Introduction
Burcucumber (Sicyos angulatus L.), a member of the Cucurbitaceae, is an annual plant (vine) originating in eastern North America and now found in several geographic regions of the world (Kurokawa et al. Reference Kurokawa, Kobayashi and Senda2009; Lee et al. Reference Lee, Kim, Cho and Lee2015; Önen et al. Reference Önen, Özaslan and Tad2015a, Reference Önen, Tad and Özaslan2015b). The species reproduces by seeds and has enormous seed production potential, which favors its invasion success in various environmental conditions. A single plant can produce 716 to 18,197 g biomass and 4,500 to 78,000 seeds under favorable conditions (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a; Smeda and Weller Reference Smeda and Weller2001). The species has been recognized as “invasive” in Europe, where it is continuously expanding its range (Hulina Reference Hulina1996). Sicyos angulatus infests several crops, including corn (Zea mays L.), soybean [Glycine max (L.) Merr.], American hazelnut (Corylus americana Walter), and cucurbits (Cucumis spp.), leading to severe economic losses in terms of yield reduction and management costs incurred (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a, Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001b, Reference Esbenshade, Curran and Roth2009; Gibson et al. Reference Gibson, Johnson and Hillger2005; Önen et al. Reference Önen, Özaslan and Tad2015a, Reference Önen, Tad and Özaslan2015b; Shimizu Reference Shimizu1999; Watanabe et al. Reference Watanabe, Kurokawa, Sasaki, Nishida, Onoue and Yoshimura2002). The species mainly invades natural habitats, riversides, forest fringes, and other open areas. It completely covers underlying native vegetation and blocks sunlight due to its rapid growth (Esbenshade et al. Reference Esbenshade, Curran, Roth, Hartwig and Orzolek2001a; Farooq et al. Reference Farooq, Tad, Önen, Gunal, Caldiran and Özaslan2017; Önen et al. Reference Önen, Özaslan and Tad2015a, Reference Önen, Tad and Özaslan2015b; Watanabe et al. Reference Watanabe, Kurokawa, Sasaki, Nishida, Onoue and Yoshimura2002). The weed can also damage infrastructure such as electric cables and telephone lines (Önen et al. Reference Önen, Özaslan and Tad2015a). Moreover, the presence of the species alongside rivers and stream banks restricts water flow, thus creating various problems for stream managers (Lee et al. Reference Lee, Kim, Cho and Lee2015).
The distribution of exotic species is strongly mediated by regional climatic conditions; however, little is known about the impact of regional environmental conditions such as temperature, pH, salinity, and water stress on exotic species’ growth and development (Skálová et al. Reference Skálová, Moravcová, Dixon, Kindlmann and Pyšek2015). Seed germination is of great significance among life-history traits, since high germination percentages are an important contributor to population establishment and persistence (Mandák Reference Mandák2003; Radford and Cousens Reference Radford and Cousens2000). Seed germination, being the first transition step in the life cycle of annual plant species, influences several aspects of plant biology, such as population dynamics, geographical distribution, and responses to climate change (Bykova et al. Reference Bykova, Chuine, Morin and Higgins2012; Cochrane et al. Reference Cochrane, Yates, Hoyle and Nicotra2015; Huang et al. Reference Huang, Liu, Bradford, Huxman and Venable2016). Various environmental factors such as temperature, light intensity, pH, salinity, moisture availability, and seed burial depth affect weed seed germination and emergence (Awan et al. Reference Awan, Chauhan and Sta Cruz2014; Baskin and Baskin Reference Baskin and Baskin2014; Chauhan Reference Chauhan2016; Chauhan and Johnson Reference Chauhan and Johnson2009).
