



Introduction
Deinotheriidae is a family of browsing graviportal proboscideans with brachydont, bilophodont cheek teeth (except for trilophodont deciduous fourth premolars and first molars), and downturned lower tusks (Harris, Reference Harris, Maglio and Cooke1978; Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa, Delmar, Werdelin and Sanders2010). Deinotheres originated in Africa in the late Oligocene (Sanders et al., Reference Sanders, Kappelman and Rasmussen2004) and subsequently dispersed into Eurasia by the early Miocene (Tassy, Reference Tassy, Lindsay, Fahlbusch and Mein1989; Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa, Delmar, Werdelin and Sanders2010; Antoine et al., Reference Antoine, Métais, Orliac, Crochet, Flynn, Marivaux, Rajpar, Roohi, Welcomme, Wang, Flynn and Fortelius2013). Very little morphological change occurred during their evolutionary history except for an increase in body size (Gräf, Reference Gräf1957; Harris, Reference Harris1973, Reference Harris1976; Tobien, Reference Tobien1988; Huttunen, Reference Huttunen2002b; Poulakakis et al., Reference Poulakakis, Lymberakis and Fassoulas2005; Pickford and Pourabrishami, Reference Pickford and Pourabrishami2013). Three genera are widely considered valid: the primitive and diminutive Chilgatherium Sanders, Kappelman, and Rasmussen, Reference Sanders, Kappelman and Rasmussen2004, and the larger Prodeinotherium Éhik, Reference Éhik1930 and Deinotherium Kaup, Reference Kaup1829 (Sanders et al., Reference Sanders, Gheerbrant, Harris, Saegusa, Delmar, Werdelin and Sanders2010). Deinotherium is stratigraphically younger (and generally larger) than Prodeinotherium (Harris, Reference Harris, Maglio and Cooke1978), with some species, e.g., Deinotherium giganteum Kaup, Reference Kaup1829, exceeding 10 tons in weight (Larramendi, Reference Larramendi2016). There are thought to be three valid species of Deinotherium: Deinotherium giganteum from western and central Eurasia, Deinotherium indicum Falconer, Reference Falconer1845 from the Indian subcontinent, and Deinotherium bozasi Arambourg, Reference Arambourg1934 from East Africa (Sahni and Tripathi, Reference Sahni and Tripathi1957; Harris, Reference Harris1973; Huttunen, Reference Huttunen2002b; but see Pickford and Pourabrishami, Reference Pickford and Pourabrishami2013, for an alternative taxonomic scheme). Deinotherium bozasi, the last known species, went extinct in the early Pleistocene (ca. 1 Ma).
South Asian deinotheres are known largely from isolated teeth in lower to upper Miocene deposits in the Siwalik Group of Indo-Pakistan, the Manchars of Sind in eastern Pakistan, the Gaj Series in the Bugti Hills of western Pakistan, the Dharmsala Group of northern India, Kutch and Perim (Piram) Island in western India, and in the Dang Valley in Nepal (Falconer, Reference Falconer1845, Reference Falconer and Murchison1868; Lydekker, Reference Lydekker1876, Reference Lydekker1880; Pilgrim, Reference Pilgrim1912, Reference Pilgrim1917; Forster-Cooper, Reference Forster-Cooper1922; Palmer, Reference Palmer1924; Sahni and Tripathi, Reference Sahni and Tripathi1957; Dehm, Reference Dehm1963; Khan et al., Reference Khan, Raghavendra Rao, Ganju and Sankaran1971; Sahni and Mishra, Reference Sahni and Mishra1975; West et al., Reference West, Lukacs, Munthe and Hussain1978; Barry et al., Reference Barry, Lindsay and Jacobs1982; Sahni and Gupta, Reference Sahni and Gupta1982; Raza et al., Reference Raza, Barry, Grant and Martin1984; Vasishat, Reference Vasishat1985; Tiwari et al., Reference Tiwari, Verma and Bhandari2006; Bhandari et al., Reference Bhandari, Mohabey, Bajpai, Tiwari and Pickford2010; Sankhyan and Sharma, Reference Sankhyan and Sharma2014; Kapur et al., Reference Kapur, Pickford, Chauhan and Thakkar2019). The taxonomic validity of the large species, Deinotherium indicum from the Tortonian of Perim Island in the Gulf of Cambay and Haritalyangar in the Siwalik Hills, is generally accepted. However, several species have been described from smaller teeth recovered from uncertain stratigraphic contexts in the Bugti Hills, Siwalik Hill, Potwar Plateau, and the Manchars: Antoletherium of Falconer, Reference Falconer and Murchison1868, Prodeinotherium pentapotamiae Lydekker, Reference Lydekker1876, Deinotherium sindiense Lydekker, Reference Lydekker1880, Deinotherium naricum Pilgrim, Reference Pilgrim1908, and Deinotherium orlovii Sahni and Tripathi, Reference Sahni and Tripathi1957; the validity of these taxa is still debated (Lydekker, Reference Lydekker1880; Gräf, Reference Gräf1957; Sahni and Tripathi, Reference Sahni and Tripathi1957; Harris, Reference Harris1973; Huttunen, Reference Huttunen2002b; Pickford and Pourabrishami, Reference Pickford and Pourabrishami2013; Bhandari et al., Reference Bhandari, Pickford and Tiwari2015).
