in the Didymoceras cheyennense Zone (upper Campanian), AMNH loc. 3418 and AMNH loc. 3529; a partial arm from AMNH loc. 3495 is lost. Additional asteroid specimens were found at an undocumented seep site in the 1990s, their present location unknown. All asteroid specimens were found on the flanks of the main core micritic masses (Fig. 3).
Echinoderm paleoecology of Pierre Shale cold seep environments has been studied on the basis of skeletal elemental chemical analyses and stable carbon isotopes (Kato et al., Reference Kato, Oji and Shirai2017). A crinoid species recovered from seep settings was found to be both morphologically and geochemically differentiated as to argue that this species is endemic and specialized to the seep environments, whereas Western Interior Seaway echinoids also based on morphologic and geochemical data were judged to have been a part of the background fauna and not restricted to seep settings. Geochemical data are not available for B. brezinai but its morphology does not differ in any obvious manner from morphologies of species of Betelgeusia that are not associated with seep settings, and therefore the new asteroid, like the echinoids, likely is a part of the background fauna.
Terminology
Terminological usage for the asteroids follows Spencer and Wright (Reference Spencer and Wright1966) and Blake (Reference Blake2013). The primary skeleton forms the body wall. The accessory skeleton includes the generally abundant spines, granules, and pedicellariae seated on all primary ossicles except ambulacrals. Ambulacral ossicles form a double series along the axis of the arm and serve as the foundation for the water vascular system. Ambulacrals are vaulted to form the ambulacral furrow, and the mouth angle ossicles (MAO) are the proximal-most ossicles of the ambulacral series. Ambulacral ossicles articulate with and are immediately dorsal to the adambulacral ossicles. Marginal ossicles form either a single inferomarginal series or a double inferomarginal plus superomarginal series, one or both positioned near the margin of the asteroid. A primary circlet of abactinals can be differentiated on the dorsal disk, and dorsal midarm ossicles can be enlarged and/or otherwise differentiated to form a carinal series. The madreporite is the outlet of the water vascular system, on the dorsal disk surface of radiasterids. No, few, or many actinals occur between the inferomarginals and adambulacrals.
Repository and institutional abbreviation
All specimens are housed in the collections at American Museum of Natural History (AMNH), Division of Paleontology.
Systematic paleontology
Order Paxillosida Perrier, Reference Perrier1884
Family Radiasteridae Fisher, Reference Fisher1916
Known genera
Betelgeusia Blake and Reid, Reference Blake and Reid1998 (middle Cretaceous); Gephyreaster Fisher, Reference Fisher1910 (extant); Indiaster Rao, Reference Rao1957 (Middle Jurassic); Radiaster Perrier, Reference Perrier1881 (extant).
Remarks
Although only two extant genera are assigned to the Radiasteridae, taxonomic interpretations of the family have differed (Fisher Reference Fisher1911, Reference Fisher1919; Clark, Reference Clark1946; Blake, Reference Blake1987; Clark and Downey, Reference Clark and Downey1992). Two extant genera, Radiaster Perrier, Reference Perrier1881 (=Mimaster), represented by four species, and monospecific Gephyreaster Fisher, Reference Fisher1910 (not Gephyriaster Spencer and Wright, Reference Spencer and Wright1966) have been assigned to the family. Because of ambiguities in the literature as well as the relatively robust construction of Gephyreaster as compared to Radiaster, Blake (Reference Blake1987) and Blake and Reid (Reference Blake and Reid1998) limited treatment of extant Radiasteridae to Radiaster itself.
