



Introduction
The conodont-based Lochkovian (Lower Devonian) subdivision has been extensively investigated by worldwide Devonian conodont researchers. The first comprehensive subdivision scheme was presented by Klapper (Reference Klapper1977), who subdivided the Lochkovian in western North America into the hesperius Zone, eurekaensis Zone, which already was defined by Klapper and Murphy (Reference Klapper and Murphy1975), Ozarkodina n. sp. D Zone (= delta Zone), and pesavis Zone, which already was defined by Fåhræus (Reference Fåhræus1971). However, great difficulty was encountered in the application of this central Nevada scheme to the Lochkovian in other places around the world, as suggested by Valenzuela-Ríos (Reference Valenzuela-Ríos1994a, Reference Valenzuela-Ríosb) and Valenzuela-Ríos and Murphy (Reference Valenzuela-Ríos and Murphy1997). On the basis of the comparison of conodont records from the Spanish Central Pyrenees with those from central Nevada, Valenzuela-Ríos (Reference Valenzuela-Ríos1994a) subdivided the delta and pesavis zones into five and two zones, respectively. Later, Valenzuela-Ríos (Reference Valenzuela-Ríos1994b) discussed the different uses of the pesavis Zone and proposed a new subdivision of this Zone and the uppermost part of the delta Zone into the pandora β-gilberti and gilberti-eosulcatus zones. Valenzuela-Ríos and Murphy (Reference Valenzuela-Ríos and Murphy1997) subdivided the Lochkovian into the lower, middle, and upper parts based upon the evolution and global records of the genus Ancyrodelloides Bischoff and Sannemann, Reference Bischoff and Sannemann1958. The lower boundaries of the middle and upper Lochkovian were suggested to be defined by the first occurrences of Lanea omoalpha Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999, and Masarella pandora Murphy et al., Reference Murphy, Matti and Walliser1981 β morphotype, respectively. The middle Lochkovian consists, in ascending order, of the omus α-eleanorae, eleanorae-trigonicus, and trigonicus-pandora β zones. Subsequently, the omus α-eleanorae Zone was further subdivided into the omoalpha-transitans and transitans-eleanorae zones (Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999). Another alternative zonal scheme proposed by Slavík et al. (Reference Slavík, Carls, Hladil and Koptíková2012) in the Prague Synform was similar to that of Murphy and Valenzuela-Ríos (Reference Murphy and Valenzuela-Ríos1999) in the upper half of the middle Lochkovian and upper Lochkovian zonation, but the lower Lochkovian and lower half of the middle Lochkovian zonation consists, in ascending order, of the hesperius-optima, optima-breviramus, breviramus-omoalpha, omoalpha-carlsi, carlsi-eoeleanorae, eoeleanorae-boucoti, and boucoti-transitans zones. In addition, they shifted the position of the lower boundary of the middle Lochkovian to the entry of A. carlsi (Boersma, Reference Boersma1973) (= Lanea carlsi in their paper). Meanwhile, Corradini and Corriga (Reference Corradini and Corriga2012) proposed a Lochkovian zonation for Sardinia and the Carnic Alps that differs from those in the Spanish Central Pyrenees and the Prague Synform in that the lower Lochkovian and lower half of the middle Lochkovian are subdivided into the hesperius and carlsi zones. By comparing records from the Spanish Central Pyrenees with those from the Prague Synform, Valenzuela-Ríos et al. (Reference Valenzuela-Ríos, Slavík, Liao, Calvo, Hušková and Chadimová2015) subdivided the former trigonicus-pandora β Zone into the trigonicus-kutscheri and kutscheri-pandora β zones. The recommended standard Lochkovian conodont zonation is mainly established on the basis of these peri-Gondwana subdivision schemes (Becker et al., Reference Becker, Marshall, Da Silva, Gradstein, Ogg, Schmitz and Ogg2020).
The Lochkovian zonation of Klapper (Reference Klapper1977) was widely applied in China (C.Y. Wang, Reference Wang1981, Reference Wang1982; Li, Reference Li1987; Wang and Zhang, Reference Wang and Zhang1988; Liao and Xia, Reference Liao and Xia1996; Xia, Reference Xia1997; Wang et al., Reference Wang, Zhang, Rong, Wang and Cai2001; P. Wang, Reference Wang2006), despite the subsequent recognition that this scheme is not globally applicable (Valenzuela-Ríos and Murphy, Reference Valenzuela-Ríos and Murphy1997). Accordingly, it is clear that these previously reported Lochkovian conodont zonations in China cannot be precisely correlated with those in the Spanish Central Pyrenees, central Nevada, the Prague Synform, Sardinia, and the Carnic Alps. It should be noted that the strata in most of the studied Lochkovian sections in China are discontinuous or poorly exposed, resulting in only one to two zones of Klapper (Reference Klapper1977) being reported from one section in most cases. Furthermore, all of the sections located in remote or mountainous areas do not belong to the South China Block, but to other blocks or terranes. According to Lu and Chen (Reference Lu and Chen2016), the widely distributed and extensively developed Devonian carbonate succession in the South China Block started in the early Emsian, while the Lochkovian is mainly represented by sandstone, siltstone, and mudstone. Recently, Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018) studied the Lower Devonian conodont biostratigraphy at the well-known Alengchu section (western Yunnan), but only a few Lochkovian taxa, without important and unequivocal zonal markers, were collected, thus precluding a precise and high-resolution conodont-based subdivision. This locality probably represents one of the few places where the Lower Devonian carbonate succession is continuously developed. Therefore, the goal of this research is to update the Lochkovian conodont biostratigraphy at the Alengchu section, which will furnish important Chinese materials for intercontinental correlation, and to test the global relevance of peri-Gondwana conodont-based subdivision schemes in China.
Geological setting
The Alengchu section is named after a small village, Alengchu, along the Jinshajiang River that is situated ~50 km northeast of Lijiang, western Yunnan (Fig. 1). The Lijiang area, plus the adjacent Yanbian or Panzhihua area (southern Sichuan), the geographically so-called Yanbian-Lijiang or Panzhihua-Lijiang district, was usually thought by tectonic geologists to constitute the southwestern margin of the South China Block (Zhang et al., Reference Zhang, Guo, Wang, Li, Dong, Liu, He, Chen, Lu and Yao2013). However, mainly based on the comparison of the Silurian conodonts, brachiopods, and graptolites between the Panzhihua-Lijiang district and the South China Block, Rong et al. (Reference Rong, Chen, Su, Ni, Zhan, Chen, Fu, Li and Fan2003) suggested that this district is not a southwestern component of the South China Block. Later, in a study of the Silurian conodont biostratigraphy at the Baizitian section, Panzhihua, Wang et al. (Reference Wang, Wang, Yang and Xie2009) proposed another hypothesis that the Panzhihua-Lijiang district probably belongs to neither the South China Block nor the Lhasa Block, but to an unknown small terrane. This opinion on the provenance of the Panzhihua-Lijiang district was recently further supported by Wang et al. (Reference Wang, Rong, Tang, Wang and Zhang2016), who noted the great differences in the tectono-geographical location, stratigraphic sequence, sedimentary development, and faunal composition among the Panzhihua-Lijiang district, the South China Block, and the Lhasa Block.

Figure 1. Location of the study area, with the star showing the site of the Alengchu section in western Yunnan, China.
The Lower Devonian is continuously developed and well exposed at the Alengchu section. Yu and Liao (Reference Yu and Liao1978) first subdivided the Lower Devonian in ascending order into the Shanjiang, Alengchu, and Banmandaodi formations. For the underlying Silurian part, two lithological units, consisting in ascending order of the Longmaxi and Baizitian formations, was adopted by Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018). The Silurian Baizitian Formation, which is marked by thick-bedded nodular limestone, is conformably overlain by the Devonian Shanjiang Formation, which is composed of thin- to medium-bedded bioclastic limestone intercalated with siltstone and shale. The Alengchu Formation is represented by medium- to thick-bedded limestone intercalated with shale. The overlying Banmandaodi Formation consists mainly of sandstone, shale, and limestone. Benthic and pelagic fossils, including rugose corals, tentaculites, and conodonts, are abundant throughout the whole succession of this section. By subdividing the Lower Devonian rugose corals at the Alengchu section into five assemblages, Yu and Liao (Reference Yu and Liao1978) pointed out that the rugose coral fauna in Lijiang shows a great difference from the contemporaneous rugose coral fauna in the South China Block. Subsequently, the Lower Devonian tentaculites from the Shanjiang Formation and lower part of the Alengchu Formation were studied by Jiang (Reference Jiang1980). Wang (Reference Wang1982) was the first to briefly report the conodont fauna from the Shanjiang Formation to the Banmandaodi Formation and made a preliminary determination on the ages of these lithological units. Recently, Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018) re-studied this section and demonstrated that the Shanjiang Formation can be correlated with the Lochkovian and Pragian, whereas the Alengchu Formation belongs to the lower part of the Emsian.
Materials and methods
Twenty-four limestone samples, each weighing 3–4 kg, were collected in 2019 from the uppermost part of the Baizitian Formation to the lower part of the Shanjiang Formation at the Alengchu section (Table 1; Fig. 2). The samples were crushed mechanically into small pieces of ~2–5 cm in diameter and dissolved in dilute (8–12%) acetic acid. The insoluble residues were then washed, air-dried, and finally concentrated by heavy-liquid separation using sodium polytungstate. Specimens coated with gold were photographed using a Scanning Electron Microscope (SEM) in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences.
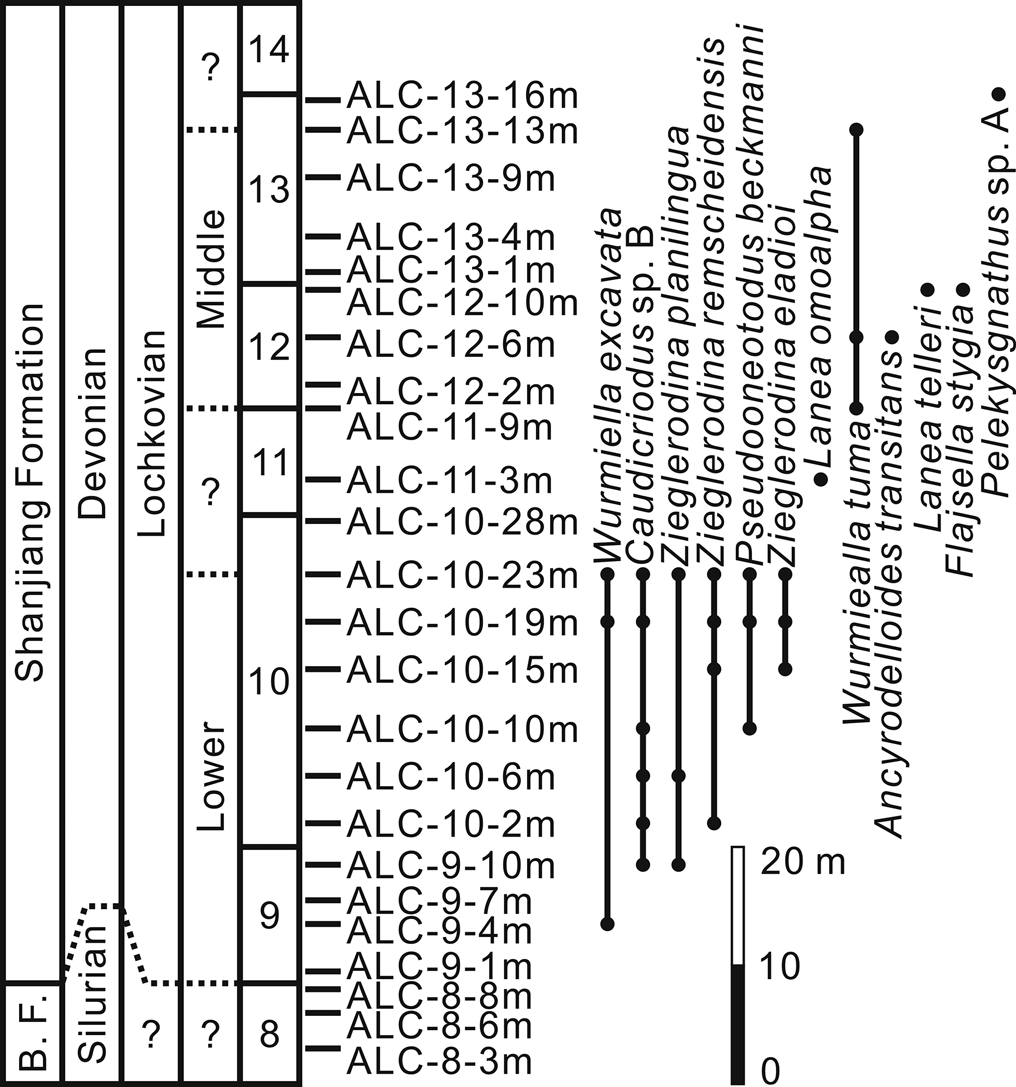
Figure 2. Distribution of conodonts in the lower part (Bed 9 to Bed 13) of the Shanjiang Formation at the Alengchu section, Lijiang, western Yunnan. Abbreviation: B.F., Baizitian Formation.
