


1. Introduction
The Plesiosauria are a group of large marine predators known from latest Triassic to the end of Late Cretaceous time, when they become extinct during the biological crisis of the Cretaceous–Palaeogene (K–Pg) boundary (e.g. Vincent et al. Reference Vincent, Bardet, Pereda Suberbiola, Bouya, Amaghzaz and Meslouh2011). Soon after their first appearance in the fossil record at the end of Triassic time, they underwent a spectacular radiation and became widespread both in time and space, being highly systematically and ecologically diversified throughout the Jurassic–Cretaceous periods and found in all continents (Ketchum & Benson, Reference Ketchum and Benson2010).
During Late Cretaceous time the Plesiosauria were particularly diversified and mainly represented by Plesiosauroidea, known by the three families Elasmosauridae, Polycotylidae and Leptocleididae, as well as by the declining Pliosauroidea, known by the Pliosauridae only (Ketchum & Benson, Reference Ketchum and Benson2010). Cretaceous pliosaurs are only represented by: Kronosaurus Longman, Reference Longman1924 from Aptian–Albian Australia and Colombia; Brachauchenius Williston, Reference Williston1903 from the Turonian Western Interior Seaway and Barremian Colombia (Hampe, Reference Hampe2005); and Megacephalosaurus eulerti Schumacher, Carpenter & Everhart, Reference Schumacher, Carpenter and Everhart2013 (FHSM VP321) and Schumacher, Reference Schumacher2008 (paratype UNSM 50136) from Turonian Kansas (see Ketchum & Benson, Reference Ketchum and Benson2010 for details, and online Supplementary Material Table S3 available at http://journals.cambridge.org/geo).
Compared to other continents, the fossil record of plesiosaurs is scarce in Africa where only five valid taxa are known. All described from Cretaceous outcrops, these are: the leptocleidid Leptocleidus capensis (Andrews, Reference Andrews1911) from Early Cretaceous (Valanginian) South Africa (Andrews, Reference Andrews1911; Cruickshank, Reference Cruickshank1997); the elasmosaurid Zarafasaura oceanis Vincent et al. Reference Vincent, Bardet, Pereda Suberbiola, Bouya, Amaghzaz and Meslouh2011 from latest Cretaceous (Maastrichtian) Morocco (Vincent et al. Reference Vincent, Bardet, Pereda Suberbiola, Bouya, Amaghzaz and Meslouh2011); and the polycotylids Thililua longicollis Bardet, Suberbiola & Jalil, Reference Bardet, Suberbiola and Jalil2003a and Manemergus anguirostris Buchy, Metayer & Frey, Reference Buchy, Metayer and Frey2005 and the elasmosaurid Libonectes atlasense Buchy, Reference Buchy2005 described from the lower Upper Cretaceous (Turonian) sediments of the Goulmima outcrop, southern Morocco (Bardet, Suberbiola & Jalil, Reference Bardet, Suberbiola and Jalil2003a ; Buchy, Reference Buchy2005; Buchy, Metayer & Frey, Reference Buchy, Metayer and Frey2005).
The Goulmima outcrop today represents the richest plesiosaur site from Africa, with the two major clades of Cretaceous plesiosaurs represented as well as the elasmosaurids and the polycotylids. Here we describe a new plesiosaur specimen from this outcrop belonging to the Pliosauridae, a clade previously undescribed from this area.
Abbreviations: FHSM – The Sternberg Museum of Natural History, Hays, Kansas (USA); MNHN – Muséum National d’Histoire Naturelle, Paris (France); UNSM – University of Nebraska State Museum, Lincoln, Nebraska, USA.
2. Geographical and geological context
In North Africa, the ‘middle’ part of the Cretaceous (Aptian–Turonian) deposits is represented by a classical trilogy forming in the landscape the recognizable cliff ‘Hamada’ that crops out extensively SW–NE from the Gulf of Agadir (Morocco) to the Gulf of Gabès (Tunisia) (Choubert & Faure-Muret, Reference Choubert and Faure-Muret1962).
The series classically includes, from the base to the top: (1) red sandstones with crossed stratifications (Ifezouane Formation); (2) lagunar gypsous marly sandstones and green marls (Aoufous Formation) (these two units are grouped into the classical ‘Continental Intercalaire’ of the French authors or Kem Kem beds, Aptian–Albian in age); and (3) a marine calcareous massive bar (Akrabou Formation), representing the beginning of the Cenomanian–Turonian transgression in the area.