Molecular methodologies are of increasing importance in the study of evolution and biogeography, these approaches potentially particularly useful for complex settings and evolutionary histories (Hardy et al., Reference Hardy, Carr, Hardman, Steinke, Corstorphine and Mah2010) such as those represented here by both seep occurrences and problematic taxa such as the Radiasteridae. As true of other invertebrates, phylogenetic relationships among major subdivisions of the crown-group Asteroidea are undergoing significant revision with evolving molecular results. At writing, Radiaster has not yet been treated in a molecular phylogenetic analysis whereas Gephyreaster clustered with Pseudarchaster Sladen, Reference Sladen1889, the two assigned to the Pseudarchasteridae within an otherwise traditional paxillosidan and notomyotidan (Benthopectinidae) complex (Mah and Foltz, Reference Mah and Foltz2011a). Middle Cretaceous Betelgeusia Blake and Reid (Reference Blake and Reid1998) was the first fossil radiasterid to be recognized. Indiaster Rao, Reference Rao1957 was originally assigned to the Goniasteridae but later transferred to the Radiasteridae (Blake and Reboul, Reference Blake and Reboul2011), the genus originally described as “probably of Upper Bathonian age” (Middle Jurassic) (Rao, Reference Rao1957, p. 213). Given molecular analysis together with the closing of the morphologic gap between Radiaster and Gephyreaster as based on species of Betelgeusia, five fossil and extant genera, Betelgeusia, Gephyreaster, Indiaster, Pseudarchaster, and Radiaster, form a monophyletic complex. Molecular positioning of Radiaster and subsequent status evaluation of the terms ‘Radiasteridae’ and ‘Pseudarchasteridae’ are left for the future.
Genus Betelgeusia Blake and Reid, Reference Blake and Reid1998
Type species
Betelgeusia reidi Blake and Reid, Reference Blake and Reid1998, by monotypy.
Other species
B. exposita Blake and Jagt, Reference Blake and Jagt2005; B. orientalis Blake and Reboul, Reference Blake and Reboul2011.
Emended diagnosis
Radiasterid with relatively robust, fasciolate marginals; fascicular expression, particularly superomarginal expression, varied among species. Abactinals subgranular to robust paxillae, not tabulate. Actinal ossicles many, small, aligned in strongly imbricate series that extend from inferomarginals to adambulacrals; well-defined grooves separate exposed crowns of adjacent series. Accessories varied: granules, robust short spinelets, or elongate delicate spinelets; pedicellariae not recognized.
Remarks
A range of marginal, accessory, and abactinal variation is developed among the four species of Betelgeusia, although generic subdivision is not deemed justified. Indiaster is incompletely known, but unlike Betelgeusia, the actinal ossicles appear to be relatively robust, rectangular, and at most, only weakly imbricate. The two marginal series of Indiaster appear to be similar, robust, and closely fitted, the two series not distinctive as in Betelgeusia. Enough remains of the Indiaster specimens to suggest that accessory development was subdued. All primary ossicles of extant Radiaster are small and delicate, the marginals blade-like or subpaxilliform, and paxillae are delicate. Spinelets are small. Diagnostic separation of Gephyreaster and Radiaster (=Mimaster) was based on presence of stout tabulate paxillae and a peculiar MAO accessory arrangement in the former (Fisher, Reference Fisher1911, p. 175), criteria that also serve to separate Gephyreaster from Betelgeusia. In Gephyreaster as in Betelgeusia, marginals and, in particular, inferomarginals are comparatively robust and closely fitted, although smaller accessories are similar between Gephyreaster and some Betelgeusia.
The strongly imbricate, paddle-like actinals of Betelgeusia aligned in well-defined series form a distinctive generic character complex. The almost upright actinal occurrences of some fragments of B. brezinai (Fig. 5.10, 5.11) might have been accentuated taphonomically, but not significantly so from natural positioning assumed during normal life activities.
Betelgeusia brezinai new species
2017 asteroid; Kato et al., p. 223, fig. 2I.
Holotype
Locality AMNH 3529, holotype AMNH-FI 111823: a small individual in overall good condition and exposing both surfaces, although distorted through sediment compaction and breakage, and partially obscured by foreign shell fragments. Remaining arm radii R=18, 12, 12, 4, and 11 mm; r of three remaining interbrachia 8–10 mm.
Paratypes
Locality AMNH-FI 3529, paratypes: an illustrated, nearly complete specimen exposed in ventral aspect, AMNH-FI 111824, an arm tip with both surfaces exposed, paratype AMNH-FI 111825; five illustrated small fragments, AMNH-FI 111825–AMNH-FI 111830; seven partial paratypes, relatively complete to fragmentary, AMNH-FI 111831–AMNH-FI 111837; and 14 small to tiny paratype fragments of larger specimens, AMNH-FI 111838–AMNH-FI 111851.