Significant variation in germination requirements has been recorded for different populations of the same species arising from different climatic regions, and these variations have been correlated with the climatic conditions prevailing at the sites of origin (Hernández-Verdugo et al. Reference Hernández-Verdugo, Oyama and Vázquez-Yanes2001; Kollmann and Bañuelos Reference Kollmann and Bañuelos2004; Leiblein-Wild et al. Reference Leiblein-Wild, Kaviani and Tackenberg2014; Montague et al. Reference Montague, Barrett and Eckert2008). Exploring the variations in seed germination requirements could provide valuable insights for future range expansion potential of invasive plant species (Dlugosch et al. Reference Dlugosch, Anderson, Braasch, Cang and Gillette2015; Kurokawa et al. Reference Kurokawa, Kobayashi and Senda2009). Sicyos angulatus can grow under a wide range of environmental conditions, indicating that different populations exhibit variations in life-history traits (Dlugosch et al. Reference Dlugosch, Anderson, Braasch, Cang and Gillette2015; Kurokawa et al. Reference Kurokawa, Kobayashi and Senda2009). The variations in life-history traits have been reported to result from the genetic diversity in different populations of S. angulatus. However, population differences are often ignored at regional scales, and studying these differences could provide valuable insights for the future spread of the species.
Turkey has vast climatic diversity due to irregular topography, which divides the country into seven geographic regions (Sensoy Reference Sensoy2014; Sensoy et al. Reference Sensoy, Demircan, Ulupinar and Balta2008). The Black Sea is the most humid region, receiving the highest amount of rainfall throughout the year. The first report of S. angulatus in Turkey dates back to 1999 in the Black Sea region (Terzioglu and Ansin Reference Terzioglu and Ansin1999), after which the species has been reported by different researchers at different times in the Black Sea region (Önen et al. Reference Önen, Özaslan and Tad2015a, Reference Önen, Tad and Özaslan2015b; Önen et al. Reference Önen, Özaslan, Günal, Akyol and Caldiran2013; Terzioglu et al. Reference Terzioglu, Coşkunçelebi and Başkent2014). The species has been observed along roadsides in the Black Sea region in different studies (Figure 1), and naturalized populations spread on an area of 3 ha have been observed in the region (Önen et al. Reference Önen, Özaslan and Tad2015a, Reference Önen, Tad and Özaslan2015b). Although recent reports have indicated that S. angulatus has already naturalized in the Black Sea region and is causing massive losses to native biodiversity and agriculture (Önen et al. Reference Önen, Özaslan and Tad2015a, Reference Önen, Tad and Özaslan2015b; Önen et al. Reference Önen, Özaslan, Günal, Akyol and Caldiran2013), no information is available regarding the range expansion potential and germination biology of different populations of the species in Turkey.

Figure 1 Distribution range and locations of burcucumber (Sicyos angulatus) populations collected from the Black Sea region, Turkey.
This study was aimed at determining the influence of different environmental factors on the seed germination biology of three populations of S. angulatus naturalized in the Black Sea region of Turkey (Figure 1). It was hypothesized that different populations will vary in terms of: (1) seed germination requirements, (2) sensitivity to osmotic stress and high salinity, and (3) seedling emergence from various seed burial depths. The results of this study will offer valuable insights for the future spread of S. angulatus in Turkey, as well as increasing our understanding of the species germination biology at a regional scale.
Materials and Methods
Study Area
The study area was the Black Sea region of Turkey, which has a warm, humid climate with average winter and summer temperatures of 7 and 23 C, respectively (Sensoy Reference Sensoy2014; Sensoy et al. Reference Sensoy, Demircan, Ulupinar and Balta2008).
Seed Collection and Seed Dormancy Release
In September 2015, mature seeds of different populations were collected from 40 mother plants per location in the Dernekpazarı, Dereli, and Çamlıhemşin districts of the Black Sea region (Table 1). The seeds were stored in glass jars at room temperature (25 C) up to the initiation of experiments. Data on soil properties of the seed collection sites and the information related to geographic locations and climatic conditions are given in Table 1 and Figure 1.