Table 1. Occurrence of conodont taxa from the lower part of the Shanjiang Formation at the Alengchu section.

Repository and institutional abbreviation
All specimens described and illustrated herein are deposited in the collections of the Nanjing Institute of Geology and Palaeontology (NIGP), Chinese Academy of Sciences.
Systematic paleontology
Nomenclature of the Pa and I elements proposed by Murphy and Valenzuela-Ríos (Reference Murphy and Valenzuela-Ríos1999, text-fig. 2) and Murphy (Reference Murphy2016, fig. 1), respectively, is followed herein.
Order Ozarkodina Dzik, Reference Dzik1976
Family Spathognathodontidea Hass, Reference Hass1959
Genus Ancyrodelloides Bischoff and Sannemann, Reference Bischoff and Sannemann1958
Type species
Ancyrodelloides trigonica Bischoff and Sannemann, Reference Bischoff and Sannemann1958.
Ancyrodelloides transitans (Bischoff and Sannemann, Reference Bischoff and Sannemann1958)
Figure 3.1, 3.2
- Reference Bischoff and Sannemann1958
Spathognathodus transitans Bischoff and Sannemann, p. 107, pl. 13, figs. 4, 5, 12, 14.
- Reference Serpagli, Gnoli, Mastandrea and Olivieri1978
Spathognathodus transitans Bischoff and Sannemann; Serpagli et al., pl. 27, fig. 5.
- Reference Lane and Ormiston1979
Ozarkodina transitans (Bischoff and Sannemann); Lane and Ormiston, p. 58, pl. 1, fig. 41, pl. 2, figs. 4, 5, 8, 9, 12, 13, pl. 3, fig. 21.
- Reference Klapper and Johnson1980
Ozarkodina transitans (Bischoff and Sannemann); Klapper and Johnson, p. 450.
- Reference Mastandrea1985a
Ozarkodina transitans (Bischoff and Sannemann); Mastandrea, p. 255, pl. 2, fig. 2, pl. 5, figs. 14, 15, 17–19.
- Reference Schönlaub1985
Ancyrodelloides transitans (Bischoff and Sannemann); Schönlaub, pl. 2, figs. 22, 24, pl. 3, fig. 1.
- Reference Barca, Gnoli, Oliviei and Serpagli1986
Ancyrodelloides transitans (Bischoff and Sannemann); Barca et al., pl. 31, fig. 6.
- Reference Wang and Zhang1988
Ancyrodelloides transitans (Bischoff and Sannemann); Wang and Zhang, pl. 1, fig. 11.
- Reference Wilson1989
Ancyrodelloides transitans (Bischoff and Sannemann) beta morph Lane and Ormiston; Wilson, p. 137, pl. 12, figs. 18, 19.
- Reference Ziegler1991
Ancyrodelloides transitans (Bischoff and Sannemann); Ziegler, p. 25, pl. 1, fig. 4.
- Reference Bardashev and Ziegler1992
Cruciodus transitans (Bischoff and Sannemann); Bardashev and Ziegler, pl. 2, figs. 14, 15, 19, 25.
- Reference Barrick and Klapper1992
Ancyrodelloides transitans (Bischoff and Sannemann); Barrick and Klapper, p. 48, pl. 4, fig. 13, pl. 5, figs. 1–5.
- Reference Valenzuela-Ríos1994a
Ancyrodelloides transitans (Bischoff and Sannemann); Valenzuela-Ríos, p. 41, pl.1, figs. 11, 14–18, 20, pl. 2, figs. 2, 4, 5, 8, 16. [with synonymy list]
- Reference Lazreq and Ouanaimi1998
Ancyrodelloides transitans (Bischoff and Sannemann); Lazreq and Ouanaimi, pl. 1, figs. 10, 11, 13.
- Reference Slavík1998
Ancyrodelloides transitans (Bischoff and Sannemann); Slavík, p. 159, pl. 1, figs. 3, 4.
- Reference Valenzuela-Ríos and García-López1998
Ancyrodelloides transitans (Bischoff and Sannemann); Valenzuela-Ríos and García-López, pl. 1, figs. 6, 8–10.
- ?Reference Corradini, Leone, Loi and Serpagli2001
Ancyrodelloides transitans (Bischoff and Sannemann); Corradini et al., pl. 1, fig. 3.
- Reference Farrell2003
Ancyrodelloides transitans (Bischoff and Sannemann) alpha morphotype Lane and Ormiston; Farrell, p. 132, pl. 5, figs. 10–12.
- non Reference Wang2006
Ancyrodelloides transitans (Bischoff and Sannemann); Wang, p. 213, pl. 3, figs. 10–12.
- Reference Jansen, Lazreq, Plodowski, Schemm-Gregory, Schindler, Weddige, Becker and Kirchgasser2007
Ancyrodelloides transitans (Bischoff and Sannemann); Jansen et al., fig. 3.9, 3.10.
- Reference Suttner2007
Ancyrodelloides transitans (Bischoff and Sannemann); Suttner, p. 29, pl. 21, figs. 3, 6–9.
- Reference Corradini and Corriga2012
Ancyrodelloides transitans (Bischoff and Sannemann); Corradini and Corriga, fig. 6N.
- Reference Slavík, Carls, Hladil and Koptíková2012
Ancyrodelloides transitans (Bischoff and Sannemann); Slavík et al., fig. 6.10.
- Reference Mavrinskaya and Slavík2013
Ancyrodelloides transitans (Bischoff and Sannemann); Mavrinskaya and Slavík, fig. 5S, T.
- Reference Corriga, Corradini and Walliser2014b
Ancyrodelloides transitans (Bischoff and Sannemann); Corriga et al., fig. 5X.
- Reference Corradini, Pondrelli, Simonetto, Corriga, Spaletta, Suttner, Kido, Mossoni and Serventi2016
Ancyrodelloides transitans (Bischoff and Sannemann); Corradini et al., pl. 4, fig. 11.
- Reference Schönlaub, Corradini, Corriga and Ferretti2017
Ancyrodelloides transitans (Bischoff and Sannemann); Schönlaub et al., pl. 3, fig. 14.
- Reference Slavík2017
Ancyrodelloides transitans (Bischoff and Sannemann); Slavík, fig. 6.10.
- Reference Valenzuela-Ríos and Liao2017
Ancyrodelloides transitans (Bischoff and Sannemann); Valenzuela-Ríos and Liao, fig. 4.7, 4.8.
- Reference Valenzuela-Ríos, Liao and Martínez-Pérez2017a
Ancyrodelloides transitans (Bischoff and Sannemann); Valenzuela-Ríos et al., fig. 6.1.
- Reference Valenzuela-Ríos, Liao and Calvo2017b
Ancyrodelloides transitans (Bischoff and Sannemann); Valenzuela-Ríos et al., fig. 4.4.
Holotype
BiSa 1958/26 from the Transgressionshorizont at Schübelebene near Elbersreuth, Frankenwald, Germany (Bischoff and Sannemann, Reference Bischoff and Sannemann1958, pl. 13, fig. 5).
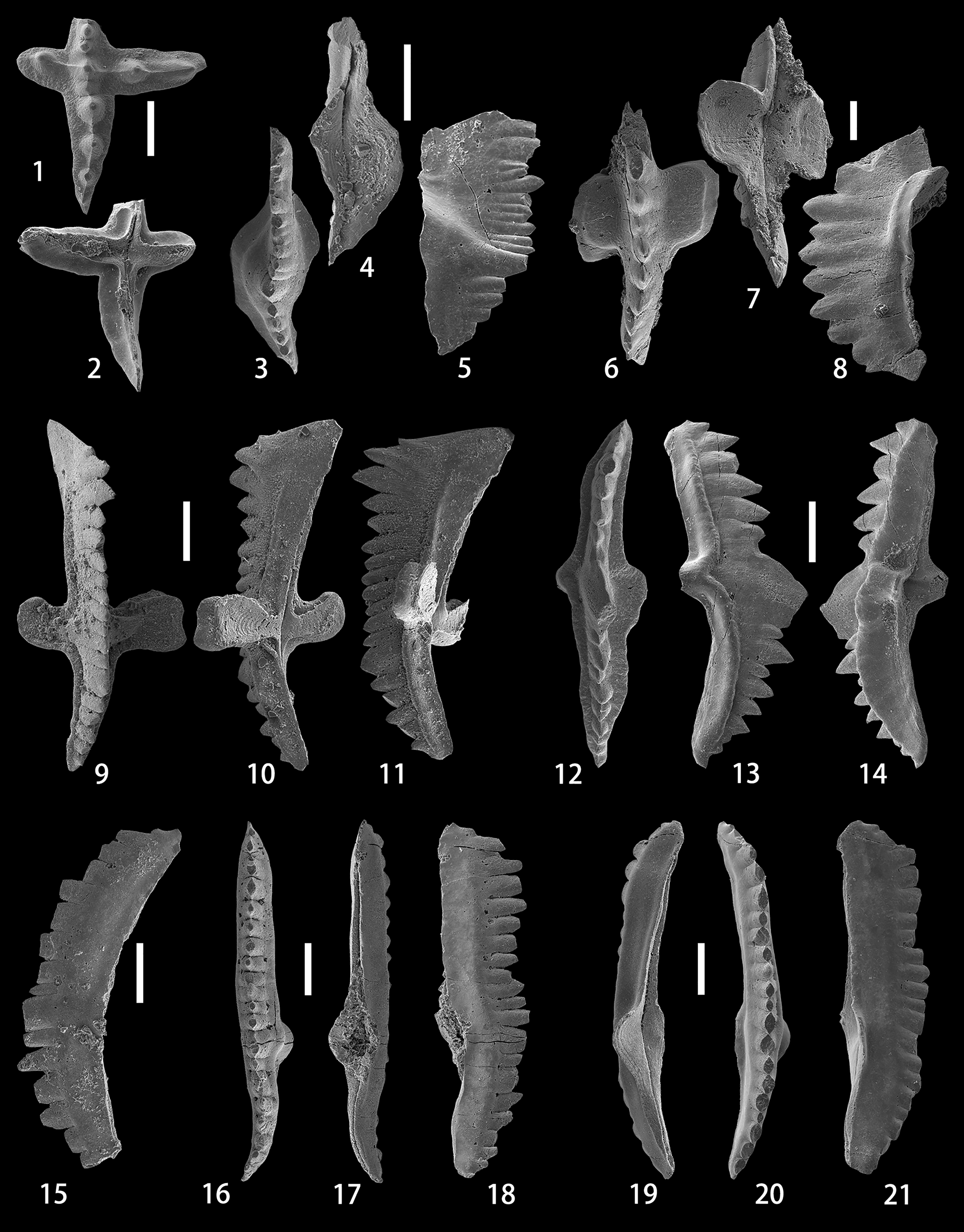
Figure 3. (1, 2) Ancyrodelloides transitans (Bischoff and Sannemann, Reference Bischoff and Sannemann1958), upper and lower views of the Pa element, NIGP 175046, sample ALC-12-6m. (3–5) Flajsella stygia (Flajs, Reference Flajs1966), upper, lower, and lateral views of the Pa element, NIGP 175047, sample ALC-12-10m. (6–8) Lanea omoalpha Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999, upper, lower, and lateral views of the Pa element, NIGP 175048, sample ALC-11-3m. (9–14) Lanea telleri (Schulze, Reference Schulze1968); (9–11) upper, lower, and lateral views of the Pa element, NIGP 175049, sample ALC-12-10m; (12–14) upper, lateral, and lower-lateral views of the Pb element, NIGP 175050, sample ALC-12-10m. (15) Wurmiella excavata (Branson and Mehl, Reference Branson and Mehl1933), lateral view of the Pa element, NIGP 175051, sample ALC-10-19m. (16–18) Wurmiella tuma (Murphy and Matti, Reference Murphy and Matti1983), upper, lower, and lateral views of the Pa element, NIGP 175052, sample ALC-11-9m. (19–21) Wurmiella aff. W. tuma (Murphy and Matti, Reference Murphy and Matti1983), lower, upper, and lateral views of the Pa element, NIGP 175053, sample ALC-12-6m. All scale bars represent 200 μm.