Locality AMNH 3418, a single paratype, AMNH-FI 111852, exposed in ventral aspect, on a small block of fossiliferous carbonate matrix. Two arms are nearly complete, R=19 mm; truncated R between about 9 and 12 mm, interbrachial radius 10 mm. Inferomarginals of interbrachia are complete; a single superomarginal series is visible, but no abactinals; no accessory ossicles remain. Approximately 14 inferomarginals between the interbrachial plane and arm tip of nearly complete arm R=19 mm, r~9 mm. About 13 to 15 marginal pairs between interbrachial midline and terminal.
Occurrences
Locality AMNH-FI 3529: Didymoceras cheyennense Zone, Pierre Shale, upper Campanian, Late Cretaceous; Pennington County, South Dakota. Exact locality information is available to qualified researchers from the American Museum of Natural History. All available specimens but one were collected at locality 3529.
The locality is a low-profile mound consisting of tan-weathered shales at the top with less-weathered black and dark-gray shales at its base. A 1–2 m main carbonate micritic mass (‘core’) is exposed with associated concretions and cemented shell hash carbonates on the core flanks; weathered carbonates are scattered downslope. The original Pierre Shale surrounding the seep was eroded during the Paleocene through early Eocene, exposing the carbonates to weathering. The seep was later reburied by Chadron Formation deposition during the middle to late Eocene, composite sediments including volcanic ash from the west and alluvial gravels and clay from the Black Hills. The seep was reexposed as a result of weathering during the Pleistocene and Holocene. The seep supported a highly diverse fauna including ammonites, crinoids (Lakotacrinus brezinai Hunter et al., Reference Hunter, Larson, Landman and Oji2016 and an unknown feather star), two types of tubeworms, two types of bryozoans, various gastropods and bivalves (including teredinids and inoceramids), unidentified crabs, a ghost shrimp, and echinoids (Larson et al., Reference Larson, Brezina, Landman, Garb and Handle2014). Preservation of the asteroids is varied, including both articulated and fragmentary specimens, exposing both surfaces, the suite recovered both from an approximately 3×5 m area in the carbonate shell hash and loose in the weathered tan shale (Fig. 3).
Locality AMNH-FI 3418: Didymoceras cheyennense Zone, Pierre Shale, upper Campanian, Late Cretaceous; Custer County, South Dakota. Exact locality information is available to qualified researchers from the American Museum of Natural History.
A vertically exposed carbonate seep cross section measuring 13 m high and 20 m wide, surrounded by black and gray shales with orange and yellow partings near the carbonates (Landman et al., Reference Landman, Cochran, Larson, Brezina, Garb and Harries2012). The base is buried beneath slumped black and dark-gray shale, but the roughly cylindrical carbonate ‘core’ and its conduit-like structures are readily visible. A detailed map of the seep face (Landman et al., Reference Landman, Cochran, Larson, Brezina, Garb and Harries2012, fig. 2) and site photographs (Larson et al., Reference Larson, Brezina, Landman, Garb and Handle2014, fig. 9a, b) have been published. The seep contains a diverse fauna of well-preserved fossils including many ammonites, a nautiloid, bivalves, gastropods, the articulate crinoid Lakotacrinus brezinai Hunter et al., Reference Hunter, Larson, Landman and Oji2016, tube worms, sponges, fish, and crabs (Landman et al., Reference Landman, Cochran, Larson, Brezina, Garb and Harries2012; Larson et al., Reference Larson, Brezina, Landman, Garb and Handle2014).
Diagnosis
Betelgeusid in which ossicles of marginal series are similar, relatively robust, rectangular in outline. Marginal fascioles narrow, deep; intermarginal articular ridges subdued, closely fitted, overlapping flanges not strongly differentiated. First actinal row overlapping onto adambulacrals. First adambulacrals strongly overlapping MAO. Accessories include granules and short, robust spinelets; elongate, delicate spines lacking.
Description
Disk large, interbrachia broadly rounded, arms triangular, tapering abruptly to rounded tips; overall body form low arched in life. Abactinals small but proportionately robust, granular to weakly paxilliform. Abactinal size diminishing distally and radially but otherwise uniform, abactinals aligned in longitudinal and transverse rows; no apparent differentiation of carinal series or primary circlet. Abactinals covered by closely fitted, uniform, small, robust spinelets or granules; abactinals and granules forming closely fitted surface lacking distinct gaps suggestive of respiratory papulae. Madreporite not recognized, although potentially occurring in lost portions of available disks.