Table 1 Geographic location, soil properties, and climatic data of different burcucumber (Sicyos angulatus) populations collected from the Black Sea region, Turkey.Footnote a

a EC, electrical conductivity; OM, organic matter; P, phosphorus content; PET, potential evapotranspiration.
b Aridity index>1 indicates areas are humid, while areas having an aridity index>2 are very humid according to Camci Çetin et al. (Reference Camci Çetin, Karaca, Haktanir and Yildiz2007).
Sicyos angulatus seeds possess a hard, water-impermeable seed coat that imparts physical dormancy to the seeds (Lim et al. Reference Lim, Kwon, Kwon, Yoon and Choi1994; Qu et al. Reference Qu, Baskin and Baskin2010). Therefore, seeds of all populations were mechanically scarified using a scalpel to release seed dormancy before germination tests, following Özaslan et al. (Reference Özaslan, Tad, Önen and Farooq2016).
General Procedure for Germination Test
The mechanically scarified seeds of all populations were incubated in petri dishes (90 by 15 mm) on two layers of filter paper. Four milliliters of deionized water or a treatment solution (for salinity, pH, and osmotic potential experiments) was used to moisten the filter paper at the initiation of the experiments. Petri dishes, each containing 25 seeds, were incubated at 25 C (except for the temperature experiment) and 12-h light and dark period (except for the complete-light and complete-dark treatments of the photoperiod experiment).
Seed germination was observed daily (except in the photoperiod experiment), and radical protrusion was the criterion for germination. Germinated seeds were removed from the petri dishes. Moisture was supplied to the seeds according to requirements using deionized water. All experiments were conducted in a factorial design with S. angulatus populations as the main factor and experimental treatments as the subfactor and were terminated 30 d after initiation. The viability of nongerminated seeds was tested using a triphenyl tetrazolium chloride test (Sawma and Mohler Reference Sawma and Mohler2002), and germination was adjusted as viability-adjusted germination following Weller et al. (Reference Weller, Florentine, Sillitoe, Grech and McLaren2016). All treatments in each experiment had five replications (1 petri dish/pot per replication), and all experiments were repeated over time. The viability-adjusted germination (%), computed separately for each replication (i.e., petri dish/pot), was used in statistical analysis and interpretation of results. The fraction of nongerminating viable seeds was nearly 5% in all populations, which excluded the chances of any biased effects for using viability-adjusted germination data in the analyses.
Effect of Photoperiod on Seed Germination
Germination of S. angulatus populations was tested under three light/dark regimes at 12/12 h, 24/0 h, and 0/24 h. Light was supplied in incubators with cool, white fluorescent lamps at 350 µEm−2 s−1 intensity. The petri dishes for the 0/24-h light/dark treatment were wrapped in two layers of aluminum foil to exclude light.
Effect of Temperature on Seed Germination
To test the effect of temperature on seed germination, mechanically scarified seeds of all populations were incubated under nine different constant-temperature regimes (5, 10, 15, 20, 25, 30, 35, 40, and 45 C) for 30 d. The temperature range selected mostly corresponds to the temperatures that the species might experience in Turkey; however, some extreme temperatures (5 and 45 C) were also included in the study to infer how the species will behave under extremely high and low temperatures.
Effect of pH on Seed Germination
Germination of all populations was investigated under a range of pH (4.0, 5.0, and 6.0 for acidic medium; 7.0 as neutral medium; and 8.0, 9.0, 10.0, and 11.0 as alkaline medium). The pH levels were selected to represent the pH of soils in the country. Soils of Turkey vary extensively in pH, but alkaline soils pH >7.5 are predominant (62.78%) (Eyüpoğlu Reference Eyüpoğlu1999). To achieve the desired level of pH, treatment solutions were prepared following Chauhan et al. (Reference Chauhan, Gill and Preston2006).