The specimen MNHN GOU 11 comes from the Goulmima region, located in the southern slope of the High Atlas of Morocco (Fig. 1a). Although its exact location remains unknown because this specimen was purchased, fieldwork has allowed us to locate several fossiliferous localities near the villages of Tadighourst and Asfla, north of Goulmima (Er-Rachidia Province), where the specimen could potentially come from (personal observation by NB).

Figure 1. (a) Palaeogeographical location of the Goulmima area in southern Morocco (modified from Bardet, Suberbiola & Jalil, Reference Bardet, Suberbiola and Jalil2003a ). (b) Probable stratigraphical range (Mammites horizon) of Brachauchenius lucasi specimen MNHN GOU 11 (modified from Cavin et al. Reference Cavin, Tong, Boudad, Meister, Piuz, Tabouelle, Aarab, Amiot, Buffetaut, Dyke, Hua and Le Loeuff2010).
In the Goulmima area, the Cenomanian–Turonian limestone bar is easily identified. It is famous for its Turonian nodulous fossiliferous levels, including ammonites as well as marine vertebrates such as bony fishes (Cavin, Reference Cavin2001; Cavin et al. Reference Cavin, Tong, Boudad, Meister, Piuz, Tabouelle, Aarab, Amiot, Buffetaut, Dyke, Hua and Le Loeuff2010) and large marine reptiles, including the basal mosasauroid Tethysaurus Bardet, Suberbiola & Jalil, Reference Bardet, Suberbiola and Jalil2003b and the three plesiosaur taxa mentioned in the previous section (see Bardet et al. Reference Bardet, Houssaye, Rage and Pereda Suberbiola2008 for details). Specimen MNHN GOU 11 was probably preserved in such one large nodule, although it remains impossible to confirm as it was purchased already prepared (NB, personal observation).
The fossils are generally very well preserved in small to very large concretion nodules that mainly occur near the top of a Cenomanian–Turonian limestone bar (Fig. 1b). The main fossiliferous levels are located in the ‘Unit 4’ (lower Turonian) of Ferrandini et al. (Reference Ferrandini, Philip, Babinot and Tronchetti1985), now reappraised by Ettachfini & Andreu (Reference Ettachfini and Andreu2004) as the ‘Unit T2a’ of the Akrabou Formation (middle Turonian). Ammonites are rather frequent in these levels, especially belonging to the genus Mammites (see Cavin et al. Reference Cavin, Tong, Boudad, Meister, Piuz, Tabouelle, Aarab, Amiot, Buffetaut, Dyke, Hua and Le Loeuff2010 for details).
The Goulmima area was the centre of a basin which underwent large subsidence during the Cenomanian–Turonian transgression. It corresponds to an open marine carbonated platform (Fig. 1) related to the maximum of the Cenomanian–Turonian transgressive phase (Ferrandini et al. Reference Ferrandini, Philip, Babinot and Tronchetti1985), with dominant Tethyan but also Central Atlantic influences (Ettachfini & Andreu, Reference Ettachfini and Andreu2004; Cavin et al. Reference Cavin, Tong, Boudad, Meister, Piuz, Tabouelle, Aarab, Amiot, Buffetaut, Dyke, Hua and Le Loeuff2010).
3. Systematic palaeontology
Plesiosauria de Blainville, Reference Blainville de1835
Pliosauroidea (Seeley, Reference Seeley1874) Welles, Reference Welles1943
Pliosauroidea Seeley, Reference Seeley1874
Brachauchenius Williston, Reference Williston1903
Brachauchenius lucasi Williston, Reference Williston1903
Holotype: USNM 4989, skull, mandible, vertebrae and ribs; Ottawa County, Kansas (USA); Greenhorn Limestone, Turonian (Schumacher & Everhart, Reference Schumacher and Everhart2005).
Referred specimen: MNHN GOU 11, an incomplete mandible; Goulmima region, Er-Rachidia province (Morocco); probably Akrabou Formation (NB, personal observation), middle Turonian (Ettachfini & Andreu, Reference Ettachfini and Andreu2004).
4. Description
4.a. Mandible
The mandible preserved only both dentaries, broken into 18 parts but remaining in connection. The right ramus is more complete, but the left is only partially preserved. No suture is visible and it is impossible to say if the splenial was preserved. The anterior part of the mandible is large and robust, measuring 945mm in length (Table 1). This indicates that the whole mandible measured roughly 1500mm in total length.