Marginal ossicles in two series; series paired, ossicles of both series robust, block-like, rectangular in vertical outline. Inferomarginals protrude slightly laterally beyond margins of superomarginals, more strongly so at interbrachia, protrusion less distinct more distally. Marginal fascioles narrow, well defined, angular. Superomarginal dorsal surface weakly arched longitudinally; transverse profile broadly rounded adradially, more sharply curved toward arm margin. Superomarginal ossicular surfaces closely covered by uniform, closely spaced small granules; small, thickened spinelets can occur at abradial edge, preservational textures suggesting spinelets might not have been developed on all ossicles or all specimens. Both longitudinal and transverse inferomarginal ventral profiles nearly flat, transverse profile curving sharply near abradial margin, abradial inferomarginal edge upright, tightly rounded. Ventral inferomarginal surfaces bearing about five rows of small pustules; accessories entirely granulate, or with scattered spinelets; abradial margin granulate or bearing short, robust spinelets. Elongate spines lacking from both marginal series.
Actinal fields large, two ossicular series aligned with each inferomarginal, although arrangement locally somewhat irregular. Ten to 12 actinals in those inferomarginal series arising at interbrachial midline in specimens R~18–20 mm. Actinal series diminishing uniformly in ossicular number, terminating at about seventh or eighth marginal, at which point inferomarginal series abut adambulacrals. Supplemental actinal series intercalated on disk away from inferomarginals. Actinals of each series aligned, successive series separated by a distinct but shallow groove; actinal grooves approximately, not sharply, aligned with inferomarginal fascioles and tissue gaps between successive adambulacrals. Actinals small, exposed surfaces subcircular, bearing one or more pustules; ossicles strongly overlapping, imbricated toward furrow. Actinal series aligned with interbrachial inferomarginals diverge toward adambulacrals leaving triangular area immediately distal to mouth-angle pair occupied by somewhat irregularly arranged actinals. Actinal ossicular surfaces bearing small spinelets or granules. Actinals adjacent to adambulacrals raised, bearing spinelets differentiated to form part of subadambulacral spinelet series.
Adambulacrals approximately aligned with actinal rows. Adambulacrals broadly rectangular in outline, wider than long; proximal-adradial extremity angled toward mouth area. Tissue gaps between sequential adambulacrals well defined. Furrow accessory ridge V-shaped, directed toward furrow; furrow spine ridge clearly set off from subambulacral spine bases and rows, these two or three in number. Abradial margin of adambulacral overlapped by first actinal. Adambulacral spinelets short, robust. Ambulacrals not exposed in available specimens.
Mouth-angle pair narrow, outline elliptical to weakly ovate, broader end of oval directed toward mouth area. Mouth-angle pair surface broadly arched, bearing spinelets similar to those of adambulacrals. First adambulacrals enlarged, abutting adradial lateral faces of mouth-angle pair, together forming robust, bud-like configuration. Actinal series nearly reaching distal edge of mouth-angle pair.
Etymology
The species is named for Jamie Brezina, who discovered all but one of the available specimens, made them available for research, and donated them to the collections of the American Museum of Natural History.
Remarks
Known species of Betelgeusia are generally similar in overall form, expressions of the abactinal, actinal, and, insofar as is known, ambulacral skeletons, including the mouth frame; they differ largely in marginal ossicular form and accessory development. Betelgeusia brezinai n. sp. is most similar to B. exposita, the latter known from a single incomplete specimen exposed in ventral view. The holotype of B. exposita is not significantly distorted or disrupted, and internal morphology is not available. Marginal ossicles of B. brezinai are more robust and fascioles less sharply defined than in B. exposita; although data are limited, actinals appear to be fewer in B. exposita and the first actinal does not overlap onto the adambulacral; the first adambulacral is proportionately smaller in B. exposita. Superomarginal accessories of B. brezinai are granules rather than spinelets as in B. exposita, and although exposure is limited, superomarginals do not appear to be inset in B. exposita. Marginals and proximal actinals of B. orientalis are similar to those of B. exposita rather than like those of B. brezinai, and MAO pair of B. orientalis are comparatively narrow and keel-like. Overall ossicular form of B. reidi is more delicate than that of B. brezinai, and spines of the former are much elongate and slender whereas those of B. orientalis are intermediate between those of B. reidi and B. exposita.