Effect of Salt Stress on Seed Germination
Nondormant seeds of all populations were incubated under seven levels of salinity to test the effects of salt stress on seed germination. Salt concentrations of 50, 100, 150, 200, 400, and 600 mM sodium chloride (NaCl) along with a control (deionized water only) were included in the salinity experiment. The salinity levels were chosen to represent the saline soil conditions of Turkey (Eyüpoğlu Reference Eyüpoğlu1999).
Effect of Osmotic Stress on Seed Germination
Seed germination of all populations was tested under eight osmotic potentials (−0.2, −0.4, −0.6, −0.8, −1.0, −1.2, −1.4, and −1.6 MPa) along with a control (0 MPa). The osmotic potentials were selected to mimic the possible osmotic stress conditions that species may encounter due to salinity and aridity problems in Turkey. The osmotic solutions were prepared by dissolving appropriate amounts of polyethylene glycol 8000 (PEG-8000) in deionized water, following Chauhan et al. (Reference Chauhan, Gill and Preston2006).
Effect of Seed Burial Depth on Seedling Emergence
The effect of various seed burial depths on seedling emergence of all populations was tested in a greenhouse experiment. Mechanically scarified seeds of all populations were sown at various soil depths (0, 0.5, 1, 2, 4, 6, 8, 10, 12, and 15 cm) in 8.8 L plastic pots (20-cm diameter, 18-cm height). A total of 25 seeds were sown per pot, and each treatment had five replications. The seeds were placed either on the surface (for 0-cm depth) or buried in the soil according to the treatments. The soil used in the pot experiment was a clay loam composed of 27.5% sand, 35% silt, and 37.5% clay, with 1.7% organic matter, 0.3 dS m−1 salinity, 10.99 carbonate, and a pH of 8.18 (Durak and Karan Reference Durak and Karan2017). There was no previous seedbank of S. angulatus in the soil. Seedling emergence (based on cotyledon visibility above the soil) was recorded daily for 30 d. The greenhouse was maintained at 25 C with 12-h light/dark conditions. The pots were initially irrigated with a mist sprinkler, but were subirrigated thereafter.
Statistical Analyses
The recorded data on seed germination (viability adjusted germination %) and seedling emergence (%) were analyzed using different statistical techniques. The homogeneity of variance was visually inspected by plotting residuals to test the variance in data before statistical analyses. ANOVA was used to test the differences among experimental runs of each experiment separately. The ANOVA indicated no differences among experimental runs; therefore, the data of both runs were pooled (so there were 10 replications in total). Petri dishes/pots instead of individual seeds were taken as a replication in the statistical analyses. A two-way ANOVA was then used to test for differences among populations and treatments; this was done separately for each experiment. Least significant difference was used post hoc to separate the means of the photoperiod experiment (Steel et al. Reference Steel, Torrie and Dickey1997).
The germination data of osmotic and salt stress experiments typically exhibit an exponential decay fashion. Thus, a three-parameter sigmoid model is best fit to these sorts of data and extensively used in the literature to analyze such data (Awan et al. Reference Awan, Chauhan and Sta Cruz2014; Chauhan Reference Chauhan2016). Therefore, a three-parameter sigmoid model was fitted to the final germination percentage values of osmotic and salt stress experiments. The model was
 $$G\,{\equals}\,G_{{\rm max}} /\left( {1{\plus}e\,\left[ {{\minus}x{\minus}T_{{50}} } \right)/G_{{\rm rate}} } \right]$$
$$G\,{\equals}\,G_{{\rm max}} /\left( {1{\plus}e\,\left[ {{\minus}x{\minus}T_{{50}} } \right)/G_{{\rm rate}} } \right]$$
In the model, G is the cumulative percentage germination at time x, G max is the maximum germination (%), T 50 is the osmotic potential or salinity level required for 50% inhibition of maximum germination, and G rate indicates the slope.