We here report the first occurrence of Deinotherium indicum from the Tapar beds in Kutch, western India. Deinothere remains are rare in Kutch. Lydekker (Reference Lydekker1876) described two specimens collected by Mr. F. Fedden (Wynne, Reference Wynne1872) from the village of Sambera (Sambda)—an incomplete right M1 (GSI A3) and a right M2 (GSI A10), as Prodeinotherium pentapotamiae based on their small size. Lydekker (Reference Lydekker1885) also reported a third specimen of P. pentapotamiae from Kutch—an incomplete left M1 (GSI A4). Since these early discoveries, the only other deinothere specimens from Kutch are a proximal fragment of a humerus from Sambera (Sahni and Mishra, Reference Sahni and Mishra1975) and dental remains referred to Deinotherium sindiense from Tapar (Bhandari et al., Reference Bhandari, Pickford and Tiwari2015) and Palasava (Kapur et al., Reference Kapur, Pickford, Chauhan and Thakkar2019). This new find of Deinotherium indicum from Kutch expands the geographic distribution of this species, hitherto only known from two or three localities. This specimen also increases our understanding of the variation in dental morphology of South Asian deinotheres, thereby allowing for a reassessment of previously defined dental apomorphies associated with this group. It also clarifies the taxonomy of Neogene deinotheres from Kutch and helps constrain the age of the Tapar beds to the late Miocene.
Geological setting
Stratigraphic information
The Neogene deposits from Kutch have been classified as the Khari Nadi, Chhasra, and Sandhan formations (Biswas, Reference Biswas1992; Catuneanu and Dave, Reference Catuneanu and Dave2017). The Khari Nadi and Chhasra formations (early Miocene) have been interpreted to be shallow-marine environments, whereas the Sandhan Formation (middle Miocene to Pliocene) is thought to represent a more terrestrial coastal setting (Catuneanu and Dave, Reference Catuneanu and Dave2017). The Tapar beds of the Kutch Basin in western India, known for their rich vertebrate fauna, consist of medium- to coarse-grained sandstones, siltstones, and mudstones with intermittent conglomerate beds (Fig. 1.3) and have been considered to form part of the Khari Nadi Formation in the past (Bhandari et al., Reference Bhandari, Mohabey, Bajpai, Tiwari and Pickford2010, Reference Bhandari, Pickford and Tiwari2015, Reference Bhandari, Kay, Williams, Tiwari, Bajpai and Hieronymus2018; Patnaik et al., Reference Patnaik, Sharma, Mohan, Williams, Kay and Chatrath2014). However, this is problematic because the Tapar beds are lithologically different from the Khari Nadi Formation. Unlike the latter, the Tapar sequence has abundant coarse-grained sandstone beds ingrained with pebbly clasts, calcareous nodules, agate pebbles, mudclasts, and conglomerates. These beds bear the greatest similarity to the Sandhan Formation (Biswas, Reference Biswas1992; Catuneanu and Dave, Reference Catuneanu and Dave2017) and likely represent a fluvial regime. The specimen of Deinotherium indicum described herein was eroding from a layer of pseudoconglomerate sandwiched between layers of micaceous sandstone in the Tapar beds (Fig 1.3). The pseudoconglomerate consists of mud clasts and soil concretions derived from paleosol horizons and lacks the typical pebbles and cobbles of a true conglomerate. This pseudoconglomerate is equivalent to the fossiliferous conglomerate described by Bhandari et al. (Reference Bhandari, Kay, Williams, Tiwari, Bajpai and Hieronymus2018).