Superorder Forcipulatacea
Subfamily Pedicellasterinae? Perrier, Reference Perrier1884
Genus indeterminate species indeterminate
Material
Locality AMNH 3529, one small arm fragment, hypotype AMNH-FI 111853.
Remarks
In a traditional arrangement of the Forcipulatacea (Spencer and Wright, Reference Spencer and Wright1966), the Pedicellasterinae is separated from the Asteriinae by absence of an adoral carina (the proximal-most adambulacrals abutting across the interbrachium immediately distal to the mouth-angle pair to form the ‘adoral carina’), and podial arrangement is biserial at least distally, rather than quadriserial throughout. The molecular phylogeny of Mah and Foltz (Reference Mah and Foltz2011b) introduced significant changes to forcipulate classification, the pedicellasterids separating into a more basal cluster, and a six-armed cluster that was imbedded among more derived forcipulates.
In the Pierre Shale fragment, the cross-shaped body wall ossicles with overlapping articular flanges are typical of Forcipulatacea. The shape of the single available ambulacral (Fig. 5.14), exposed in dorsal aspect, documents an arm that had only two rows of tube feet, a generalized configuration nevertheless typical of pedicellasterids in the traditional sense. Adambulacral form is typical of pedicellasterids, although not diagnostic. Familial assignment is traditional; available data are not sufficient for familial assignment following Mah and Foltz (Reference Mah and Foltz2011b) or for generic and species assignments.
Diversification of the Paxillosida
The extant Radiasteridae based on morphologic data and represented by Radiaster is assigned to the order Paxillosida (Blake, Reference Blake1987; Gale, Reference Gale2011); in the literature, the Paxillosida traditionally has been envisioned as a major subdivision of the extant Asteroidea (Spencer and Wright, Reference Spencer and Wright1966), the order including the comparatively familiar fossil and extant Luidia Forbes, Reference Forbes1839 and Astropecten Gray, Reference Gray1840. The molecular analysis of Mah and Foltz (Reference Mah and Foltz2011a) recognized paxillosidan affinities for both the second known extant radiasterid genus, Gephyreaster, and Pseudarchaster Sladen, Reference Sladen1889, the latter traditionally considered to be a goniasterid valvatidan rather than a paxillosidan (Spencer and Wright, Reference Spencer and Wright1966). Drawing on these results, Radiaster here is aligned with Gephyreaster, and together with Pseudarchaster, the three are accepted as a monophyletic paxillosidan cluster. Mah and Foltz (Reference Mah and Foltz2011a) argued for a broadly sister-group status between the Paxillosida plus the Notomyotida (monofamilial, Benthopectinidae), and the bulk of the traditional Valvatida including the Goniasteridae plus many extant members of the Spinulosida sensu Spencer and Wright (Reference Spencer and Wright1966). Goniasterids and Pseudarchaster are widely distributed in modern oceans, and in part because of a comparatively robust construction, goniasterids predominate in the Cretaceous chalk asteroid faunas of Europe.
Although a number of Triassic crown-group asteroids are known, the Paxillosida has not been recorded from pre-Jurassic rocks (Gale, Reference Gale2011, familial summary, p. 57) whereas Hess (Reference Hess1987, p. 907) found that the paxillosidan astropectinids radiated in the Middle Jurassic. Two Jurassic astropectinids have been recognized, the relatively species-rich Pentasteria Valette, Reference Valette1929 and monospecific Advenaster Hess, Reference Hess1955. Pentasteria has been discussed by a number of authors, most notably by H. Hess, whose many papers provide perspective on both this genus and Advenaster (Hess, Reference Hess1955, Reference Hess1960a, Reference Hess1960b, Reference Hess1968, Reference Hess1970, Reference Hess1972, Reference Hess1987; Enay and Hess, Reference Enay and Hess1970). Another Jurassic genus, Archastropecten Hess, Reference Hess1955, was later relegated to a subgeneric status within Pentasteria (Hess, Reference Hess1960a).
