











Crossref Citations
This article has been cited by the following publications. This list is generated based on data provided by Crossref.
Li, An-Xing
Wu, Xiang-Yun
Xie, Ming-Quan
Lin, Rui-Qing
Gasser, Robin B.
and
Zhu, Xing-Quan
2005.
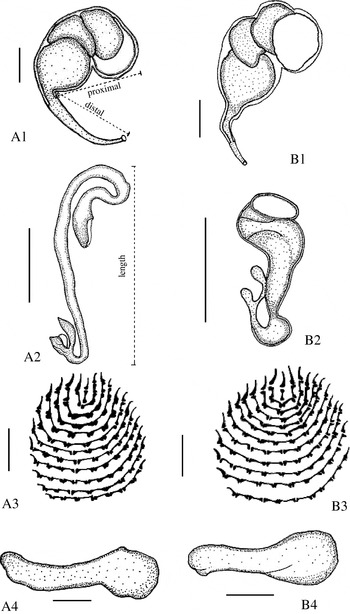
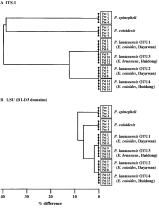
PCR-based identification and delineation of members within the Pseudorhabdosynochus lantauensis complex (Monogenea: Diplectanidae).
Parasitology Research,
Vol. 98,
Issue. 1,
p.
34.
VILAS, R.
CRISCIONE, C. D.
and
BLOUIN, M. S.
2005.
A comparison between mitochondrial DNA and the ribosomal internal transcribed regions in prospecting for cryptic species of platyhelminth parasites.
Parasitology,
Vol. 131,
Issue. 06,
p.
839.
WU, X. Y.
ZHU, X. Q.
XIE, M. Q.
and
LI, A. X.
2006.
The radiation of Haliotrema (Monogenea: Dactylogyridae: Ancyrocephalinae): molecular evidence and explanation inferred from LSU rDNA sequences.
Parasitology,
Vol. 132,
Issue. 05,
Sun, H. Y.
Zhu, X. Q.
Xie, M. Q.
Wu, X. Y.
Li, A. X.
Lin, R. Q.
and
Song, H. Q.
2006.
Characterization of Cryptocaryon irritans isolates from marine fishes in Mainland China by ITS ribosomal DNA sequences.
Parasitology Research,
Vol. 99,
Issue. 2,
p.
160.
Wu, X. Y.
Zhu, X. Q.
Xie, M. Q.
Wang, J. Q.
and
Li, A. X.
2007.
The radiation of Thaparocleidus (Monogenoidea: Dactylogyridae: Ancylodiscoidinae): phylogenetic analyses and taxonomic implications inferred from ribosomal DNA sequences.
Parasitology Research,
Vol. 102,
Issue. 2,
p.
283.
Wu, Xiang-Yun
Zhu, Xing-Quan
Xie, Ming-Quan
and
Li, An-Xing
2007.
The evaluation for generic-level monophyly of Ancyrocephalinae (Monogenea, Dactylogyridae) using ribosomal DNA sequence data.
Molecular Phylogenetics and Evolution,
Vol. 44,
Issue. 2,
p.
530.
Waeschenbach, Andrea
Webster, Bonnie L.
Bray, Rodney A.
and
Littlewood, D.T.J.
2007.
Added resolution among ordinal level relationships of tapeworms (Platyhelminthes: Cestoda) with complete small and large subunit nuclear ribosomal RNA genes.
Molecular Phylogenetics and Evolution,
Vol. 45,
Issue. 1,
p.
311.
Lin, Rui-Qing
Zhu, Xing-Quan
Wei, Dong-Xia
Deng, Yan
Liu, Wei
Song, Hui-Qun
Li, An-Xing
and
Lun, Zhao-Rong
2007.
Characterization of Oesophagostomum spp. from pigs in China by PCR-based approaches using genetic markers in the internal transcribed spacers of ribosomal DNA.
Parasitology Research,
Vol. 101,
Issue. 2,
p.
351.
Schoelinck, Charlotte
and
Justine, Jean-Lou
2011.
Four species of Pseudorhabdosynochus (Monogenea: Diplectanidae) from the camouflage grouper Epinephelus polyphekadion (Perciformes: Serranidae) off New Caledonia.