Table 1. Measurements of the different elements of Brachauchenius lucasi specimen MNHN GOU 11. NA – not applicable.

The symphysis is of medium size, being 250mm length dorsally and 145mm ventrally (Figs 2, 3; online Supplementary Material Video S1 available at http://journals.cambridge.org/geo), which corresponds to 26% of the preserved dentary. The maximum ramus height is 120mm on the posterior part of the ramus and decreases slowly forward, being 68mm high just before the symphysis. No diastema is visible between the last symphysal alveolus and the next.

Figure 2. (a) Photograph, with the localization and detail of the three teeth (1–3); (b) 3D reconstruction; and (c) interpretative drawing of Brachauchenius lucasi specimen MNHN GOU 11 in dorsal view, with the numeration of the different elements measured.
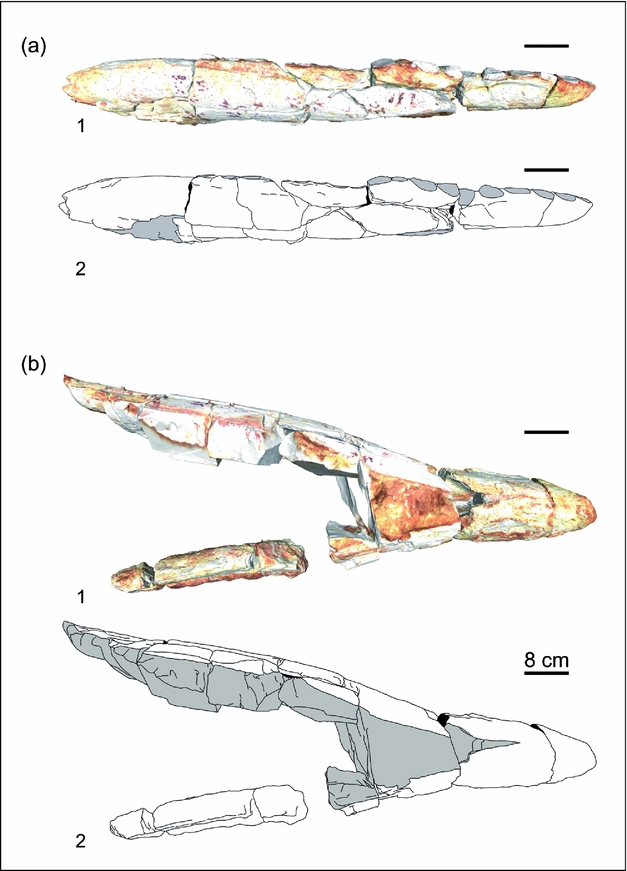
Figure 3. (a) 3D reconstruction and (b) interpretative drawing of Brachauchenius lucasi specimen MNHN GOU 11 in (1) lateral and (2) ventral views.
The right dentary bears 24 alveoli, 6 of which are located within the symphysis. All alveoli have roughly the same diameter, averaging about 32mm × 35mm (Table 1). The alveoli are lingually bordered by a rim that extends anteriorly up to the symphysis and that bears the alveoli for the replacement teeth. As in all other plesiosaurs, the replacement teeth are located postero-lingually to the primary alveoli.
4.b. Dentition
No tooth is complete, but several are partially preserved. In the alveolus 9 of the right dentary ramus, the root of a mature tooth is eroded and broken at the base of the crown; it is 25mm × 35mm in diameter, which permits the maximum width of mature tooth roots of this specimen to be estimated (Fig. 2).
Two replacement teeth are preserved on alveoli 2 and 7 of the right ramus (Fig. 2). The first is very small (5mm length) and still located in the replacement alveolus, while the 7th alveolus is largest, of length 25mm. It has a rounded cross-section and the enamel is ornamented with heavily striae all around and some branching striae toward the base of the crown. The teeth are large, posteriorly recurved rounded cones. Thanks to the various longitudinal dentary fractures, we can observe that the alveoli are relatively deep (50mm). The symphysal teeth are more closely spaced than in the remainder of the ramus, with an interval of 2–3mm versus 5mm.