Description
The anterior process is broken, with the two posteriormost oval and pointed denticles preserved. The posterior process is arched downwards posteriorly, slightly deflected inwards, and consists of four stout denticles and one tiny denticle with an oval cross-section, descending in height towards the posterior end and connected by a thin ridge. The platform on the posterior process is poorly developed and bench-like shaped, while it is almost undeveloped on the remnant of the anterior process. The outer process is twice as long as the inner process. Both processes meet the longitudinal axis of the unit at an angle of ~90°. The inner process has a rounded termination and bears only one thick denticle that has an oval cross-section, whereas the outer process has two stout denticles. A thin ridge extends from the tip of the denticle on the inner process via the cusp to the two denticles on the outer process. The platform on the inner and outer processes is poorly developed and narrow. Being only slightly higher than adjacent denticles on the anterior and posterior processes, the cusp is not prominent and almost the same size as the denticles on the inner and outer processes. On the lower side, the shallow and medium-sized basal cavity expands beneath the lateral processes and almost reaches the slightly pointed terminations on both the inner and outer sides. The basal cavity continues posteriorly as a narrow groove that runs to the distal end of the unit.
Material
One specimen from sample ALC-12-6m.
Remarks
Murphy and Matti (Reference Murphy and Matti1983, p. 21, text-fig. 5) pointed out the great difference in the shape of lateral terminations of the basal cavity between A. trigonicus Bischoff and Sannemann, Reference Bischoff and Sannemann1958, and A. transitans. Ancyrodelloides trigonicus has a small pit beneath the cusp that tapers to a point laterally for a short distance; in contrast, A. transitans has a shallow or moderately deep basal cavity that is bluntly rounded laterally and may reach the terminations of lateral processes. The present specimen has a basal cavity greatly expanding beneath the lateral processes, thus resembling A. transitans; however, both the inner and outer terminations are not bluntly rounded but somewhat pointed, especially the outer one. Specimens with a similar shape of the lateral terminations of the basal cavity were also illustrated as A. transitans by Schönlaub (Reference Schönlaub1980, pl. 4, fig. 17) and Valenzuela-Ríos (Reference Valenzuela-Ríos1994a, pl. 2, figs. 5, 8, 16).
Genus Flajsella Valenzuela-Ríos and Murphy, Reference Valenzuela-Ríos and Murphy1997
Type species
Spathognathodus stygius Flajs, Reference Flajs1966.
Flajsella stygia (Flajs, Reference Flajs1966)
Figure 3.3–3.5
- Reference Flajs1966
Spathognathodus stygius Flajs, p. 204, pl. 5, figs. 16, 17.
- Reference Schulze1968
Pathognathodus seebergensis Schulze, p. 227, pl. 17, fig. 4.
- Reference Lane and Ormiston1979
Ozarkodina stygia (Flajs); Lane and Ormiston, p. 57, pl. 1, figs. 45, 46, pl. 2, figs. 10, 11.
- Reference Murphy and Matti1983
Ozarkodina stygia (Flajs); Murphy and Matti, p. 10, pl. 3, figs. 1, 2, 7, 8.
- Reference Wang and Ziegler1983
Ozarkodina stygia (Flajs); Wang and Ziegler, pl. 2, fig. 1.
- Reference Valenzuela-Ríos1994a
Ozarkodina stygia (Flajs); Valenzuela-Río, p. 66, pl. 6, figs. 17–19.
- Reference Valenzuela-Ríos and Murphy1997
Flajsella stygia (Flajs); Valenzuela-Ríos and Murphy, p. 139, figs. 8.26–8.28, ?9.17, 9.18, 9.26–9.30. [with synonymy list]
- Reference Valenzuela-Ríos and García-López1998
Flajsella stygia (Flajs); Valenzuela-Ríos and García-López, pl. 1, fig. 4.
- Reference Valenzuela-Ríos2002
Flajsella stygia (Flajs); Valenzuela-Ríos, pl. 1, fig. 8.
- Reference Suttner2007
Flajsella stygia (Flajs); Suttner, p. 31, pl. 18, figs. 12, 13.
- Reference Murphy and Valenzuela-Ríos2017
Flajsella stygia (Flajs); Murphy and Valenzuela-Ríos, pl. 1, figs. 1–5, 25, 28, 29.
- Reference Schönlaub, Corradini, Corriga and Ferretti2017
Flajsella stygia (Flajs); Schönlaub et al., pl. 3, figs. 5, 6.
- Reference Takahashi, Agematsu, Rahman and Sashida2017
Flajsella stygia (Flajs); Takahashi et al., p. 994, fig. 5.9.
- Reference Valenzuela-Ríos, Liao and Martínez-Pérez2017a
Flajsella stygia (Flajs); Valenzuela-Ríos et al., fig. 6.5.
- Reference Corradini, Corriga, Pondrelli, Serventi, Simonetto and Ferretti2019
Flajsella stygia (Flajs); Corradini et al., fig. 12G.
Holotype
Specimen 2337/115/1 from the limestone at 1280 m from the south portal of the Plöcken tunnel, Carnic Alps, Austria (Flajs, Reference Flajs1966, pl. 5, fig. 17).
Description
The anterior part of the free blade is broken; the straight posterior part clearly bends to join the cusp at the proximal end. The posterior carina joins the anterior carina at the cusp in an obvious offset. In front of the stout cusp, characterized by an elliptical cross-section, the remnant of the anterior blade consists of two comparatively wider and probably higher denticles anteriorly and nine narrower and smaller denticles posteriorly that are of approximately the same height. Therefore, the highest point of the blade probably lies in the anterior half of the anterior carina. The short posterior carina is slightly deflected inwards, and is composed of four denticles descending steeply from the cusp. The basal cavity is large and deep, with a broken inner lobe. The lower margin in the posterior part of the blade is characteristically straight.
Material
One specimen from sample ALC-12-10m.
Remarks
Although the anterior part of the anterior blade is broken, the obvious offset of the straight anterior carina and slightly deflected posterior carina at the cusp, the steeply descending posterior denticles, and the highest point of the blade probably located in the anterior half of the anterior blade permit assignment of our specimen to F. stygia. Flajsella schulzei (Bardashev, Reference Bardashev1989) differs from F. stygia in the straight anterior carina that is in line with the posterior carina, and the almost symmetrical basal cavity. Flajsella stygia also shares a close similarity in the outline of the carinae around the cusp with F. streptostygia Valenzuela-Ríos and Murphy, Reference Valenzuela-Ríos and Murphy1997, which, however, is highly distinguishable by the curved anterior carina and the angular junction of carinae at the cusp.
Genus Lanea Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999
Type species
Ozarkodina eleanorae Lane and Ormiston, Reference Lane and Ormiston1979.
Lanea omoalpha Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999
Figure 3.6–3.8
- Reference Murphy and Matti1983
Ancyrodelloides omus alpha morph Murphy and Matti, p. 16, pl. 2, figs. 18–20.
- ?Reference Mawson and Talent1994
Ancyrodelloides omus alpha morph Murphy and Matti; Mawson and Talent, p. 51, figs. 11H–L.
- Reference Valenzuela-Ríos1994a
Ancyrodelloides omus morphotype delta Valenzuela-Ríos, p. 40, pl. 1, fig. 10.
- Reference Murphy and Valenzuela-Ríos1999
Lanea omoalpha Murphy and Valenzuela-Ríos, p. 327, pl. 1, figs. 10–19, 23, 27–29, pl. 2, figs. 12–14.
- Reference Talent and Mawson1999
Ancyrodelloides omus Murphy and Matti; Talent and Mawson, p. 73, pl. 5, figs. 6, 7, 9, pl. 7, fig. 13, pl. 12, fig. 10.
- Reference Wang, Zhou, Yan, Wu, Zhao and Qian2000
Ancyrodelloides omus Murphy and Matti; Wang et al., pl. 2, fig. 5.
- Reference García-López, Sanz-López, Sarmiento, García-López and Bastida2002
Lanea omoalpha Murphy and Valenzuela-Ríos; García-López et al., pl. 3, fig. 11.
- Reference Farrell2004
Lanea omoalpha Murphy and Valenzuela-Ríos; Farrell, p. 976, pl. 8, figs. 12, 13.
- Reference Barrick, Meyer and Ruppel2005
Lanea omoalpha Murphy and Valenzuela-Ríos; Barrick et al., p. 117, pl. 1, figs. 5–8, 13, 14.
- non Reference Jin, Qian and Wang2005
Lanea omoalpha Murphy and Valenzuela-Ríos; Jin et al., pl. 2, figs. 6–8, 11, 12.
- Reference Valenzuela-Ríos, Liao, Martínez-Pérez, Castelló, Botella, Gámez, Liñán and Valenzuela-Ríos2005
Lanea omoalpha Murphy and Valenzuela-Ríos; Valenzuela-Ríos et al., pl. 1, figs. e, f.
- Reference Wang, Minjin, Ziegler, Munchtsetseg, Gereltsetseg and Undarya2005b
Lanea omoalpha Murphy and Valenzuela-Ríos; Wang et al., fig. 3.3, 3.4.
- Reference Wang2006
Lanea omoalpha Murphy and Valenzuela-Ríos; Wang, p. 214, pl. 4, figs. 1, 2, pl. 5, figs. 4, 5, 8, 9.
- non Reference Jansen, Lazreq, Plodowski, Schemm-Gregory, Schindler, Weddige, Becker and Kirchgasser2007
Lanea omoalpha Murphy and Valenzuela-Ríos; Jansen et al., fig. 3.6.
- Reference Suttner2007
Lanea omoalpha Murphy and Valenzuela-Ríos; Suttner, p. 31, pl. 19, figs. 5, 8. 10.
- Reference Wang, Wang, Yang and Xie2009
Lanea omoalpha Murphy and Valenzuela-Ríos; Wang et al., pl. 2, fig. 22.
- Reference Drygant and Szaniawski2012
Lanea omoalpha Murphy and Valenzuela-Ríos; Drygant and Szaniawski, p. 858, fig. 13M, P, T.
- Reference Slavík, Carls, Hladil and Koptíková2012
Lanea cf. omoalpha Murphy and Valenzuela-Ríos; Slavík et al., fig. 6.14.
- Reference Mavrinskaya and Slavík2013
Lanea omoalpha Murphy and Valenzuela-Ríos; Mavrinskaya and Slavík, fig. 5A–C.
- ?Reference Corriga, Corradini, Schönlaub and Pondrelli2016
Lanea omoalpha Murphy and Valenzuela-Ríos; Corriga et al., fig. 5Q.
- Reference Mathieson, Mawson, Simpson and Talent2016
Ancyrodelloides omus Murphy and Matti; Mathieson et al., p. 614, fig. 21A–F.
- Reference Schönlaub, Corradini, Corriga and Ferretti2017
Lanea omoalpha Murphy and Valenzuela-Ríos; Schönlaub et al., pl. 3, fig. 20.
- Reference Valenzuela-Ríos and Liao2017
Lanea omoalpha Murphy and Valenzuela-Ríos; Valenzuela-Ríos and Liao, fig. 4.2, 4.3.
- Reference Valenzuela-Ríos, Liao and Calvo2017b
Lanea omoalpha Murphy and Valenzuela-Ríos; Valenzuela-Ríos et al., fig. 4.8.
- Reference Corradini, Corriga, Pondrelli, Serventi, Simonetto and Ferretti2019
Lanea omoalpha Murphy and Valenzuela-Ríos; Corradini et al., fig.12Q.
- Reference Hušková and Slavík2020
Lanea omoalpha Murphy and Valenzuela-Ríos; Hušková and Slavík, fig. 7D.
- Reference Medici, Malferrari, Savioli and Ferretti2020
Lanea omoalpha Murphy and Valenzuela-Ríos; Medici et al., fig. 2H.
Holotype
Specimen UCR 8762, I/1, from the Windmill Limestone at SP VII Section, Coal Canyon, northern Simpson Park Range, Nevada, USA (Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999, pl. 1, figs. 17–19).
Description
The anterior blade is broken, only the last denticle, characterized by a lenticular cross-section, was preserved. The posterior blade consists of seven denticles that descend gradually from the second denticle to the posterior end, thus forming an arched upper profile in lateral view. Situated above the apex of the basal cavity, the cusp is slightly enlarged but not prominently high. Platform lobes are slightly asymmetrical and characteristically subquadrate-rounded, with a well-developed terrace extending from the blade at ~90° on both sides and meeting the brim, which is much smaller and narrower than the terrace, at an obtuse angle. The wide and open basal cavity extends anteriorly beneath the anterior blade as an open and deep groove, and posteriorly as an open but somewhat shallow groove that gradually tapers to the posterior termination. In lateral view, a weak pinch zone is developed close to the lower margin of the anterior and posterior blades.