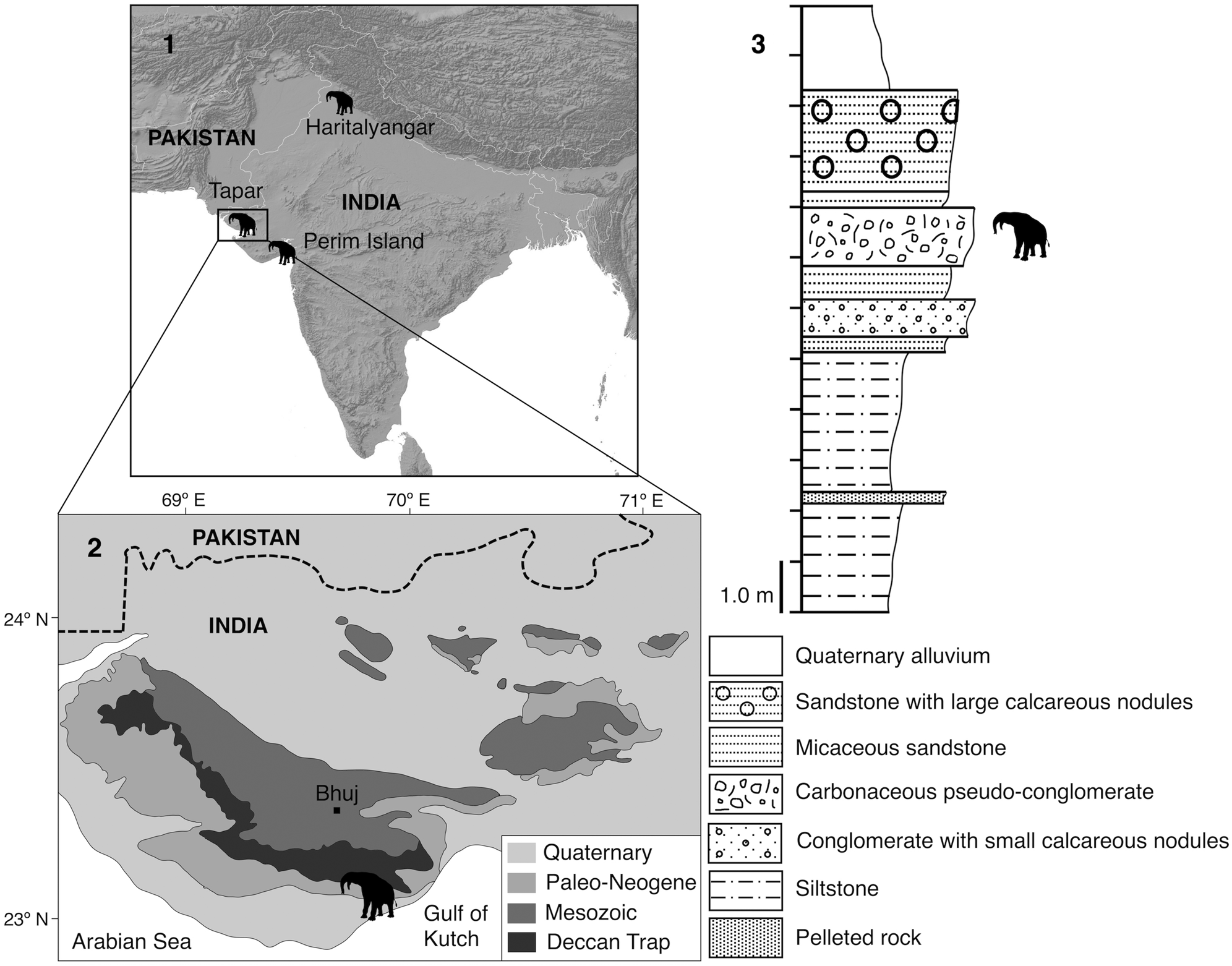
Figure 1. Geographic distribution of Deinotherium indicum in the Indian subcontinent: (1) locator map showing the three major localities where D. indicum was found; (2) geological map of Kutch; (3) stratigraphic column of the Tapar locality.
Locality information
Specimen PU KT-1 was found at Tapar, Kutch (23°15′12.28″N, 70°08′46.20″E), in the state of Gujarat, western India (Fig. 1.2).
Materials and methods
Specimen PU KT-1 is a right p4. It was collected in the Tapar beds by NPS, RP, KMS, NAS and YPS and cleaned with brushes. The specimen is now accessioned in the paleontological collections of the Department of Geology, Panjab University, Chandigarh. Measurements were taken using digital calipers.
Repositories and institutional abbreviations
BGR/LBEG = Bundesanstalt für Geowissenschaften und Rohstoffe/Landesamt für Bergbau Energie und Geologie Hannover, Hannover, Germany; GSI = Geological Survey of India, Kolkata, India; IMM = Nei Mongol Museum, Hohhot, China; NHMUK = The Natural History Museum, London, UK; PU = Panjab University, Chandigarh, India; SNSB-BSPG = Staatliche Naturwissenschaftliche Sammlungen Bayerns-Bayerische Staatssammlung für Paläontologie und Geologie, Munich, Germany. Upper case letters P and M refer to upper premolars and molars, whereas lower case letters p and m refer to mandibular premolars and molars, respectively; numbers following these letters indicate tooth position.
Systematic paleontology
Proboscidea Illiger, Reference Illiger1811
Deinotheroidea Bonaparte, Reference Bonaparte1845
Deinotheriidae Bonaparte, Reference Bonaparte1845
Deinotheriinae Bonaparte, Reference Bonaparte1845
Deinotherium Kaup, Reference Kaup1829
Type species
Deinotherium giganteum Kaup, Reference Kaup1829.
Other species
Deinotherium indicum Falconer, Reference Falconer1845 and Deinotherium bozasi Arambourg, Reference Arambourg1934.