5. Discussion
5.a. Systematical attribution
The specimen MNHN GOU 11 has been compared with a selection of Plesiosauria for which the mandible is preserved. The most recent works on the subject, mainly O’Keefe (Reference O’Keefe2001), Smith & Vincent (Reference Smith and Vincent2010), Ketchum & Benson (Reference Ketchum and Benson2010, Reference Ketchum and Benson2011a , Reference Ketchum and Benson b ), Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) and Schumacher, Carpenter & Everhart (Reference Schumacher, Carpenter and Everhart2013) have been used for this comparative purpose (online Supplementary Material Tables S1 and S2, available at http://journals.cambridge.org/geo.)
Because of its long mandible, MNHN GOU 11 could belong either to Pliosauroidea or to polycotylid Plesiosauroidea and can reasonably be excluded from other Plesiosauroidea (Plesiosauridae, Elasmosauridae, Leptocleididae) which all possess a small and gracile head as well as short mandibular symphysis. As polycotylids have already been described from Turonian deposits of Goulmima (Bardet, Suberbiola & Jalil, Reference Bardet, Suberbiola and Jalil2003a ; Buchy, Metayer & Frey, Reference Buchy, Metayer and Frey2005), the possibility that MNHN GOU 11 could belong to this clade cannot be excluded and the specimen has been compared to taxa of both clades.
The Polycotylidae have a long and tapered mandible like some Pliosauridae (e.g. Marmornectes), but their symphysis is generally more elongated and more slender than that of pliosauroids in general and of MNHN GOU 11 in particular (Figs 2–4). Moreover, polycotylids bear more slender teeth (Bardet, Suberbiola & Jalil, Reference Bardet, Suberbiola and Jalil2003a ), even on taxa with robust teeth such as Eopolycotulus (Albright III, Gillette & Titus, Reference Albright III, Gillette and Titus2007b ) or Dolichorhynchops (O’Keefe, Reference O’Keefe2008), that differ considerably from the large robust teeth of MNHN GOU 11. Mandibular symphysis of Polycotylidae usually encompasses at least 12 (Pahasapasaurus) and up to 20 alveoli (Dolichorhynchops), much longer and more slender than the symphysis of MNHN GOU 11. Only 9 symphyseal tooth positions are known in Manemergus, but again it is much more slender and elongate and never exhibits the slight spoon-shaped inflation of the MNHN GOU 11 symphysis.

Figure 4. Comparison between (a) Goulmima specimen MNHN GOU 11 and a selection of (b) Rhomaleosauridae, (c) Polycotylidae and (d) Pliosauridae for which the mandible is well known. (a) Drawing of Brachauchenius lucasi specimen MNHN GOU 11 in (upper) dorsal view and (lower) ventral view. (b) Drawing of Rhomaleosaurus cramptoni in (upper, from Cruickshank, Reference Cruickshank1996) dorsal view and (lower, from Smith & Dyke, Reference Smith and Dyke2008) ventral view. (c) Drawing of Dolichorhynchops herschelensis in (upper) ventral view and (lower) dorsal view (from Sato, Reference Sato2005). (d) Gallardosaurus itturraldei drawing in ventral view (from Gasparini, Reference Gasparini2009). Polyptychodon hudsoni photo in dorsal view (from Welles & Slaughter, Reference Welles and Slaughter1963). Brachauchenius lucasi photo in dorsal view (from Albright III, Gillette & Titus, Reference Albright, Gillette and Titus2007a ). Photographs of Simolestes vorax in (upper, from Andrews, Reference Andrews1913) ventral view and (lower, from Tarlo, Reference Tarlo1960) dorsal view. Photograph of Liopleurodon ferox in (upper) dorsal view and (lower) ventral view (Andrews, Reference Andrews1913). Pliosaurus brachydeirus drawing in dorsal view (from Noè, Smith & Walton, Reference Noè, Smith and Walton2004). Peloneustes philarchus photograph (upper, from Tarlo, Reference Tarlo1960) in dorsal view and outline (lower, from Lydekker, Reference Lydekker1889) in dorsal view. Scale bars: 10cm.
The Pliosauroidea have large head and mandible, massive toothed jaws and elongated mandibular symphysis (O’Keefe, Reference O’Keefe2001), all characters that fit rather well with the mandible from Morocco. On this basis, MNHN GOU 11 is therefore referred to the Pliosauroid clade.
Pliosauroids include two families, the Pliosauridae (Seeley, Reference Seeley1874) and the Rhomaleosauridae (Nopsca, Reference Nopsca1928; Ketchum & Benson, Reference Ketchum and Benson2011a ).