Material
One specimen from sample ALC-11-3m.
Remarks
The broken specimen depicted herein is quite large (1.33 mm long) and probably represents an ontogenetically gerontic form. The arrangement of enlarged denticles on the posterior blade, the unconstricted large basal cavity, the large platform terrace that joins the brim at an obtuse angle, and the weakly developed pinch zone allow the identification of this broken specimen as L. omoalpha. The specimen closely resembles one broken specimen illustrated by Murphy and Valenzuela-Ríos (Reference Murphy and Valenzuela-Ríos1999, pl. 2, figs. 12–14) in the arrangement of denticles on the blade and the lower and upper profiles of the blade. This specimen resembles L. eoeleanorae Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999, in possessing a well-developed terrace that occupies most of the lobe surface; however, the obtuse angle junction of the terrace and brim, the clearly visible platform lobes in oral view, and the absence of a distinct brim sulcus in lateral view differ from the corresponding features of L. eoeleanorae.
Lanea telleri (Schulze, Reference Schulze1968)
Figure 3.9–3.14
- Reference Schulze1968
Spathognathodus steinhornensis telleri Schulze, p. 229, pl. 17, figs. 18, 19.
- Reference Murphy and Valenzuela-Ríos1999
Lanea telleri (Schulze); Murphy and Valenzuela-Ríos, p. 330, pl. 2, figs. 24–40. [with synonymy list]
- Reference Valenzuela-Ríos2002
Lanea telleri (Schulze); Valenzuela-Ríos, pl. 1, fig. 6.
- Reference Suttner2007
Lanea telleri (Schulze); Suttner, p. 32, pl. 19, figs. 11, 12.
- Reference Corradini, Pondrelli, Simonetto, Corriga, Spaletta, Suttner, Kido, Mossoni and Serventi2016
Lanea telleri (Schulze); Corradini et al., pl. 3, fig. 10.
- Reference Schönlaub, Corradini, Corriga and Ferretti2017
Lanea telleri (Schulze); Schönlaub et al., pl. 3, fig. 18.
- Reference Valenzuela-Ríos, Liao and Martínez-Pérez2017a
Lanea telleri (Schulze); Valenzuela-Ríos et al., fig. 6.3.
Holotype
Specimen Tüb. Co 1313/37 from P. 10/Probe 1 at Seeberggebiet, Karawanken Alps, southern Austria (Schulze, Reference Schulze1968, pl. 17, fig. 19).
Description
The robust Pa element is deflected slightly inwards in the posterior part and arched gently downwards anteriorly and posteriorly, thus the anterior and posterior ends of the lower margin are the lowest. At the anterior end of the blade, there are two high denticles, which are followed by about nine palisade-type and almost uniform (in height rather than in width) denticles anteriorly, and six gradually descending denticles posteriorly. The cusp is not prominent, but the first denticle on the posterior blade is enlarged. Located entirely posterior to the middle point of the blade, the terraced platform lobes are asymmetrical, thus the rectangular outer lobe is much wider than the semi-circular inner lobe. The anterior blade is longer than the posterior blade. The edge of the platform is rimmed by a narrow marginal ridge, which extends along margins of the anterior and posterior ledges. On the lower side, the constricted basal cavity is dumbbell shaped, extending anteriorly and posteriorly as appressed grooves.
The angulate pectiniform Pb element is as robust as the Pa element, and slightly deflected laterally at the anterior end. The posteriorly reclined cusp is extremely wide and high, anterior to which are six palisade, back-curved, and more or less uniform denticles, and posterior to which are eight irregular and comparatively smaller denticles. The anterior process is as long as the posterior one. A narrow and asymmetrical platform is developed on the inner and outer sides of the blade, and extends along the anterior and posterior processes as a bench at the base of the denticles. On the lower side, the basal cavity is also constricted and runs anteriorly and posteriorly as appressed grooves.
Material
One Pa element and one Pb element from sample ALC-12-10m.
Remarks
Our Pa element conforms to the description of the Pa element given by Murphy and Valenzuela-Ríos (Reference Murphy and Valenzuela-Ríos1999, p. 332) in having a constricted basal cavity, a rimed platform, well-developed benches along the anterior and posterior blades, and a posteriorly situated basal cavity. Murphy and Valenzuela-Ríos (Reference Murphy and Valenzuela-Ríos1999) also illustrated two specimens of Pb element from the Simpson Park Range section VII, which our Pb element closely resembles in possessing benches on the anterior and posterior processes. However, our specimen differs in having fewer but stouter denticles anterior to the wide and high cusp.
Genus Wurmiella Murphy, Valenzuela-Ríos, and Carls, Reference Murphy, Valenzuela-Ríos and Carls2004
Type species
Ozarkodina excavata tuma Murphy and Matti, Reference Murphy and Matti1983.
Wurmiella excavata (Branson and Mehl, Reference Branson and Mehl1933)
Figure 3.15
- Reference Branson and Mehl1933
Ozarkodina simplex Branson and Mehl, p. 52, pl. 3, figs. 46, 47.
- Reference Wang, Li, Rong and Dong1983
Ozarkodina excavata excavata (Branson and Mehl); Wang, p. 154, pl. 1, figs. 10, 11, 14–16, pl. 2, figs. 24, 25.
- Reference Wang, Weddige, Ziegler and Minjin2005a
Ozarkodina excavata excavata (Branson and Mehl); Wang et al., pl. 1, fig. 2.
- Reference Corriga and Corradini2009
Wurmiella excavatus (Branson and Mehl); Corriga and Corradini, fig. 4C.
- Reference Suttner2009
Ozarkodina excavata excavata (Branson and Mehl); Suttner, p. 78, pl. 1, fig. 15. [with synonymy list]
- Reference Corradini and Corriga2010
Wurmiella excavata (Branson and Mehl); Corradini and Corriga, p. 248, pl. 2, figs. 9–25.
- Reference Corriga, Corradini and Walliser2014b
Wurmiella excavata (Branson and Mehl); Corriga et al., fig. 6A.
- Reference Corradini, Pondrelli, Simonetto, Corriga, Spaletta, Suttner, Kido, Mossoni and Serventi2016
Wurmiella excavata (Branson and Mehl); Corradini et al., pl. 3, figs. 14, 15, pl. 4, figs. 1, 2.
- Reference Schönlaub, Corradini, Corriga and Ferretti2017
Wurmiella excavata (Branson and Mehl); Schönlaub et al., pl. 2, fig. 3.
- Reference Takahashi, Agematsu, Rahman and Sashida2017
Wurmiella excavata (Branson and Mehl); Takahashi et al., p. 995, fig. 5.12, 5.13.
- Reference Chen, Männik, Wang, Fang, Chen, Ma and Zhang2020
Wurmiella excavata (Branson and Mehl); Chen et al., figs. 5bb, cc, 6a–c, e, f, 7o–q, t.
- Reference Hušková and Slavík2020
Wurmiella excavata (Branson and Mehl); Hušková and Slavík, fig. 7H.
Neotype
Specimen 13151 from the Bainbridge Formation at Lithium, Missouri, USA (Rexroad and Crag, Reference Rexroad and Craig1971, pl. 80, fig. 7).
Description
Specimens of the Pa element have a long, straight or twisted blade bearing 11–15 denticles (partially broken posteriorly in some specimens), and are usually arched downwards posteriorly, resulting in a rounded lower margin in lateral view. The irregular denticles are normally stout or palisade, and erect or posteriorly reclined. The cusp is slightly or remarkably wider than other denticles and situated in the posterior half. The small lobes are slightly to clearly asymmetrical (in some specimens one lobe is more developed and laterally extended than the opposite lobe). The small and moderately deep basal cavity extends anteriorly as an open and slightly deep groove that turns to be appressed at the anterior termination of the blade, and posteriorly as an open but somewhat shallow groove that may be inverted at the posterior end. In lateral view, ledges are absent or weakly developed at the base of the denticles.
Material
Eight specimens from samples ALC-9-4m (1), ALC-10-19m (3), and ALC-10-23m (4).
Remarks
This species shares a close similarity with W. tuma (Murphy and Matti, Reference Murphy and Matti1983) and W. wurmi (Bischoff and Sannemann, Reference Bischoff and Sannemann1958). However, W. tuma differs from W. excavata in having a higher density of denticles on the blade, poorly developed cusp, and strongly developed ledges at the base of denticles. Wurmiella wurmi differs from W. excavata in possessing a biconcave or convexo-concave lower margin and well-developed ledges.
Wurmiella tuma (Murphy and Matti, Reference Murphy and Matti1983)
Figure 3.16–3.18
- Reference Wang1982
Spahognathodus wurmi Bischoff and Sannemann; Wang, p. 445, pl. 1, figs. 25–27, 29, 30.
- Reference Murphy and Matti1983
Ozarkodina excavata tuma Murphy and Matti, p. 7, pl. 1, figs. 3–9.
- Reference Murphy and Cebecioglu1986
Ozarkodina tuma Murphy and Matti; Murphy and Cebecioglu, pl. 1, fig. 1.1–1.7, 1.18–1.22.
- Reference Valenzuela-Ríos1994a
Ozarkodina tuma Murphy and Matti; Valenzuela-Ríos, p. 70, pl. 4, figs. 2, 5.
- Reference Murphy, Valenzuela-Ríos and Carls2004
Wurmiella tuma (Murphy and Matti); Murphy et al., p. 11, fig. 2.16–2.28.
Holotype
Specimen UCR8536/2 from the Bastille Limestone at the Mill Canyon Section, central Nevada, USA (Murphy and Matti, Reference Murphy and Matti1983, pl. 1, figs. 3, 4).
Description
Specimens of the Pa element are long and consist of 20 small and palisade denticles of a lenticular cross-section, of which the straight anterior blade and weakly or clearly deflected posterior blade bear 12 and 8 denticles, respectively. Two to three denticles close to the anterior end of the blade and the cusp posterior to the middle point of the blade are somewhat enlarged, but not prominently high. The lower margin is characteristically stepped. The slightly deep and asymmetrical basal cavity, with small, inflated, and shouldered lobes, is situated posterior to the mid-length of the blade. The basal cavity extends anteriorly as an open and deep groove, reaching the anterior end of the blade, and posteriorly for a short distance as a flush groove, which is followed by a clearly inverted keel to the posterior end. In lateral view, ledges are strongly developed on both sides of the blade at the base of denticles.
Material
Four specimens from samples ALC-11-9m (1), ALC-12-6m (1), and ALC-13-13m (2).
Remarks
Specimens are similar to the representative Pa elements of W. tuma in the high number of denticles on the blade, the well-developed ledges, the asymmetrical and deep basal cavity with inflated lobes, and the straight or angular lower margin. According to Murphy and Matti (Reference Murphy and Matti1983, pl. 1, figs. 3–9), representative Pa elements of W. tuma have an open and deep posterior groove tapering to the end; in contrast, the illustrated specimen herein differs slightly in the development of a short groove that turns into an inverted keel at the end of the posterior blade. On the basis of the profile of the lower margin, W. tuma differs greatly from W. wurmi.
The ledges and inner shoulder in one specimen provisionally designated here as W. aff. W. tuma (Fig. 3.19–3.21) are weakly or barely developed, as opposed to W. tuma in which the ledges and inner shoulder are strongly developed. Moreover, the basal cavity of W. aff. W. tuma is shallow and extends posteriorly as a clearly inverted keel. Both of these features differ greatly from the corresponding features of W. tuma.
Genus Zieglerodina Murphy, Valenzuela-Ríos, and Carls, Reference Murphy, Valenzuela-Ríos and Carls2004
Type species
Spathognathodus remscheidensis Ziegler, Reference Ziegler1960.
Zieglerodina eladioi (Valenzuela-Ríos, Reference Valenzuela-Ríos1994a)
Figure 4.1–4.8
- Reference Wang, Li, Rong and Dong1983
Ozarkodina remscheidensis remscheidensis (Ziegler); Wang, p. 155, pl. 2, fig. 22.
- Reference Valenzuela-Ríos1994a
Ozarkodina eladioi Valenzuela-Ríos, p. 59, pl. 5, figs. 1–35. [with synonymy list]
- Reference Sanz-López, Perret and Vachard2006
“Ozarkodina” eladioi Valenzuela-Ríos; Sanz-López et al., fig. 5.14, 5.15.