Diagnosis
Large deinotheres. Dental formulae as for the family; tendency for the development of subsidiary styles on P3 and P4, and for simplification of the postmetaloph ornamentation of M2 and M3 when compared to Prodeinotherium. The skull rostrum not parallel to the mandibular symphysis and nearly horizontally aligned; rostral trough and external nares wide; preorbital swelling sited anteriorly on the rostrum; skull roof short and narrow at the temporal fossae; occiput sloping gently posteriorly; occipital condyles elevated above the level of the external auditory meatus; paroccipital process very elongate. Postcranial skeleton with cursorial modifications to graviportal structure; scapular spine reduced with no acromion or metacromion; carpals and tarsals narrow with dolichopodous metapodials exhibiting functional tetradactyly. (Modified from Harris, Reference Harris1973.)
Deinotherium indicum Falconer, Reference Falconer1845
Figures 2, 3.1–3.4
- Reference Falconer1845
Dinotherium indicum Falconer, p. 360, pl. 14, figs. 1, 1a.
- Reference Falconer and Murchison1868
Dinotherium perimense Falconer, p. 415.
- Reference Osborn1936
Deinotherium indicum; Osborn, p. 94 (emended spelling).
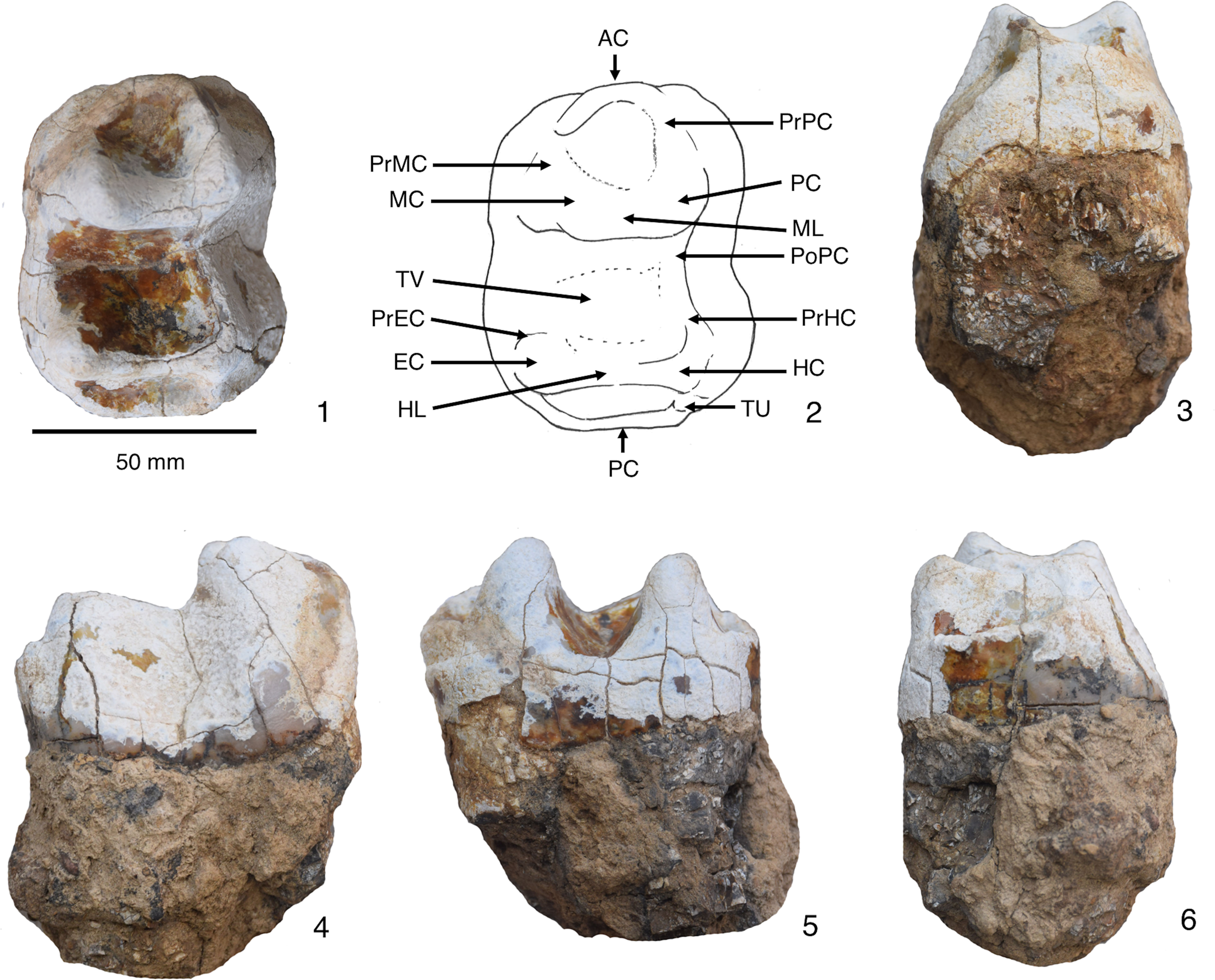
Figure 2. Deinotherium indicum, PU KT-1, right p4: (1) occlusal view; (2) line drawing of the occlusal view; (3) anterior view; (4) buccal view; (5) lingual view; (6) posterior view. AC = anterior cingulum; EC = entoconid; HC = hypoconid; HL = hypolophid; MC = metaconid; ML = metalophid; PC = protoconid; PoPC = post-protocristid; PrEC = pre-entocristid; PrHC = pre-hypocristid; PrMC = pre-metacristid; PrPC = pre-protocristid; TU = tubercle; TV = transverse valley.