Temperature, pH, and seed burial experiments generally indicate an increasing trend followed by a decreasing trend. Thus, a three-parameter Gaussian model, which best describes the data of these experiments, was used for the analysis (Awan et al. Reference Awan, Chauhan and Sta Cruz2014; Chauhan Reference Chauhan2016). Therefore, a three-parameter Gaussian model was fitted to the final germination percentage data for the temperature, pH, and seed burial experiments. The model was:
 $$G\,{\equals}\,a{\times}e\,\left[ {{\minus}0.5{\minus}\left\{ {\left( {x{\minus}b} \right)/c} \right\}^{2} } \right]$$
$$G\,{\equals}\,a{\times}e\,\left[ {{\minus}0.5{\minus}\left\{ {\left( {x{\minus}b} \right)/c} \right\}^{2} } \right]$$
The Gaussian model gives a “bell-curve.” In Equation 2, a corresponds to the height of the curve’s peak (maximum germination or seedling emergence); b is the position of the center of the peak (temperature, pH, or depth of seed burial to achieve maximum germination or seedling emergence); and c is the width of the bell, which indicates the range over which germination occurs. ANOVA was performed using SPSS v. 21.0 (IBM 2012), while sigmoid and Gaussian models were fit using SigmaPlot v. 13.0.
Results and Discussion
Light/Dark and Temperature
Seed germination was altered by different light/dark conditions and interactions among populations and light/dark conditions, while populations had nonsignificant (P=0.30) effects on seed germination (Table 2). The highest values (87.0%, 90.5%, and 82.2% for the Dernekpazarı, Dereli, and Çamlıhemşin populations, respectively) of the final germination percentage were recorded under 12/12-h light/dark for all populations. The seeds of all populations had similar germination under 0/24- and 24/0-h light/dark conditions (Figure 2). The seeds of all populations exhibited >50% germination under continuous-light and continuous-dark treatments, indicating that S. angulatus seeds are not strictly photoblastic. Overall, germination percentage was higher in the 12-h light/dark condition than in either the 24/0- or 0/24-h light/dark regime. Although some seeds can germinate in darkness, the highest germination was recorded in seeds receiving both light and dark each day. Seeds of all populations proved nonphotoblastic, indicating their ability to emerge from various seed burial depths. The nonphotoblastic nature of the seeds could be linked with the low number of hours of sunshine received per day in the Black Sea region. Sicyos angulatus can germinate throughout the growing season (European and Mediterranean Plant Protection Organization [EPPO] 2010), indicating that soil depth and light have little effect on seed germination. These results of the photoperiod experiment are also supported by the seed burial experiment, in which seedling emergence was recorded from almost all the burial depths included in the study (data discussed later in the section Seed Burial Depth).

Figure 2 The effect of photoperiod (12/12, 24/0, and 0/24 h) on seed germination of different burcucumber (Sicyos angulatus) populations collected from the Black Sea region, Turkey. Means sharing the same letters are statistically similar (P>0.05). The vertical bars represent the standard error of the mean.
Table 2 ANOVA of different burcucumber (Sicyos angulatus) populations collected from the Black Sea region, Turkey, and experimental treatments (photoperiods, constant temperatures, pH levels, salinity, and osmotic potentials) on final germination percentage, and seed burial depths on seedling emergence percentage.

a df, degrees of freedom.