Indiaster, sourced from Pachham Island (Kutch, Gujarat, India), was based on two specimens and a fragment thought derived from one of the larger specimens. The specimens unfortunately “are not traceable” and “probably misplaced” (personal communication, D. Srivastava, 2009), and available morphologic data are limited. Published photographs of Rao show both surfaces but only a single surface of each specimen, that in ventral aspect later reproduced by Spencer and Wright (Reference Spencer and Wright1966, fig. 56.2). Five ossicular outline drawings accompanied the original photographs. The specimens appear abraded, limiting potential information. Specimen sizes appear to approximate those of Betelgeusia brezinai. When described, Indiaster was assigned to the Goniasteridae whereas here, following Blake and Reboul (Reference Blake and Reboul2011), it is transferred to the Radiasteridae.
Betelgeusia currently is known only from Cretaceous occurrences. The type species, B. reidi, is represented by many specimens from several formations of the complex middle Cretaceous (Albian, Cenomanian) of Texas; B. exposita is known from a single incomplete Late Cretaceous (Maastrichtian) specimen from Europe; and B. orientalis is known from numerous Early Cretaceous (Barremian) specimens from Morocco.
Overall form and ossicular morphologies of Mesozoic fossils together with those of extant astropectinids and pseudarchasterids (Fisher, Reference Fisher1911, Reference Fisher1919; Hess, Reference Hess1955, Reference Hess1968, Reference Hess1972, Reference Hess1987; Blake and Jagt, Reference Blake and Jagt2005; Blake and Reboul, Reference Blake and Reboul2011) suggest a hypothetical configuration for a primordial paxillosidan species, although no species is considered adequately documented as to serve as an outgroup exemplar for phylogenetic analyses. The posited basal paxillosidan species would have been low-arched with a relatively large disk, broadly curved interbrachia, and triangular, comparatively abruptly tapering arms. A differentiated dorsal midarm series of abactinals together with a differentiated ring or primary circlet of dorsal disk ossicles are typical of most crown-group asteroid clades, but lacking from radiasterids and astropectinids, and generally only weakly differentiated in Pseudarchaster. Interpretation of carinal or primary circlet presence in fossils and even some modern specimens can be difficult because specimen symmetry and sediment compaction can elevate the midarm series as to suggest carinals, and the eye can interpret disk ossicles as concentric in arrangement, suggesting a circlet. Accepting the Paxillosida as a sister group of a part of the terminologically traditional Valvatida (Mah and Foltz, Reference Mah and Foltz2011a), abactinal differentiation was likely plesiomorphic within early paxillosidans, and retained in Pseudarchaster.
In the hypothetical basal paxillosidan, two robust, similar, abutted marginal series would have defined the ambitus and separated an abactinal field of uniform, relatively robust paxilliform abactinals from the more or less enlarged field of actinals, these abutted and aligned in series. Except for B. reidi, available data for ambulacral and adambulacral internal morphology of Mesozoic radiasterids are limited, but adambulacrals likely were rectangular in the basal paxillosidan and bore well-developed spines; granules or small spinelets were common on other ossicles. If pedicellariae were differentiated, blades were simple and spinelet-like.
The strong but generically distinctive marginal differentiation seen in Astropecten and Luidia did not occur in Jurassic Indiaster or Pentasteria whereas complex marginals, ambulacrals, and delicate paxillae suggestive of Astropecten occur in middle Cretaceous B. reidi but not in younger Cretaceous B. brezinai (Blake and Reid, Reference Blake and Reid1998, fig. 8.17; Fig. 5.7 herein), the latter then an inferred more stemward configuration surviving in similar form today not only in Pseudarchaster but also in some astropectinids (Plutonaster Sladen, Reference Sladen1889; Tethyaster Sladen, Reference Sladen1889), goniasterids (Mediaster Stimpson, Reference Stimpson1857; Ceramaster Verrill, Reference Verrill1899), and other valvatidan families (Ganeriidae, Odontasteridae). Expressions of B. reidi might be either basal among those paxillosidans that led to Astropecten and Luidia, or homoplastic. Strongly imbricate actinals of Betelgeusia (Fig. 5.10, 5.11) and Radiaster are not known among the Jurassic paxillosidans.