Rhomaleosauridae were unearthed from Upper Triassic – Middle Jurassic rocks of Western Europe, Argentina and Canada (online Supplementary Material Tables S2 and S3, available at http://journals.cambridge.org/geo). This family is characterized by a short, reinforced and keeled spatulate symphysis (Smith & Vincent, Reference Smith and Vincent2010; Fig. 4), like that of the pliosaurid Simolestes Andrews, Reference Andrews1909 from the Jurassic rocks of Europe and India (see Godefroit, Reference Godefroit1994). This special mandible morphology tip is therefore quite different from MNHN GOU 11.
Pliosauridae comprise at least 11 genera (see Ketchum & Benson, Reference Ketchum and Benson2011a ) of which the mandible morphology is known (or can be partially deduced by the skull morphology) in Thalassiodracon Storrs & Taylor, Reference Storrs and Taylor1996 from the lower Lower Jurassic deposits of England; Hauffiosaurus O’Keefe, Reference O’Keefe2001 from the upper Lower Jurassic rocks of Germany; Pliosaurus Owen, Reference Owen1841, Liopleurodon Sauvage, Reference Sauvage1873, Peloneustes Lydekker, Reference Lydekker1889, Simolestes Andrews, Reference Andrews1909 and Marmonectes Ketchum & Benson, Reference Ketchum and Benson2011a from the Middle–Upper Jurassic rocks of mainly western Europe; Polyptychodon Owen, Reference Owen1841 from the lower Upper Cretaceous deposits of England and Texas; Megacephalosaurus Schumacher, Carpenter & Everhart, Reference Schumacher, Carpenter and Everhart2013 from the middle Turionan rocks of Kansas; Kronosaurus Longman, Reference Longman1924 from the upper Lower Cretaceous deposits of Australia and Colombia and finally Brachauchenius Williston, Reference Williston1903, from the lower Upper Cretaceous unit of the Western Interior Way.
Simolestes has a short and spatulate symphysis very different from that of MNHN GOU 11. The same occurs with the very short and plesiosauroid-like mandible of the basal Thalassiodracon. On the contrary, Hauffiosaurus, Peloneustes and Marmonectes have a very long and slender mandibular symphysis including around 12 pairs of teeth (Ketchum & Benson, Reference Ketchum and Benson2011b ), therefore very different from MNHN GOU 11. Liopleurodon has a robust and moderately long mandibular symphysis like that of the specimen from Morocco; however, it exhibits distinct heterodonty (Tarlo, Reference Tarlo1960; Noè, Smith & Walton, Reference Noè, Smith and Walton2004) unlike that observed in MNHN GOU 11, which exhibits alveoli roughly similar in size. As far as Pliosaurus is concerned, P. macromerus (Phillips, Reference Phillips1871), P. brachyspondylus (Owen, Reference Owen1841) and P. brachydeirus (Owen, Reference Owen1841) bear teeth with trihedral or sub-trihedral cross-sections (Tarlo, Reference Tarlo1960; Ketchum & Benson, Reference Ketchum and Benson2011a ; Knusten, Reference Knusten2012), unlike the circular section of the teeth of the Morocco specimen. All the other species of Pliosaurus – P. portentificus Noè, Smith & Walton, Reference Noè, Smith and Walton2004; P. andrewsi Tarlo, Reference Tarlo1960; P. kevani Benson et al. Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013; P. carpenteri Benson et al. Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013; and P. westburyensis Benson et al. Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013 (Tarlo, Reference Tarlo1960; Noè, Smith & Walton, Reference Noè, Smith and Walton2004; Benson et al. Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) – present a number of symphyseal teeth (8–15 teeth) larger than that observed on the Goulmina specimen (6 teeth). Kronosaurus shows only 3–4 symphysal teeth (Longman, Reference Longman1924; Druckenmiller, Reference Druckenmiller2006), compared to the 6 found in MNHN GOU 11. The new genus Megacephalosaurus described by Schumacher, Carpenter & Everhart (Reference Schumacher, Carpenter and Everhart2013) is very similar to the specimen from Morocco, but its size is significantly larger (50%).