Figure 3. Deinothere p4s from the Indian subcontinent: (1) PU KT-1, Deinotherium indicum, right p4; (2) GSI A585, Deinotherium indicum, right p4; (3) GSI A586, Deinotherium indicum, left p4; (4) GSI A596, Deinotherium indicum, right p4; (5) GSI A20, Antoletherium, left p4; (6) GSI A22, Prodeinotherium pentapotamiae, left p4; (7) SNSB-BSPG 1956 II 226, P. pentapotamiae, right p4. Figures 3.2, 3.3, and 3.4 modified from Sahni and Tripathi (Reference Sahni and Tripathi1957); Figures 3.5 and 3.6 modified from Lydekker (Reference Lydekker1880).
Holotype
Hypolophid and posterior cingulum of a left m2 (NHMUK PV OR 14756) from a ferruginous conglomerate along the shoreline, Perim (Piram) Island, Gulf of Cambay, Gujrat, India (Falconer, Reference Falconer1845, pl. 14, figs. 1, 1a).
Diagnosis
Very large deinothere with thick enamel on dentition. Mandible deep and wide, with a slightly convex labial surface, and a more circular cross section toward the posterior; m2 with transverse valley between the protolophid and metalophid and well-developed talonid; m3 bounded by tubercles on either side; m1 with tubercle on the buccal surface at the opening of the transverse valley. (Modified from Sahni and Tripathi, Reference Sahni and Tripathi1957.)
Occurrence
Tortonian (late Miocene) of Perim Island in the Gulf of Cambay, Gujarat, India (21°37′N, 72°22′30″E), Haritalyangar in the state of Himachal Pradesh, India (31°32′N, 76°38′E), and Tapar in Kutch, India (23°15′12.28″N, 70°08′46.20″E) (Fig. 1.1).
Description
Specimen PU KT-1 is a slightly worn isolated right p4 from a large deinothere (Fig. 2). The tooth is subrectangular in occlusal view and bilophodont. The anteroposterior length is 76.0 mm, the transverse diameter of the metalophid is 55.0 mm, and that of the hypolophid is 61.0 mm. Maximum crown height is 51.0 mm. The posterior width is greater than the anterior. The enamel is weathered and cracked over the entire occlusal surface, but the four main cusps (protoconid, hypoconid, metaconid, and entoconid) are well defined. The metalophid and hypolophid are anteriorly concave with the latter being slightly longer and narrower than the former. These two lophids are connected by the post-protocristid, which descends into the transverse valley and joins the pre-hypocristid. The longitudinal ridge formed by the post-protocristid and pre-hypocristid is buccally positioned so that the transverse valley is wide and deep and covers most of the mesiolingual surface. The transverse valley is blocked completely on the buccal side and open on the lingual side. The pre-entocristid is low and descends toward the transverse valley. A very small tubercle is present on the posterobuccal surface. The cingulum is absent on both buccal and lingual sides, but a well-developed but weakly crenulated cingulum is present at the anterior edge. The posterior cingulum is also well developed, weakly crenulated, and fused with a post-hypocristid.
Remarks
The present specimen is the largest deinothere tooth known from Kutch, much larger than the teeth described by Lydekker (Reference Lydekker1880) and Kapur et al. (Reference Kapur, Pickford, Chauhan and Thakkar2019) from this region. Complete and unworn lower dentition from South Asian deinotheres are rare. The hypodigm of Deinotherium indicum from Perim Island includes a mandibular fragment with p3–m3, but with broken crowns (NHMUK PV OR 14739a; Falconer, Reference Falconer and Murchison1868, pl. 33, fig 5). The incomplete nature of the lower dentition from the Perim Island specimen prevents a morphological comparison to PU KT-1, but the p4s are very similar in size (Table 1). Sahni and Tripathi (Reference Sahni and Tripathi1957) described three additional p4s referred to Deinotherium indicum. Two p4s (GSI A585, A586) from Haritalyangar are well worn and therefore unsuitable for a morphological comparison; they are however similar in size to PU KT-1 (Fig. 3.2, 3.3, Table 1). The third p4 (GSI A596) is from an unknown locality, but bears many morphological features in common with PU KT-1 including the longitudinal ridge being more buccally positioned resulting in a wide and deep transverse valley, and a well-developed anterior cingulum with a strong C-shaped extension (Fig. 3.1, 3.4). The main difference between these two specimens is in the placement of the tubercle. Sahni and Tripathi (Reference Sahni and Tripathi1957) reported that the tubercle on GSI A596 is placed next to the longitudinal ridge on the buccal side (they erroneously listed this specimen as a left p4 when in fact it is a right p4). In contrast, the tubercle on PU KT-1 is located posterobucally near the posterior cingulum (Fig. 2).