Systematic Parasitology,
Vol. 79,
Issue. 1,
p.
41.
Blasco-Costa, Isabel
Míguez-Lozano, Raúl
Sarabeev, Volodimir
and
Balbuena, Juan Antonio
2012.
Molecular phylogeny of species of Ligophorus (Monogenea: Dactylogyridae) and their affinities within the Dactylogyridae.
Parasitology International,
Vol. 61,
Issue. 4,
p.
619.
Wang, C.R.
Gao, J.F.
Zhu, X.Q.
and
Zhao, Q.
2012.
Characterization of Bunostomum trigonocephalum and Bunostomum phlebotomum from sheep and cattle by internal transcribed spacers of nuclear ribosomal DNA.
Research in Veterinary Science,
Vol. 92,
Issue. 1,
p.
99.
Yoon, Gil Ha
Al-Jufaili, Sarah
Freeman, Mark A
Bron, James E
Paladini, Giuseppe
and
Shinn, Andrew P
2013.
Omanicotyle heterospina n. gen. et n. comb. (Monogenea: Microcotylidae) from the gills of Argyrops spinifer (Forsskål) (Teleostei: Sparidae) from the Sea of Oman.
Parasites & Vectors,
Vol. 6,
Issue. 1,
Chaudhary, A.
and
Singh, H.S.
2013.
Description of two new species of the genus Thaparocleidus Jain, 1952 (Monogenea, Dactylogyridae) from freshwater fish in India: morphological and molecular phylogenetic evidence.
Journal of Helminthology,
Vol. 87,
Issue. 2,
p.
160.
Patrelle, Cécile
Ferté, Hubert
and
Jouet, Damien
2014.
Identification of Chabertiidae (Nematoda, Strongylida) by PCR-RFLP based method: A new diagnostic tool for cross transmission investigation between domestic and wild ruminants in France.
Infection, Genetics and Evolution,
Vol. 28,
Issue. ,
p.
15.
Chaudhary, Anshu
Verma, Chandni
and
Singh, Hridaya Shanker
2014.
Ribosomal DNA as molecular markers and their applications in the identification of fish parasites (Platyhelminthes: Monogenea) from India.
Asian Pacific Journal of Tropical Disease,
Vol. 4,
Issue. 1,
p.
82.
Sarabeev, Volodimir
and
Desdevises, Yves
2014.
Phylogeny of the Atlantic and Pacific species of Ligophorus (Monogenea: Dactylogyridae): Morphology vs. molecules.
Parasitology International,
Vol. 63,
Issue. 1,
p.
9.
Messu Mandeng, Françoise D.
Bilong Bilong, Charles F.
Pariselle, Antoine
Vanhove, Maarten P. M.
Bitja Nyom, Arnold R.
and
Agnèse, Jean-François
2015.
A phylogeny of Cichlidogyrus spp. (Monogenea, Dactylogyridea) clarifies a host-switch between fish families and reveals an adaptive component to attachment organ morphology of this parasite genus.
Parasites & Vectors,
Vol. 8,
Issue. 1,
Chotnipat, S
Miller, TL
Knuckey, RM
and
Hutson, KS
2015.
Molecular and morphological evidence for the widespread distribution of Laticola paralatesi infecting wild and farmed Lates calcarifer in Australia.
Diseases of Aquatic Organisms,
Vol. 113,
Issue. 3,
p.
195.
Igeh, Patience C
Dos Santos, Quinton M
and
Avenant-Oldewage, Annemariè
2017.
Redescription ofCichlidogyrus philander(Monogenea, Ancyrocephalidae) using scanning electron microscopy (SEM) and molecular analysis.
Parasite,
Vol. 24,
Issue. ,
p.
49.
Dmitrieva, Evgenija V.
Sanna, Daria
Piras, M. Cristina
Garippa, Giovanni
and
Merella, Paolo
2018.
Xenoligophoroides cobitis (Ergens, 1963) n. g., n. comb. (Monogenea: Ancyrocephalidae), a parasite of Gobius cobitis Pallas (Perciformes: Gobiidae) from the Mediterranean and Black seas.
Systematic Parasitology,
Vol. 95,
Issue. 7,
p.
625.