Finally, MNHN GOU 11 presents similarities to both Polyptychodon Owen, Reference Owen1841 and Brachauchenius Williston, Reference Williston1903. The systematical situation of Polyptychodon remains problematic (see following section) and direct comparisons cannot be made with this taxon. Brachauchenius includes only the species B. lucasi Williston, Reference Williston1903, described from Turonian rocks of the Western Interior Seaway of North America and upper Barremian deposits of northern South America (Colombia) (Hampe, Reference Hampe2005). It is characterized by 21 (Carpenter, Reference Carpenter1996) to 25 (Albright III, Gillette & Titus, Reference Albright, Gillette and Titus2007a ) dentary teeth. Teeth are regular in size along the series and are robust and heavily striated all around, with some branching of the striation toward the base of the crown (Albright III, Gillette & Titus, Reference Albright, Gillette and Titus2007a ). This combination of characters is comparable with that observed in MNHN GOU 11. The only minor difference observed is that the alveoli of MNHN GOU 11 occupy almost the total dentary width, a character not reported in any other taxon. However, as the external surface of MNHN GOU 11 bones is somewhat eroded, this could partly be a preservational bias.
Based on the characters described above, the specimen MNHN GOU 11 is referred to the pliosaurid genus Brachauchenius and, this genus being represented by only one species, de facto to Brachauchenius lucasi Williston, Reference Williston1903.
5.b. Synonymy and the taxonomic problem of Polyptychodon Owen, Reference Owen1841
This problem has already been noted by other authors (e.g. Albright III, Gillette & Titus, Reference Albright, Gillette and Titus2007a ; Schumacher, Reference Schumacher2008) and has a special importance in our work because Polyptychodon is described for outcrops of the same age as our specimen. It is therefore very important to understand better this taxon, in particular to compare it correctly with other Plesiosauria.
Polyptychodon was described for the first time by Owen in 1841 in his Odontography from specimens from the Greensands of England. At that time, he proposed two new species P. interruptus and P. continuus. P. interruptus is based on a unique isolated tooth and another one included in a mandibular fragment, to which a skull fragment was later referred (Owen, Reference Owen1851, pl. 4, fig. 1). P. continuus is based on few isolated teeth and isolated and fragmentary postcranial material. Among all this material, one tooth was chosen as the type; the reason for this choice remains unclear and is not explained by Owen. More than one century later Welles & Slaughter (Reference Welles and Slaughter1963) described a new species of Polyptychodon, P. hudsoni, from Turonian deposits of Texas (USA) on the basis of an incomplete skull and mandible and isolated teeth, as well as (possibly) a caudal vertebra found later at the same place. These three species are therefore known from Cretaceous (Aptian–Santonian) Europe for P. continuus and P. interuptus (Owen, Reference Owen1861; Persson, Reference Persson1963; Welles & Slaughter, Reference Welles and Slaughter1963; Papazzoni, Reference Papazzoni2003) and Turonian North America for P. hudsoni. It should be noted that P. interruptus has also been found in Asia (Sepkoski, Reference Sepkoski2002). Teeth bearing a “Polyptychodon morphology” are common in the Albian–Cenomanian outcrops of Europe (personal observation by NB).
Different problems arise from this genus. Firstly, as noted by Welles & Slaughter (Reference Welles and Slaughter1963) and Schumacher (Reference Schumacher2008), it is based on very poor material, mainly isolated teeth. Moreover, these teeth are non-diagnostic as compared to some other pliosaurs, being large cones with circular cross-section and striated enamel which are all characteristics applying to several pliosaur taxa. Welles & Slaughter (Reference Welles and Slaughter1963) proposed the designation of the skull fragment referred to P. interruptus by Owen (Reference Owen1851) as the new type of this species. As for most nineteenth-century palaeontological descriptions, Owen's publications (Reference Owen1841, Reference Owen1851) were very ambiguous as far as the designation of the holotype and referred specimens are concerned. To the isolated teeth originally described (also not necessarily from the same individual), he associated several bone elements without any homologous elements of comparison; his only justification was that they had the same stratigraphical origin. At this time, when only Polyptychodon had been described from Upper Cretaceous deposits of England, this reasoning seemed logical (another common situation in nineteenth-century palaeontological descriptions).
Another problem arises from the proposition of the new species P. hudsoni by Welles & Slaughter (Reference Welles and Slaughter1963). As noted by Albright III, Gillette & Titus (Reference Albright, Gillette and Titus2007a ), Welles & Slaughter described the new specimen from Texas without comparing it to pliosaurid taxa already known from Cretaceous rocks of North American, especially Brachauchenius lucasi. Further, Williston (Reference Williston1907) referred to this species a specimen (USNM 2361, see Schumacher, Carpenter & Everhart, Reference Schumacher, Carpenter and Everhart2013) recovered from the same stratigraphical unit as their P. hudsoni specimen. It is noteworthy that P. hudsoni resembles B. lucasi in having teeth with branching striations, but differs from the holotype of Polyptychodon, P. interruptus, which lacks branching striations.