Seed germination was influenced by population and constant temperatures; however, their interaction was nonsignificant (P=0.34) (Table 2). The nonsignificant interactions could be the result of similar climatic conditions faced by the populations. The seeds of all populations were able to germinate under all temperatures included in the study, except at 5 C (Figure 3). Seed germination increased with increasing temperature, reached a maximum, and then started to decline. More than 50% of the seeds from all populations were able to germinate at temperatures between 10 and 35 C. The optimum temperatures predicted by the model for germination were 24.5, 24.4, and 24.5 C for the Dernekpazarı (93.5%), Dereli (99.5%), and Çamlıhemşin (88.3%) populations, respectively (Figure 3). Overall, seeds collected from Dereli exhibited the highest germination rates, whereas those from Çamlıhemşin exhibited the lowest germination rates under all temperature regimes. The results of the current study are in agreement with the results of Mann et al. (Reference Mann, Rieck and Witt1981), who reported that 25 to 30 C is optimum for germination of S. angulatus. The temperature requirements of S. angulatus can be linked to climatic conditions of the Black Sea region, which has higher precipitation and a warmer climate than other regions in Turkey (Camci Çetin et al. Reference Camci Çetin, Karaca, Haktanir and Yildiz2007; Sensoy Reference Sensoy2014; Sensoy et al. Reference Sensoy, Demircan, Ulupinar and Balta2008). Temperature requirements of S. angulatus indicate that it can germinate in most of the climatic regions in Turkey. Although seeds were able to germinate over a wide range of temperatures, germination alone cannot guarantee the survival and reproduction of a species, as seedling and subsequent growth stages are highly sensitive to adverse environmental conditions (DiTommaso Reference DiTommaso2004).

Figure 3 The effect of different constant-temperature regimes on seed germination of different burcucumber (Sicyos angulatus) populations collected from the Black Sea region, Turkey. The lines represent a three-parameter Gaussian model fit to the final germination percentage data, while vertical bars represent the standard error of the mean.
pH
Different populations and pH levels altered seed germination, but their interaction was nonsignificant (P=0.15) (Table 2). Seeds of all populations exhibited higher germination at slightly acidic and basic pH levels, while a significant reduction in germination was observed under highly acidic and alkaline pH levels (Figure 4). The maximum germination predicted by the model was under pH of 6.9, 6.8, and 6.8 for the Dernekpazarı (76.5%), Dereli (79.5%), and Çamlıhemşin (71.4%) populations, respectively. The Dereli population exhibited the highest germination under all pH levels, while the lowest germination was observed for seeds of the Çamlıhemşin population. The populations studied have been reported to behave similarly for seed dormancy (Özaslan et al. Reference Özaslan, Tad, Önen and Farooq2016); however, slight differences were observed for seed germination. These slight differences are thought to be the result of differences in climatic conditions and soil properties of the seed collection sites (Table 1). Soils in the Dereli site had significantly higher clay and calcium carbonate content and a higher pH level compared with Dernekpazarı and Çamlıhemşin soils.

Figure 4 The effect of different pH levels on seed germination of different burcucumber (Sicyos angulatus) populations collected from the Black Sea region, Turkey. The lines represent a three-parameter Gaussian model fit to the final germination percentage data, while vertical bars represent the standard error of the mean.
Water and Salinity Stress
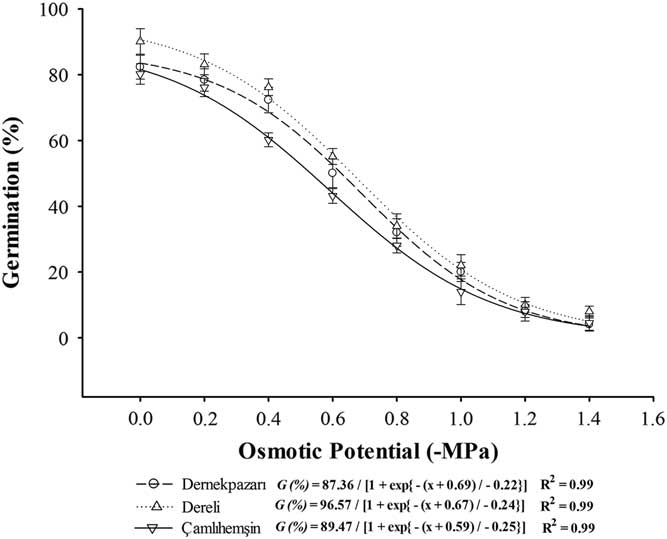