Table 1. A comparison of deinothere p4s from Asia. Lp4 = left p4; Rp4 = right p4; * = incomplete specimen; + = measurement was greater on the complete tooth. All measurements in mm.

Three smaller p4s have also been reported from the Indian subcontinent (Fig. 3.5–3.7, Table 1). Two have been referred to Prodeinotherium pentapotamiae (SNSB-BSPG 1956 II 226 and GSI A22) and the third belongs to the specimen of Antoletherium (GSI A20). The specimen of Antoletherium is a left mandibular fragment with p4–m2 (Lydekker, Reference Lydekker1880, pl. 29, figs. 2, 3). The anterobuccal surface of the p4 is damaged, therefore we cannot comment on the structure of the cingulum (Fig. 3.5). However, much like PU KT-1, the tooth is subrectangular with the hypolophid being longer than the metalophid. The longitudinal ridge, in contrast, although positioned somewhat buccally, bends mesially, in effect dividing the transverse valley into two halves. Sahni and Tripathi (Reference Sahni and Tripathi1957) reported a tubercle on the buccal surface formed in the valley between the metalophid and hypolophid, but this structure is not clear in Lydekker's (Reference Lydekker1880, pl. 29, figs. 2, 3) illustration (Fig. 3.5). This tooth is also considerably smaller than PU KT-1 (Table 1). The morphology seen on the p4 of Antoletherium is reflected in the left p4 (GSI A22) referred to P. pentapotamiae by Lydekker (Reference Lydekker1880; Fig. 3.6) and a right p4 (SNSB-BSPG 1956 II 226) described by Dehm (Reference Dehm1963) from Cheskewala, 14 km SW of Chinji Village (Fig. 3.7). On both of these teeth, the longitudinal ridge appears to bisect the transverse valley into two halves, in contrast to PU KT-1. The p4 of Antoletherium (GSI A20) and of P. pentapotamiae (GSI A22) each lack a posterior cingulum, and GSI A22 lacks any dental tubercles. Both of these teeth are also smaller than PU KT-1 (Table 1). SNSB-BSPG 1956 II 226 and GSI A22 both lack a strong anterior cingulum (Fig. 3.6, 3.7).
Discussion
The first deinothere teeth from Tapar were reported by Bhandari et al. (Reference Bhandari, Pickford and Tiwari2015) and referred to Deinotherium sindiense based on their small size. However, these remains are too fragmentary to be systematically diagnostic, therefore, until additional evidence is presented, we consider Deinotherium indicum to be the only species of deinothere from the Tapar Beds. Assessing Neogene deinothere taxonomy from isolated teeth is challenging because very little dental variation exists between the genera Prodeinotherium and Deinotherium and among their species (Gräf, Reference Gräf1957; Harris, Reference Harris1973; Huttunen, Reference Huttunen2002a). Based on its size, PU KT-1 is comparable to teeth referred to Deinotherium indicum from Perim Island, Haritalyangar, and an unknown locality (Table 1), Deinotherium giganteum p4s from Europe (Depéret, Reference Depéret1887; Gräf, Reference Gräf1957; Tobien, Reference Tobien1988; Gasparik, Reference Gasparik1993; Huttunen, Reference Huttunen2002a; Vergiev and Markov, Reference Vergiev and Markov2010; Pickford and Pourabrishami, Reference Pickford and Pourabrishami2013), and Deinotherium bozasi p4s from Kenya (Harris, Reference Harris1976). It is larger than p4s referred to P. pentapotamiae and Antoletherium from South Asia (Table 1), a p4 referred to P. cf. P. pentapotamiae from Thailand (Sickenberg, Reference Sickenberg1971), a p4 from Chinese Prodeinotherium sinense Qiu et al., Reference Qiu, Wang, Li, Deng and Sun2007 (Table 1), p4s from P. bavaricum (von Meyer, Reference von Meyer1833), and P. hungaricum Éhik, Reference Éhik1930 from Europe (Gräf, Reference Gräf1957; Gasparik, Reference Gasparik1993; Huttunen, Reference Huttunen2002a; Huttunen and Göhlich, Reference Huttunen and Göhlich2002; Koufos et al., Reference Koufos, Zouros and Mourouzidou2003; Pickford and Pourabrishami, Reference Pickford and Pourabrishami2013; Tóth and Hyžný, Reference Tóth and Hyžný2013), and P. hobleyi (Andrews, Reference Andrews1911) from East Africa (Harris, Reference Harris1973). PU KT-1 is however smaller than the p4s referred to Deinotherium proavum Eichwald, Reference Eichwald1835 from Europe, although it is morphologically similar (Pickford and Pourabrishami, Reference Pickford and Pourabrishami2013). Nonetheless, we choose not to synonymize similarily sized Old World deinotheres until more crainal and postcranial remains are recovered from India. Based on size and geographic provenance, we refer this tooth to Deinotherium indicum.