- Reference Benfrika, Bultynck and Hassani2007
Ozarkodina remscheidensis remscheidensis (Ziegler); Benfrika et al., fig. 8A.
- Reference Corriga, Corradini, Schönlaub and Pondrelli2016
Zieglerodina eladioi (Valenzuela-Ríos); Corriga et al., p. 266, fig. 5A, B.
- Reference Schönlaub, Corradini, Corriga and Ferretti2017
Zieglerodina eladioi (Valenzuela-Ríos); Schönlaub et al., pl. 3, fig. 2.
- Reference Corriga and Corradini2019a
Zieglerodina eladioi (Valenzuela-Ríos); Corriga and Corradini, p. 182, figs. 2, 3.
- Reference Corradini, Corriga, Pondrelli and Suttner2020
Zieglerodina eladioi (Valenzuela-Ríos); Corradini et al., fig. 12C.
Holotype
Specimen MPZ 8072 from Bed 6A of Litosoma A at the Gerri 1.1 Section, Spanish Central Pyrenees (Valenzuela-Ríos, Reference Valenzuela-Ríos1994a, pl. 5, fig. 1).

Figure 4. (1–8) Zieglerodina eladioi (Valenzuela-Ríos, 1994a); (1–3) upper, lateral, and lower-lateral views of the Pa element, NIGP 175054, sample ALC-10-19m; (4, 5) upper and lower-lateral views of the Pa element, NIGP 175055, sample ALC-10-23m; (6–8) upper, lateral, and lower-lateral views of the Pa element, NIGP 175056, sample ALC-10-23m. (9–14) Zieglerodina planilingua (Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999); (9–11) upper, lateral, and lower views of the Pa element, NIGP 175057, sample ALC-9-10m; (12–14) upper, lateral, and lower views of the Pa element, NIGP 175058, sample ALC-10-23m. (15–22) Zieglerodina remscheidensis (Ziegler, Reference Ziegler1960); (15, 16) upper and lateral views of the Pa element, NIGP 175059, sample ALC-10-2m; (17–19) upper, lateral, and lower views of the Pa element, NIGP 175060, sample ALC-10-19m, the width and height of the cusp are reduced because of breakage; (20–22) upper, lateral, and lower views of the Pa element, NIGP 175061, sample ALC-10-23m. All scale bars represent 200 μm.
Description
Specimens of the Pa element have a short posterior blade bearing four denticles, the anteriormost one of which is usually small and followed by two higher and larger denticles before a lowest and small denticle at the posterior end. The anterior blade constitutes 5–6 denticles that are of similar size and of more or less the same height, except for the anteriormost one or two, which may be slightly higher. The high cusp is almost twice as wide as adjacent denticles, but only slightly wider than the stoutest denticle on the posterior blade. Normally, the axis of the blade is straight. The basal cavity, which is situated posterior to the middle length of the blade, has two wide, asymmetrical, and strongly expanded lateral lobes. The basal cavity extends anteriorly as a narrow and deep groove reaching the anterior end of the blade, and as a wide and deep groove tapering towards the posterior end.
Material
Seven specimens from samples ALC-10-15m (1), ALC-10-19m (2), and ALC-10-23m (4).
Remarks
According to the original description introduced by Valenzuela-Ríos (Reference Valenzuela-Ríos1994a), Z. eladioi is mainly characterized by the short posterior blade possessing two larger denticles, a comparably longer anterior blade whose denticles vary little in size, and two wide, slightly asymmetrical lobes, all of which are clearly observed in our specimens. A great many specimens of this species were previously identified as Z. remscheidensis (Ziegler, Reference Ziegler1960), which, however, has distinctly high and strong denticles at the anterior end of the blade and more numerous denticles on the blade. Zieglerodina mashkovae (Drygant, Reference Drygant1984) differs greatly from Z. eladioi in possessing straight upper and lower margins, a symmetrical and approximately centrally situated basal cavity, numerous (5–7) denticles of nearly equal height on the anterior and posterior blades, and a large cusp. Zieglerodina shoenlaubi Corradini et al., Reference Corradini, Corriga, Pondrelli, Serventi, Simonetto and Ferretti2019, which resembles Z. eladioi in possessing a short posterior blade, is distinguishable by having small denticles alternating with large ones on the blade.
Zieglerodina planilingua (Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999)
Figure 4.9–4.14
- Reference Wang1982
Spathognathodus remscheidensis Ziegler; Wang, p. 444, pl. 1, figs. 13, 14.
- Reference Murphy and Valenzuela-Ríos1999
“Ozarkodina” planilingua Murphy and Valenzuela-Ríos, p. 326, pl. 1, figs. 1–9.
- Reference García-López, Sanz-López, Sarmiento, García-López and Bastida2002
Ozarkodina planilingua Murphy and Valenzuela-Ríos; García-López et al., pl. 3, fig. 10.
- Reference Farrell2003
“Ozarkodina” planilingua Murphy and Valenzuela-Ríos; Farrell, p. 137, pl. 7, figs. 20–23.
- Reference Barrick, Meyer and Ruppel2005
Ozarkodina planilingua Murphy and Valenzuela-Ríos; Barrick et al., p. 120, pl. 1, figs. 3, 4, 11, 12.
- ?Reference Wang, Weddige, Ziegler and Minjin2005a
“Ozarkodina” planilingua Murphy and Valenzuela-Ríos; Wang et al., pl. 1, figs. 10, 15.
- Reference Wang, Minjin, Ziegler, Munchtsetseg, Gereltsetseg and Undarya2005b
“Ozarkodina” planilingua Murphy and Valenzuela-Ríos; Wang et al., fig. 3.5, 3.6.
- Reference Wang2006
Lanea omoalpha Murphy and Valenzuela-Ríos; Wang, p. 214, pl. 5, figs. 12–15.
- Reference Corriga and Corradini2009
Ozarkodina planilingua Murphy and Valenzuela-Ríos; Corriga and Corradini, fig. 4J, K.
- Reference Corradini and Corriga2010
Ozarkodina planilingua Murphy and Valenzuela-Ríos; Corradini and Corriga, pl. 3, figs. 14–16.
- Reference Drygant and Szaniawski2012
Zieglerodina planilingua (Murphy and Valenzuela-Ríos); Drygant and Szaniawski, p. 856, fig. 13A–D.
- Reference Corriga, Corradini, Haude and Walliser2014a
Zieglerodina planilingua (Murphy and Valenzuela-Ríos); Corriga et al., fig. 3C.
- Reference Corriga, Corradini and Walliser2014b
Zieglerodina planilingua (Murphy and Valenzuela-Ríos); Corriga et al., p. 197, fig. 6H–L.
- Reference Corradini, Pondrelli, Simonetto, Corriga, Spaletta, Suttner, Kido, Mossoni and Serventi2016
Zieglerodina planilingua (Murphy and Valenzuela-Ríos); Corradini et al., pl. 3, fig. 12.
- Reference Corriga, Corradini, Schönlaub and Pondrelli2016
Zieglerodina planilingua (Murphy and Valenzuela-Ríos); Corriga et al., fig. 5O.
- Reference Zhen, Hegarty, Percival and Pickett2017
“Ozarkodina” planilingua Murphy and Valenzuela-Ríos; Zhen et al., fig. 5a, ?b.
- Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018
Lanea planilingua (Murphy and Valenzuela-Ríos); Wang et al., fig. 4A–F.
- Reference Hušková and Slavík2020
Lanea planilingua (Murphy and Valenzuela-Ríos); Hušková and Slavík, fig. 7E.
- Reference Medici, Malferrari, Savioli and Ferretti2020
Zieglerodina planilingua (Murphy and Valenzuela-Ríos); Medici et al., fig. 2L.
- Reference Corradini, Corriga, Pondrelli and Suttner2020
Zieglerodina planilingua (Murphy and Valenzuela-Ríos); Corradini et al., fig. 12J.
Holotype
Specimen UCR 8758, I/3, from the Windmill Limestone at SP VII Section, Coal Canyon, northern Simpson Park Range, Nevada, USA (Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999, pl. 1, figs. 7–9).
Description
Specimens of the Pa element are usually straight and consist of 11–16 stout denticles of a lenticular cross-section. At the anterior end of the blade, there are 2–3 high denticles followed by 3–5 smaller and lower denticles before the prominent and enlarged cusp. Posterior to the cusp are one or two small denticles, which are succeed posteriorly by three larger and higher denticles. The denticle at the posterior end of the blade is the lowest. The slightly asymmetrical, semi-circular or subquadrate-rounded lobes have a small terrace that extends from the blade at various angles and meet the brim at an obtuse angle, and are positioned slightly posterior to the middle length of the blade. In all specimens, the brim surface of the lobe is larger than the terrace. The basal cavity is wide, moderately deep, and unconstricted, extending anteriorly and posteriorly as deep grooves that taper gradually to distal ends. In lateral view, a weak pinch zone is present close to the lower margin of the blade.
Material
Seven specimens from samples ALC-9-10m (1), ALC-10-6m (1), and ALC-10-23m (5).
Remarks
The arrangement of the denticles on the blade, the outline of the basal cavity, and the development and shape of the terrace allow us to assign our specimens to Z. planilingua. This species is morphologically like Z. remscheidensis and L. omoalpha, the former of which does not possess the wide basal cavity and the characteristic small terrace on lobes, whereas the latter of which has a more developed and larger terrace than Z. planilingua. Moreover, the arrangement of denticles on the posterior blade in L. omoalpha is different from that in Z. planilingua.
Zieglerodina remscheidensis (Ziegler, Reference Ziegler1960)
Figure 4.15–4.22
- Reference Ziegler1960
Spathognathodusremscheidensis Ziegler, p. 194, pl. 13, figs. 1, 2, 4, 5, 7, 8, 10, 14.
- Reference Walliser1964
Spathognathodus steinhornensis remscheidensis Ziegler; Walliser, p. 87, pl. 9, fig. 24.
- non Reference Clark and Ethington1966
Spathognathodusremscheidensis Ziegler; Clark and Ethington, pl. 84, figs. 12, 14.
- non Reference Fåhræus1969
Spathognathodus steinhornensisremscheidensis Ziegler; Fåhræus, pl. 1, figs. 12, 15, 16.
- non Reference Druce1970
Spathognathodusremscheidensis Ziegler; Druce, p. 57, pl. 8, fig. ?4, pl. 9, fig. 9.
- non Reference Barnett1971
Spathognathodusremscheidensis Ziegler; Barnett, p. 288, pl. 35, figs. 1–20, pl. 36, figs. 1–24.
- Reference Link and Druce1972
Spathognathodus steinhornensis Ziegler; Link and Druce, p. 92, pl. 10, figs. 1–7, text-fig. 60.
- Reference Mashkova1972
Spathognathodus steinhornensisremscheidensis Ziegler; Mashkova, p. 83, pl. 2, fig. 20.
- non Reference Savage1973
Spathognathodusremscheidensis Ziegler; Savage, p. 329, pl. 34, figs. 9–29, 33–42, text-fig. 28.
- Reference Klapper and Murphy1975
Ozarkodina remscheidensis remscheidensis (Ziegler); Klapper and Murphy, p. 41, pl. 7, figs. 22, 25–30.
- non Reference Carls1975
Ozarkodina remscheidensis (Ziegler); Carls, pl. 2, figs. 16–18.
- non Reference Mehrtens and Barnett1976
Ozarkodina remscheidensis remscheidensis (Ziegler); Mehrtens and Barnett, p. 497, pl. 1, figs. 7, 10.
- Reference Chatterton and Perry1977
Ozarkodina remscheidensis remscheidensis (Ziegler); Chatterton and Perry, p. 786, pl. 3, figs. 28–32; pl. 4, fig. 36.
- non Reference Wang1979
Spathognathodus cf.remscheidensis Ziegler; Wang, p. 403, pl. 1, figs. 23, 24.
- Reference Chlupáč, Kriz and Schönlaub1980
Ozarkodina remscheidensis remscheidensis (Ziegler); Chlupáč et al., pl. 18, figs. 11, 14–26, pl. 19, figs. 7, 13, 14, 18–21.
- non Reference Pickett1980
Ozarkodina remscheidensis (Ziegler); Pickett, fig. 13D–G.
- Reference Schönlaub1980
Ozarkodina remscheidensis remscheidensis (Ziegler); Schönlaub, pl. 1, figs. 8, ?15, pl. 3, fig. 21, pl. 4, fig. 2, pl. 6, fig. 4, pl. 7, fig. 29.
- Reference Serpagli and Mastandrea1980
Ozarkodina remscheidensis remscheidensis (Ziegler); Serpagli and Mastandrea, figs. 8, 12.