Two possibilities can therefore be envisaged for Polyptychodon as follows.
-
1) Polyptychodon is a nomen dubium because is it not based on any diagnostical material. In this case, the systematical position of P. hudsoni should be revised and this species, if valid, should be referred to another genus (possibly Brachauchenius, or to a new genus). It is interesting to note that in their recent phylogenetic work, Benson et al. (Reference Benson, Evans, Smith, Sassoon, Moore-Faye, Ketchum and Forrest2013) refer at least some specimens originally described as Polyptychodon to Brachauchenius.
-
2) Polyptychodon is a valid genus. In this case, as noted by Schumacher (Reference Schumacher2008), a paratype bearing diagnostic characters, for example the skull fragment mentioned by Owen (Reference Owen1851), must be defined. It should be adequate to prove that isolated specimens actually belong to the same species, and to permit the erection of a new and more rigorous diagnosis. Pending the discovery of new and more complete specimens, this is difficult in the current state of knowledge of this taxon.
5.c. Palaeobiogeography and palaeoecological interpretations
Brachauchenius lucasi was previously known only from several Cenomanian–Turonian outcrops from the Western Interior Seaway of North America (Williston, Reference Williston1907; Carpenter, Reference Carpenter1996; Hampe, Reference Hampe2005; Liggett et al. Reference Liggett, Shimada, Bennett and Schumacher2005; Albright III, Gillette & Titus, Reference Albright, Gillette and Titus2007a ) and from upper Barremian deposits of northern South America (Colombia) (Hampe, Reference Hampe2005). The description of a new specimen of Brachauchenius lucasi in the Turonian outcrops of Morocco therefore allows the greatly enlarged palaeobiogeographical distribution of this taxon to North Africa (Fig. 5). Moreover, some affinities between North American and North African faunas have been already noted for other vertebrate taxa, more specifically teleostean fishes (Cavin et al. Reference Cavin, Tong, Boudad, Meister, Piuz, Tabouelle, Aarab, Amiot, Buffetaut, Dyke, Hua and Le Loeuff2010).

Figure 5. Palaeobiogeographical map of the Turonian distribution of the Plesiosauria.
The sedimentary unit in which MNHN GOU 11 was likely fossilized corresponds to a shallow marine environment related to the maximum of the Cenomanian–Turonian transgressive phase (e.g. Cavin, Reference Cavin1999). The faunal relationships between the long distant Western Interior Seaway and the Goulmima Basin could have been due to the occurrence of W–E-aligned currents within the Tethys during early Late Cretaceous time. These currents probably facilitated the dispersion of other contemporary marine faunas, such as the teleostean fish, but also other marine reptiles such as squamates (Caldwell & Cooper, Reference Caldwell and Cooper1999; Bardet et al. Reference Bardet, Houssaye, Rage and Pereda Suberbiola2008).
The Turonian Goulmima outcrop is well known for its high diversity of marine vertebrates, including teleostean fish (Cavin, Reference Cavin1997, Reference Cavin1999, Reference Cavin2001) and marine reptiles. The latter are represented by two major clades, the Plesiosauria and the Squamata (Bardet, Suberbiola & Jalil, Reference Bardet, Suberbiola and Jalil2003a , Reference Bardet, Suberbiola and Jalil b ; Buchy, Reference Buchy2005; Buchy, Metayer & Frey, Reference Buchy, Metayer and Frey2005). The Plesiosauria are the most diverse and up until now were represented by three different Plesiosauroid taxa, two polycotylids (Thililua and Manemergus) and an elasmosaurid (Libonectes morgani). The description of this specimen belonging to the pliosaurid Brachauchenuis lucasi allows the faunal list of the Turonian Goulmima outcrops to be improved. Regarding the small to medium size of all the other taxa, Brachauchenuis lucasi was probably the top predator of the area.
Acknowledgements
NB thanks François Escuillié (Eldonia, Gannat, France) for the donation of the specimen and for providing the geographical and stratigraphical occurrence data which were acquired during fieldwork in the Goulmima area in 2001. We also thank Florent Goussard for help with the 3C treatment of the specimen and Renaud Vacant for assistance with the preparation the specimen.
Declaration of interest
None
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0016756815000321