The presence of Deinotherium indicum at Tapar is helpful in refining the age of the locality. It has been suggested previously that the Tapar locality is early Miocene in age (Bhandari et al., Reference Bhandari, Mohabey, Bajpai, Tiwari and Pickford2010). However, similar-sized p4s referred to Deinotherium indicum have been recovered from Haritalyangar (Table 1), a locality that has been magnetostratigraphically dated to 10.1–8.6 Ma (Pillans et al., Reference Pillans, Williams, Cameron, Patnaik, Hogarth, Sahni, Sharma, Williams and Bernor2005), and thus we can infer a similar age range for Tapar. The faunal assemblage from Tapar includes Muridae gen. indet. sp. indet. of Illiger, Reference Illiger1811, an indeterminate ‘insectivore,’ ?Amphicyonidae sp. of Haeckel, Reference Haeckel1866, Sivapithecus sp. of Pilgrim, Reference Pilgrim1910, Gomphotherium sp. of Burmeister, Reference Burmeister1837, Brachypotherium sp. of Roger, Reference Roger1904, a large rhinocerotid, hipparionine horses, Kachchhchoerus salinus (Pilgrim, Reference Pilgrim1926), Sanitherium schlagintweiti von Meyer, Reference von Meyer1866, Dorcatherium minus Lydekker, Reference Lydekker1876, Giraffa priscilla Matthew, Reference Matthew1929, cf. Protragocerus sp. of Depéret, Reference Depéret1887, and cf. Gazella sp. of de Blainville, Reference de Blainville1816 (Bhandari et al., Reference Bhandari, Mohabey, Bajpai, Tiwari and Pickford2010, Reference Bhandari, Pickford and Tiwari2015, Reference Bhandari, Kay, Williams, Tiwari, Bajpai and Hieronymus2018). This fauna is similar to that found at Haritalyangar and other similarly aged faunas from the Dhok Pathan Formation on the Potwar Plateau in Pakistan (Barry et al., Reference Barry, Lindsay and Jacobs1982; Pillans et al., Reference Pillans, Williams, Cameron, Patnaik, Hogarth, Sahni, Sharma, Williams and Bernor2005), further indicating a late Miocene (Tortonian) age for Tapar. This age assessment supports the correlation of the Tapar beds with the younger Samdhan Formation, rather than the older Khari Nadi Formation.
The morphology of this largely complete p4 helps resolve some of the problems surrounding the systematics of Indian deinotheres. Sahni and Tripathi (Reference Sahni and Tripathi1957) argued that an apomorphy of Deinotherium indicum is the presence of a dental tubercle (homologous to a style) on the ‘outer side’ of p4, whereas Prodeinotherium pentapotamiae (their Deinotherium pentapotamiae) lacks this structure. We interpret the ‘outer side’ to mean the buccal surface, because the only p4 that they described with a tubercle is GSI A596, a right p4 with a tubercle on the buccal margin of the occlusal surface (Fig 3.4). This apomorphy has been problematic because it has been used to synonymize much smaller and older taxa, e.g., Antoletherium (a taxon formerly synonymized with P. pentapotamiae), with the larger Deinotherium indicum (see Sahni and Tripathi, Reference Sahni and Tripathi1957). We examined the validity of this apomorphy by comparing the known p4s from the Indian subcontinent, Europe, China, and Africa. PU KT-1 has a small tubercle, but is placed posterobucally, and not in the transverse valley unlike the similarly sized p4, GSI A596. The p4 of P. pentapotamiae (GSI A22) lacks tubercles (Fig. 3.6), but the similarly sized p4 of P. sinense has a buccal tubercle (Qiu et al., Reference Qiu, Wang, Li, Deng and Sun2007, figs. 2, 3). Huttunen (Reference Huttunen2002a) also showed that in a sample of European deinothere teeth, styles or tubercles were present on both large and small p4s. Moreover, Harris (Reference Harris1973) stated that apart from the p3, the lower dentition of African Prodeinotherium and Deinotherium are identical except for size, and that the presence of mesostyles is diagnostic to the superior P3 and P4 of Deinotherium. Given the variation in the presence of tubercles or styles on the inferior dentition, particularly on the p4s of both large and small Asian deinotheres, and the lack of variation in deinothere inferior dentition from Europe and Africa, we reject the presence of tubercles as an apomorphy of Deinotherium indicum as stated by Sahni and Tripathi (Reference Sahni and Tripathi1957). We agree with Harris (Reference Harris1973) and Huttunen (Reference Huttunen2002a) that the major differences between Deinotherium and Prodeinotherium inferior dentition lie primarily in size and not in morphology.