- Reference Uyeno1980
Ozarkodina remscheidensis remscheidensis (Ziegler); Uyeno, p. 41, pl. 3, figs. 6, 7.
- non Reference Wang1981
Spathognathodus remscheidensis Ziegler; Wang, p. 81, pl. 1, figs. 1–6.
- non Reference Savage1982
Ozarkodina remscheidensis (Ziegler); Savage, p. 986, pl. 1, figs. 1–8, pl. 2, figs. 21–26.
- Reference Wang1982
Spathognathodus remscheidensis Ziegler; Wang, p. 444, pl. 1, fig. 19.
- Reference Wang, Li, Rong and Dong1983
Ozarkodina remscheidensis remscheidensis (Ziegler); Wang, p. 155, pl. 2, fig. 26.
- Reference Murphy and Matti1983
Ozarkodina remscheidensis (Ziegler); Murphy and Matti, pl. 2, figs. 4–6.
- Reference Wang and Ziegler1983
Ozarkodina remscheidensis (Ziegler); Wang and Ziegler, fig. 2.21.
- Reference Mastandrea1985a
Ozarkodina remscheidensis remscheidensis (Ziegler); Mastandrea, p. 252, pl. 1, figs. 1–3.
- Reference Mastandrea1985b
Ozarkodina remscheidensis remscheidensis (Ziegler); Mastandrea, pl. 1, fig. 1.
- Reference Schönlaub1985
Ozarkodina remscheidensis remscheidensis (Ziegler); Schönlaub, pl. 1, figs. 8, 9, 15.
- Reference Mawson1986
Ozarkodina remscheidensis remscheidensis (Ziegler); Mawson, p. 49, pl. 6, fig. 3.
- Reference Kuwano1987
Ozarkodina remscheidensis (Ziegler); Kuwano, pl. 2, figs. 2, 3.
- non Reference Li1987
Ozarkodina remscheidensis remscheidensis (Ziegler); Li, p. 364, pl. 164, figs. 1–4.
- non Reference Jeppsson1989
Ozarkodina steinhornensisremscheidensis Ziegler; Jeppsson, p. 28, pl. 2, figs. 6–11.
- non Reference Sorentino1989
Ozarkodina remscheidensis remscheidensis (Ziegler); Sorentino, p. 93, pl. 3, figs. 1–9, 12, 13.
- Reference Wilson1989
Ozarkodina remscheidensis remscheidensis (Ziegler); Wilson, p. 139, pl. 11, figs. 8, 9.
- non Reference Bischoff and Argent1990
Ozarkodina remscheidensis remscheidensis (Ziegler); Bischoff and Argent, p. 457, pl. 4, figs. 1, ?2.
- Reference Olivieri and Serpagli1990
Ozarkodina remscheidensis remscheidensis (Ziegler); Olivieri and Serpagli, pl. 4, fig. 10.
- Reference Weyant and Morzadec1990
Ozarkodina remscheidensis (Ziegler); Weyant and Morzadec, pl. 1, figs. 6, 7.
- non Reference Uyeno1990
Ozarkodina remscheidensis remscheidensis (Ziegler); Uyeno, p. 92, pl. 4, figs. 4–6, pl. 5, figs. 1–3, 30, 68, pl. 6, fig. 11, pl. 13, figs. 11, 12, 20, pl. 15, figs. 27, 28, 32–34, pl. 6, figs. 34, 35, pl. 17, figs. 10, 32.
- non Reference Uyeno1991
Ozarkodina remscheidensis remscheidensis (Ziegler); Uyeno, pl. 1, fig. 13.
- non Reference Barrick and Klapper1992
Ozarkodina remscheidensis remscheidensis (Ziegler); Barrick and Klapper, p. 49, pl. 4, figs. 8, 9, pl. 6, figs. 5–16.
- non Reference Bardashev and Ziegler1992
Ozarkodina remscheidensis remscheidensis (Ziegler); Bardashev and Ziegler, pl. 1, figs. 5, 6.
- Reference Mawson and Talent1994
Ozarkodina remscheidensis remscheidensis (Ziegler); Mawson and Talent, fig. 13O.
- Reference Valenzuela-Ríos1994a
Ozarkodina remscheidensis remscheidensis (Ziegler); Valenzuela-Ríos, p. 55. [with synonymy list]
- Reference Barrick and Noble1995
Ozarkodina remscheidensis remscheidensis (Ziegler); Barrick and Noble, fig. 6.9, 6.12, 6.15, 6.16.
- non Reference Colquhoun1995
Ozarkodina remscheidensis remscheidensis (Ziegler); Colquhoun, pl. 1, figs. 9, 10.
- non Reference Sloan, Talent, Mawson, Simpson, Brock, Engelbretsen, Jell, Aung, Pfaffenritter, Trotter and Withnall1995
Ozarkodina remscheidensis remscheidensis (Ziegler); Sloan et al., pl. 12, figs. 9, 10.
- Reference Miller and Aldridge1997
Ozarkodina remscheidensis remscheidensis (Ziegler); Miller and Aldridge, p. 46, pl. 1, figs. 21, 24.
- non Reference Talent and Mawson1999
Ozarkodina remscheidensis remscheidensis (Ziegler); Talent and Mawson, pl. 5, figs. 13–18, pl. 7, fig. 12, pl. 8, fig. 1.
- non Reference Benfrika1999
Ozarkodina remscheidensis remscheidensis (Ziegler); Benfrika, p. 315, pl. 1, fig. 6.
- Reference García-López, Sanz-López, Sarmiento, García-López and Bastida2002
Ozarkodina remscheidensis (Ziegler); García-López et al., pl. 2, figs. 19, 20.
- Reference Farrell2003
Ozarkodina remscheidensis remscheidensis (Ziegler); Farrell, p. 137, pl. 8, figs. 1–4, 6, 8–15.
- Reference Mawson, Talent, Molloy and Simpson2003
Ozarkodina remscheidensis remscheidensis (Ziegler); Mawson et al., p. 90, pl. 2, figs. 2, 4–6, 10.
- Reference Farrell2004
Ozarkodina remscheidensis remscheidensis (Ziegler); Farrell, p. 974, pl. 10, figs. 2–4, 7–9.
- Reference Murphy, Valenzuela-Ríos and Carls2004
Zieglerodina remscheidensis (Ziegler); Murphy et al., p. 13, fig. 3.1–3.9, 3.11, ?3.14–3.16, 3.19, 3.20–3.25.
- non Reference Barrick, Meyer and Ruppel2005
Ozarkodina remscheidensis (Ziegler); Barrick et al., p. 120, pl. 1, figs. 2, 9, 10.
- Reference Jin, Qian and Wang2005
Ozarkodina remscheidensis remscheidensis (Ziegler); Jin et al., pl. 2, figs. 9, 10.
- non Reference Trotter and Talent2005
Ozarkodina remscheidensis remscheidensis (Ziegler); Trotter and Talent, p. 42, pl. 18, figs. 1, 3.
- ?Reference Corradini, Simonetto, Serventi, Rigo and Calligaris2005
Ozarkodina remscheidensis remscheidensis (Ziegler); Corradini et al., fig. 5b.
- Reference Wang2006
Ozarkodina remscheidensis remscheidensis (Ziegler); Wang, p. 212, pl. 5, figs. 10, 11.
- non Reference Benfrika, Bultynck and Hassani2007
Ozarkodina remscheidensis remscheidensis (Ziegler); Benfrika et al., fig. 8A.
- Reference Suttner2007
Ozarkodina remscheidensis remscheidensis (Ziegler); Suttner, p. 41, pl. 17, figs. 7, 8, 10.
- non Reference Gaetani, Mawson, Sciunnach and Talent2008
Ozarkodina remscheidensis remscheidensis (Ziegler); Gaetani et al., p. 281, pl. 1, fig. 1.
- Reference Suttner2009
Ozarkodina remscheidensis remscheidensis (Ziegler); Suttner, p. 79, pl. 1, figs. 9, ?10.
- Reference Corradini and Corriga2010
Zieglerodina remscheidensis (Ziegler); Corradini and Corriga, pl. 3, figs. 10, 11, ?12, 13.
- Reference Corradini and Corriga2012
Zieglerodina remscheidensis (Ziegler); Corradini and Corriga, fig. 6H.
- Reference Drygant and Szaniawski2012
Zieglerodina remscheidensis (Ziegler); Drygant and Szaniawski, p. 853, fig. 12A–G.
- Reference Corriga, Corradini, Haude and Walliser2014a
Zieglerodina remscheidensis (Ziegler); Corriga et al., fig. 3G.
- Reference Corriga, Corradini and Walliser2014b
Zieglerodina remscheidensis (Ziegler); Corriga et al., fig. 6F, G.
- Reference Corradini, Pondrelli, Simonetto, Corriga, Spaletta, Suttner, Kido, Mossoni and Serventi2016
Zieglerodina remscheidensis (Ziegler); Corradini et al., pl. 3, figs. 20–22.
- Reference Corriga, Corradini, Schönlaub and Pondrelli2016
Zieglerodina remscheidensis (Ziegler); Corriga et al., fig. 5I, J.
- Reference Mathieson, Mawson, Simpson and Talent2016
Ozarkodina remscheidensis (Ziegler); Mathieson et al., p. 634, figs. 27A–F, I, J, U, V, 32K.
- non Reference Zhen, Hegarty, Percival and Pickett2017
Zieglerodina remscheidensis (Ziegler); Zhen et al., fig. 5c, d.
- Reference Corradini, Corriga, Pondrelli and Suttner2020
Zieglerodina remscheidensis (Ziegler); Corradini et al., fig. 12H.
Holotype
Specimen G88b from the “Ockrige Kalke” of the basal part of the Hüinghäuser beds, Untenrüden Bed e, Rhenish Slate Mountains, Germany (Ziegler, Reference Ziegler1960, pl. 13, fig. 4).
Description
Specimens of the Pa element are normally straight or weakly deflected, and made up of 12–15 palisade or triangular denticles. At the anterior end of the blade is a small denticle, which is followed by one to two very high and wide denticles. The cusp is stout and prominent, but is not as high as the anteriormost denticles. Denticles between the high anteriormost part and the cusp are comparatively lower and smaller. The posterior blade possesses 5–6 denticles that vary in size, but all are lower than those at the anteriormost part of the blade. Two to three denticles posterior to the cusp are much smaller and lower, posterior to which is a high denticle, almost equal in height to the cusp. The basal cavity, occupying approximately one-quarter of the length of the blade, lies posterior to the middle point of the blade. Normally, it has rounded and symmetrical lobes that bear no ornamentation; however, an incipient and small terrace-like structure is present in some specimens (Fig. 4.15). Beneath the anterior and posterior blades, a narrow groove extends to distal ends of the blade. The lower margin is more or less straight, but the anterior lower margin may rise diagonally.
Material
Eleven specimens from samples ALC-10-2m (2), ALC-10-15m (2), ALC-10-19m (2), and ALC-10-23m (5).
Remarks
According to the original description given by Ziegler (Reference Ziegler1960), Z. remscheidensis is mainly characterized by having 1–2 denticles behind the anterior edge and a single denticle (the cusp) that is especially high above the basal cavity, and a basal cavity positioned in the anterior part of the posterior half of the blade.
Since first named by Ziegler (Reference Ziegler1960), Z. remscheidensis has been widely reported from the uppermost Silurian to the lowermost Devonian around the world. However, a great number of previously illustrated specimens do not conform to the representative specimens of this species, but have a centrally situated basal cavity and denticles on the blade of little variance in size and of the same height. In the establishment of the new genus Zieglerodina, Murphy et al. (Reference Murphy, Valenzuela-Ríos and Carls2004) emphasized the common characteristics of this species and restricted its diagnosis to specimens morphologically resembling the holotype (Ziegler, Reference Ziegler1960, pl. 13, fig. 4). In the present paper, the taxonomic revision proposed by Murphy et al. (Reference Murphy, Valenzuela-Ríos and Carls2004) is adopted; therefore, the identification of specimens with more or less regular denticulation or a centrally positioned basal cavity as Z. remscheidensis is not accepted herein (for details, see the synonymy list).