Different osmotic potentials, salinity stress level, and population affected (P < 0.01) seed germination, while their interactions were nonsignificant (Table 2). Overall, seeds from the Dereli population exhibited higher germination compared with the other populations; however, all populations exhibited similar sensitivity to salinity and osmotic potential (Figures 5 and 6). The osmotic potentials required to inhibit 50% of maximum germination predicted by the model were −0.69, −0.67, and −0.59 MPa for the Dernekpazarı, Dereli, and Çamlıhemşin populations, respectively (Figure 5). Similarly, seeds of all populations germinated at between 0 to 300 mM NaCl concentration. Germination of all populations was >50% in NaCl concentrations up to 150 mM, with germination exceeding 25% even at 230 mM NaCl. The salinity levels required to inhibit 50% of maximum germination predicted by the model were 189, 188, and 182 mM NaCl for the Dernekpazarı, Dereli, and Çamlıhemşin populations, respectively (Figure 6). Sicyos angulatus has been proven to be highly sensitive to salinity and water stress for growth and fecundity traits as well (Farooq et al. Reference Farooq, Tad, Önen, Gunal, Caldiran and Özaslan2017). Thus, the sensitivity of seeds to water and salinity stress indicates that S. angulatus could not expand its range to the areas having low water availability and relatively saline and highly acidic or alkaline soils. Similarly, S. angulatus is unable to withstand drought at the germination stage (EPPO 2010). The sensitivity of S. angulatus can be explained as an adaptive response to the soils of the region, which are nonsaline, having almost neutral pH. As all populations arise from the same region (Table 1), they have adapted to existing soil and climatic conditions. However, there could be some maternal effects as well, which need to be explored in detail.

Figure 5 The effect of different osmotic stress levels on seed germination of different burcucumber populations (Sicyos angulatus) collected from the Black Sea region, Turkey. The lines represent a three-parameter sigmoid model fit to the final germination percentage data, while vertical bars represent the standard error of the mean.

Figure 6 The effect of different salinity levels on seed germination of different burcucumber (Sicyos angulatus) populations collected from the Black Sea region, Turkey. The lines represent a three-parameter sigmoid model fit to the final germination percentage data, while the vertical bars represent the standard error of the mean.
Seed Burial Depth
Seedling emergence differed among populations and seed burial depths; however, their interaction remained nonsignificant (P=0.13) (Table 2). Seedling emergence was expected to decrease with an increasing seed burial depth; however, an initial increase up to 2 cm and then a sharp decline was observed in seedling emergence for all populations (Figure 7). The seeds of all populations were able to emerge from 0- to 12-cm depth, and more than 20% seedling emergence was predicted by the model between 0 and 6 cm. The highest seedling emergence was predicted from 2.2-, 2.0-, and 2.1-cm soil depths for the Dernekpazarı, Dereli, and Çamlıhemşin populations, respectively (Figure 7). Seedling emergence of all populations was severely hampered beyond 6-cm burial depth. The reason for low seedling emergence from the surface may be poor soil and seed contact and water imbibition from one side of the seed only, which resulted in low water uptake. Lower water uptake and seedling emergence of seeds placed on the surface have been reported (Messersmith et al. Reference Messersmith, Curran, Roth, Hartwig and Orzolek2000) due to water impermeability of seeds (Qu et al. 2010, Reference Qu, Baskin and Baskin2012). Low emergence of various weedy and invasive species with increasing soil depth has been well described (Awan et al. Reference Awan, Chauhan and Sta Cruz2014; Chauhan Reference Chauhan2016; Chauhan et al. Reference Chauhan, Gill and Preston2006; Chauhan and Johnson Reference Chauhan and Johnson2009; Özaslan et al. Reference Özaslan, Farooq, Önen, Ozcan, Bukun and Gunal2017). The results of the current study are in agreement with Messersmith et al. (Reference Messersmith, Curran, Roth, Hartwig and Orzolek2000), who reported a sharp decline in seedling emergence for S. angulatus beyond 5-cm soil depth. Although, it has been reported that S. angulatus cannot germinate from soil depths of more than 10 cm (Messersmith et al. Reference Messersmith, Curran, Roth, Hartwig and Orzolek2000), our study reports that a small proportion of the seeds are able to emerge even from 15-cm seed burial depth. These results indicate that weed species have the potential to adapt for seedling emergence from deeper soil depths.