Based on our analysis of the morphological variation in deinothere p4s, we revise Sahni and Tripathi's (Reference Sahni and Tripathi1957) taxonomy of Deinotherium indicum and exclude Antoletherium from the list of synonymized species. Antoletherium was described by Falconer (Reference Falconer and Murchison1868) based on a drawing of a mandibular fragment with p4–m2 discovered by Lieutenant Garnett at Kushalgarh near Attock in what is now Pakistan. The stratigraphic provenance of the Kushalgarh fossils is unknown (Theobald, Reference Theobald1881), but biostratigraphically, the fossils are inferred to correlate with the Lower to the lowermost Middle Siwaliks or middle Miocene (Flynn et al., Reference Flynn, Lindsay, Pilbeam, Raza, Morgan, Barry, Badgley, Behrensmeyer, Cheema, Rajpar, Opdyke, Wang, Flynn and Fortelius2013). Despite the small size of Antoletherium, Sahni and Tripathi (Reference Sahni and Tripathi1957) used the presence of a buccal tubercle on the p4 (not seen in Lydekker's Reference Lydekker1880 illustration) along with other tubercles on the m1 and m2 to synonymize it with Deinotherium indicum. However, because we do not regard tubercles on the p4 as a valid apomorphy, and the small P. sinense also has tubercles on m1–m3 (Qiu et al., Reference Qiu, Wang, Li, Deng and Sun2007), the morphological criterion used by Sahni and Tripathi (Reference Sahni and Tripathi1957) is too variable and cannot be used diagnostically for the large genus Deinotherium. The older stratigraphic context for Antoletherium provides additional evidence against its synonymy with Deinotherium indicum. A general trend in deinothere evolution is a temporal increase in size across successive taxa (Gräf, Reference Gräf1957; Harris, Reference Harris, Maglio and Cooke1978; Tobien, Reference Tobien1988; Huttunen, Reference Huttunen2002b; Poulakakis et al., Reference Poulakakis, Lymberakis and Fassoulas2005; Pickford and Pourabrishami, Reference Pickford and Pourabrishami2013), further indicating that the specimen of Antoletherium represents a distinct older and smaller taxon. Moreover, Antoletherium bears the greatest metric similarlty to the middle Miocene P. pentapotamiae (Table 1). Thus, we accept Lydekker's (Reference Lydekker1880) original synonymization of Antoletherium with the latter.
Previously, only small deinotheres (Prodeinotherium pentapotamiae and Deinotherium sindiense) have been documented from Kutch (Lydekker, Reference Lydekker1880; Bhandari et al., Reference Bhandari, Pickford and Tiwari2015; Kapur et al., Reference Kapur, Pickford, Chauhan and Thakkar2019). Small specimens of deinotheres referred to Deinotherium sindiense have been recovered from Palasava (Kapur et al., Reference Kapur, Pickford, Chauhan and Thakkar2019) and Pasuda (Bhandari et al., Reference Bhandari, Pickford and Tiwari2015), and although direct dates are not available, the associated fauna suggest a middle Miocene (Langhian–Serravalian, ca. 15.9–11.6 Ma) age. Small deinothere remains from the undated locality of Sambera have been referred to P. pentapotamiae (GSI A10; Lydekker, Reference Lydekker1876). Prodeinotherium pentapotamiae is commonly found in the middle Miocene Lower Siwaliks (Dehm, Reference Dehm1963; West et al., Reference West, Lukacs, Munthe and Hussain1978), suggesting a similar middle Miocene age for Sambera as well. Determining whether multiple species of small deinotheres existed during the middle Miocene is beyond the scope of this study, but based on isolated dentition alone, Deinotherium sindiense and P. pentapotamiae are very similar to one another in size (Supplemental Table 1). However, we follow Bhandari et al. (Reference Bhandari, Pickford and Tiwari2015) and reject the hypothesis proposed by Sahni and Tripathi (Reference Sahni and Tripathi1957) that Deinotherium sindiense is synonymous with Deinotherium indicum considering its far smaller size (Supplemental Table 1) and the fragmentary nature of the holotype (Lydekker, Reference Lydekker1880) precluding a detailed morphological comparison. The confirmed presence of Deinotherium indicum at Tapar shows that ecosystems in western India experienced a replacement of smaller deinotheres by larger species in the late Miocene, just as has been seen in the northern part of the Indian subcontinent (Sahni and Tripathi, Reference Sahni and Tripathi1957) and elsewhere in the world (Harris, Reference Harris and Harris1983).
Acknowledgments
The authors would like to thank A.C. Pritchard, D. DeMar, J. Saarinen, and M.T. Carrano for helpful discussions. We thank P. Brewer and N. Gabriel for providing photographs of Perim Island deinothere specimens from NHMUK, and G. Rößner and M. Schellenberger for providing photographs of the Siwalik deinothere material housed at the SNSB-BSPG. Lastly, we thank H. Zhang, an anonymous reviewer, and H.-D. Sues and the editorial staff for feedback that helped improve this manuscript. Financial support was provided by the Government of India, Ministry of Earth Science (MoES/ P.O. (Geoscience)/46/2015) and Department of Science and Technology, Science and Engineering Research Board (SERB HRR/2018/000063).
Accessibility of supplemental data
A tabular list of specimens and measurements used for comparison is available in Supplemental Data 1. Data are available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.dfn2z34wz.