Order Prioniodontida Dzik, Reference Dzik1976
Family Icriodontidae Müller and Müller, Reference Müller and Müller1957
Genus Caudicriodus Bultynck, Reference Bultynck1976
Description
The I element has a narrow and long spindle that bears 4–7 transverse ridges posterior to the anteriormost denticle, is more or less straight, and gradually narrows anteriorly in the anterior two-thirds, but keeps a constant width in the posterior one-third, thus resulting in a carrot-shaped outline. Median row denticles are distinct only in small specimens, but are completely fused with lateral row denticles in large ones. A weak or thin longitudinal crest runs from the anterior end of the spindle to the posterior end along the middle-line of the spindle. The transverse ridges in the posterior part of the spindle are separated by a constant and equal space, which becomes much greater in the anterior half of the spindle, especially between the first and second transverse ridges. Posterior to the spindle, two laterally compressed denticles are higher than those on the spindle. The first one is slightly or greatly smaller, from which an anteriorly directed and unornamented inner spur emanates. The posteriormost denticle is more prominent, from which a posteriorly directed process branches to the lateral side at an angle of ~120° with the main axis of the spindle. This outer process, which is moderately long (Fig. 5.7) or commonly broken (Fig. 5.1, 5.4), is denticulate and bears a well-developed thin ridge. On the lower side, the basal cavity is wide and deeply excavated in the posterior part.
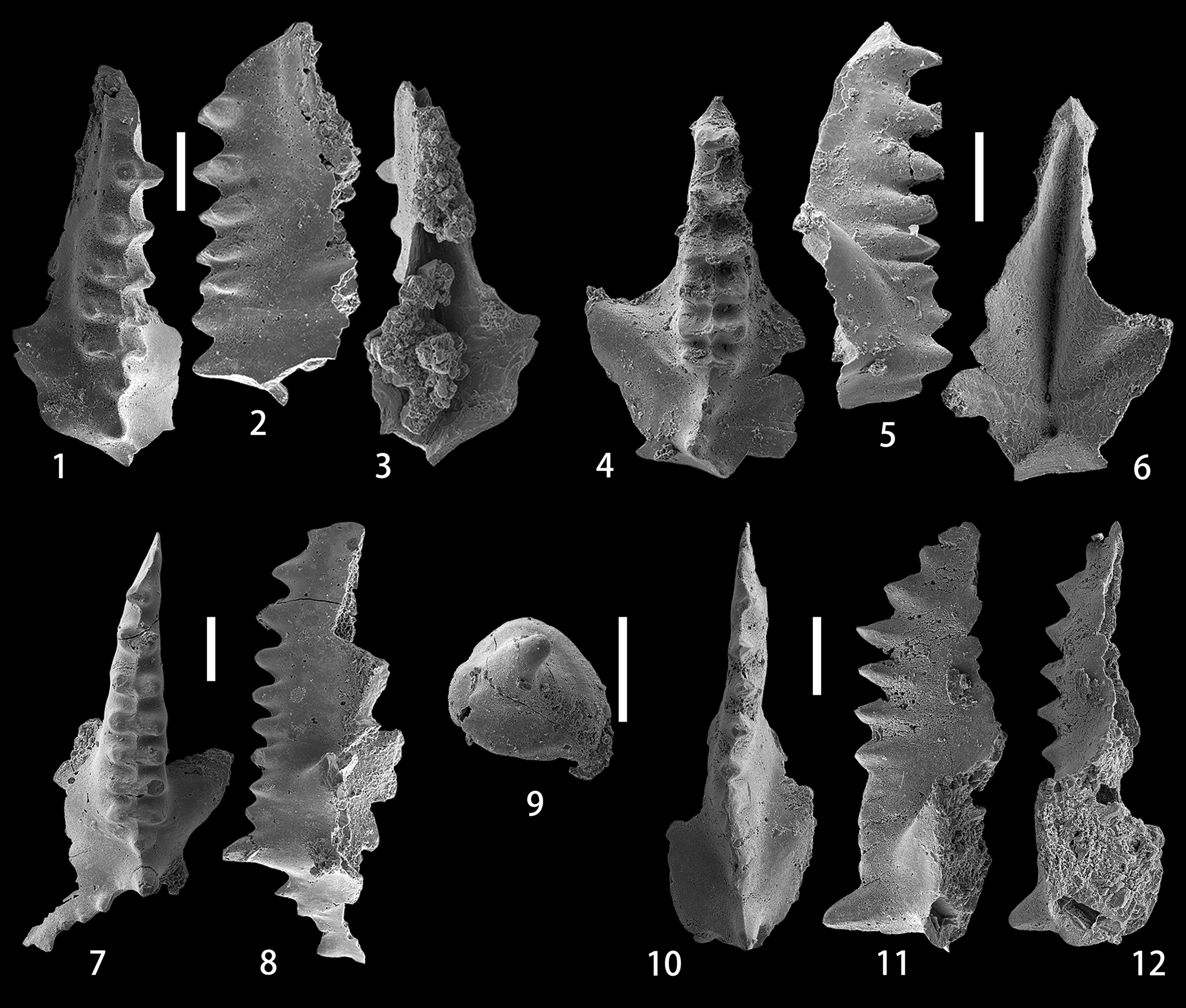
Figure 5. (1–8) Caudicriodus sp. B; (1–3) upper, lateral, and lower views of the I element, NIGP 175062, sample ALC-9-10m; (4–6) upper, lateral, and lower views of the I element, NIGP 175063, sample ALC-10-6m; (7, 8) upper and lateral views of the I element, NIGP 175064, sample ALC-10-19m. (9) Pseudooneotodus beckmanni (Bischoff and Sannemann, Reference Bischoff and Sannemann1958), upper view, NIGP 175065, sample ALC-10-10m. (10–12) Pelekysgnathus sp. A, upper, lateral, and lower views of the I element, NIGP 175066, sample ALC-13-16m. All scale bars represent 200 μm.
Material
Fifteen specimens from samples ALC-9-10m (5), ALC-10-2m (2), ALC-10-6m (4), ALC-10-10m (1), ALC-10-19m (1), and ALC-10-23m (2).
Remarks
Our specimens are characterized by the numerous fused transverse ridges on the spindle, a feature normally observed in representative I elements of C. hesperius (Klapper and Murphy, Reference Klapper and Murphy1975) and C. woschmidti (Ziegler, Reference Ziegler1960). Unlike C. woschmidti, which has a short spindle with wide transverse ridges and a relatively restricted basal cavity, the Alengchu specimens possess a much narrower and longer spindle and a comparatively larger basal cavity. Caudicriodus hesperius is easily distinguishable by the extremely long posteriorly directed lateral process; in contrast, the posterior process is normally broken in most of the specimens (Fig. 5.1, 5.4) or much shorter in the intact specimen (Fig. 5.7). Moreover, the basal cavity in our specimens is less expanded than that of C. hesperius.
Genus Pelekysgnathus Thomas, Reference Thomas1949
Type species
Pelekysgnathus inclinatus Thomas, Reference Thomas1949.
Pelekysgnathus sp. A
Figure 5.10–5.12
Description
The specimen of the I element has a long and low blade bearing six small denticles in the anterior part, which is followed by an undenticulate and fused area that takes almost one-third of the length of the unit before a high, enlarged, and erect cusp at the posterior end. Three denticles at the anteriormost part are clearly proclined, whereas the following three denticles are more or less erect and of the same height. In upper view, the longitudinal axis of the blade is slightly sigmoidal. A weak and anteriorly directed spur runs diagonally from the cusp. On the lower side, the basal cavity is narrow in the anterior half of the blade and abruptly widens in the posterior half.
Material
One specimen from sample ALC-13-16m.
Remarks
The longer and lower blade in our specimen is similar to that of P. serratus elongatus Carls and Gandl, Reference Carls and Gandl1969. However, in contrast to the latter subspecies, which is distinguished by the distinctly erect denticles in the anterior part and normally two posteriorly reclined cusps in the posterior part, the Alengchu specimen has three proclined denticles at the anterior part of the blade, an undenticulate area in the posterior half of the blade, and one erect cusp. These features readily distinguish this specimen from P. serratus serratus Jentzsch, Reference Jentzsch1962.
Order Protopanderontida Sweet, Reference Sweet1988
Family Protopanderodontidae Lindström, Reference Lindström1970
Genus Pseudooneotodus Drygant, Reference Drygant1974
Type species
Oneotodus? beckmanni Bischoff and Sannemann, Reference Bischoff and Sannemann1958.
Pseudooneotodus beckmanni (Bischoff and Sannemann, Reference Bischoff and Sannemann1958)
Figure 5.9
- Reference Bischoff and Sannemann1958
Oneotodus? beckmanni Bischoff and Sannemann, p. 98, pl. 15, figs. 22–25.
- Reference Corradini2007
Pseudooneotodus beckmanni (Bischoff and Sannemann); Corradini, p. 142, pl. 1, figs. 1–7. [with synonymy list]
- Reference Corriga, Corradini and Walliser2014b
Pseudooneotodus beckmanni (Bischoff and Sannemann); Corriga et al., fig. 5L.
- Reference Han, Zhang, Wang, Wu and Hou2014
Pseudooneotodus beckmanni (Bischoff and Sannemann); Han et al., pl. 2, figs. 4–10.
- Reference Corradini, Pondrelli, Simonetto, Corriga, Spaletta, Suttner, Kido, Mossoni and Serventi2016
Pseudooneotodus beckmanni (Bischoff and Sannemann); Corradini et al., pl. 3, fig. 18.
- Reference Corriga, Corradini, Schönlaub and Pondrelli2016
Pseudooneotodus beckmanni (Bischoff and Sannemann); Corriga et al., fig. 5S.
- Reference Mathieson, Mawson, Simpson and Talent2016
Pseudooneotodus beckmanni (Bischoff and Sannemann); Mathieson et al., p. 597, fig. 8Q–S. [with synonymy list]
- Reference Schönlaub, Corradini, Corriga and Ferretti2017
Pseudooneotodus beckmanni (Bischoff and Sannemann); Schönlaub et al., pl. 3, fig. 17.
- Reference Takahashi, Agematsu, Rahman and Sashida2017
Pseudooneotodus beckmanni (Bischoff and Sannemann); Takahashi et al., p. 995, fig. 5.4.
- Reference Corradini, Corriga, Pondrelli, Serventi, Simonetto and Ferretti2019
Pseudooneotodus beckmanni (Bischoff and Sannemann); Corradini et al., fig. 12S.
Holotype
Specimen BiSa 1958/85 from the Tentakulitenkalk at Schübelebene near Elbersreuth, Frankenwald, Germany (Bischoff and Sannemann, Reference Bischoff and Sannemann1958, pl. 15, fig. 25).
Description
The short and low conical element has a single apical denticle that is posteriorly reclined. The base of the unit is broad and subtriangular with rounded corners. The lateral surface is distinctly smooth.
Material
Five specimens from samples ALC-10-10m (1), ALC-10-19m (2), and ALC-10-23m (2).
Remarks
The morphological variation of P. beckmanni has been extensively studied by Corradini (Reference Corradini2007), who also disputed Barrick's (Reference Barrick1977) apparatus reconstruction of this species and further suggested, on the basis of material from Sardinia and the Carnic Alps, that this species has a unimembrate apparatus. The broad, subtriangular outline of the base and the single curved tip allow the assignation of our specimens to this species.
Conodont biostratigraphy
In the present study, only 14 samples yielded conodont elements, of which the state of preservation is mostly poor. In general, the abundance is low throughout the lower part of the Shanjiang Formation. All samples from the uppermost part of the underlying Baizitian Formation were completely barren, whereas 64 well-preserved specimens that are assignable to 12 species belonging to 8 genera (Ancyrodelloides, Flajsella, Lanea, Wurmiella, Zieglerodina, Caudicriodus, Pelekysgnathus, and Pseudooneotodus) were obtained from the lower part of the Shanjiang Formation (Table 1; Fig. 2). Based on the stratigraphic ranges of conodonts, one chronostratigraphic interval (the lower Lochkovian) and one conodont zone (the middle Lochkovian transitans-pandora β Zone) are recognized from the lower part of the Shanjiang Formation at the Alengchu section (Fig. 2). The binomial nomenclatural system for time-stratigraphic units described by Murphy (Reference Murphy1977) is applied herein to parts of the investigated interval.
The interval ranging from the uppermost part of Bed 9 to the upper part of Bed 10 belongs to the lower Lochkovian. Caudicriodus sp. B is recorded in sample ALC-9-10m, which is located ~10 m above the lower boundary of Bed 9, and ranges upwards into sample ALC-10-23m, which is located 23 m above the lower boundary of Bed 10. Three samples were also taken from the lower and middle parts of Bed 9, but only one specimen, assignable to Wurmiella excavata, which has been widely documented from the lower Silurian to Lower Devonian, was recovered below the lowest occurrence of Caudicriodus sp. B. Caudicriodus sp. B shows similarity with C. hesperius and C. woschmidti in the development of fused transverse ridges on the spindle, a unique feature that is recorded in the early evolution of Caudicriodus during the early Lochkovian. As noted earlier, Caudicriodus sp. B differs from these two species in the shape of the basal cavity, the outline of the spindle, and/or the length of the posteriorly directed later process. Although the occurrence of Caudicriodus sp. B suggests a preliminary correlation of the interval from the uppermost part of Bed 9 to the upper part of Bed 10 with the lower Lochkovian, the precise level of the Silurian-Devonian boundary needs to be investigated further. In addition, other associated fauna includes Pseudoneotodus beckmanni, W. excavata, Zieglerodina eladioi, Z. planilingua, and Z. remscheidensis; the latter three have their highest occurrences in sample ALC-10-23m.