Figure 7 The effect of different seed burial depths on seedling emergence of different burcucumber (Sicyos angulatus) populations collected from the Black Sea region, Turkey. The lines represent a three-parameter Gaussian model fit to the final seedling emergence percentage data, while the vertical bars represent the standard error of the mean.
Seeds of all populations were able to withstand a wide range of temperatures, but they were sensitive to higher levels of pH and water and salinity stress, with slight differences among populations. These differences could be explained by the climatic conditions and soil properties of the sampling sites (Table 1). Overall, the results indicate that S. angulatus has a limited range expansion potential in Turkey and is unsuited to even slightly harsh climatic conditions. Saline and alkaline areas were about 2% of the total area of the Turkish soils (Sönmez Reference Sönmez2004), which now have increased to 5.27% following the irrigation projects initiated after the 1970s (Doğan Reference Doğan2014). Similarly >20% area of the country is arid and semiarid (Camci Çetin et al. Reference Camci Çetin, Karaca, Haktanir and Yildiz2007), which is unsuitable for the species. Climate change is expected to increase aridity and salinity in the country (Dudu and Çakmak Reference Dudu and Çakmak2018), which will further halt the spread of this species to new areas. Thus, management efforts should be concentrated on the currently invaded range. In arable lands, a no-tillage approach seems a viable option to check S. angulatus seedling emergence (Messersmith et al. Reference Messersmith, Curran, Roth, Hartwig and Orzolek2000). The seeds remaining on the soil surface will be unable to imbibe water and germinate. The no-tillage option would be viable for the management of S. angulatus in agricultural lands of irrigated areas in Turkey. However, the Black Sea region is the most humid and mountainous region of the country, and seed germination of S. angulatus is known to be stimulated by rainfall (Smeda and Weller Reference Smeda and Weller2001); therefore, a no-tillage approach does not seem a viable management option against S. angulatus in this case. Deep tillage to bury the seeds at sufficient depth, subsequent shallow tillage, and effective management of emerging seedlings could reduce the soil seedbank in the long run. These management options need to be explored in arable lands of the Black Sea region.
Although seeds were collected from the same region, the slight differences in germination suggest that the populations included in the study may have undergone differential adaptations. The population differences arise either due to their origin in different climates or genetic diversity (Gioria and Pyšek Reference Gioria and Pyšek2017). The slight variation in different populations could be explained by earlier findings, which reported significant variations for life-history traits among different populations of S. angulatus (Jeon et al. Reference Jeon, Kang, Yoon, Lee and Chung2003; Kurokawa et al. Reference Kurokawa, Kobayashi and Senda2009; Watanabe et al. Reference Watanabe, Kurokawa, Sasaki, Nishida, Onoue and Yoshimura2002). Therefore, genetic variation, maternal effects, and prevailing climatic conditions could be the potential drivers of the observed variations. However, there is no evidence in the studied populations to support a genetic basis for this inference, which needs to be explored.
The current study adds valuable information to our understanding of the germination biology of the species, which will be helpful in the development of suitable management practices and modeling the potential spread of S. angulatus in Turkey.
Acknowledgments
The current study was funded by the Scientific and Technological Council of Turkey (TUBITAK) with grant number 113 O 790 as a part of the COST Action (TD 1209—European Information System for Alien Species). The authors are grateful to Carol C. Baskin, University of Kentucky, USA, for her valuable comments and suggestions on the earlier drafts of this article. No conflicts of interest have been declared.