Expression of the termini of the tube feet might have been emergent within early paxillosidans, but fossilized tube feet are unavailable. Traditionally, termini of the tube feet of all taxa assigned to the Paxillosida are pointed whereas in the remainder of the crown-group Asteroidea, termini are expanded and suckered or disk-like; expressions, however, likely were more complex than the simple two-fold terminology would suggest (Vickery and McClintock, Reference Vickery and McClintock2000; Santos et al., Reference Santos, Haesaerts, Jangoux and Flammang2005). Unlike other paxillosidans, Gephyreaster, Radiaster, and Pseudarchaster have disk-like tube feet. In separating Radiaster (=Mimaster Sladen, Reference Sladen1882) from the astropectinids Plutonaster and Leptychaster Smith, Reference Smith1876, the essential difference in the structure of the tube feet was thought to be “a more important barrier [to combining taxa] than is commonly supposed,” the large sucking disks of Pseudarchaster found to ally this genus with Radiaster and Gephyreaster (Fisher, Reference Fisher1911, p. 175). It was, however, also pointed out, “In passing,” that presence of “conical tube feet with a little button (as in the Astropectinidae) at the tip” occur in Radiaster cognatus (Fisher, Reference Fisher1911, p. 175). However, as discussed in some detail by Clark (Reference Clark1962, p. 11), even familial assignment of this species and genus (under Mimastrella Fisher, Reference Fisher1916) is unclear, Clark including Mimastrella in the Astropectinidae but also finding aspects that “strongly suggest an affinity with the family Solasteridae.” Regardless of ultimate species disposition, complimenting Fisher’s observation on tube feet of ‘R. cognatus,’ occurrence of expanded termini were reported in the early ontogeny of a species of Astropecten (Oguro et al., Reference Oguro, Komatsu and Kano1976). Modern expressions together with the phylogeny of Mah and Foltz (Reference Mah and Foltz2011a) and the complexities noted by Vickery and McClintock (Reference Vickery and McClintock2000) and Santos et al. (Reference Santos, Haesaerts, Jangoux and Flammang2005) suggest an emergent status for tube foot differentiation in early paxillosidans, rather than a single simple ordinal-level apomorphy.
Actinal ossicular alignment immediately adjacent to the mouth frame of B. brezinai appears to have been somewhat irregular, reminiscent of corresponding expressions in both Triassic Trichasteropsis Eck, Reference Eck1879 (Blake and Hagdorn, Reference Blake and Hagdorn2003) and a species of Pycinaster Spencer, Reference Spencer1907 (Blake et al., Reference Blake, Donovan, Mah and Dixon2015). Abundant actinals, uncommon among Paleozoic asteroids, might have arisen as a space-filling device for disk enlargement, the ossicles only later and homoplastically becoming more strongly aligned, perhaps to form efficient water channels or to enhance body flexibility (Blake and Hotchkiss, Reference Blake and Hotchkiss2004).
Taxonomic changes
A new asteroid species, Betelgeusia brezinai (Radiasteridae), is described; four species of Betelgeusia, all Cretaceous in age, now are known. A second radiasterid genus, Indiaster, is Jurassic in age. Fossil radiasterids have been recorded from Africa, western Europe, India, and North America. A single fragment is assigned to the forcipulate Pedicellasterinae. Data are insufficient for formal phylogenetic analysis but enough is known to reconstruct many likely aspects of morphology of the ancestral species of Paxillosida.
Acknowledgments
J. Brezina collected the bulk of the specimens, advised on the geology, and donated the specimens to the American Museum of Natural History. The Maude family permitted property access. All localities were discovered and mapped by J. Brezina working with The American Museum of Natural History and other research institutions as part of a hydrocarbon seep mapping project. N. Landman, M. Garb, K. Cochran, and numerous students assisted in the field. N. Landman obtained necessary permits for collecting on public grasslands. C. Mah and J. Jagt provided valuable manuscript reviews, and we are indebted to J. Jin and S. Zamora for editorial work.