Lanea omoalpha is recorded at a level 3 m above the base of Bed 11. Its lowest occurrence was previously proposed to define the base of the middle Lochkovian and the base of the omoalpha-transitans Zone (Valenzuela-Ríos and Murphy, Reference Valenzuela-Ríos and Murphy1997; Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999). As shown by Murphy and Valenzuela-Ríos (Reference Murphy and Valenzuela-Ríos1999, text-fig. 1), this taxon ranges from the base of the omoalpha-transitans Zone into the transitans-eleanorae Zone in Nevada. However, Corradini and Corriga (Reference Corradini and Corriga2012) documented a much lower occurrence of this species in the lower Lochkovian in the Carnic Alps; a similar situation was also encountered in the Prague Synform (Slavík et al., Reference Slavík, Carls, Hladil and Koptíková2012). In order to eliminate the possibility of confusion in the biostratigraphical correlation caused by the range of L. omoalpha, Corradini and Corriga (Reference Corradini and Corriga2012) accepted the suggestion of Slavík (Reference Slavík2011) and Slavík et al. (Reference Slavík, Carls, Hladil and Koptíková2012) that L. carlsi (= Ancyrodelloides carlsi) acts as a better marker for the lower boundary of the middle Lochkovian. However, Murphy and Valenzuela-Ríos (Reference Murphy and Valenzuela-Ríos2017) preferred A. transitans to mark the base of the middle Lochkovian because of its distinctive features and wider geographic distribution—a suggestion subsequently supported by Corriga and Corradini (Reference Corriga and Corradini2019b). Accordingly, L. omoalpha has a much longer range from the upper part of the hesperius-carlsi Zone into the lower part of the transitans-trigonicus Zone (Corriga et al., Reference Corriga, Corradini and Walliser2014b; Corriga and Corradini, Reference Corriga and Corradini2019b). Therefore, it is difficult to make a precise biostratigraphical determination for the lower part of Bed 11 with only the single occurrence of L. omoalpha in sample ALC-11-3m.
Ancyrodelloides transitans was recorded together with Wurmiella tuma in sample ALC-12-6m, which was collected from a level ~6 m above the base of Bed 12. The lowest occurrence of the latter species in sample ALC-11-9m is almost 6 m lower than that of the former one. Lanea telleri and Flajsella stygia enter together at a slightly higher level, ~10 m above the base of Bed 12. From the overlying strata, only two specimens of W. tuma and one specimen of Pelekysgnathus sp. A were recovered from the upper part of Bed 13. In contrast to the underlying strata, the interval ranging from sample ALC-11-9m to sample ALC-13-13m contains a distinctly low abundance of conodonts. According to Murphy et al. (Reference Murphy, Valenzuela-Ríos and Carls2004), W. tuma normally has a relatively short stratigraphical range from the upper eleanorae-trigonicus Zone to the lower trigonicus-pandora β Zone; however, it extends upwards into the pandora β-gilberti Zone in the Carnic Alps and Sardinia (Corradini and Corriga, Reference Corradini and Corriga2012, fig. 5). Viewed as the important zonal marker of the transitans-eleanorae Zone or transitans-trigonicus Zone, A. transitans has its lowest occurrence at the base of this zone and its highest occurrence in the trigonicus-pandora β Zone (Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999, text-fig. 1; Valenzuela-Ríos and Liao, Reference Valenzuela-Ríos and Liao2012; Valenzuela-Ríos et al., Reference Valenzuela-Ríos, Slavík, Liao, Calvo, Hušková and Chadimová2015, fig. 4). Lanea telleri is restricted to the lower part of the trigonicus-pandora β Zone or the kutscheri-pandora β Zone in the Spanish Central Pyrenees (Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999, text-fig. 4; Valenzuela-Ríos et al., Reference Valenzuela-Ríos, Slavík, Liao, Calvo, Hušková and Chadimová2015, fig. 4), but it also extends downwards into the eleanorae-trigonicus Zone or the upper part of the transitans-trigonicus Zone in Nevada (Murphy and Valenzuela-Ríos, Reference Murphy and Valenzuela-Ríos1999, text-fig. 1). In both Nevada and the Spanish Central Pyrenees, F. stygia enters in the upper half of the transitans-trigonicus Zone (Valenzuela-Ríos and Murphy, Reference Valenzuela-Ríos and Murphy1997, fig. 4; Valenzuela-Ríos and Liao, Reference Valenzuela-Ríos and Liao2012), and has its highest occurrence in the lower part of the kutscheri-pandora β Zone (Valenzuela-Ríos et al., Reference Valenzuela-Ríos, Slavík, Liao, Calvo, Hušková and Chadimová2015, fig. 4). Additionally, somewhat similar ranges of A. transitans, L. telleri, and F. stygia were also observed by Corradini and Corriga (Reference Corradini and Corriga2012, fig. 5) in the Carnic Alps and Sardinia. Taking the stratigraphical ranges of these four species into consideration, it is suggested that the interval from the uppermost part of Bed 11 (sample ALC-11-9m) to the upper part of Bed 13 (sample ALC-13-13m) is correlated with the transitans-pandora β Zone. This interval definitely belongs to the middle Lochkovian, the upper limit of which is defined by the entry of Masarella pandora β morphotype (Valenzuela-Ríos and Murphy, Reference Valenzuela-Ríos and Murphy1997).
One specimen of Pelekysgnathus sp. A was obtained in sample ALC-13-16m from the uppermost part of Bed 13. It differs greatly from previously reported Lochkovian species of this genus. The lack of other important zonal markers in sample ALC-13-16m precludes precise determination of the conodont biostratigraphy for the uppermost part of Bed 13.
Discussion
Wang (Reference Wang1982) first reported the Lochkovian conodont fauna from the Shanjiang Formation or the Lower Devonian at Alengchu and nearby Xiapanluo, which, however, consists of a few specimens of Caudicriodus woschmidti, Spathognathodus remscheidensis, and S. wurmi Bischoff and Sannemann, Reference Bischoff and Sannemann1958. Recently, Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018) restudied the Lower Devonian conodont biostratigraphy at the Alengchu section, and roughly defined the bases of the Lochkovian, Pragian, and Emsian. Although few species, of which most have a rather long stratigraphical range and thus are of little biostratigraphical significance, were recorded from the lower part of the Shanjiang Formation, Wang et al. (2018, p. 294) applied the threefold subdivision of the Lochkovian first proposed by Valenzuela-Ríos and Murphy (Reference Valenzuela-Ríos and Murphy1997; later modified by Slavík et al., Reference Slavík, Carls, Hladil and Koptíková2012) to the studied succession at the Alengchu section. The lower part of the Lochkovian corresponds to an interval from the base of Bed 9 (also the base of the Shanjiang Formation) to the middle part of Bed 10; the middle part of the Lochkovian starts from the middle part of Bed 10 and ends close to the base of Bed 15; and Bed 15 to Bed 17 were assigned to the upper part of Lochkovian (Wang et al., Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018, fig. 3).
Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018, fig. 6T) obtained only one small icriodontid specimen with three transverse rows of distinct nodes on the short but narrow spindle, a narrow basal cavity, and a well-developed but short posterior lateral process in their sample ALC10-1 from the basal part of Bed 10. They provisionally and equivocally assigned it to Caudicriodus hesperius. Taking lithological characters of Bed 9 with respect to the underlying Baizitian Formation and the overlying remaining part of the Shanjiang Formation into consideration, they temporarily located the Silurian-Devonian boundary at the base of the Shanjiang Formation.
In the present study, several adult specimens of Caudicriodus sp. B were obtained in a lower level from the uppermost part of Bed 9, which is ~2 m below the level of sample ALC10-1 of Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018). This species shows a close similarity with C. hesperius and C. woschmidti in the development of the characteristic transverse ridges on the spindle. Moreover, Ma et al. (Reference Ma, Wang and Zhang2017) recognized an intermediate δ13Ccarb positive excursion in the basal part of the Shanjiang Formation at the Alengchu section, which indicates that the Silurian-Devonian boundary probably is situated in the basal part of the Shanjiang Formation (Bed 9). It should be noted that, as demonstrated by C.Y. Wang et al. (Reference Wang, Wang, Yang and Xie2009) and Y. Wang et al. (Reference Wang, Rong, Tang, Wang and Zhang2016), the uppermost Silurian or the Pridoli is probably missing at the Baizitian section, which is also situated in the Panzhihua-Lijiang district. Whether the same situation occurs at the Alengchu needs to be investigated. The entry of Wurmiella tuma within the middle part of Bed 9 (third sample ALC9-1c) of H.H. Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018) is noteworthy because this level is slightly higher than the supposed Silurian-Devonian boundary of Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018). Wurmiella tuma has a lowest occurrence in the middle Lochkovian transitans-trigonicus Zone; thus the reported occurrence by Wang et al. (2018, without illustrations) in such a lower level needs to be examined.
Mainly on the basis of the occurrences of dacryoconarids and the genus Pandorinellina Müller and Müller, Reference Müller and Müller1957, Wang et al. (2018, without illustrations) placed the base of the middle part of the Lochkovian roughly in the middle part of Bed 10. As mentioned by Slavík (Reference Slavík2011) and Slavík et al. (Reference Slavík, Carls, Hladil and Koptíková2012), the phylogenetically oldest dacryoconarids were recorded in the Prague Synform and Carnic Alps within the short range of Lanea carlsi (= Ancyrodelloides carlsi), whose entry was suggested as a marker of the beginning of the middle Lochkovian. Moreover, the origination of Pandorinellina also was demonstrated by Murphy et al. (Reference Murphy, Valenzuela-Ríos and Carls2004) in the middle Lochkovian. However, our study indicates that the upper part of Bed 10 still belongs to the lower Lochkovian. The reported occurrence of Pandorinellina in sample ALC10-4 of Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018) needs to be verified. More importantly, specimens of the zonal marker A. carlsi have not been identified at the Alengchu section. Therefore, the accurate position of the lower boundary of the middle Lochkovian is still unknown at this section.
The base of the upper part of the Lochkovian was correlated by Wang et al. (Reference Wang, Ma, Slavík, Wei, Zhang and Lü2018) with the base of Bed 15, which is slightly higher than the lowest occurrence of Masarella pandora alpha morphotype. Due to the absence of important index taxa from Bed 14 to Bed 17 in the present study, a detailed discussion of the upper Lochkovian is not possible.
Conclusions
The main results of this study on the Lochkovian conodonts at the Alengchu section, western Yunnan is summarized below.
A conodont fauna consisting of 12 taxa assigned to Ancyrodelloides, Flajsella, Lanea, Wurmiella, Zieglerodina, Caudicriodus, Pelekysgnathus, and Pseudooneotodus was obtained from the lower part (Bed 9 to Bed 13) of the Shanjiang Formation at the Alengchu section. Most of these taxa have been rarely reported in China.
Due to the scarcity of index fossils, the interval ranging from the uppermost part of Bed 9 to the upper part of Bed 10 is assigned roughly to the lower Lochkovian. The Silurian-Devonian boundary is probably located in Bed 9 from the basal part of the Shanjiang Formation.
The interval covering the uppermost part of Bed 11 to the upper part of Bed 13 is correlated with the middle Lochkovian transitans-pandora β Zone. This is the first time that the peri-Gondwana conodont-based subdivision schemes have been partially tested in China. Flajsella and L. telleri are documented for the first time in South China.
Acknowledgments
We thank J.I. Valenzuela-Ríos (Valencia) and D.J. Over (Geneseo) for providing important and useful comments that greatly improved the manuscript. T.H. Gao is acknowledged for reading and polishing the manuscript. This work is supported by the Strategic Priority Research Program (B) of the Chinese Academy of Sciences (XDB26000000), the Second Tibetan Plateau Scientific Expedition and Research (No. 2019QZKK0706), the National Natural Science Foundation of China (41872003, 41702009, 91855205), and the Youth Innovation Promotion Association Chinese Academy of Sciences.






